
Abstract
Several studies have shown the effect of climatic oscillations on fisheries. Small pelagic fish are of special global economic importance and very sensitive to fluctuations in the physical environment in which they live. The main goal of this study was to explore the relationship between the North Atlantic Oscillation (NAO), the East Atlantic pattern (EA), and the Arctic Oscillation (AO) on the landings and first sale prices of the most representative small pelagic commercial species of the purse-seine fisheries in the Gulf of Cadiz (North East Atlantic), the European anchovy Engraulis encrasicolus and the European sardine Sardine pilchardus. Generalised linear models (GLMs) with different data transformations and distribution errors were generated to analyse these relationships. The best results of the models were obtained by applying a moving average of order 3 to the dataset with a double weighted median. Our results demonstrate relationships between NAO, AO, and EA and European anchovy and sardine landings. These cause an indirect effect on the first sale price in markets through catch variations, which affect the price according to the law of supply and demand. The limitations of this study and management implications are discussed.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The effect of climatic variability of large-scale climatic phenomena on small pelagic fisheries is a well-described fact in scientific literature (for example Chávez et al. 2003; Alheit et al. 2009; Checkley et al. 2017; Báez et al. 2019a; Báez et al. 2021). Understanding the effect of these variations is essential for better management of fish stocks (Leitão et al. 2014; Leitão 2015), as the climatic variables that affect the ocean can alter feeding patterns, growth, and the migratory behaviour of the fisheries’ target species (Miller et al. 2010). Many authors have shown that climatic oscillations can affect the catchability of species (Rubio et al. 2015), their physical condition (Basilone et al. 2006; Báez et al. 2019b), and fishing effort (Rubio et al. 2015), amongst others (for example, see Báez et al. 2021 for a review). Moreover, recent studies have shown that climatic oscillation could explain first sale fish market prices (Fernández et al. 2020). The first sale price of a fishery product is the price that the first buyer in the market chain has paid for the product. This is established in Spain through public action at the fish market.
According to Barnston and Livezey (1987) and Wang et al. (2005), the three most important interannual sources of climatic variability patterns in the Northern Hemisphere that affect the Atlantic Ocean are North Atlantic Oscillation (NAO), East Atlantic pattern (EA), and Arctic Oscillation (AO).
The NAO is defined by the redistribution of atmospheric masses between the subtropical high surface pressure centre located near the Azores (Azores High) and the centre of low surface pressures near Iceland (Icelandic Low) (Hurrell and Deser 2009; Báez et al. 2021). The NAO phases determine the strength and direction of the westerly winds. When the pressures of the subtropical high and the polar low intensify, the NAO is in its positive phase. This event increases the number and intensity of the disturbances that cross the Atlantic towards north-western Europe, leading to hot and wet winters in north-western Europe, whilst decreasing rainfall in winter in the Iberian Peninsula. When the NAO is in its negative phase, the opposite happens (Hurrell 1995). Sánchez et al. (2007) showed how on a regional scale in the Gulf of Cadiz, the positive and negative phases of the NAO are associated with upwellings and downwellings, which in turn are related to temperature variations. The NAO shows both intra- and inter-annual variation (Hurrell 1995).
The EA consists of a North–South dipole that spans the North Atlantic Ocean, with centres near 55°N, 20–35°W and 25–35°N, 0–10°W. It is structurally similar to the NAO, and its anomaly centres are found to the southeast of it. A positive EA phase is associated with an increase in the mean rainfall in northern Europe and Scandinavia and a decrease in rainfall in southern Europe and is also associated with an increase in temperatures in the north of the Iberian Peninsula (Sáenz et al. 2001) and vice versa (Rodríguez-Puebla et al. 1998, 2001). The negative phase is associated with upwellings on the west coast of the Iberian Peninsula (De Castro et al. 2008).
The AO was described by Thompson and Wallace (1998) as the main dominant mode of variability in the Northern Hemisphere. It is the dominant pattern of non-seasonal variations in atmospheric pressure north of 20°N and is characterised by anomalies in pressure of positive or negative magnitudes in the Arctic and anomalies of opposite magnitudes located between 37 and 45°N. The positive phase is characterised by a strengthening of the polar vortex from the surface to the lower stratosphere, resulting in storms in the North Atlantic and a shift of droughts to the Mediterranean. During the negative phase, the continental cold air spreads through western Europe whilst the storms intensify in the Mediterranean region (Ambaum et al. 2001; Rodríguez-Puebla et al. 2002). Different studies have shown that there is a connection between the NAO and the AO (Overland et al. 2010; Báez et al. 2013a). During winter, both AO and NAO tend to be in a positive phase when the stratospheric vortex is strong (Douville 2009).
The Gulf of Cadiz is located to the southwest of the Iberian Peninsula and, due to its geographical position, presents a continuous exchange of water masses between the Atlantic Ocean and the Mediterranean Sea through the Strait of Gibraltar, which makes it a highly dynamic area (Vargas et al. 2002). The mouths of the important rivers such as the Guadalquivir or the Guadiana, amongst others, turn the waters around them into very productive areas through a major contribution in nutrients (Uriarte et al. 1996; Pérez-Rubín et al. 1997; García-Lafuente and Ruiz 2007). Together with other physical–chemical and ecological processes, this contributes to making the Gulf of Cadiz a spawning, breeding, and juvenile area for many important fishery species such as the European anchovy Engraulis encrasicolus (Linnaeus, 1758) and the European sardine Sardina pilchardus (Walbaum, 1792) (Pérez-Rubín et al. 1997, 1999; García-Isarch et al. 2003a, b; Baldó et al. 2006).
In general, small pelagic fishes are a group of special economic importance for all countries. However, due to the peculiar hydrological and hydrodynamic processes in the Gulf of Cadiz, small pelagic fisheries represent an important economic resource (Baldó et al. 2006). The fishing resources of this area are characterised by the seasonal alternation of various types of fishing gear with different target species, which are related to the variation in abundance of the species in accordance with the season. These variations are affected by both biological factors (dependent on the ecology of the species) and economic factors (dependent on the market value) (Ruíz et al. 2017). In the purse-seine fleet of the Gulf of Cadiz, the main target species is the European anchovy that inhabits these waters due to its high first sale price, but it also captures other species of great fishing interest, mainly European sardine (the second most important species caught by purse seiners from the Gulf of Cádiz) (Millán 1992; Casimiro-Soriguer et al. 2000; Ruíz et al. 2017; ICES 2020). Since 2002, the low recruitment of the European anchovy has kept the population at historical critical levels (ICES 2008). Over the last 15 years, the Iberian Peninsula European sardine stock decreased severely due to prolonged low recruitment and high catch levels, with serious social and economic impacts on the Portuguese and Spanish purse seine fisheries (Silva et al. 2019). After a last good recruitment in 2004, during the years 2006 to 2010, European sardine recruitment decreased to very low levels affecting catches in subsequent years (ICES 2011; Garrido et al. 2017).
