
Abstract
We recently identified oncologic miR-182 as a new regulator of pulmonary artery hypertension (PAH) that targets myeloid-associated differentiation marker (Myadm), which is expressed in bone marrow stem cells and multipotent progenitors. Both miR-182 and Myadm are expressed in the cardiopulmonary system and correlated with the balance between the bone morphogenetic protein (BMP) and the transforming growth factor (TGF)-β signalling pathways, which are disturbed in PAH. We hypothesize that miR-182/Myadm are involved in BMP-TGF-β-signalling way in PAH. Hypoxia triggered pathological progression in cardiopulmonary PAH in vivo and in vitro; these changes were accompanied by strongly dowregulated BMP/SMAD1/5/8 expression and enhanced TGF-β/SMAD2/3 signalling pathway, favouring SMAD4/SMAD2 transcript formation and inhibiting the PAH negative regulator Id1 expression. miR-182 gain-of-function significantly inhibited the pathological progression in hypoxia-induced PAH (HPH) in vivo and in vitro, with a restoration of the balance in BMP-TGF-β signalling pathway. This recovery was abrogated by overexpression of Myadm. Conversely, loss-of-function of miR-182 increased the pathological progression of HPH followed by severe disturbance of BMP and TGF-β signal transduction and reduced Id1 expression, which was restored by Myadm knockdown. We also showed that the miR-182/Myadm relate BMP-TGF-β pathway is associated with NOS3/NO/cGMP via the crosstalk between endothelial cells and smooth muscle cells. Our findings further support the therapeutic significance of miR-182/Myadm in PAH via the balance of BMP- and TGF-β-associated mechanisms.










Similar content being viewed by others

References
Aranda JF, Reglero-Real N, Kremer L, Marcos-Ramiro B, Ruiz-Sáenz A, Calvo M, Enrich C, Correas I, Millán J, Alonso MA (2011) MYADM regulates Rac1 targeting to ordered membranes required for cell spreading and migration. Mol Biol Cell 22:1252–1262. https://doi.org/10.1091/mbc.E10-11-0910
Bai Y, Sun L, Du L, Zhang T, Xin W, Lan X, Du G (2013) Association of circulating levels of asymmetric dimethylarginine (ADMA) with carotid intima-media thickness: evidence from 6168 participants. Ageing Res Rev 12:699–707. https://doi.org/10.1016/j.arr.2012.02.003
Calvier L, Boucher P, Herz J, Hansmann G (2019) LRP1 deficiency in vascular SMC leads to pulmonary arterial hypertension that is reversed by PPARγ activation. Circ Res 124:1778–1785. https://doi.org/10.1161/CIRCRESAHA.119.315088
Chen J, Deng Y, Ao L, Song Y, Xu Y, Wang CC, Choy KW, Tony Chung KH, Du Q, Sui Y, Yang T, Yang J, Li H, Zou C, Tang T (2019) The high-risk HPV oncogene E7 upregulates miR-182 expression through the TGF-β/Smad pathway in cervical cancer. Cancer Lett 460:75–85. https://doi.org/10.1016/j.canlet.2019.06.015
Chen M, Shen H, Zhu L, Yang H, Ye P, Liu P, Gu Y, Chen S (2019) Berberine attenuates hypoxia-induced pulmonary arterial hypertension via bone morphogenetic protein and transforming growth factor-β signaling. J Cell Physiol 234:17482–17493. https://doi.org/10.1002/jcp.28370
Ding C, Ding X, Zheng J, Wang B, Li Y, Xiang H, Dou M, Qiao Y, Tian P, Xue W (2020) miR-182-5p and miR-378a-3p regulate ferroptosis in I/R-induced renal injury. Cell Death Dis 11:929. https://doi.org/10.1038/s41419-020-03135-z
Gao Y, Pan L, Zhao L, Dang X (2021) HDAC1 promotes artery injury through activation of VAV3 by binding to miR-182-5p in atherosclerotic mice model. Cell Signal 78:109840. https://doi.org/10.1016/j.cellsig.2020.109840
Heo J, Yang HC, Rhee WJ, Kang H (2020) Vascular smooth muscle cell-derived exosomal microRNAs regulate endothelial cell migration under PDGF stimulation. Cells 9:639. https://doi.org/10.3390/cells9030639
