
Abstract
Key message
Prediction of tree growth based on size or mass as proposed by the Metabolic Scaling Theory is an over-simplification and can be significantly improved by consideration of stem and crown morphology.
Tree growth and metabolic scaling theory, as well as corresponding growth equations, use tree volume or mass as predictors for growth. However, this may be an over-simplification, as the future growth of a tree may, in addition to volume or mass, also depend on its past development and aspects of the current inner structure and outer morphology. The objective of this evaluation was to analyse the effect of selected structural and morphological tree characteristics on the growth of common tree species in Europe. Here, we used eight long-term experiments with a total of 24 plots and extensive individual measurements of 1596 trees in monospecific stands of European beech (Fagus sylvatica L.), Norway spruce (Picea abies (L.) Karst.), Scots pine (Pinus sylvestris L.) and sessile oak (Quercus petraea (Matt.) Liebl.). Some of the experiments have been systematically surveyed since 1870. The selected plots represent a broad range of stand density, from fully to thinly stocked stands. We applied linear mixed models with random effects for analysing and modelling how tree growth and productivity are affected by stem and crown structure. We used the species-overarching relationship \(\mathrm{iv}={{a}_{0}\times v}\) between stem volume growth, \(\mathrm{iv}\)and stem volume, \(v,\) as the baseline model. In this model \({a}_{0}\) represents the allometric factor and α the allometric exponent. Then we included tree age, mean stem volume of the stand and structural and morphological tree variables in the model. This significantly reduced the AIC; RMSE was reduced by up to 43%. Interestingly, the full model estimating \(\mathrm{iv}\) as a function of \(v\) and mean tree volume, crown projection area, crown ratio and mean tree ring width, revealed a \(\alpha \cong 3/4\) scaling for the relationship between \(\mathrm{iv}\propto {v}^{\alpha }\). This scaling corresponded with Kleiber’s rule and the West-Brown-Enquist model of the metabolic scaling theory. Simplified approaches based on stem diameter or tree mass as predictors may be useful for a rough estimation of stem growth in uniform stands and in cases where more detailed predictors are not available. However, they neglect other stem and crown characteristics that can have a strong additional effect on the growth behaviour. This becomes of considerable importance in the heterogeneous mixed-species stands that in many countries of the world are designed for forest restoration. Heterogeneous stand structures increase the structural variability of the individual trees and thereby cause a stronger variation of growth compared with monocultures. Stem and crown characteristics, which may improve the analysis and projection of tree and stand dynamics in the future forest, are becoming more easily accessible by Terrestrial laser scanning.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
In the past, many works have been published about the course of tree growth and its dependency on tree age or tree size (von Bertalanffy 1951; Richards 1959; Coomes and Allen 2007). Tree growth theory (von Bertalanffy 1951) and corresponding growth equations (Zeide 1993) used tree volume or mass as predictors for growth. Current stem volume or stem mass represent the accumulated past growth of a tree and are certainly better predictors for its current growth than tree age. Holding all other factors fixed, the bigger the present size of a tree the more extended the meristems for building new cells and the higher the growth rate. Thus the current size represents, together with other variables, the ecological legacy of the past individual tree growth. Other characteristics, such as the past tree ring pattern (Camarero et al. 2018) or crown morphology (Mäkelä and Valentine 2006), may further determine a tree’s growth and vitality in the future. An estimation of the metabolic rate and growth of a tree, only based on body size or mass, as proposed by the metabolic scaling theory and model by West, Brown and Enquist (1997) (WBE model), may be useful for a rough species-overarching estimation of stem growth in uniform stands and if more detailed predictors are not available. The WBE model proposed the 3/4 scaling between plant growth and plant mass. It has been derived from the fractally structured internal pipe system of plants (West et al. 1997; Enquist et al. 1998). Based on the 3/4 scaling of allometric ideal plants Enquist et al. (2009) and West et al. (2009) later extended their scaling theory to the stand level and the self-thinning under demographic equilibrium conditions. Among others, Kozłowski and Konarzewski (2004) and Muller-Landau et al. (2006) argued that the WBE model is an over-simplification of tree allometry. Especially in heterogeneous stands with a wide variation of stem and crown allometries, tree growth may be co-determined by other tree attributes in addition to tree mass (Pretzsch 2014; Pretzsch et al. 2015).
A huge leap forward was the inclusion of individual tree-specific structural and morphological information for growth prediction. Well know exemplars for this approach are eco-physiological models (Landsberg 2011) and, in particular, structural–functional models (Sievänen et al. 2000, Grote and Pretzsch 2002) which use e.g., sapwood area, leaf area, or crown length as predictors of tree growth. Whereas eco-physiological processes and functions are often much more difficult to measure and less representative, structural and morphological traits are easier to access and may support the bridging of process-based and empirical approaches to modelling tree growth (Mäkelä and Valentine 2006). Structural and morphological traits, such as crown projection area, crown ratio and mean tree ring width, may contain ecological legacy information about the past of the tree that is relevant for its present and future growth. It can be made use of the fact that in seasonal forests tree ring pattern and crown structure store information about growth rhythm in trees (Lüttge and Hertel 2009). The tree ring pattern or crown structure can be harnessed for quantifying the tree’s past development by appropriated metrics (Pretzsch 2021). Tree-ring, crown, or root morphology patterns represent a structural legacy imbedded in stem, crown, and root (Netzer et al. 2019; Ogle et al. 2015). This legacy may affect the tree’s functioning and growth, e.g., via light interception, hydraulic conduction, or water and nutrient uptake. In this way, the differences in structure may cause specific differences in the functioning and growth curve patterns. Such information about structural and morphological traits is often available for a larger number of trees and can be used for model parameterization or evaluation.
The objective of this study was to analyse the effects of selected structural and morphological characteristics on the growth of common tree species. Such detailed structural and morphological traits may contain information of the past growth that is “ecologically memorised” by the tree and determines its present and future growth (Berger and Hildenbrandt 2000; Camarero et al. 2018). Based on individual tree growth and allometric stem and crown characteristics of European beech (Fagus sylvatica L.), Norway spruce (Picea abies (L.) Karst.), Scots pine (Pinus sylvestris L.), and sessile oak (Quercus petraea (Matt.) Liebl.), we answer the following three questions:
-
Q1: how does tree volume growth depend on tree volume and tree age in general and at defined tree ages or in defined stand development phases?