The Spanish fleet includes approximately 87 vessels with a length range between 10 and 25 m (Ruíz et al. 2017). From 2002 to 2004, the large tonnage purse seine fleet returned to fishing. However, to counterbalance this increase in fishing effort, authorities used a combination of fishery closures and reductions in the number of purse-seine vessels (ICES 2007). The Gulf of Cadiz purse-seine fleet mainly operates from the home-base harbours of Barbate, Punta Umbría, and Isla Cristina. Vessels from Barbate harbour are relatively large and until 1999 could also fish in Moroccan waters outside of the EU Economic Exclusive Zone (EEZ) (the agreement between Morocco and the EU was terminated on 30 November 1999 and was not renewed again until 2006) (García del Hoyo et al. 2004; García-Isarch et al. 2012). These conditions along with environmental pressures make landings from European anchovy and European sardine notably fluctuate jeopardising the biological stability of the species and its economic sustainability, risking the jobs of nearly 800 employees (Ruíz et al. 2017).
The main aim of this present study is to determine the relationship between the most important interannual climatic oscillations in the Northern Hemisphere (i.e., climatic oscillations NAO, AO, and EA) and the European anchovy and sardine landings, and first sale prices in the Gulf of Cádiz. In the same way, this study tries to use the climatic oscillations as a proxy to perform ecological predictions, to establish the conceptual bases of the relationships, and to know and anticipate the fisheries’ level of resilience to climate oscillation variability.
Material and methods
Study area
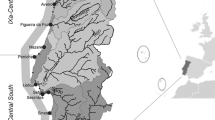
For this study, the Gulf of Cadiz has been limited to the waters between the meridian of Punta Marroquí (near Tarifa) and Cape San Vicente, covering approximately 300 km of coastline (Bellido et al. 2000). The study area is included in the ICES subdivision IX.a-South (Fig. 1) which is the area where the purse seine fleet fish.

Gulf of Cadiz (ICES subdivision IX.a-South, Southwest Spain)
Fisheries data
European anchovy and European sardine landings’ data has been used as a proxy for species abundance (García et al. 2003). Annual landings data (t) and first sale prices (€/kg) of the species from 1985 to 2017 were extracted from the Fisheries Information System Database of the Junta de Andalucía (Andalusia Regional Government) (Sistema de Información Andaluz de Comercialización y Producción Pesquera of the Junta de Andalucía 2020). The first sale prices were standardised with the annual Consumer Price Index (CPI) provided by the Spanish National Institute of Statistics (Instituto Nacional de Estadística, 2020) performing a restatement of stockholders’ equity (Fernández et al. 2020). Hereinafter, this variable is named as standard first sale price.
Atmospheric oscillation data
Three climate indices have been selected: North Atlantic Oscillation (NAO), Arctic Oscillation (AO), and the East Atlantic pattern (EA). Monthly NAO, AO, and EA indices values were taken from the website of the National Oceanic and Atmospheric Administration (NOAA): https://www.noaa.gov/. Six variables were extracted from each climatic index: annual index (average of all months of the year), winter index (average for the months December to March), summer index (average for the months June to August), and 3 lagged indices (using different lags to compare fishing data from one year with climatic indices from previous years, in order to analyse the effect on the early life stages and the influence on recruitment). Up to a 3-year lag between the influence of environmental conditions and anchovy landings was used based on the age classes that were captured the most in the study area (ICES 2020) which could suggest a knock-on effect from the environmental oscillations. Each variable was also square or cube transformed. The use of variables with quadratical and cubical transformations could show potentially, ecologically meaningful relationships with landings. The database was finally composed of 108 climate variables. The synopsis of the variables and sub-variables used in this study is summarised in Table 1.
Generalised linear models (GLM)
Two types of GLM (linear and logistic) were performed with 7 different transformations of the data, obtaining 7 different smoothing methods (Table 2). Data smoothing is a statistical technique that consists of removing outliers from a data set to make a pattern more visible. This improves the fit of the residuals to the family of GLMs and removes noise from the data (Tong 1976).
Linear GLMs were performed using the gamma error distribution for continuous variables, and logistic regressions were performed using the binary error distribution for response variables composed of 1 s and 0 s (Zuur et al. 2009).
First, GLM models were generated using landings (in tons, t) as the response variable and the climatic indices as explanatory variables. To find the explanatory variables that best fit each model, the step forward methodology was used, carried out as follows: (1) A battery of models were created with the response variable and with each of the climate oscillations as explanatory variables; (2) the model with the lowest AICc and the highest percentage of deviance explained was selected; and (3) if any of the explanatory variables were not introduced in a significant way, they were eliminated from the model.
To create bioeconomic models, we used the ecological variables selected in the previous GLM models. In this way, ecological interpretations can be made from the bioeconomic models, and therefore ecological management can be linked to economical implications. The bioeconomic models were fitted using the same error distribution and link as the ecological model. A stepwise backward selection (using the “step” function) was performed. This method iteratively removes the least contributive predictors and stops when it has found the most parsimonious model (Crawley 2012).
In Online Resource 1, we provide a detailed explanation on data analysis, bioeconomic models, and model evaluation. All statistical models were generated with R version 4.0 and RStudio version 1.1.463 (RStudio Team 2016; R Core Team 2020).
Results
A preliminary exploration was performed for each species (Online Resource 2). A total of 1520 GLMs were performed for both species. The best results of the linear regression models are shown in Table 3.
The best results of the European anchovy ecological GLMs were obtained using the gamma error structure and the inverse link function. The three climatic variables were significant in all of the ecological models, except WMA_all (i.e., the transformation moving average of order 3 where the median of all variables is double weighted), which only included the variables AO and EA. The variable EA was introduced 3-year lagged into all of the ecological models and, except for the WMA_all model, the NAO variable was introduced into all of the linear ecological GLMs. In most models, NAO was introduced as the sub-variable NAO winter with a 3-year lag. The results showed an increase in the deviance and goodness of fit when both the response and explanatory variables are smoothed. The WMA_all and MA_all models (i.e., the transformation moving average of order 3 applied to all variables) explained the highest deviance percentages (~ 80 and ~ 76%, respectively). Multicollinearity was not found in any ecological model (VIF < 10). The Raw (without data transformation), MA_all, and WMA_all models did not present autocorrelation in the residuals. The ecological models that best adjusted to the data were the WMA_all and MA_all models, with an observed vs predicted R2 of 0.804 and 0.752, respectively.
For European sardine, all linear GLMs were better fitted using the identity link function, except for the Raw model that used the inverse link function. The three climatic indices were introduced significantly in all the models except in the MA_resp (i.e., the transformation moving average of order 3 applied only to the response variables), which only included the variables AO and EA. The NAO climatic index was always entered as the sub-variable NAOw1, corresponding to the winter NAO with 1-year lag. As in the European anchovy models, smoothing all variables in the models (_all) improved the explained deviance and goodness of fit. The WMA_all model explained the highest percentage of deviance (~ 75%) followed by the MA_all model (~ 68%). The model with the lowest percentage of explained deviance was Raw with 40.16%. No model presented multicollinearity. The Raw, MA_all, and WMA_all models did not show residual autocorrelation. The model that best fit the data was WMA_all (R2 = 0.703) followed by the MA_all model (R2 = 0.685).
The best results of the logistic GLMs are shown in Table 4.