Kumar R, Mickael C, Kassa B, Gebreab L, Robinson JC, Koyanagi DE, Sanders L, Barthel L, Meadows C, Fox D, Irwin D, Li M, McKeon BA, Riddle S, Dale Brown R, Morgan LE, Evans CM, Hernandez-Saavedra D, Bandeira A, Maloney JP, Bull TM, Janssen WJ, Stenmark KR, Tuder RM, Graham BB (2017) TGF-β activation by bone marrow-derived thrombospondin-1 causes Schistosoma- and hypoxia-induced pulmonary hypertension. Nat Commun 8:15494. https://doi.org/10.1038/ncomms15494
Lavoie JR, Ormiston ML, Perez-Iratxeta C, Courtman DW, Jiang B, Ferrer E, Caruso P, Southwood M, Foster WS, Morrell NW, Stewart DJ (2014) Proteomic analysis implicates translationally controlled tumor protein as a novel mediator of occlusive vascular remodeling in pulmonary arterial hypertension. Circulation 129:2125–2135
Lee MY, Tsai KB, Hsu JH, Shin SJ, Wu JR, Yeh JL (2016) Liraglutide prevents and reverses monocrotaline-induced pulmonary arterial hypertension by suppressing ET-1 and enhancing eNOS/sGC/PKG pathways. Sci Rep 6:31788. https://doi.org/10.1038/srep31788
Li N, Hwangbo C, Jaba IM, Zhang J, Papangeli I, Han J, Mikush N, Larrivée B, Eichmann A, Chun HJ, Young LH, Tirziu D (2016) miR-182 modulates myocardial hypertrophic response induced by angiogenesis in heart. Sci Rep 6:21228. https://doi.org/10.1038/srep21228
Li Q, Xing W, Gong X, Wang Y (2018) Long non-coding RNA urothelial carcinoma associated 1 promotes proliferation, migration and invasion of osteosarcoma cells by regulating microRNA-182. Cell Physiol Biochem 51:1149–1163. https://doi.org/10.1159/00049549
Oliveira SDS, Chen J, Castellon M, Mao M, Raj JU, Comhair S, Erzurum S, Silva CLM, Machado RF, Bonini MG, Minshall RD (2019) Injury-induced shedding of extracellular vesicles depletes endothelial cells of Cav-1 (Caveolin-1) and enables TGF-β (transforming growth factor-β)-dependent pulmonary arterial hypertension. Arterioscler Thromb Vasc Biol 39:1191–1202. https://doi.org/10.1161/ATVBAHA.118.312038
Pettersson M, Dannaeus K, Nilsson K, Jönsson JI (2000) Isolation of MYADM, a novel hematopoietic-associated marker gene expressed in multipotent progenitor cells and up-regulated during myeloid differentiation. J Leukoc Biol 67:423–431. https://doi.org/10.1002/jlb.67.3.423
Pekarova M, Koudelka A, Kolarova H, Ambrozova G, Klinke A, Cerna A, Kadlec J, Trundova M, Sindlerova Svihalkova L, Kuchta R, Kuchtova Z, Lojek A, Kubala L (2015) Asymmetric dimethyl arginine induces pulmonary vascular dysfunction via activation of signal transducer and activator of transcription 3 and stabilization of hypoxia-inducible factor 1-alpha. Vascul Pharmacol 73:138–148. https://doi.org/10.1016/j.vph.2015.06.005
Qiu Y, Luo X, Kan T, Zhang Y, Yu W, Wei Y, Shen N, Yi B, Jiang X (2014) TGF-β upregulates miR-182 expression to promote gallbladder cancer metastasis by targeting CADM1. Mol Biosyst 10:679–685. https://doi.org/10.1039/c3mb70479c
Song L, Liu L, Wu Z, Li Y, Ying Z, Lin C, Wu J, Hu B, Cheng SY, Li M, Li J (2012) TGF-beta induces miR-182 to sustain NF-kappaB activation in glioma subsets. J Clin Invest 122:3563–3578. https://doi.org/10.1172/JCI62339
Sun L, Bai Y, Zhao R, Sun T, Cao R, Wang F, He G, Zhang W, Chen Y, Ye P, Du G (2016) Oncological miR-182-3p, a novel smooth muscle cell phenotype modulator, evidences from model rats and patients. Arterioscler Thromb Vasc Biol 36:1386–1397. https://doi.org/10.1161/ATVBAHA.115.307412
Sun L, Hu C, Wang Z, Zhang X (2020) MiR-182 inhibits kidney fibrosis by regulating transforming growth factor β1/Smad3 pathway in autosomal dominant polycystic kidney disease. IUBMB Life 72:1340–1348. https://doi.org/10.1002/iub.2255