-
Q2: how do stem and crown characteristics (e.g., mean annual growth in the past, crown ratio, crown projection area) determine individual tree growth?
-
Q3: how does the growing area efficiency (tree growth per crown projection area) depend on tree size and various stem and crown characteristics?
Finally, we discuss the implications of structure-growth relationships for forest ecology and management, and for silvicultural guidelines.
Material and methods
Location and site characteristics of the selected stands
We used eight long-term experiments with 24 plots and extensive individual measurements of 1596 trees in monospecific stands of European beech, Norway spruce, Scots pine, and sessile oak (Table 1). The experiments belong to a research network, which was established by far-sighted researchers in the late nineteenth century to procure growth and yield data as a quantitative basis for sustainable forest management (described, among others, by von Ganghofer (1881)). In the beginning, measurements on these experiments only included stem diameter, tree height and tree removal (Verein Deutscher Forstlicher Versuchsanstalten 1873, 1902). Later also tree coordinates, crown dimensions, and tree vitality have been measured (Pretzsch 2017; Pretzsch et al. 2019). Some of the experiments selected for this study have been systematically surveyed since 1870. The selected 24 plots (three plots per experiment) represent a broad range of stand density, from fully to thinly stocked stands. Most of the stands were established by regular planting. To cover different tree ages, we selected one experiment in an earlier and one in an advanced stand development phase for each tree species. For details of the locations and site characteristics see Table 1.
The eight experiments represent growth conditions in the plains and highlands of Central and Southern Germany (Table 1). The experiments are located between 400 and 780 m a.s.l. The long-term mean temperature and mean annual precipitation exhibit a broad range of climate conditions (6.8–8.0 °C and 680–1120 mm year−1, respectively). The distribution of the experiments over five eco-regions and seven geological zones is reflected in the spectrum of soil types. The poorest soils are pseudogley soils derived from Coburg sandstone, while the most fertile soils are parabrown soils from diluvial loess-loam in the pre-alpine highlands. Located mainly between Frankfurt, Regensburg and Garmisch-Partenkirchen, the majority of the experimental stands are stocking on soils of mediocre fertility in the Southern German gradual layer area “Schichtstufenlandschaft” and on soils of rich fertility in the pre-alpine highlands.
Table 2 provides an overview of all tree and stand variables, constants and coefficients used in this study. According to the standard of the International Union of Forest Research Organizations (Johann 1993; Kramer and Akça 1995) we used lowercase letters for tree variables and uppercase letters for stand variables.
Repeated surveys resulted in the data suitable for calculation of all common stand characteristics for each of the up to 21 successive survey periods. Table 3 gives an overview of the stand characteristics of the last survey. The reported stand-level data were derived from the successive inventories of the tree diameters, tree heights, and records of the removal trees. As explained in more detail further below, we included unthinned plots, where we recorded the natural mortality, and we also included thinned plots where we recorded the trees removed by heavy thinning. We used standard evaluation methods according to the DESER-norm recommended by the German Association of Forest Research Institutes (in German “Deutscher Verband Forstlicher Forschungsanstalten”) (Johann 1993; Biber 2013). For estimating the merchantable stem volume in dependence on tree diameter, tree height and form factor, we used the approach by Franz et al. (1973) with the equations and coefficients published by Pretzsch (2002, p. 170, Table 7.3) Table 4.
Each of the four species was represented by experimental plots in young and mature stands. The stand ages ranged from 44 to 188 years at the time of the last survey (Table 3). From each of the eight experiments, we included plots without thinning, with moderate, and with heavy thinning. The plots without active thinning represented the stand development under self-thinning conditions. Compared with the stand basal area of the unthinned plots (100%), the density of the moderately thinned plots was kept at a level of about 70%. The stand basal area on the heavily thinned plots was kept at a level of about 50%. The setpoint stand basal areas of about 70 and 50% of the unthinned plots were achieved by continuously removing trees in the course of the successive surveys. The broad variation of stand ages and stand densities covered by the 24 plots (four species \(\times\) two stand ages \(\times\) three stand density variants) was important for the representativeness of the results. We were aiming at general species-, age-, and stand density-overarching results and tried to avoid a case study character, focussed on one or just a few selected stands, only.
Table 3 reports the mean stand characteristics of the three differently thinned plots per each experiment. To quantify the site quality of the included experimental stands we used the site index based on the species-specific yield tables by (Assmann and Franz 1963, 1965); Wiedemann (1943); (Schober 1967, 1975) and Jüttner (1955). Based on the stand ages and the dominant tree heights of the last survey and based on the height-age-relationships of the abovementioned yield tables we calculated the dominant heights of the stand age of 100 years by extrapolation. This standard procedure of site indexing (Skovsgaard and Vanclay 2008) resulted in site indices of 24.2–39.4 m at age 100. Notice, that the site indexes are rather similar within the pairs of young and old stands of each species. Certainly, the tree number per hectare decreases with age and is much higher for conifers with mostly slim crowns compared with deciduous species with more extended and more space requiring crowns. Mean stem diameters ranged between 14.4 and 58.6 cm and mean heights between 15.0 and 40.3 m. The standing stem volumes reached maximum values of 1404 m3ha−1 at the last survey. In the last survey periods, the mean periodic stem volume growth ranged between 6.7–24.5 m3ha−1year−1 and the total yield until the last survey was 767–2141m3ha−1. The density on the three plots per site ranged between unthinned conditions to heavy thinning. Thus the plots represent fully to thinly stocked conditions. All stands were without canopy gaps, caused by damages.
The broad variation of growth and yield characteristics due to species, age and site conditions was intentional, as we aimed at generalisable results regarding the effect of the stem and crown structure on individual tree growth. In summary, the dataset covered ecologically different tree species (light demanding to shade tolerant), different site conditions (average to excellent sites), different tree ages (middle-aged to old), varying stand densities (unthinned to heavily thinned), and a broad range of social tree positions (dominated to predominant trees). Any spatial and temporal autocorrelations due to the nested data levels, experiment, plot, and tree, were taken into account by the random effects in models 1–8 (see Sect. “Statistical analyses and models”).