For European anchovy, the logistic models significantly introduced the variable EA but not the variable NAO. The variable AO was only introduced in the Logit_mean model (i.e., binary transformation of the response variable according to whether the value is higher (1) or lower (0) than the mean). The Logit_mean model explained 48.44% of the deviance against the 22.66% of the Logit_cp model (i.e., binary transformation of the response variable according to whether the value of the observation was higher (1) or lower (0) than the mean in the corresponding changepoint period). The Logit_mean model did not present multicollinearity amongst variables. The residuals of the models showed not to be autocorrelated. The logistic models showed to be well fitted according to the Hosmer–Lemeshow statistic (p > 0.05), though the AUC value of the Logit_mean model was higher than that of Logit_cp, showing the model had a better fit. The Logit_mean model obtained lower accuracy (~ 36%) and higher precision (75%) than the Logit_cp model (~ 55 and ~ 67%, respectively).
For European sardine, the variables NAO and EA were introduced in both logistic GLMs, whilst the variable AO was only introduced in the Logit_cp (i.e., binary transformation of the response variable according to whether the value is higher (1) or lower (0) than the mean in the corresponding changepoint period) model. The Logit_mean model (i.e., binary transformation of the response variable according to whether the value of the observation was higher (1) or lower (0) than the total mean) presented a higher percentage of explained deviance (~ 55%) than the Logit_cp model (~ 52%). The residuals of the models showed not to be autocorrelated. The models showed to be well fitted according to the Hosmer–Lemeshow statistic (p > 0.05), though the AUC value of the Logit_mean model was higher than that of Logit_cp, showing a better fit. The Logit_mean model obtained lower accuracy (~ 42%) and precision (~ 86%) than the Logit_cp model (~ 45 and ~ 94%, respectively).
Models that presented an explained deviance of less than 50% were discarded. Finally, after comparing the amount of deviance explained by the model and the goodness-of-fit, the model that best fits both species’ landings is the WMA_all model, which corresponds to the transformation of the data by means of a moving average whose median is double weighted. For European anchovy, the partial effects plot (Fig. 2) shows that negative values of the EA variable 3 years prior to the landings positively affect the response variable (and vice versa) and extreme values (both positive and negative) of AO during the summer affects landings of the same year negatively. For European sardine, partial effects plot (Fig. 3) shows how negative values of the NAO variable and positive AO and EA values have a positive effect on European sardine landings.

Partial effects plots of the European anchovy WMA_all model (Moving average of order 3 where the median of all the variables is double weighted). Key: Desc_ANE, European anchovy landings; AOs_sq, summer Arctic Oscillation squared; EAs3_cb, summer East Atlantic pattern with 3-year lag and cubed

Partial effects of the European sardine WMA_all model (Moving average of order 3 where the median of all variables is double weighted). Key: Desc_PIL, European sardine landings; NAOw1, winter North Atlantic Oscillation with 1-year lag; AOs1_cb, summer Arctic Oscillation with 1-year lag and cubed; EAw3_sq, winter East Atlantic pattern with 3-year lag and squared
Bioeconomic models
In a first step, in the case of the European anchovy, residual autocorrelation was found in all bioeconomic models performed. The bioeconomic model that best fit the data was WMA_all. Similarly, in the case of the European sardine, the step function did not return any bioeconomic model with significant explanatory variables, so no valid bioeconomic model could be generated.
For this reason, in a second step to improve and fix the assumptions of the bioeconomic models, these were repeated (using the same methodology as for the ecological ones) with two modifications: (1) splitting the dataset into two parts (1985–1999 and 2000–2017) and (2) including landings and first sale prices of the other species as potential explanatory variables. The hope was that by splitting the series after the year with the highest catch landings (and therefore the highest supply), the variability within each data subset would be smoothed. Furthermore, landings were included as explanatory variables as according to García del Hoyo (1997) the European sardine fishery is less economically profitable than the European anchovy; therefore, their landings could be negatively dependent on the European anchovy fishery.
The resulting models describing first sale prices had significant explanatory variables and no autocorrelation; these results can be found in Table 5.
Discussion
Ecological models
In the case of European anchovy landings, we found that negative values of the EA variable 3 years prior to the landings would positively affect the response variable, and vice versa. In addition, extreme values (both positive and negative) of AO during the summer seem to negatively affect the landings of the same year (Fig. 2).
The summer East Atlantic pattern is the second dominant mode of summer low-frequency variability in the Euro-Atlantic region. A positive EA phase is associated with an increase in the mean rainfall in northern Europe and Scandinavia, and a decrease in rainfall in southern Europe and is also associated with an increase in temperatures in the north of the Iberian Peninsula (Sáenz et al. 2001) and vice versa (Rodríguez-Puebla et al. 1998, 2002). The onset of the European anchovy spawning period in the Gulf of Cadiz is related to the seasonal warming of the sea surface waters and the beginning of the stratification of the water column (Palomera 1992; García and Palomera 1996; Motos et al. 1996; Kideys et al. 1999; Millán 1999; Baldó et al. 2006). However, the biological characteristics of the small pelagic fishes make them highly sensitive to environmental forcing and extremely variable in their abundance (Alheit et al. 2009, 2012). In recent years, an increase in abundance and a gradual expansion northward of Round sardinella (Sardinella aurita) have been documented along the western Mediterranean coasts in relation to a progressive increase in sea water temperature (Sabatés et al. 2006, 2013; Tsikliras 2008); this species has been also found in the Gulf of Cadiz (Pérez-Rubín and Mafalda 2004). Round sardinella is a thermophilic species particularly frequent in the warmer waters of the eastern and southwestern Mediterranean basins (Sabatés et al. 2013). The Round sardine reproduces in summer, from late June to September, when surface waters reach the highest temperature of the year (Palomera and Sabatés 1990; Somarakis et al. 2002). Spawning periods of the European anchovy and Round sardinella coincide during the summer, and their temperature-related cohabitation has already been documented (e.g., Palomera and Sabatés 1990; Maynou et al. 2008; Zarrad et al. 2012; Karachle and Stergiou 2013; Diankha et al. 2015). Their cohabitation has also been demonstrated specifically in our study area, the Gulf of Cadiz (Perez-Rubín and Mafalda 2004). The partial spatial coincidence and the same vertical distribution of larvae, as well as their morphological similarity, may result in competition for food, as has been shown by other authors (Morote et al. 2008; Schismenou et al. 2008; Macías et al. 2014; Maynou et al. 2014; Albo-Puigserver et al. 2019; Effrosynidis et al. 2020). The numerical abundance of European anchovy versus Round sardinella has also been shown to be one of the reasons for displacement between species, so different temperature windows caused by different climatic indices may favour the presence of one species or another (Palomera and Sabatés 1990; Raab et al. 2013; Diankha et al. 2015). Mellado-Cano et al. (2019) demonstrated how the different phases of the EA regulate extreme high temperature events led by the NAO, even reversing the patterns of climate variability. The consideration of the EA as a temperature regulator together with the NAO can explain over 50% of the variation in temperature and can cause differences of up to 2 and 3 °C (Moore and Renfrew 2011). Therefore, we conclude that the inhibition of extremely warm temperatures by the EA in its negative phase could benefit the European anchovy by displacing a competitor, the Round sardinella. This results in a favourable spawning period for European anchovy, which has a knock-on effect that is reflected in the catches 3 years later.