Sun L, Lin P, Chen Y, Yu H, Ren S, Wang J, Zhao L, Du G (2020) miR-182-3p/Myadm contribute to pulmonary artery hypertension vascular remodeling via a KLF4/p21-dependent mechanism. Theranostics 10:5581–5599. https://doi.org/10.7150/thno.44687
Tao J, Barnett JV, Watanabe M, Ramírez-Bergeron D (2018) Hypoxia supports epicardial cell differentiation in vascular smooth muscle cells through the activation of the TGFβ pathway. J Cardiovasc Dev Dis 5:19. https://doi.org/10.3390/jcdd5020019
Upton PD, Davies RJ, Tajsic T, Morrell NW (2013) Transforming growth factor-beta(1) represses bone morphogenetic protein-mediated Smad signaling in pulmonary artery smooth muscle cells via Smad3. Am J Respir Cell Mol Biol 49:1135–1145. https://doi.org/10.1165/rcmb.2012-0470OC
Yang J, Li X, Al-Lamki RS, Southwood M, Zhao J, Lever AM, Grimminger F, Schermuly RT, Morrell NW (2010) Smad-dependent and Smad-independent induction of Id1 by prostacyclin analogues inhibits proliferation of pulmonary artery smooth muscle cells in vitro and in vivo. Circ Res 107:252–262. https://doi.org/10.1161/CIRCRESAHA.109.209940
Yu J, Lei R, Zhuang X, Li X, Li G, Lev S, Segura MF, Zhang X, Hu G (2016) MicroRNA-182 targets SMAD7 to potentiate TGFβ-induced epithelial-mesenchymal transition and metastasis of cancer cells. Nat Commun 7:13884. https://doi.org/10.1038/ncomms13884
Zhang X, Xiao C, Liu H (2018) Ganoderic acid A protects rat H9c2 cardiomyocytes from hypoxia-induced injury via up-regulating miR-182-5p. Cell Physiol Biochem 50:2086–2096. https://doi.org/10.1159/000495053
Zhang T, Kawaguchi N, Tsuji K, Hayama E, Furutani Y, Sugiyama H, Nakanishi T (2020) Silibinin upregulates CXCR4 expression in cultured bone marrow cells (BMCs) especially in pulmonary arterial hypertension rat model. Cells 9:1276. https://doi.org/10.3390/cells9051276
Zeller T, Schurmann C, Schramm K, Müller C, Kwon S, Wild PS, Teumer A, Herrington D, Schillert A, Iacoviello L, Kratzer A, Jagodzinski A, Karakas M, Ding J, Neumann JT, Kuulasmaa K, Gieger C, Kacprowski T, Schnabel RB, Roden M, Wahl S, Rotter JI, Ojeda F, Carstensen-Kirberg M, Tregouet DA, Dörr M, Meitinger T, Lackner KJ, Wolf P, Felix SB, Landmesser U, Costanzo S, Ziegler A, Liu Y, Völker U, Palmas W, Prokisch H, Guo X, Herder C, Blankenberg S, Homuth G (2017) Transcriptome-wide analysis identifies novel associations with blood pressure. Hypertension 70:743–750. https://doi.org/10.1161/HYPERTENSIONAHA.117.09458
Funding
This work was supported by National Natural Science Foundation (Grant number: 82070056), the Open Project of the National Clinical Research Center for Geriatric Diseases (NCRCG-PLAGH-2019025), and the grant from the National Key Technologies R&D Program for New Drugs of China (no. 2018ZX09J18109-004).
Author information
Authors and Affiliations
Contributions
LS, YB, KH and HX designed the study and produced the initial draft of the manuscript. LS, YC, JW and TL contributed to the haemodynamic measurements. YC and X produced animal models of PAH. LS, YB and CL carried all in vitro experiments and collected the data. YB carried out data analyses. LS, YB, and KH contributed to funding acquisition. LS, and YB contributed to response of the manuscript. All authors have read and approved the final submitted manuscript.
Corresponding authors
Ethics declarations
Conflict of interest
The author declares that they have no competing interest.
Ethics approval
Not applicable.
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Availability of data and materials
Supplementary data to this article can be found online.
Supplementary Information
Below is the link to the electronic supplementary material.