Repeated measurements at the tree and stand level
In addition to the diameter at breast height (\({d}_{1.3}\), cm) and tree height (\(h\), m), the height to crown base (\(\mathrm{hcb}\), m) and the crown radii in eight cardinal directions were measured according to standards described by Pretzsch (2009, pp. 115–118). All variables were repeatedly measured in the past. Based on these variables we calculated the crown length (\(\mathrm{cl}\), m) and crown ratio (\(\mathrm{cr}\), m) (\(\mathrm{cr}=\mathrm{cl}/h\)). The eight radii were used to calculate the crown projection area in m2 \(\mathrm{cpa}={\stackrel{-}{\mathrm{cr}}}^{2}\times\) with \({\stackrel{-}{\mathrm{cr}}}^{2}=\sqrt{({r}_{1}^{2}+{r}_{2}^{2}+\dots +{r}_{8}^{2})/8}\). We further calculated the mean tree ring width in cm/year as \(\mathrm{mrw}={d}_{1.3}/2/\mathrm{tree\,age}\) (Fig. 1). These were the main structural and morphological variables that we used as predictors for estimating the tree growth in the respective following periods. For overview of the individual tree characteristics on the eight long-term experiments see Table 4.

Visualisation of variables used in this study for quantifying tree structure and crown morphology. d1.3 stem diameter at height 1.3 m, h tree height, hcb height to crown base, cl crown length, cr crown radius, cpa crown projection area, mrw mean ring width (= d1.3/2/tree age)
Statistical analyses and models
To analyse the effect of characteristics such as tree age, mean ring width and crown size on tree growth, we applied linear mixed effect models with nested random effects. In this way, we account for any spatial and temporal autocorrelation effects. The fixed effect variables such as stem volume, tree age, crown projection area, crown ratio, and mean ring width represented the influence of the trees' past and present characteristics on its growth. The fixed effects were covered by the parameters \({a}_{0}\)−\({a}_{\mathrm{n}}\). The random effects on \({a}_{0}\) (intercept) at the experiment, plot, survey, and tree level took into consideration any spatial (several plots per experiment, several trees per plot) and temporal (several successive surveys, repeated measurements at the same tree) autocorrelations. The random effect \({b}_{\mathrm{i}}, {b}_{\mathrm{ij}}, {b}_{\mathrm{ijk}}, {b}_{\mathrm{ijkl}}\) covered the level of the experiment, the plot, the survey and the tree. We described the respective model alternatives and selected the variable combinations based on the root mean square error, RMSE, and the AIC criterion (Akaike 1981). The following numbers of the models refered to the results in the text (Models 1–8). In Table 5 we restricted the results to the characteristics of the fixed effects.
Model 1
was fitted to annual stem volume increment, iv, of trees on long-term experiments with known stem volume, v, at the beginning of the respective measurement period. This model reflected the general dependency of tree growth on tree size. By pooling the data of all four tree species and different tree and stand development phases we addressed the overarching allometric relationship \(iv\sim {v }^{\alpha }\).
Model 2
Using this model we analysed the relationship between stem volume growth and stem volume for each plot and survey separately. In this way, we analysed the drift of the \(\mathrm{iv}\sim {v }^{\alpha }\) allometry with progressing stand development (see Supplement Table 1).
Models 3.1 and 3.2
Models 4.1 and 4.2
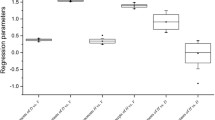
The models 3a and 3b and 4a and 4b revealed the drift of the allometric factor a and allometric exponent α with progressing stand development. The progressing stand development was represented by stand age and mean stem volume of the stand, respectively.
Model 5
Model 5 was similar to model 1 but included tree age as a predictor variable. Using this model we tried to reveal how the allometric \(\mathrm{iv}\sim {v }^{\alpha }\) relationship was co-determined by tree age.
Model 6
Model 6 was similar to model 2 but included mean stem volume as a predictor variable instead of tree age. Using this model we tried to reveal how the allometric \(\mathrm{iv}\sim {v }^{\alpha }\) relationship was co-determined by mean tree stem volume as a proxy for the stand development phase.
Model 7
By this model, we tried to reveal how the allometric \(\mathrm{iv}\sim {v }^{\alpha }\) relationship was co-determined by mean tree stem volume (as a proxy for the stand development phase), by crown projection area and crown ratio (as indicators for the current tree structure), and by mean ring width (as a proxy for the tree’s past growth).
Models 8.1–8.3
These three models represented auxillary relationships which reflected how crown projection area, crown ratio and mean tree ring width depended on the current stem volume. By insertion of these auxillary allometric relationships (relationships between cpa and v, cr and v, and mrw and v) model 7 may be transformed to model 1, and vice versa.
The statistical software R 3.4.1 (R Core Team 2018) was used for all calculations, in particular the function lme from the package nlme (Pinheiro et al. 2018).
Results
Stem volume growth depending on stem volume and tree age (Q1)
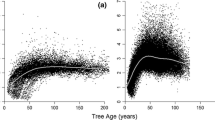
The regression analysis of the relationship between stem volume growth and stem volume (model 1) yielded an allometric exponent \({a}_{1}=1.44\pm 0.02\) (see Fig. 2a and Table 5). Interestingly this is not at all in accordance with the general \(\alpha =3/4\) scaling rule proposed by Kleiber (1947) and the metabolic scaling theory by West et al. (1997).