On the other hand, the AO values that are affecting European anchovy landings are occurring without a time lag. Therefore, the AO is not having an effect on biomass, but on fishing effort. The AO values play an important role in determining extreme conditions such as frozen precipitations, strong winds, and extreme weather events in the Gulf of Cadiz (Rangel-Buitrago and Anfuso 2012; Cabrero et al. 2019). This suggests that extreme AO values result in the reduction of catch per unit effort and fishing effort due to adverse weather conditions.
In the case of European sardine landings, our results show that negative NAO during the previous winter and positive AO during the previous summer favour European sardine landings.
The negative phases of NAO induce major precipitation in southern Europe (Trigo et al. 2002, 2004; Hurrell et al. 2003; Vicente-Serrano et al. 2011; Báez et al. 2013a). Survival of European sardine larvae is closely related to vertical mixing and, consequently, to wind stress as a contributing mechanism (Lloret et al. 2004). Atmospheric disturbances affect marine sedimentation by a transfer of energy from the air to the sea, and from it to the seabed (De Luque 2008). This energy agitates the waters favouring the mixing of deep and superficial waters, increasing the contribution of nutrients to the surface (Báez et al. 2013b), affecting primary production positively, and this in turn affects the abundance of European sardine (Vargas-Yáñez et al. 2020). The increase in precipitation by the negative NAO also leads to an increase in runoff from the Iberian Peninsula (Trigo et al. 2004; Báez et al. 2013a). The fertilisation and local planktonic production by these plumes of continental freshwater support the growth and survival of the fish larvae of this species and avoid starvation (Chícharo et al. 2003; Santos et al. 2007). The Guadalquivir estuary (located at the mouth of the Guadalquivir and Guadiana rivers) is considered an important nursery area for many different species (Baldó et al. 2006). Therefore, the rainfall regime and the flow of the rivers, driven in turn by negative NAO phases, could be of great importance on a regional scale (Trigo et al. 2004). Moreover, Guisande et al. (2001) indicate the advantageous effect of the NAO in its negative phase on the recruitment of the European sardine due to the fact that the prevailing winds from the south drive the flow of water from the sea to the coast avoiding larval drift offshore, as well as the nutritional benefit from the mixture of nutrients in the column caused by a negative NAO phase.
Positive phases of the summer AO produce warmer conditions in the Iberian Peninsula, increasing dry winds (Marshal et al. 2001; Hall et al. 2014; Baldwin et al. 2007). Within the Atlantic Iberian waters, European sardine grows and improves in condition during spring and summer when temperature is close to the annual maxima and plankton production is high (Silva et al. 2008). Thus, the hydrographical conditions derived from a positive AO during the previous summer favour the biological conditions of the spawners during the following winter.
The climatic variable EA has been included as the same sub-variable previously introduced in the European anchovy model but here it was squared. The fact that it is squared implies that extreme values of the winter EA 3 years prior to the catches are beneficial to European sardine landings (Fig. 3). European sardine recruitment is greatly impaired when the temperature tends to be above or below the optimal range (Garrido et al. 2017). The impact of temperature on European sardine landings lead by the AO has been documented (Báez et al. 2019a). Therefore, we suggest that the positive effect of the variable EAw3 on European sardine landings is because this variable in its extreme values may be detrimental to European anchovy, with the consequent redirection of fishing effort from European anchovy to European sardine as the second main target species.
Bioeconomic models
Our results show that the prices of European anchovy during the first sale stage are conditional on the price of European sardine and the climatic subvariate EA 3-year lag. At the same stage, the first sale price of European sardine depends on the prices and landings of European anchovy. The results of these models present a high amount of explained variance (> 80%) and a high fit (R2 predicted vs. adjusted > 0.80) which means that the resulting models can be considered as valid as well as good results. The inclusion of the EA variable in the model increases the percentage of explained variance by 9%, thus confirming the indirect effect of climate on the price of European anchovy. García del Hoyo (1997) found that the price of European anchovy during this period was sufficiently elastic to not depend on landings. Casimiro-Soriguer et al. (2000) studied the price of European sardine and anchovy during this period obtaining similar results in a similar period for European sardine first sale prices, depending only on his own landings.
During the second period (2000–2017), the results show that the price of European anchovy is dependent on its own landings and that of European sardine. During this period the European sardine prices are dependent only on its own landings. The results for European anchovy are considered acceptable although they have lower explained deviance and fit (~ 50%). These results confirm the relationship between European anchovy landings-standard first sale prices-climatic oscillations during both periods. The results in the European sardine bioeconomic model are considered insufficient to determine the factors modulating European sardine prices during the second period. There are very influential factors in the price of European sardine such as the demand by the canning industry or the high seasonal nature of the demand for its fresh consumption, especially during the summer months, where it can triple in value (Casimiro-Soriguer et al. 2000). Prices are not only dependent on the law of supply and demand but are also conditioned by multiple factors (biological, social, economic, institutional, commercial factors, etc.).
Limitations of this study
Fishing effort is not available for the study period. Thereby, the catch per unit of effort could not be used as a proxy for stock abundance. However, given that the fleet in the fishing area has not changed greatly during the study period (ICES 2018), standardisation by unit effort may be dispensable (see OR3). Furthermore, landings have been used in other similar studies (García et al. 2003; Báez and Real 2011; Keller et al. 2014).
The patterns and processes reflected by climate indices are still unclear and difficult to discover (Straile and Stenseth 2007). Nevertheless, using global climate indices such as NAO, AO, or EA, the biological effects may exhibit a longer delay than with respect to any single local climate variables independently, which makes it possible for ecologists to anticipate them and make predictions (Báez et al. 2021). The mechanisms by which these climate indices act remain unclear, although there are well-established plausible mechanisms in the literature that could explain the results found, as detailed above.
Climatic oscillations are called “packages of weather” due to the effect on multiple weather variables simultaneously (Stenseth et al. 2003). The link between climatic oscillations and the corresponding ecosystem response are called teleconnections (Heffernan et al. 2014). Present results show that a significant part of the variability in interannual landings of European anchovy and European sardine stocks could have a relationship with the large-scale climate oscillations NAO, AO, and EA.
In summary, our results reveal the impact of short-term climatic oscillations on European anchovy and European sardine landings. Fishing is primarily an economic activity, and climate variability can also affect economic performance by driving changes in catch prices, due to the effect on supply. Finally, the planet is experiencing global warming. According to most forecasts, the climatic indices will become more and more extreme (for example Báez et al. 2021), and as has been highlighted in this study, the extreme values of the climatic oscillations in most models are those that have the greatest impact. Thereby, those climatic oscillations should be incorporated into fishery management, and future studies should focus on finding the mechanisms involved at the regional level. The bioeconomic models created allow for ecological interpretations, and therefore it is possible to link ecological management with economical implications. In this context, in agreement with Báez et al. (2021), input-based control measurements should be preferred in these highly variable and unpredictable situations.
Data availability
The datasets generated and/or analysed during the current study are available in the Sistema de Información Andaluz de Comercialización y Producción Pesquera of the Junta de Andalucía repository, http://www.juntadeandalucia.es/agriculturaypesca/idapes/servlet/FrontController.
Code availability
All the R scripts will be accessible in the supplementary information.