395_2021_892_MOESM1_ESM.tif
Fig. SI1 The study design of Hypoxia-induced pulmonary hypertension in rats. a. A preliminary experiment was established by placing the rats in hypobaric hypoxic chambers (0.5 atm) with simulated altitude of 18,000 ft (10% O2) to mediate a hypoxia-mediated pulmonary hypertension. At the beginning, 32 rats were placing in the chambers. After hypoxia exposure (about 0 days, 7 days, 14 days and 21 days), rats were weighed and anaesthetized for the right ventricular systolic pressure (RVSP) and pulmonary artery pressure (PAP) examination by right-sided heart catheterization to check whether the HPH model is successfully established or not. Two rats died because of intolerance of hypoxia on Day 21. b. The schematic diagram of miR-182/Myadm treatment in rats in pulmonary artery remodelling in hypobaric hypoxic chambers. On D0, Rats treated with either recombinant adenovirus expressing Myadm, or shRNA against Myadm or control virus at a dose of 2×1010 pfu/kg followed by tail vein injection with the miR-182 mimic-overexpressing adenovirus or miR-182 inhibitor-overexpressing adenovirus control at a dose of 2×1010 pfu/kg were then placed in the hypobaric hypoxic chambers to be exposed to hypoxia (10% PO2) for 3 weeks. The chamber was flushed with a mixture of room air and nitrogen and was opened on the alternate day during health check and feeding. Normoxic control rats (normoxia) were kept at sea level altitude under the same light–dark cycle received injections with an equal volume of DMSO vehicle. On Day12, rats were re-treated with the recombinant adenovirus as indicated in the panel c. c. Grouping (treated with virus as indicated), exposure to normoxia (or hypoxia) and numbers (N) of rats in each group at the beginning of experiment and the numbers of rats died from intolerance of hypoxia or weakness or anaesthetization. (TIF 12097 KB)
395_2021_892_MOESM2_ESM.tif
Fig. S2 miR-182 and Myadm regulate migration in response to hypoxia and TGF-β in PA-SMCs. a-b: Quiescent rat PA-SMCs were transfected with recombinant adenovirus expressing the Myadm gene (Ad-Myadm) or control vector and then transfected with the recombinant adenovirus of miR-182 mimic (miR-182) at a concentration of 30 MOI per cell. c-d: Quiescent rat PA-SMCs were transfected with recombinant adenovirus expressing the specific shRNA of Myadm gene (shRNA) or control virus and then transfected with recombinant adenovirus containing miR-182 inhibitor (miR-182 inhibitor) at a concentration of 30 MOI per cell. MOI: multiplicity of infection. Then, the cells were stimulated with TGF-β (5 ng/mL) or exposed to hypoxia (0.1% PO2) for the next 24 h for cell migration detection using a wound-healing assay. The images are shown at 10× magnification. Bar indicates 100 μm. Four different fields of migration were photographed with a video camera system, and migration was analysed using Image-Pro Plus 5.1 software at the intersection of the previously marked line and the wound edge before and after exposure to hypoxia or treatment with TGF-β for 24 h. Migration was expressed as the change in covered area. The results are expressed as the mean ±SD. n=6 *p<0.05, **p<0.01 vs the PA-SMCs infected with control virus and exposed to normoxia or treated with solvent control. #p<0.05, ##<0.01 vs the PA-SMCs infected with control virus in response to TGF-β or hypoxia exposure. $ p<0.05, $$<0.01 vs the PA-SMCs infected with miR-182/miR-182 inhibitor-overexpressing virus in response to TGF-β or hypoxia exposure. n=6 per group. (TIF 5258 KB)
395_2021_892_MOESM3_ESM.tif
Fig. S3 miR-182/Myadm regulates PASMC proliferation in response to hypoxia. Quiescent rat PA-SMCs were transfected with different recombinant adenoviruses as indicated. Then, cells were exposed to hypoxic conditions (0.1% PO2) for the next 48 h to detect cell proliferation using an EdU incorporation assay. Red fluorescence indicated positive cells (DNA incorporation), and DAPI was employed to detect the nuclei. The images are shown at 20×magnification. The results are expressed as the mean ± SD. *p<0.05, **p<0.01 vs the PA-SMCs infected with control virus. #p<0.05, ##<0.01 vs the PA-SMCs infected with control virus and subjected to hypoxia. $ p<0.05, $$<0.01 vs the PA-SMCs infected with miR-182/miR-182 inhibitor-overexpressing virus and subjected to hypoxia. n=6 per group. (TIF 5267 KB)