Dependency of stem volume increment, \(\mathrm{iv}\), on stem volume and tree age at the beginning of the respective growth periods visualised for a pooled dataset of Norway spruce, Scots pine, European beech and sessile oak. a Measurement and model of the \(\mathrm{iv}\sim v\) relationship (see Table 5, model 1), b overall relationship \(\mathrm{iv}\sim v\) compared with the \(\mathrm{iv}\sim v\) relationship of each included survey period. The black dots represent the mean iv and mean v values for each survey, c allometric factor \({a}_{\mathrm{t}}^{^{\prime}}\) and d allometric exponent \({\alpha }_{\mathrm{t}}\) plotted over tree age; the relationship \({a}_{\mathrm{t}}^{^{\prime}}\sim \mathrm{age}\) was highly significant (solid black line in (c)) whereas their was no significant relationship \({\alpha }_{\mathrm{t}}\sim \mathrm{age}\) (broken black line in (d))
To trace the \(\mathrm{iv}\sim v\) relationship to the level of the single survey and the inter-individual relationship between \(\mathrm{iv}\) and \(v\) at a given point in time, we fitted the regression \(\mathrm{ln}\left(\mathrm{iv}\right)={a}_{\mathrm{t}}+{ \alpha }_{\mathrm{t}}\times \mathrm{ln}(v)\) to the stem growth and stem volume data of every individual survey separately (see model 2 and statistical characteristics in Supplement Table 1). The results were visualised by the delogarithmic relationships \(\mathrm{iv}={a}_{\mathrm{t}}\times {v}^{{\alpha }_{\mathrm{t}}}\) in Fig. 2b. The temporal \(\mathrm{iv}\sim v\) relationships were also progressive (\({\alpha }_{\mathrm{t}}\gg 1\)). Obviously at a given point in time, the tall trees benefit overproportionally in growth due to their preferential access to light and their outcompeting effect on lower neighbours. Supplement Table 1 shows that all temporal inter-individual \(\mathrm{iv}\sim v\) relationships were highly significant regarding their intercepts \({a}_{\mathrm{t}}\) and slopes \({\alpha }_{\mathrm{t}}\). The slopes ranged mainly between \({\alpha }_{\mathrm{t}}=0.99- 1.85\).
Both \({a}_{\mathrm{t}}\) and \({\alpha }_{\mathrm{t}}\) were used for exploring the development of these coefficients with progressing stand development via models 3.1 and 3.2. The results were shown in Fig. 2c, d and Table 5. We also carried out analogous regression analyse on the basis of the variable \({v}_{\mathrm{mean}}\) instead of tree age. The calculations yielded similar results as the regressions analyses based on tree age (see Supplement Fig. 2).
We found a highly significant (p < 0.001) negative correlation between the allometric factor \({a}_{\mathrm{t}}\) and tree age (corr = − 0.94) and also between the allometric factor \({a}_{\mathrm{t}}^{^{\prime}}\) and mean tree volume, \({v}_{\mathrm{mean}}\), (corr = − 0.80) (Fig. 2c and Supplement Fig. 2a). In contrast, the allometric exponent \({a}_{\mathrm{t}}^{^{\prime}}\) was independent of both tree age (Fig. 2d and Supplement Fig. 2b) and mean tree volume. The respective Pearson correlation coefficients were r = 0.24 and r = 0.14 (both non-significant).
The results of the \({a}_{\mathrm{t }}\sim \mathrm{ age}\) and \({\alpha }_{\mathrm{t}} \sim \mathrm{ age}\) regression and the analogous regressions based on \({v}_{\mathrm{mean}}\) were shown in Table 5 and Supplement Fig. 2. Both evaluations showed that the allometric factor may decrease with tree age. This means that the scaling remained similar but the level of the curves decreased with increasing age (Fig. 2c, d, Supplement Fig. 2).
To scrutinize the contribution of tree age or mean tree volume to the estimation of \(\mathrm{iv }\)we analysed the relationships \(\mathrm{iv}=f(v, \mathrm{age})\) and \(\mathrm{iv}=f(v, {v}_{\mathrm{mean}})\), in addition to the baseline relationship \(\mathrm{iv}=f(v)\) (see models 5 and 6). For model parameters see Table 5. The RMSE was 0.67 in case of \(\mathrm{iv}=f(v)\), 0.48 for \(\mathrm{iv}=f(v, \mathrm{age})\), and 0.40 also for \(\mathrm{iv}=f(v, {v}_{\mathrm{mean}})\). Thus the RMSE could be reduced by 28–40% by adding the tree age or mean tree volume as a predictor. It was an important finding that mean tree size had a similar predictive power as tree age (see Supplement Fig. 2 and Table 5). Tree or stand age is often not available but mean tree volume is rather easy accessible and mostly available when monitoring, inventorying, or modelling tree and stand growth.
Figure 3a showed the dependency of stem volume increment, \(\mathrm{iv}\), on stem volume and tree age at the beginning of the respective growth periods. Here, we visualised the results of the overarching evaluation for all species and how it developed with progressing tree age (see Table 5, Model 5). Figure 3b showed the dependency of stem volume increment, \(\mathrm{iv}\), on stem volume and mean stem volume at the beginning of the respective growth periods (see Table 5, Model 6).

Dependency of stem volume increment, iv, on a stem volume and tree age and b stem volume and men stem volume of the stand at the beginning of the respective growth periods. Overarching analysis for all species. a Model of the \(\mathrm{iv}\sim v, \mathrm{age}\) relationship (see Table 5, model 5) for tree ages of 50, 100, and 150 years, and b model of the \(\mathrm{iv}\sim v,{v}_{\mathrm{mean}}\) relationship (see Table 5, model 6) for stands with mean tree volume of vmean = 1, 2, and 3 m3
Stem volume growth depending on stem and crown characteristics (Q2)
Based on the model \(\mathrm{iv}=f(v, {v}_{\mathrm{mean}})\), we included variables of the individual tree structure and morphology to further improve the prediction of \(\mathrm{iv}\). The inclusion of \(\mathrm{cpa}\), \(\mathrm{cr}\), and \(\mathrm{mrw}\) (see Table 5, model 7) showed the best results regarding the reduction of the AIC criterion and RMSE. Compared with the baseline relationship (\(\mathrm{iv}=f(v)\)), the relationships \(\mathrm{iv}=f(v, {v}_{\mathrm{mean}})\) and \(\mathrm{iv}=f(v, {v}_{\mathrm{mean}}, \mathrm{cpa}, \mathrm{cr},\mathrm{ mrw})\) further reduced the AIC and RMSE, respectively. Interestingly, the tree species added as categorial variable, and the site index did not significantly contribute to the model.