References
Albo-Puigserver M, Borme D, Coll M, Tirelli V, Palomera I, Navarro J (2019) Trophic ecology of range-expanding round sardinella and resident sympatric species in the NW Mediterranean. Mar Ecol Prog Ser 620:139–154
Alheit J, Pohlmann T, Casini M, Greve W, Hinrichs R, Mathis M, O’Driscoll K et al (2012) Climate variability drives anchovies and sardines into the North and Baltic Seas. Prog Oceanogr 96:128–139. https://doi.org/10.1016/j.pocean.2011.11.015
Alheit J, Roy C, Kifani S (2009) Decadal-scale variability in populations. In: Alheit J, Oozeki Y, Roy C (eds) Checkley Jr. Cambridge University Press, Cambridge, pp 64–87
Ambaum MHP, Hoskins BJ, Stephenson DB (2001) Arctic Oscillation or North Atlantic Oscillation? J Clim 14(16):3495–3507. https://doi.org/10.1175/1520-0442(2001)014%3c3495:AOONAO%3e2.0.CO;2
Arthun M, Bogstad B, Daewel U, Keenlyside NS, Sando AB, Schrum C, Ottersen G (2018) Climate based multi-year predictions of the Barents Sea cod stock. PLoS ONE 13(10):e0206319. https://doi.org/10.1371/journal.pone.0206319
Báez JC, Real R (2011) The North Atlantic Oscillation affects landings of anchovy Engraulis encrasicolus in the Gulf of Cadiz (south of Spain). J Appl Ichthyol 27(5):1232–1235. https://doi.org/10.1111/j.1439-0426.2011.01796.x
Báez JC, Gimeno L, Real R (2021) North Atlantic Oscillation and fisheries management during global climate change. Rev Fish Biol Fish. https://doi.org/10.1007/s11160-021-09645-z
Báez JC, Gimeno L, Gómez-Gesteira M, Ferri-Yáñez F, Real R (2013a) Combined effects of the North Atlantic Oscillation and the Arctic Oscillation on sea surface temperature in the Alboran Sea. PLoS ONE 8(4):e62201. https://doi.org/10.1371/journal.pone.0062201
Báez JC, Macías M, De Castro M, Gómez-Gesteira M, Gimeno L, Real R (2013b) Analysis of the effect of atmospheric oscillations on physical condition of prereproductive bluefin tuna from the Strait of Gibraltar. Anim Biodivers Conserv 36(2):225–233
Báez JC, Santamaría MTG, García A, González JF, Hernández E, Ferri-Yáñez F (2019a) Influence of the arctic oscillations on the sardine off Northwest Africa during the period 1976–1996. Vie Et Milieu 69(1):71–77
Báez JC, Muñoz-Exposito P, Gómez-Vives MJ, Godoy-Garrido D, Macías D (2019b) The NAO affects the reproductive potential of small tuna migrating from the Mediterranean Sea. Fish Res 216:41–46
Baldó F, García-Isarch E, Jiménez MP, Romero Z, Sánchez-Lamadrid A, Catalán IA (2006) Spatial and temporal distribution of the early life stages of three commercial fish species in the northeastern shelf of the Gulf of Cadiz. Deep Sea Res Part II 53(11–13):1391–1401. https://doi.org/10.1016/j.dsr2.2006.04.004
Baldwin MP, Dameris M, Shepherd TG (2007) How will the stratosphere affect climate change? Science 316(5831):1576–1577. https://doi.org/10.1126/science.1144303
Barnston AG, Livezey RE (1987) Classification, seasonality and persistence of low-frequency atmospheric circulation patterns. Mon Weather Rev 115(6):1083–1126. https://doi.org/10.1175/1520-0493(1987)115%3c1083:CSAPOL%3e2.0.CO;2
Basilone G, Guisande C, Patti B, Mazzola S, Cuttitta A, Bonanno A, Vergara AR, Maneiro I (2006) Effect of habitat conditions on reproduction of the European anchovy (Engraulis encrasicolus) in the Strait of Sicily. Fish Oceanogr 15(4):271–280. https://doi.org/10.1111/j.1365-2419.2005.00391.x
Bellido JM, Pierce GJ, Romero JL, Millán M (2000) Use of the frequency analysis methods to estimate growth of anchovy (Engraulis encrasicolus L. 1758) in the Gulf of Cadiz (SW Spain). Fish Res 48:107–115. https://doi.org/10.1016/S0165-7836(00)00183-1
Cabrero Á, González-Nuevo G, Gago J, Cabanas JM (2019) Study of sardine (Sardina pilchardus) regime shifts in the Iberian Atlantic shelf waters. Fish Oceanogr 28(3):305–316
Casimiro-Soriguer M, Junqueira R, López MA, Moreira P, Pescador E, Cabrera-Castro R, Alcântara P, Marques TJ, Hernand, JA (coord.) (2000) Bio-socioeconomic study of the sardine and anchovy fisheries in the South Atlantic Iberian Region (ICES IXa). Commission of the European comminities DGXIV/C/1–2000, pp 220
Chavez FP, Ryan J, Lluch-Cota SE, Ñiquen M (2003) From anchovies to sardines and back: multidecadal change in the Pacific Ocean. Science 299(5604):217–221
Checkley DM Jr, Asch RG, Rykaczewski RR (2017) Climate, anchovy, and sardine. Ann Rev Mar Sci 9:469–493. https://doi.org/10.1146/annurev-marine-122414-033819
Chícharo MA, Esteves E, Santos AMP, dos Santos A, Peliz Á, Ré P (2003) Are sardine larvae caught off northern Portugal in winter starving? An approach examining nutritional conditions. Mar Ecol Progr Ser 257:303–309. https://doi.org/10.3354/meps257303
Crawley MJ (2012) The R book. Wiley
De Luque L (2008) El impacto de eventos catastróficos costeros en el litoral del Golfo de Cádiz. Revista Atlántica-Mediterránea De Prehistoria y Arqueología Social 10:131–153
De Castro M, Gómez-Gesteira M, Lorenzo MN, Alvarez I, Crespo AJC (2008) Influence of atmospheric modes on coastal upwelling along the western coast of the Iberian Peninsula, 1985 to 2005. Climate Res 36(2):169–179. https://doi.org/10.3354/cr00742
Deyle ER, Fogarty M, Hsieh CH, Kaufman L, MacCall AD, Munch SB, Perretti CT et al (2013) Predicting climate effects on Pacific sardine. Proc Nat Acad Sci 110(16):6430–6435. https://doi.org/10.1073/pnas.1215506110
Diankha O, Thiaw M, Sow BA, Brochier T, Gaye AT, Brehmer P (2015) Round sardinella (Sardinella aurita) and anchovy (Engraulis encrasicolus) abundance as related to temperature in the Senegalese waters. Thalassas 31(2):9–17
Douville H (2009) Stratospheric polar vortex influence on Northern Hemisphere winter climate variability. Geophys Res Lett 36:1–5. https://doi.org/10.1029/2009GL039334
Effrosynidis D, Tsikliras A, Arampatzis A, Sylaios G (2020) Species distribution modelling via feature engineering and machine learning for pelagic fishes in the Mediterranean sea. Appl Sci 10(24):8900
Fernández IL, Báez JC, Rubio CJ, Muñoz P, Camiñas JA, Macías D (2020) Climate oscillations effects on market prices of commercially important fish in the northern Alboran Sea. Int J Biometeorol 64:689–699. https://doi.org/10.1007/s00484-020-01859-3
García del Hoyo JJ (1997) Análisis Económico de la Pesca de Cerco en la Región Suratlántica Española. Papeles De Economia Española 71:231–251
