
Abstract
Sesame (Sesamum indicum L.) is an important oilseed crop with significant economic importance in many developing countries. This study rigorously investigated the impact of various applications and doses of two specific plant growth regulators, paclobutrazol (PAC) and mepiquat chloride (MC), on the development, yield components, oil content, and fatty acid composition of sesame plants across two consecutive growing seasons. The research revealed that while paclobutrazol application increased the plant height it also resulted in a slight decrease in oil content. The paclobutrazol treatment resulted in the highest seed yield (21.3 g/plant), which was significantly different from both the MC (13.1 g/plant) and PAC + MC (14.6 g/plant) treatments. In contrast, the mepiquat chloride treatment resulted in the lowest oil content. The combined application of both regulators showed intermediate effects. In addition, variations in fatty acid content were observed between applications and doses, with significant differences in saturated and unsaturated fatty acid levels. The findings provide valuable insights into the potential benefits and implications of using growth regulators in sesame cultivation, especially in the context of mechanized harvesting and oil quality optimization.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Sesame (Sesamum indicum L.), belonging to the Pedaliaceae family, is an ancient oilseed crop. Due to its extreme oil and nutritional value, sesame is often referred to as the “queen of oilseed crops.” The oil content in sesame can vary based on the cultivar, with some reaching as high as 62.7% (Uzun et al. 2008). This high oil content makes sesame as significant as other oil crops like oilseed rape (Sharafi et al. 2015), soybean (Yao et al. 2020), and peanut (Yol et al. 2017). Sesame oil is known for its resistance to rancidity, primarily due to its fatty acid composition and unique antioxidants. Notably, oleic and linoleic acids make up approximately 80% of the total fatty acids which increase health benefits (Were et al. 2006). The oil is also valued for its nutritional content, which includes proteins, vitamins, amino acids, and antioxidants such as sesamin, sesamolin, and sesamol (Ghafoorunissa and MVV 2004; C and SA 2015).
Despite an annual production of 6.35 million tonnes across 12.50 million hectares (FAO, 2021), sesame faces a significant gap between supply and demand due to consistently low productivity and yield. This is attributed to the crop’s wild characteristics (Ashri 1998; Uzun and Çağırgan 2006; Yol et al. 2021), lack of superior cultivars (Yol and Uzun 2012), and challenges from both abiotic and biotic stress factors (Ustun et al. 2017), including drought, waterlogging, salt, and heat stress exacerbated by climate change (Ciaffi et al. 1996; Raza et al. 2018). Seed shattering is another significant issue, reducing productivity and limiting sowing areas (Langham and Wiemers 2002; Qureshi et al. 2022), though losses can be mitigated through the selection of resistant cultivars or the use of innovative agronomic techniques, such as employing plant growth regulators (PGRs) or micronutrients (Shah et al. 2021).
Plant growth regulators (PGRs), including both promotive and inhibitory agents, can regulate plant height, suppress excessive vegetative growth, or enhance flowering, with the triazole compound being recognized for its growth regulatory properties and ability to enhance abiotic stress tolerance (Arteca 2014; Suman et al. 2017; Desta and Amare 2021). Paclobutrazol (PAC) and Mepiquat chloride (MC) are two PGRs widely used to modulate plant growth. PAC, a triazole family member, reduces plant height by inhibiting sterol and gibberellic activities (Khan et al. 2009), also serving as a stress protectant (Vineeth et al. 2016). MC, known for suppressing gibberellic acid, leads to reduced stem elongation (Tung et al. 2019). While their use has been documented in various plants, including sesame (Ahmed et al. 2023), rice (Syahputra et al. 2013), and sunflower (Koutroubas and Damalas 2015), the specific effects of PAC and MC on sesame’s growth and oil profile remain underexplored.
The efficacy of PGRs on plant development depends on the environmental conditions post-application. Favorable temperature and soil moisture levels can enhance positive plant responses (Koutroubas and Damalas 2020). Using the right concentration of growth regulators has shown promising results in increasing seed production and reducing shattering in sesame (Prakash and Ganesan 2000). However, overuse can decrease crop yield (Peng et al. 2014; Mehmood et al. 2021). While PGRs have been extensively studied in various crops (Tesfahun 2018), limited research exists on the effects of paclobutrazol and mepiquat chloride on sesame growth.