Figure 4 visualised the full model (see Table 5, model 7) \(\mathrm{iv}=f(v, {v}_{\mathrm{mean}},\mathrm{ cpa}, \mathrm{cr},\mathrm{mrw})\). According to this overarching model, the exponential relationship \(\mathrm{iv}\propto {v}^{3/4}\) remained close to \(\alpha \cong 3/4\) during the whole stand development. However, stem structure and crown morphology modified this basic relationship as shown in Fig. 4. With progressing stand development, indicated by \({v}_{\mathrm{mean}}\) in Fig. 4a, individual growth of trees with defined stem volume decreased (indicated by a2 = − 0.34 in model 7, see Table 5). The crown projection area had a strong positive effect on tree volume growth (Fig. 4b) (indicated by a3 = 0.21 in model 7, see Table 5). Increasing crown ratios slightly decreased stem volume growth (Fig. 4c) (indicated by a4 = − 0.18 in model 7, see Table 5). The past mean tree ring width, \(\mathrm{mrw}\), indicated the tree’s growth potential (combination of site index, neighborhood conditions, inner stem hydraulic). The variable \(\mathrm{mrw}\) had a strong positive effect on tree volume growth (Fig. 4d) (indicated by a5 = 1.29 in model 7, see Table 5). Notice, that when showing the effects of different structural and morphological traits on growth in Fig. 4a, b, c, d, we set the other variables constant on their overall mean level as provided by the dataset.

Dependency of stem volume increment, iv, on stem and crown characteristics shown for Norway spruce, Scots pine, European beech and sessile oak in one overarching evaluation (see underling model 7 in Table 5). Stem volume growth plotted over the individual stem volume (a) for stands with different men tree volumes as an indication for progressing stand development, (b) for trees with different crown projection areas, cpa, (c) for trees with different crown ratios, cr, and (d) for trees with a different mean ring with, mrw, in their development so far
In summary, when analysing Q2 we started with the \(\mathrm{ln}(\mathrm{iv})\sim \mathrm{ln}(v)\) relationship (see model 1, Table 5) as a baseline model with \(\mathrm{AIC}=1830\) and a root mean square error of the prediction of \(\mathrm{RMSE}=0.67\). Inclusion of the tree age (see model 2, Table 5) reduced both, the AIC to 1807 and RMSE to 0.40, which meant a strong reduction of the RMSE by 39%. Inclusion of the mean stem volume vmean (see model 3, Table 5) also reduced both, the AIC to 1820 and RMSE to 0.48, which meant a reduction of the RMSE by 29% compared with the baseline model 1. The use of vmean as a predictor was of special interest, as in contrast to the tree age, the mean stem volume is more readily available as a predictor. Inclusion of structural and morphological tree attributes reduced both statistical measures to \(\mathrm{AIC}=1761\) and \(\mathrm{RMSE}=0.38\). In the case of RMSE this means a reduction by 43% compared with the baseline model \(\mathrm{ln}(\mathrm{iv})\sim \mathrm{ln}(v)\).
Stem growth per crown projection area (Q3)
The transition from individual tree growth to productivity shown in Figs. 4, 5 was based on the relationships \(\mathrm{iv}=f(v, {v}_{\mathrm{mean}}, \mathrm{cpa}, \mathrm{cr}, \mathrm{mrw})\) and \(\mathrm{iv}/\mathrm{cpa}=f(v, {v}_{\mathrm{mean}},\mathrm{ cpa}, \mathrm{cr}, \mathrm{mrw})\). By rearrangement of model 7 \(\mathrm{iv}={e}^{{a}_{0}}\times {v}^{{a}_{1}}\times {v}_{\mathrm{mean}}^{{a}_{2}}\times {\mathrm{cpa}}^{{a}_{3}}\times {\mathrm{cr}}^{{a}_{4}}\times {\mathrm{mrw}}^{{a}_{5}}\), we arrived at the equation.

Dependency of tree growth per crown projection area, iv/cpa, on stem volume and crown characteristics according to a species-overarching evaluation for Norway spruce, Scots pine, European beech and sessile oak (model 7). a iv/cpa depending on stem volume, shown for various levels of cpa. b iv/cpa depending on cpa, shown for various levels of v. For (a, b) cpa and v were varied and the other independent variables vmean, mrw, and cr were set constant. Variables are v stem volume, cpa crown projection area, vmean mean stem volume of the stand, mrw mean ring width, cr crown ratio
\(iv/\mathrm{cpa}={e}^{{a}_{0}}\times {v}^{{a}_{1}}\times {v}_{\mathrm{mean}}^{{a}_{2}}\times {\mathrm{cpa}}^{{a}_{3}}\times {\mathrm{cr}}^{{a}_{4}}\times {\mathrm{mrw}}^{{a}_{5}}/\mathrm{cpa}\). The latter equation represented a description of tree productivity in dependence on stem and stand attributes. Its visualisation (Fig. 5) showed how the crown projection area affected tree productivity (Fig. 5a, b). For this evaluation, we kept \(\mathrm{mrw}\) and \(\mathrm{cr}\) constant at the overall mean level (\(\mathrm{mrw}\) = 0.2, = \(\mathrm{cr}\) 0.3). The figure visualized that in general the smaller the crown projection area of trees, the higher their growing area efficiency. Figure 5a showed the \(\mathrm{iv}\)/\(\mathrm{cpa}\) plotted over \(v\) for different \(\mathrm{cpa}\)-level. It revealed that productivity increased on average with increasing stem volume. However, the level was lower in the case of trees of large crown compared to small-crowned trees. Figure 5b corroborated this finding by showing the continuous decrease of tree productivity with increasing \(\mathrm{cpa}\) values. This decrease applied for trees of all stem volume level. We found no interaction that would suggest a steeper decrease of productivity for larger stems compared to smaller ones.
On the one hand, we revealed a progressive relationship (\(\gg 1\)) between \(\mathrm{iv}\) and v, [\(\mathrm{iv}=f(v)\)] by model 1 (see Fig. 2). On the other hand, we found a degressive relationship between \(iv\) and other tree attributes in addition to v, \(\mathrm{iv}=f\left(v, {v}_{\mathrm{mean}}, \mathrm{mrw}, \mathrm{cpa}, \mathrm{cr}\right)\) using model 7 (see Fig. 4). These two relationships are not contradictive. By insertion of the basic allometric relationships between cpa and v, cr and v, and mrw and v (Supplement Fig. 3) the degressive relationship of model 7 may be transformed to the progressive relationship of model 1.