García del Hoyo JJ, Castilla Espino D, Jiménez Toribio R (2004) Determination of technical efficiency of fisheries by stochastic frontier models: a case on the Gulf of Cadiz (Spain). ICES J Mar Sci 61(3):416–421. https://doi.org/10.1016/j.icesjms.2004.02.003
García-Lafuente J, Ruiz J (2007) The Gulf of Cadiz pelagic ecosystem. A Review Prog Oceanogr 74(2–3):228–251. https://doi.org/10.1016/j.pocean.2007.04.001
García-Isarch E, García A, Silva L, Sobrino I (2003a) Spatial and temporal characterisation of the fish spawning habitat off the Guadalquivir River mouth (Gulf of Cadiz, SW Spain). In: Third International Zooplankton Production Symposium. The role of zooplankton in global ecosystem dynamics: comparative studies from the world oceans. Gijón, Spain, 20–23 May 2003, pp 64–65
García-Isarch E, Millán M, Ramos F, García-Santamaría T, Burgos C. (2012) Recent past and present of the Spanish fishery of anchovy (Engraulis encrasicolus Linnaeus, 1758) in Atlantic Moroccan waters, 441–449 pp. In: García S, Tandstad M., Caramelo AM (Ed.). Science and management of small pelagics. FAO Fisheries and Aquaculture Proceedings. FAO, Roma, pp 606
García-Isarch E, Silva L, García A, Sobrino I (2003b) Distribución espacio-temporal de la acedía Dicologlossa cuneata (Moreau, 1881) en aguas de la desembocadura del Guadalquivir (Golfo de Cádiz). Boletín Del Instituto Español De Oceanografía 19:493–503
García A, Palomera I (1996) Anchovy early life history and its relation to its surrounding environment in the Western Mediterranean basin. Scientia Marina 60(Suppl. 2):155–166
García A, Cortés D, Ramírez T, Giráldez A, Carpena A (2003) Contribution of larval growth rate variability to the recruitment of the Bay of Málaga anchovy (SW Mediterranean) during the 2000–2001 spawning seasons. Sci Mar 67:477–490. https://doi.org/10.3989/scimar.2003.67n4477
Garrido S, Silva A, Marques V, Figueiredo I, Bryère P, Mangin A, Santos AMP (2017) Temperature and food-mediated variability of European Atlantic sardine recruitment. Prog Oceanogr 159:267–275. https://doi.org/10.1016/j.pocean.2017.10.006
Guisande C, Cabanas JM, Vergara AR, Riveiro I (2001) Effect of climate on recruitment success of Atlantic Iberian sardine Sardina pilchardus. Mar Ecol Prog Ser 223:243–250. https://doi.org/10.3354/meps223243
Hall R, Erdélyi R, Hanna E, Jonesa JM, Scaifec AA (2014) Drivers of North Atlantic Polar Front jet stream variability. Int J Climatol 21:1863–1898. https://doi.org/10.1002/joc.4121
Heffernan JB, Soranno PA, Angilletta MJ Jr, Buckley LB, Gruner DS, Tim HK, Kellner JR, Kominoski JS, Rocha AV, Xiao J, Harms TK, Goring SJ, Koenig LE, McDowell WH, Powell H, Richardson AD, Stow CA, Vargas R, Weathers KC (2014) Macrosystems ecology: understanding ecological patterns and processes at continental scales. Front Ecol Environ 12:5–14
Hurrell JW (1995) Decadal trends in the North Atlantic Oscillation: regional temperatures and precipitation. Science 269(5224):676–679. https://doi.org/10.1126/science.269.5224.676
Hurrell JW, Deser C (2009) North Atlantic climate variability: the role of the North Atlantic Oscillation. J Mar Syst 78(1):28–41. https://doi.org/10.1016/j.jmarsys.2009.11.002
Hurrell JW, Kushnir Y, Ottersen G, Visbeck M (2003) An overview of the North Atlantic Oscillation. In: Hurrell JW, Kushnir Y, Ottersen G, Visbeck M (eds.). The North Atlantic Oscillation climatic significance and environmental impact. Geophysical Monograph: Washington DC, pp 1–35
ICES (2007) Report of the Working Group on the Assessment of Mackerel, Horse Mackerel, Sardine, and Anchovy. ICES Headquarters, Copenhagen, 3–4 September 2007 (ICES CM 2007/ACFM:31)
ICES (2008) Report of the Working Group on Anchovy (WGANC), 13–16 June 2008. ICES Headquarters, Copenhagen. ICES CM 2008/ACOM:04
ICES (2011) Report of the ICES Advisory Committee, 2011 ICES Advice, 2011. Book 7, 118 pp
ICES (2018) Report of the Working Group on Southern Horse Mackerel, Anchovy and Sardine (WGHANSA), 26–30 June 2018, Lisbon, Portugal. ICES CM 2018/ACOM:17. 659 pp
ICES (2020) Working Group on Southern Horse Mackerel, Anchovy and Sardine (WGHANSA). ICES Scientific Reports. 2:41–655. https://doi.org/10.17895/ices.pub.5977
Instituto Nacional de Estadística (Spain) (2020) Available: https://www.ine.es, Accessed 17 Dec 2020
Karachle P, Stergiou K (2013) Feeding and ecomorphology of three clupeoids in the N Aegean Sea. Mediterr Mar Sci 15(1):9–26. https://doi.org/10.12681/mms.350
Keller S, Valls M, Hidalgo M, Quetglas A (2014) Influence of environmental parameters on the life-history and population dynamics of cuttlefish Sepia officinalis in the western Mediterranean. Estuar Coast Shelf Sci 145:31–40
Kideys AE, Gordina AD, Bingel R, Niermann U (1999) The effect of environmental conditions on the distribution of eggs and larvae of anchovy (Engraulis encrasicolus L.) in the Black Sea. ICES J Mar Sci 56(Suppl):58–64. https://doi.org/10.1006/jmsc.1999.0605
Leitão F (2015) Time series analyses reveal environmental and fisheries controls on Atlantic horse mackerel (Trachurus trachurus) catch rates. Cont Shelf Res 111:342–352. https://doi.org/10.1016/j.csr.2015.08.026
Leitão F, Alms V, Erzini K (2014) A multi-model approach to evaluate the role of environmental variability and fishing pressure in sardine fisheries. J Mar Syst 139:128–138. https://doi.org/10.1016/j.jmarsys.2014.05.013
Lloret J, Palomera I, Salat J, Solé I (2004) Impact of freshwater input and wind on landings of anchovy (Engraulis encrasicolus) and sardine (Sardina pilchardus) in shelf waters surrounding the Ebre (Ebro) River delta (north-western Mediterranean). Fish Oceanogr 13(2):102–110. https://doi.org/10.1046/j.1365-2419.2003.00279.x
Macías D, Castilla-Espino D, García del Hoyo JJ, Navarro G, Catalán IA, Renault L, Ruiz J (2014) Consequences of a future climatic scenario for the anchovy fishery in the Alboran Sea (SW Mediterranean): a modeling study. J Mar Syst 135:150–159
Marshal J, Kushnir Y, Battisti D, Chang P, Czaja A, Dickson R, Hurrell J et al (2001) North Atlantic climate variability: phenomena, impacts and mechanisms. Int J Climatol 21:1863–1898. https://doi.org/10.1002/joc.693