This study represented one of the first comprehensive analyses focusing on the impacts of PAC and MC on both the agronomic and oil quality traits of sesame, addressing a critical gap in current agricultural practices. Unlike previous studies that primarily focused on yield aspects, our research uniquely evaluated the combined effects of PAC and MC on sesame, particularly examining their role in modulating oil content and profile. By elucidating the roles of PAC and MC, our study not only filled a significant research gap but also offered practical insights that could lead to more resilient sesame cultivation practices amid changing climate conditions. Our findings revealed that strategic application of PAC and MC can significantly influence sesame’s growth dynamics and oil quality, offering novel insights for optimizing cultivation practices and improving crop resilience.
Materials and Methods
Plant Material
Turkey is known for its agricultural diversity, and sesame is an important crop, especially in regions with suitable climates for its cultivation. The country values sesame for its seeds, which are a key ingredient in traditional foods and oils. In this study, two sesame genotypes, Muganli-57 and Paloma, were utilized. Muganli-57, has yellow seeds, a high yield of about 50% oil, and its capsules shatter at harvest. It was highly demanded cultivar, registered by the West Mediterranean Agricultural Research Institute, Turkey. Paloma, sourced from the USDA GenBank with the accession number PI 599446, is characterized by curly leaves, brown seeds, and non-shattered capsules at harvest.
Climatic and Field Experiment Conditions
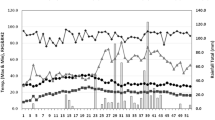
The climatic data from April to September, depicted in Fig. 1, elucidates the meteorological patterns of Antalya, highlighting the semi-arid climate of the experimental region during the study period. In 2021 and 2022, the peak temperatures for July and August were recorded at 31.3˚C and 30.6˚C, and 31.2˚C and 28.3˚C, respectively, demonstrating the region’s warm summer conditions. Precipitation levels from April to July 2021 were marginally lower than those in 2022, while humidity levels remained generally consistent across both years, with a notable exception in August. Complementing the climatic overview, a two-year field experiment was conducted at Akdeniz University’s experimental area located in Antalya, southern Turkey (coordinates: 36˚53′55.6“N 30˚38′29.5″E), during the 2021 and 2022 growing seasons. The site’s soil composition, a blend of loam and clay, is slightly alkaline, evidenced by a pH level of 7.7, and characterized by a low organic matter content, quantified at 1.23%.

Climatic condition of experimental site
Experimental Design
The study was conducted using a randomized complete block design (RCBD) with a factorial arrangement to systematically evaluate the effects of two plant growth regulators, paclobutrazol (PAC) and mepiquat chloride (MC), on the growth, yield, and oil composition of two sesame genotypes, Muganli 57 and Paloma. The experiment was carried out over two consecutive growing seasons (2021 and 2022) at Akdeniz University’s agricultural research fields in Antalya, Turkey. Before seeding, plant residues were cleared, and a drip irrigation system was set up. Standard recommended practices were consistently followed. Seeding occurred in the first week of June, with harvest in the last week of October for both years. The factorial design comprised two factors: (1) genotype, with two levels (Muganli 57 and Paloma), and (2) growth regulator treatment, with four levels for each regulator (control, low, medium, and high concentrations). Three treatments were administered: Paclobutrazol, mepiquat chloride, and a mixture of both growth regulators, each at varying doses. The treatments were:
-
Paclobutrazol:
-
A0-Control (no application)
-
A01 (200 mg/L)
-
A02 (400 mg/L)
-
A03 (600 mg/L)
-
Mepiquat chloride:
-
A0-Control (no application)
-
A01 (200 mg/L)
-
A02 (400 mg/L)
-
A03 (600 mg/L)
-
Paclobutrazol and Mepiquat chloride mixture:
-
B0-Control (no application)
-
B01 (100 mg/L PAC + 100 mg/L MC)
-
B02 (200 mg/L PAC + 200 mg/L MC)
-
B03 (300 mg/L P AC + 300 mg/L MC)
All chemicals, in liquid form, were mixed with water and applied twice: at the pre-reproductive stage (35 days post-planting) and the late-bloom stage (80 days post-sowing). The same dosages and applications were used in both years.