Discussion
Stem structure and crown morphology measurements can improve the understanding, modeling, and prediction of tree growth and can bridge the gap between statistical and ecoyphysiological modelling approaches (Mäkelä and Valentine 2006). Classical approaches of growth projection were mainly based on tree age (Richards 1959; Assmann and Franz 1965). Simple allometric approaches and metabolic scaling at least included the current total size or mass in the prediction of future growth (King 2005; Nord-Larsen and Johannsen 2007). However, generic allometric relationships neglected other tree attributes that better consider the individual tree's status, growing space, or crown structure (e.g., Coates et al. 2003). Individual eco-physiological models (Korol et al. 1995; Grote and Pretzsch 2002) and structural–functional models (Lacointe et al. 2000; Sievänen et al. 2008) conceptualized tree growth based on basic physiological mechanisms such as photosynthesis, respiration, partitioning, and allometric principles (Mäkelä et al. 2000; Landsberg 2003; Valentine and Mäkelä 2005). Latter models for the estimation of tree growth were often calibrated by annual growth rates at the tree level since information of carbon and nutrient flows as well as forest structure at higher levels of the resolution were rare (Jonard et al. 2020). This study emphasized that various aspects of stem and crown structure and morphology which strongly affect tree growth may deserve special consideration when building, initiating, calibrating or evaluating individual tree models.
This study revealed a high significance of selected structural and morphological stem and tree characteristics for the growth of common tree species in Central Europe. For this purpose, we started with the models \(\mathrm{iv}=f\left(v\right)\) and \(\mathrm{iv}=f\left(v, {v}_{\mathrm{mean}}\right)\), which only took into account the stem volume for the prediction of tree growth in the following period. The surveys in different stand development phases showed a strong inter-individual variation of \(\mathrm{iv}\) due to size-asymmetric competition. The rather simple double logarithmic equation \(\mathrm{ln}\left(\mathrm{iv}\right)={a}_{0}+\times \mathrm{ln}\left(v\right)\), which becomes \(\mathrm{iv}={a}_{0}^{^{\prime}}\times {v}^{\alpha }\), explained about 60% of the variation of annual growth (\(\mathrm{iv}\)). However, these relationships considered neither the stand structure and species nor the individual tree’s structural and morphological attributes. So, they also neglected information about the past of the tree, e.g., information about the tree structure and morphology as adapted to the social status of the tree within the stand.
Our sequence of models for estimating tree volume growth, \(\mathrm{iv}\), showed a continuously increasing accuracy from models including only stem volume, to models with mean stem volume or tree age, to the model including crown projection area, crown ratio and mean tree ring width. This underlined the relevance of structure and morphology for growth prediction. The inclusion of stem and crown information reduced the RMSE by 43% compared with the bivariate scaling. The reduction of AIC and the RMSE after the inclusion of further tree characteristics indicated that those attributes represent important additional information, affecting tree growth. We hypothesised that the structural and morphological traits contain relevant information of a tree’s past that are relevant for current and future growth. Interestingly, the inclusion of tree species via a categorial variable and site index did not improve the accuracy. Apparently, the structural and morphological traits already contain such information.
Inclusion of the crown projection area, \(\mathrm{cpa}\), crown ratio, \(\mathrm{cr}\), and mean ring width, \(\mathrm{mrw}\), even changed the relationship between tree attributes and growth from progressive to degressive (compare Fig. 3 with 4). Interestingly, the inclusion of further stem and crown characteristics beyond stem volume led to a scaling close to \(=3/4\) (\(\mathrm{iv}\propto {v}^{3/4}\)) (see Table 5, model 7, regression coefficient \({a}_{1}=0.77\pm 0.084\)). This meant that the revealed scaling was close to the overarching \(=3/4\), predicted by the constant of Kleiber (1947) and the metabolic scaling theory by West et al. (1997). Notice that the underlying regression analysis was based on 1596 trees of various ages, tree species, and stand densities; i.e., the resulting coefficients were quite well substantiated by data. This meant that the 3/4 scaling was modified by other stem and crown characteristics (Fig. 4). This substantiated the notion, that for a rough estimation of tree growth, the 3/4 scaling approach may suffice. However, for a more accurate estimation of tree growth the full model, encompassing a broader set of tree variables, representing the past and present state of structure and morphology of the tree, should be applied.
For analysing and predicting growth at the tree or stand level, information about the stand or tree characteristics (e.g., by classical inventory or TLidar) might be applied for a more accurate estimation of the growth depending on stand and tree characteristics. Stand characteristics such as age or mean tree size of the stand may be sufficient for growth predictions in uniform monospecific stands. However, individual tree information such as crown width, crown length or relative tree size may become even more relevant in heterogeneous stands with trees varying strongly in social status, crown sizes and growth (Pretzsch and Rais 2016; Pretzsch 2019).
Tree productivity can be calculated based on annual stem volume growth per crown projection area, \(\mathrm{cpa}\). Cpa may be used as a substitute for the growing area (Webster and Lorimer 2003). Crown projection area translated tree growth (\(\mathrm{iv}\)) to productivity (\(\mathrm{iv}\)/\(\mathrm{cpa}\)). As the \(\mathrm{cpa}\) increases continuously with stem volume, the productivity (\(\mathrm{iv}\)/c\(\mathrm{pa}\)) peaks and decreases earlier than \(\mathrm{iv}\). In contrast to tree growth, tree productivity decreased continuously with tree size. As crowns may overlap or stay in distance to each other (Pretzsch 2014), the real tree growing area may be lower or higher than the \(\mathrm{cpa}\). This may change the level of the relationship between stem and crown size, but not the continuous increase of \(\mathrm{cpa}\) with stem size. When concluding that big trees actively fix larger amounts of carbon compared to smaller trees (e.g., Stephenson et al. 2014), one should consider those differences between growth per individual and growth per unit of growing area (i.e., productivity). As the growing area requirement per tree continuously increases with size, tree productivity and contribution to carbon sequestration per unit area decreases although the growth may continue until advanced age and big size (Schütz 2002; Sillett et al. 2015).