395_2021_892_MOESM4_ESM.tif
Fig. SI4 miR-182/Myadm regulates proliferation in response to TGF-β. Quiescent rat PA-SMCs were transfected with different recombinant adenoviruses at a concentration of 30 MOI per cell as indicated. Then, the cells were stimulated with TGF-β (5 ng/mL). The EdU incorporation assay was used to detect proliferating cells. Red fluorescence indicated positive cells (DNA incorporation), and DAPI was employed to detect the nuclei. The images are shown at 20×magnification. The results are expressed as the mean ± SD. *p<0.05, **p<0.01 vs the PA-SMCs infected with control virus and treated with control solvent. #p<0.05, ##<0.01 vs the PA-SMCs treated with miR-182/miR-182 inhibitor-overexpressing virus and treated with TGFβ. $ p<0.05, $$<0.01 vs the PA-SMCs infected with miR-182/miR-182 inhibitor-overexpressing virus and treated with TGFβ. n=6 per group (TIF 4881 KB)
395_2021_892_MOESM5_ESM.tif
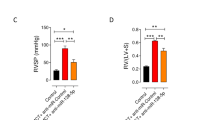
Fig. SI5 Overexpression of miR-182 regulates right ventricular remodelling and hypertrophy in rats with HPH by targeting Myadm. Rats treated with recombinant adenovirus as indicated and then were exposed to hypoxia (10% PO2) or normoxia. Whole heart samples were embedded in paraffin, sliced into 5-μm sections and stained with HE (a); the bar indicates 5 mm. Sirius red (d) and wheat germ agglutinin (e) staining followed by examination with a light microscope (Nikon) were used to detect disrupted collagen deposition and myocardial fibrosis. DAPI was employed to detect the nuclei (e). The ratio of heart weight to body weight (b) and right ventricular hypertrophy (c) (shown as the weight ratio of right ventricle to left ventricle plus interventricular septal, R/(L+S)) were detected. The results are expressed as the mean ±SD. *p<0.05, **p<0.01 vs the rats injected with control virus. #p<0.05, ##<0.01 vs the rats treated with miR-182-overexpressing virus and exposed to normoxia. $ p<0.05, $$<0.01 vs the rats treated with miR-182-overexpressing virus and subjected to hypoxia. n=6-8 per group as indicated in Fig. SI1 (TIF 6277 KB)
395_2021_892_MOESM6_ESM.tif
Fig. SI6 Knockdown of miR-182 regulates right ventricular remodelling and hypertrophy in rats with HPH via overexpression of Myadm. Rats treated with either recombinant adenovirus as indicated were exposed to hypoxia (10% PO2) or normoxia. Whole heart samples were embedded in paraffin, sliced into 5-μm sections and stained with HE (a), bar indicates 5 mm. Sirius red (d) and wheat germ agglutinin (e) staining followed by examination with a light microscope (Nikon) were used to detect disrupted collagen deposition and myocardial fibrosis. The ratio of heart weight to body weight (b) and right ventricular hypertrophy (c) (shown as the R/(L+S)) were detected. The results are expressed as the mean ±SD. *p<0.05, **p<0.01 vs the rats injected with control virus. #p<0.05, ##<0.01 vs the rats treated with miR-182 inhibitor-overexpressing virus and exposed to normoxia. $ p<0.05, $$<0.01 vs the rats treated with miR-182 inhibitor-overexpressing virus and then subjected to be exposure to hypoxia. n=6-8 per group as indicated in Fig. SI1 (TIF 12257 KB)
395_2021_892_MOESM7_ESM.tif
Fig. SI7 miR-182 regulates the expression of critical factors in the BMP and TGF-β signalling pathways in PA-SMCs in a Myadm-dependent manner in response to hypoxia. Quiescent rat PA-SMCs were transfected with different recombinant adenoviruses at a concentration of 30 MOI per cell as indicated. Then, the cells were stimulated with TGF-β (5 ng/mL) or exposed to hypoxic conditions (0.1% PO2). 24 hours later, total RNA from PASMCs was extracted, and the relative mRNA expression levels of miR-182, Myadm (a), BMP2 (b, left panels), BMP4 (c, left panels), Id1 (d) and p21/Cip1 (e) to those from the control PASMCs (X-fold) were detected. β-Actin was used as a loading control. The supernatant of PASMCs was collected, and the relative expression levels of BMP2/4 were measured using ELISA kits (b, c right panels). The results are expressed as the mean ± SD. *p<0.05, **p<0.01 vs the PA-SMCs infected with control virus and exposure to normoxia condition. #p<0.05, ##<0.01 vs the PA-SMCs treated with miR-182/miR-182 inhibitor-overexpressing virus and subjected to be exposure to hypoxia. $ p<0.05, $$<0.01 vs the PA-SMCs infected with miR-182/miR-182 inhibitor-overexpressing virus and then subjected to be exposure to hypoxia. n=6 per group. (TIF 3909 KB)