Maynou F, Sabatés A, Salat J (2014) Clues from the recent past to assess recruitment of Mediterranean small pelagic fishes under sea warming scenarios. Clim Change 126(1):175–188
Mellado-Cano J, Barriopedro D, García-Herrera R, Trigo RM, Hernández A (2019) Examining the North Atlantic Oscillation, East Atlantic pattern and jet variability since 1685. J Clim 32(19):6285–6298. https://doi.org/10.1175/jcli-d-19-0135.1
Mendenhall E, Hendrix C, Nyman E, Roberts PM, Hoopes JR, Watson JR, Lam VWY et al (2020) Climate change increases the risk of fisheries conflict. Mar Policy 117:103954. https://doi.org/10.1016/j.marpol.2020.103954
Merino G, Barange M, Blanchard JL, Harle J, Holmes R, Allen I, Allison EH et al (2012) Can marine fisheries and aquaculture meet fish demand from a growing human population in a changing climate? Glob Environ Chang 22(4):795–806. https://doi.org/10.1016/j.gloenvcha.2012.03.003
Millán M (1992) Descripción de la pesquería de cerco en la región suratlántica española y atlántico norte marroquí. Informes Técnicos del Instituto Español de Oceanografia 136(3–70)
Millán M (1999) Reproductive characteristics and condition status of anchovy Engraulis encrasicolus L. from the Bay of Cadiz (SW Spain). Fish Res 41:73–86. https://doi.org/10.1016/S0165-7836(99)00010-7
Miller K, Charles A, Barange M, Brander K, Gallucci VF, Gasalla MA, Khan A, Munro G, Murtugudde R, Ommer RE, Perry RI (2010) Climate change, uncertainty, and resilient fisheries: institutional responses through integrative science. Prog Oceanogr 87(1–4):338–346. https://doi.org/10.1016/j.pocean.2010.09.014
Moore GWK, Renfrew IA (2011) Cold European winters: interplay between the NAO and the East Atlantic mode. Atmospheric Science Letters 13(1):1–8. https://doi.org/10.1002/asl.356
Morote E, Olivar MP, Villate F, Uriarte I (2008) Diet of round sardinella, Sardinella aurita, larvae in relation to plankton availability in the NW Mediterranean. J Plankton Res 30(7):807–816. https://doi.org/10.1093/plankt/fbn039
Motos L, Uriarte U, Valencia V (1996) The spawning environment of the Bay of Biscay anchovy (Engraulis encrasicolus L.). Scientia Marina 60(Suppl 2):117–140
Overland JE, Alheit J, Bakun A, Hurrell JW, Mackas DL, Miller AJ (2010) Climate controls on marine ecosystems and fish populations. J Mar Syst 79:305–315. https://doi.org/10.1016/j.jmarsys.2008.12.009
Palomera I (1992) Spawning of anchovy Engraulis encrasicolus in the Northwestern Mediterranean relative to hydrographic features in the region. Mar Ecol Prog Ser 79:215–223
Palomera I, Sabatés A (1990) Co-occurrence of Engraulis encrasicolus and Sardinella aurita eggs and larvae in the Northwestern Mediterranean. Sci Mar 54:63–69
Pérez-Rubín J, Mafalda Jr P (2004) Abnormal domination of gilt sardine (Sardinella aurita) in the middle shelf ichthyoplankton community of Gulf of Cadiz (SW Iberian Peninsula) in summer: related changes in the hydrologic structure and implications in the larval fish and mesozooplankton assemblages finded. ICES Annual Science Conference. Vigo, pp 22–25
Pérez-Rubín J, Cano N, Arrate P, García-Lafuente J, Escánez J, Vargas M, Hernández F (1997) El Ictioplancton, el holoplancton y el medio marino en el Golfo de Cádiz, Estrecho de Gibraltar y sector noroeste del Mar de Alborán, en julio de 1994. Informes Técnicos del Instituto. Español de Oceanografía 167
Pérez-Rubín J, Lucaya NC, Prieto L, García CM, Ruíz J, Echevarria F, Corzo A et al (1999) La estructura del ecosistema pelágico en relación con las condiciones oceanográficas y topográficas en el golfo de Cádiz, estrecho de Gibraltar y mar de Alborán (sector noroeste), en julio de 1995. Informes Técnicos Del Instituto Español De Oceanografía 175:3–73
R Core Team (2020) R: a language and environment for statistical computing. R Foundation for Statistical Computing. Vienna, Austria. URL: https://www.R-project.org/
Raab K, Llope M, Nagelkerke LAJ, Rijnsdorp AD, Adriaan DR, Lorna RT, Priscilla L, Piet R, Mark DC (2013) Influence of temperature and food availability on juvenile European anchovy Engraulis encrasicolus at its northern boundary. Mar Ecol Prog Ser 488:233–245
Rangel-Buitrago N, Anfuso G (2012) Winter wave climate, storms and regional cycles: the SW Spanish Atlantic coast. Int J Climatol 33(9):2142–2156. https://doi.org/10.1002/joc.3579
Rodríguez-Puebla C, Encinas AH, Sáenz J (2001) Winter precipitation over the Iberian Peninsula and its relationship to circulation indices. Hydrol Earth Syst Sci Discuss 5:233–244
Rodríguez-Puebla C, Encinas AH, Nieto S, Garmendia J (1998) Spatial and temporal patterns of annual precipitation variability over the Iberian Peninsula. Int J Climatol 18:299–316. https://doi.org/10.1002/(SICI)1097-0088(19980315)18:3%3c299::AID-JOC247%3e3.0.CO;2-L
Rodríguez-Puebla C, Encinas A, Domínguez MF, Nieto S (2002) Impacto de índices climáticos en las variaciones de precipitación acumulada en los meses de febrero, marzo y abril. In: Guijarro, Grimalt, Laita and Alonso (Eds.). El agua y el clima 3:315–323
RStudio Team (2016) RStudio: integrated development for R. RStudio, Inc., Boston, MA. URL http://www.rstudio.com/
Rubio CJ, Macías D, Báez JC (2015) Efecto de las oscilaciones atmosféricas sobre las capturas de Grandes Migradores Pelágicos con interés pesquero. Resúmenes sobre el VIII Simposio MIA15, Malaga, 21–23 September 2015
Ruiz J, Rincón MM, Castilla D, Ramos F, García del Hoyo JJ (2017) Biological and economic vulnerabilities of fixed TACs in small pelagics: an analysis of the European anchovy (Engraulis encrasicolus) in the Gulf of Cadiz. Mar Policy 78:171–180. https://doi.org/10.1016/j.marpol.2017.01.022
Sabatés ANA, Martín P, Lloret J, Raya V (2006) Sea warming and fish distribution: the case of the small pelagic fish, Sardinella aurita, in the western Mediterranean. Glob Change Biol 12(11):2209–2219. https://doi.org/10.1111/j.1365-2486.2006.01246.x
Sabatés A, Salat J, Raya V, Emelianov M (2013) Role of mesoscale eddies in shaping the spatial distribution of the coexisting Engraulis encrasicolus and Sardinella aurita larvae in the northwestern Mediterranean. J Mar Syst 111:108–119. https://doi.org/10.1016/j.jmarsys.2012.10.002