Conclusions
The species- and site-overarching analyses of tree growth data showed that stem volume as a predictor allowed only a rough estimation of tree growth. However, the inclusion of stem structure and crown morphology variables significantly improved the prediction of tree growth. Simplifying approaches that mainly rely on stem diameter or tree mass for growth estimation may be useful for rough species-overarching upscaling in uniform stands and in cases where more detailed predictors are not available. Stem and crown characteristics, and maybe other internal stem traits (e.g., heartwood area portion, embolism, narrow ring series caused by damages or suppression phases) may become of considerable importance for assessing and better predicting tree growth in the more heterogeneous stands of the future, where tree structure varies and may cause a strong variation of the course of growth.
Author contributions statement
HP initiated and conceptualised the study, evaluated the data, wrote, and revised the manuscript.
References
Akaike H (1981) Likelihood of a model and information criteria. J Econom 16(1):3–14. https://doi.org/10.1016/0304-4076(81)90071-3
Assmann E, Franz F (1963) Vorläufige Fichten-Ertragstafel für Bayern. Forstl Forschungsanst München, Inst Ertragskd, p 104
Assmann E, Franz F (1965) Vorläufige Fichten-Ertragstafel für Bayern. Forstw Cbl 84(1):13–43
Berger U, Hildenbrandt H (2000) A new approach to spatially explicit modelling of forest dynamics: spacing, ageing and neighbourhood competition of mangrove trees. Ecol Model 132(3):287–302
Biber P (2013) Kontinuität durch flexibilität. Standardisierte datenauswertung im rahmen eines waldwachstumskundlichen informationssystems. Allgemeine Forst Jagdzeitung 184(7):167–177
Camarero JJ, Gazol A, Sangüesa-Barreda G, Cantero A, Sánchez-Salguero R, Sánchez-Miranda A, Ibáñez R (2018) Forest growth responses to drought at short-and long-term scales in Spain: squeezing the stress memory from tree rings. Front Ecol Evol 6:9
Coates KD, Canham CD, Beaudet M, Sachs DL, Messier C (2003) Use of a spatially explicit individual-tree model (SORTIE/BC) to explore the implications of patchiness in structurally complex forests. Forest Ecol Manage 186:297–310
Coomes DA, Allen RB (2007) Effects of size, competition and altitude on tree growth. J Ecol 95(5):1084–1097
Enquist BJ, Brown JH, West GB (1998) Allometric scaling of plant energetics and population density. Nature 395(6698):163–165
Enquist BJ, West GB, Brown JH (2009) Extensions and evaluations of a general quantitative theory of forest structure and dynamics. Proc Nat Acad Sci USA 106:7046–7051
Franz F, Bachler J, Deckelmann B, Kennel E, Kennel R, Schmidt A, Wotschikowsky U (1973) Bayerische Waldinventur 1970/71, Inventurabschnitt I: Großrauminventur Aufnahme- und Auswertungsverfahren. Forstl Forschungsber München 11: 143.
Grote R, Pretzsch H (2002) A model for individual tree development based on physiological processes. Plant Biology 4(02):167–180
Johann K (1993) DESER-Norm 1993. Normen der Sektion Ertragskunde im Deutschen Verband Forstlicher Forschungsanstalten zur Aufbereitung von waldwachstumskundlichen Dauerversuchen. Proc Dt Verb Forstl Forschungsanst, Sek Ertragskd, in Unterreichenbach-Kapfenhardt. 96–104
Jonard M, André F, de Coligny F, de Wergifosse L, Beudez N, Davi H, Ligot G, Ponette Q, Vincke C (2020) HETEROFOR 1.0: a spatially explicit model for exploring the response of structurally complex forests to uncertain future conditions–Part 1: carbon fluxes and tree dimensional growth. Geosci Model Dev 13:905–935
Jüttner O (1955) Eichenertragstafeln. In: Schober R (ed) (1971) Ertragstafeln der wichtigsten Baumarten. JD Sauerländer’s Verlag, Frankfurt am Main, pp 12–25, 134–138
King DA (2005) Linking tree form, allocation and growth with an allometrically explicit model. Ecol Model 185:77–91
Kleiber M (1947) Body size and metabolic rate. Physiol Rev 27(4):511–541
Korol RL, Running SW, Milner KS (1995) Incorporating intertree competition into an ecosystem model. Can J For Res 25:413–424
Kozłowski J, Konarzewski M (2004) Is West, Brown and Enquist’s model of allometric scaling mathematically correct and biologically relevant? Funct Ecol 18(2):283–289
Kramer H, Akça A (1995) Leitfaden zur Waldmeßlehre. JD Sauerländer’s Verlag, Frankfurt am Main, p 266
Lacointe A (2000) Carbon allocation among tree organs: a review of basic processes and representation in functional-structural tree models. Ann For Sci 57:521–533
Landsberg J (2003) Physiology in forest models: history and the future. Forest Biom Model Inform Sci 1:49–63
Landsberg JJ, Sands PJ (2011) Physiological ecology of forest production: principles, processes and models, vol 4. Elsevier/Academic Press, The Netherlands, p 352
Lüttge U, Hertel B (2009) Diurnal and annual rhythms in trees. Trees 23(4):683
Mäkelä A, Valentine HT (2006) Crown ratio influences allometric scaling in trees. Ecology 87(12):2967–2972
Mäkelä A, Landsberg J, Ek AR, Burk TE, Ter-Mikaelian M, Ågren GI, Oliver CD, Puttonen P (2000) Process-based models for forest ecosystem management: current state of the art and challenges for practical implementation. Tree Physiol 20:289–298
Muller-Landau HC, Condit RS, Chave J, Thomas SC, Bohlman SA, Bunyavejchewin S, Harms KE (2006) Testing metabolic ecology theory for allometric scaling of tree size, growth and mortality in tropical forests. Ecol Lett 9(5):575–588
Netzer Y, Munitz S, Shtein I, Schwartz A (2019) Structural memory in grapevines: early season water availability affects late season drought stress severity. Eur J Agron 105:96–103