395_2021_892_MOESM8_ESM.tif
Fig. SI8 miR-182 regulates the expression of critical factors in the BMP-TGF-β signalling transduction pathway in a Myadm-dependent manner in response to TGFβ in PA-SMCs. Quiescent rat PA-SMCs were transfected with different recombinant adenoviruses as indicated. Then, the cells were stimulated with TGF-β (5 ng/mL) or solvent control. The relative mRNA expression levels of BMP2 (a, left panels), BMP4 (b, left panels), Id1 (c) and p21/Cip1 (d) compared to those from the control PA-SMCs (X-fold) were detected. β-Actin was used as loading control. The supernatant of PA-SMCs was collected, and the relative expression levels of BMP2/4 were measured using ELISA kits (a, b right panels). The results are expressed as the mean ± SD. *p<0.05, **p<0.01 vs the PA-SMCs infected with control virus and treated with control solvent. #p<0.05, ##<0.01 vs the PA-SMCs treated with miR-182/miR-182 inhibitor-overexpressing virus and treated with TGF-β. $ p<0.05, $$<0.01 vs PA-SMCs infected with miR-182/miR-182 inhibitor-overexpressing virus and treated with TGF-β. n=6 per group. (TIF 3582 KB)
395_2021_892_MOESM9_ESM.tif
Fig. SI9 The change of MiR-182/Myadm in PA-ECs regulates the SMAD1/5/8-SMAD2/3 signalling transduction way in PA-SMCs. a. The schematic diagram of a coculture of human PA-EC and PA-SMC that was performed using a Transwell cell culture insert chamber as indicated. PA-ECs: pulmonary artery endothelial cells, PA-SMCs: pulmonary artery smooth muscle cells. Transwell chamber contains a membrane with 0.4 µm pores which prevented cells from direct contact. Briefly, the PA-SMCs (1×105 cells/well) were added to in the bottom of the 24-well plates. The Transwell system were inserted as indicated in panel a. the PA-ECs infected with the recombinant adenovirus vectors as indicated for 48 hours and then were seeded in the upper compartment of the Transwell inserts at a concentration of 4×104 cells/well. Hence, the coculture system contains the PA-SMCs in the bottom compartment and the PA-ECs in the upper compartment. Then, the coculture systems were exposed to hypoxia (0.1% PO2) for the next 24 h, the cell supernatant were collected for cGMP detection using ELISA kit (b) and the PA-SMC lysates were collected for the measurement of the phosphorous levels of SMAD1/5/8 and SMAD2/3 (c,d) using Western Blotting. The results are expressed as the mean ± SD. *p<0.05, **p<0.01 vs the PA-SMCs cocultured with PA-ECs infected with control virus. #p<0.05, ##<0.01 vs the PA-SMCs cocultured with PA-ECs infected with control virus and subjected to hypoxia. $ p<0.05, $$<0.01 vs the PA-SMCs cocultured with PA-ECs infected with miR-182/miR-182 inhibitor-overexpressing virus and subjected to hypoxia. n=6 per group. (TIF 5856 KB)
395_2021_892_MOESM10_ESM.tif
Fig. SI10. Western blotting images of protein expression. (a). The blotting images of Myadm, BMPR2, HIF-1α, β-Actin, GAPDH, phosphorous-SMAD2/3 (p-SMAD2/3), phosphorous-SMAD1/5/8 SMAD2/3 and SMAD1/5/8 from the lungs in HPH rats as were indicated in Fig. 6g and 7g. (b). The blotting images of Myadm, HIF1α, phosphorous-NOS3, NOS3 and GAPDH in PA-ECs in the coculture of we indicated in Fig.9d and e. (c). The blotting images of phosphorous-SMAD2/3 (p-SMAD2/3), phosphorous-SMAD1/5/8 SMAD2/3 and SMAD1/5/8, GAPDH and β-Actin in PA-SMCs as we indicated in Fig.SI10 c,d. (TIF 1511 KB)
Rights and permissions
About this article

Cite this article
Bai, Y., Wang, J., Chen, Y. et al. The miR-182/Myadm axis regulates hypoxia-induced pulmonary hypertension by balancing the BMP- and TGF-β-signalling pathways in an SMC/EC-crosstalk-associated manner. Basic Res Cardiol 116, 53 (2021). https://doi.org/10.1007/s00395-021-00892-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00395-021-00892-6