Saenz J, Zubillaga J, Rodríguez-Puebla C (2001) Interannual winter temperature variability in the north of the Iberian Peninsula. Climate Res 16:169–179. https://doi.org/10.3354/cr016169
Sánchez RF, Relvas P, Delgado M (2007) Coupled ocean wind and sea surface temperature patterns off the western Iberian Peninsula. J Mar Syst 68(1–2):103–127. https://doi.org/10.1016/j.jmarsys.2006.11.003
Santos AMP, Chícharo A, Dos Santos A, Moita T, Oliveira PB, Peliz Á, Ré P (2007) Physical-biological interactions in the life history of small pelagic fish in the Western Iberia Upwelling Ecosystem. Prog Oceanogr 74(2–3):192–209. https://doi.org/10.1016/j.pocean.2007.04.008
Schismenou E, Giannoulaki M, Valavanis VD, Somarakis S (2008) Modeling and predicting potential spawning habitat of anchovy (Engraulis encrasicolus) and round sardinella (Sardinella aurita) based on satellite environmental information. In Essential Fish Habitat Mapping in the Mediterranean. Springer, Dordrecht
Silva A, Garrido S, Ibaibarriaga L, Pawlowski L, Riveiro I, Marques V, Ramos F et al (2019) Adult-mediated connectivity and spatial population structure of sardine in the Bay of Biscay and Iberian coast. Deep Sea Res Part II 159:62–74. https://doi.org/10.1016/j.dsr2.2018.10.010
Silva A, Carrera P, Massé J, Uriarte A, Santos MB, Oliveira PB, Soares E et al (2008) Geographic variability of sardine growth across the northeastern Atlantic and the Mediterranean Sea. Fish Res 90(1–3):56–69. https://doi.org/10.1016/j.fishres.2007.09.011
Sistema de Información Andaluz de Comercialización y Producción Pesquera of the Junta de Andalucía. 2020. Available: http://www.juntadeandalucia.es/agriculturaypesca/idapes/servlet/FrontController Accessed 28 Dec 2020
Somarakis S, Drakopoulos P, Filippou V (2002) Distribution and abundance of larval fish in the northern Aegean Sea—eastern Mediterranean—in relation to early summer oceanographic conditions. J Plankton Res 24:339–357. https://doi.org/10.1093/plankt/24.4.339
Stenseth NC, Ottersen G, Hurrell JW, Mysterud A, Lima M, Chan KS, Yoccoz NG, Ådlandsvik B (2003) Studying climate effects on ecology through the use of climate indices, the North Atlantic Oscillation, El Niño Southern Oscillation and beyond. Proc R Soc Lond B 270:2087–2096. https://doi.org/10.1098/rspb.2003.2415
Straile D, Stenseth NC (2007) The North Atlantic Oscillation and ecology: links between historical time-series, and lessons regarding future climate warming. Clim Res 34:259–269. https://doi.org/10.3354/cr00702
Thompson DWJ, Wallace JM (1998) The Arctic Oscillation signature in the wintertime geopotential height and temperature fields. Geophys Res Lett 25:1297–1300
Tong H (1976) Fitting a smooth moving average to noisy data (corresp.). IEEE Transact Informat Theory 22(4):493–49
Trigo RM, Osborn TJ, Corte-Real JM (2002) The North Atlantic Oscillation influence on Europe: climate impacts and associated physical mechanisms. Climate Res 20:9–17. https://doi.org/10.3354/cr020009
Trigo RM, Pozo-Vázquez D, Osborn TJ, Castro-Diez Y, Gámiz-Fortis S, Esteban-Parra MJ (2004) North Atlantic Oscillation influence on precipitation, river flow and water resources in the Iberian Peninsula. Int J Climatol 24(8):925–944. https://doi.org/10.1002/joc.1048
Tsikliras AC (2008) Climate-related geographic shift and sudden population increase of a small pelagic fish (Sardinella aurita) in the eastern Mediterranean Sea. Mar Biol Res 4(6):477–481. https://doi.org/10.1080/17451000802291292
Uriarte A, Prouzet P, Villamor B (1996) Bay of Biscay and Ibero Atlantic anchovy populations and their fisheries. Sci Mar 60(2):237–255
Vargas-Yáñez M, Giráldez A, Torres P, González M, García-Martínez MC, Moya F (2020) Variability of oceanographic and meteorological conditions in the northern Alboran Sea at seasonal, inter-annual and long-term time scales and their influence on sardine (Sardina pilchardus, Walbaum 1792) landings. Fish Oceanogr 00:1–14. https://doi.org/10.1111/fog.12477
Vargas JM, García-Lafuente J, Delgado J (2002) Seasonal and wind-induced variability of Sea Surface Temperature patterns in the Gulf of Cadiz. J Mar Syst 38(3–4):205–219. https://doi.org/10.1016/S0924-7963(02)00240-3
Vicente-Serrano SM, Trigo RM, López-Moreno JI, Liberato MLR, Lorenzo-Lacruz J, Beguería S, Morán-Tejeda E et al (2011) Extreme winter precipitation in the Iberian Peninsula in 2010: anomalies, driving mechanisms and future projections. Climate Res 46:51–65. https://doi.org/10.3354/cr00977
Wang D, Wang C, Yang X, Lu J (2005) Winter Northern Hemisphere surface air temperature variability associated with the Arctic Oscillation and North Atlantic Oscillation. Geophys Res Lett. https://doi.org/10.1029/2005GL022952
Zarrad R, Alemany F, Jarboui O, Garcia A, Akrout F (2012) Comparative characterization of the spawning environments of European anchovy, engraulids encrasicolus, and round sardinella, sardinella aurita (Actinopterygii: clupeiformes) in the eastern coast of Tunisia. Acta Ichthyologica et Piscatoria 42(1):9–19
Zuur AF, Ieno EN, Walker N, Saveliev AA, Smith GM (2009) Mixed effects models and extensions in ecology with R. Springer, New York
Acknowledgements
Authors want to express their gratefulness to their colleagues Juan Pérez-Rubín, Víctor Sanz-Fernández, Fernando Ramos, and Marcos Llope for their helpful comments on previous drafts. Authors would also like thank Samantha Blakeman for proofreading the manuscript in English. The authors would like to thank the anonymous referee for their comments on the manuscript.
Funding
Open Access funding provided thanks to the CRUE-CSIC agreement with Springer Nature.
Author information
Authors and Affiliations
Contributions
JCB and RC-C conceived the ideas and designed methodology; JC-G collected the data; JC-G and IAC analysed the data; JC-G wrote a first draft of the manuscript. All authors contributed critically to the drafts and gave final approval for publication.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Castro-Gutiérrez, J., Cabrera-Castro, R., Czerwinski, I.A. et al. Effect of climatic oscillations on small pelagic fisheries and its economic profit in the Gulf of Cadiz. Int J Biometeorol 66, 613–626 (2022). https://doi.org/10.1007/s00484-021-02223-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00484-021-02223-9