Nord-Larsen T, Johannsen V (2007) A state-space approach to stand growth modelling of European beech. Ann For Sci 64:365–374
Ogle K, Barber JJ, Barron-Gafford GA, Patrick Bentley L, Young JM, Huxman TE, Loik ME, Tissue DT (2015) Quantifying ecological memory in plant and ecosystem processes. Ecol Lett 18(3):221–235
Pinheiro J et al. "R Core Team. (2018) nlme: linear and nonlinear mixed effects models. R package version 3.1-137." R Found. Stat. Comput. Retrieved from https://CRAN.R-project.org/package=nlme. Accessed 19 July 2018
Pretzsch H (2002) Grundlagen der Waldwachstumsforschung. Blackwell Wissenschafts, Berlin, p 414
Pretzsch H (2014) Canopy space filling and tree crown morphology in mixed-species stands compared with monocultures. For Ecol Manage 327:251–264
Pretzsch H (2017) Langfristige Ertragskundliche Versuchsflächen in Wäldern-Idee, Nutzen und Perspektiven. BFW Berichte 153:5–23
Pretzsch H (2019) The effect of tree crown allometry on community dynamics in mixed-species stands versus monocultures. A review and perspectives for modeling and silvicultural regulation. Forests 10(9):810
Pretzsch H (2021) The social drift of trees. Consequence for growth trend detection, stand dynamics, and silviculture. Eur J Forest Res. https://doi.org/10.1007/s10342-020-01351-y ((in press))
Pretzsch H, Rais A (2016) Wood quality in complex forests versus even-aged monocultures: review and perspectives. Wood Sci Technol 50(4):845–880
Pretzsch H, Forrester DI, Rötzer T (2015) Representation of species mixing in forest growth models. A review and perspective. Ecol Model 313:276–292
Pretzsch H, del Río M, Biber P, Arcangeli C, Bielak K, Brang P, Sycheva E (2019) Maintenance of long-term experiments for unique insights into forest growth dynamics and trends: review and perspectives. Eur J Forest Res 138(1):165–185
Richards FJ (1959) A flexible growth function for empirical use. J Exp Bot 10(2):290–301
Schober R (1967) Buchen-Ertragstafel für mäßige und starke Durchforstung. In: Schober R (1972) Die Rotbuche 1971 Schr Forstl Fak Univ Göttingen u Niedersächs Forstl Versuchsanst 43/44, JD Sauerländer’s Verlag, Frankfurt am Main, p 333
Schober R (1975) Ertragstafeln wichtiger Baumarten. JD Sauerländer’s Verlag, Frankfurt am Main
Schütz JP (2002) Silvicultural tools to develop irregular and diverse forest structures. Forestry 75(4):329–337
Sievänen R, Nikinmaa E, Nygren P, Ozier-Lafontaine H, Perttunen J, Hakula H (2000) Components of functional-structural tree models. Ann For Sci 57(5):399–412
Sievänen R, Perttunen J, Nikinmaa E, Kaitaniemi P (2008) Toward extension of a single tree functional-structural model of Scots pine to stand level: effect of the canopy of randomly distributed, identical trees on development of tree structure. Funct Plant Biol 35:964–975
Sillett SC, Van Pelt R, Carroll AL, Kramer RD, Ambrose AR, Trask DA (2015) How do tree structure and old age affect growth potential of California redwoods? Ecol Monogr 85(2):181–212
Skovsgaard JP, Vanclay JK (2008) Forest site productivity: a review of the evolution of dendrometric concepts for even-aged stands. Forestry 81(1):13–31
Stephenson NL, Das AJ, Condit R, Russo SE, Baker PJ, Beckman NG, Alvarez E (2014) Rate of tree carbon accumulation increases continuously with tree size. Nature 507(7490):90–93
Valentine HT, Mäkelä A (2005) Bridging process-based and empirical approaches to modeling tree growth. Tree Physiol 25(7):769–779
Verein Deutscher Forstlicher Versuchsanstalten (1873) Anleitung für Durchforstungsversuche. In: Ganghofer von A eds (1884) Das Forstliche Versuchswesen. Schmid‘sche Buchhandlung, Augsburg, vol 2, pp 247-253
Verein Deutscher Forstlicher Versuchsanstalten (1902) Beratungen der vom Vereine Deutscher Forstlicher Versuchsanstalten eingesetzten Kommission zur Feststellung des neuen Arbeitsplanes für Durchforstungs- und Lichtungsversuche. AFJZ 78:180–184
Bertalanffy von L (1951) Theoretische Biologie: II. Band, Stoffwechsel, Wachstum, 2nd edn. A Francke AG, Bern, p 418
Ganghofer von A (1881) Das Forstliche Versuchswesen, Band I. Augsburg, 1881, 505S.
Webster CR, Lorimer CG (2003) Comparative growing space efficiency of four tree species in mixed conifer–hardwood forests. For Ecol Manage 177(1–3):361–377
West GB, Brown JH, Enquist BJ (1997) A general model for the origin of allometric scaling laws in biology. Science 276(5309):122–126
West GB, Enquist BJ, Brown JH (2009) A general quantitative theory of forest structure and dynamics. Proc Nat Acad Sci USA 106:7040–7045
Wiedemann E (1943) Kiefern-Ertragstafel für mäßige Durchforstung, starke Durchforstung und Lichtung, In: Wiedemann E (1948) Die Kiefer 1948. Verlag M and H Schaper, Hannover, 337 p
Zeide B (1993) Analysis of growth equations. For Sci 39(3):594–616
Acknowledgements
This publication is part of the CARE4C project that has received funding from the European Union’s HORIZON 2020 research and innovation programme under the Marie Skłodowska-Curie grant agreement No 778322. I thank Gerhard Schütze for preparing the data of the long-term experiments, Monika Bradatsch for contributing graphical artwork, and anonymous reviewers for their constructive criticism.
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The author declares that he has no conflict of interest.
Additional information
Communicated by van der Maaten.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Pretzsch, H. Tree growth as affected by stem and crown structure. Trees 35, 947–960 (2021). https://doi.org/10.1007/s00468-021-02092-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00468-021-02092-0