Data Collection
Each treatment combination was replicated three times across separate plots within each block to mitigate the effects of potential environmental variability within the field.
To address concerns regarding the sample size, particularly the representativeness of three plants per plot for traits such as yield and its components, we adopted the following methodology: Both cultivars were planted in rows 3 m long, spaced 70 cm apart, with individual plants 20 cm apart within rows, containing a total of 15 sesame plants arranged in rows to ensure adequate spacing and uniform exposure to environmental conditions. From each plot, three plants were randomly selected for detailed measurement of yield and yield components to ensure that the sampled plants accurately reflected the plot’s overall performance. Prior to the experiment, a power analysis was conducted to determine the adequacy of the sample size for detecting statistically significant differences among treatment combinations. This analysis confirmed that three plants per plot, given the number of replicates and treatment combinations, provided sufficient power to detect differences in yield and yield components with a confidence level of 95%. The yield was measured by harvesting the seeds from the selected plants at full maturity, followed by drying and weighing. Yield components, including the number of capsules per plant, seeds per capsule, and 100-seed weight, were also recorded. These measurements were complemented by observations of plant height and branching patterns to provide a comprehensive assessment of the growth regulators’ effects.
Oil Extraction, Methylation, and Gas Chromatography
The extraction process began with drying cleaned sesame seeds at 105˚C, followed by grinding using a Retsch GM 200 grinder. The oil was then extracted using the Soxhlet apparatus with petroleum ether as the solvent (boiling range 40–60˚C). Fatty acids were converted to methyl esters as described by Garces and Mancha (1993). These esters were analyzed using gas chromatography (GC) equipped with a flame ionization detector (FID) and a mass spectrometer (Shimadzu, GCMS-QP2020NX). The analysis used a capillary column (SH-Stabilwax capillary; dimensions: 60.0 m x 0.25 mm x 0.25 m). The GC-MS/FID analysis was conducted in split mode (40:1), with an injector temperature of 250˚C and a sample volume of 1 µL. Fatty acid ratios were determined by analyzing peak areas from the GC-FID, identified using the Wiley W9N11 library on the mass spectrometer.
Statistical Analysis
Analysis of variance (ANOVA) was conducted using the PROC GLM procedure in SAS Version 9.3 (Institute 2011). Mean comparisons for agronomic and quality traits were made using the least significant differences.
Results
Effect of Growth Regulators on Agronomic Traits
The effects of different treatments on various agronomic traits of sesame are presented in Table 1. The treatments included the application of paclobutrazol (PAC), mepiquat chloride (MC), and a combination of both (PAC + MC). Based on the results, the combined application of paclobutrazol and mepiquat chloride (PAC + MC) showed a slight increase in the number of capsules, and stem length of the first capsule, but did not significantly affect plant height. The highest number of branches were recorded in control compared to application of growth regulators. However, the PAC treatment significantly outperformed the other treatments in terms of seed yield, while the MC and control treatment had the highest 100 seed weight. The number of capsules formed was the highest in the PAC + MC treatment (111.3), followed closely by MC with 106.2 capsules and PAC with 105.5 capsules. The control line had the lowest value for this trait. Similarly, the number of branches was slightly higher in the PAC + MC treatment (6.1) compared to 5.7 in the MC treatment and 5.5 in the PAC treatment.
The stem length of the first capsule was the longest in the PAC + MC treatment (73.8 cm), followed by MC (70.1 cm) and PAC (70.0 cm). In terms of plant height, the PAC treatment resulted in the tallest plants (161.5 cm), while the control treatment had the shortest plants (151.4 cm). The MC treatment plants had a height of 160.1 cm. There were significant differences in seed yield among the treatments. The PAC treatment resulted in the highest seed yield (21.3 g/plant), which was significantly different from both the MC (13.1 g/plant) and P + MC (14.6 g/plant) treatments. In terms of 100 seed weight, the MC and control treatment had the highest weight (0.3 g), which was significantly different from the PAC and PAC + MC treatment (0.2 g).
Effect of Growth Regulators on Sesame Cultivars for Agronomic Traits
The various doses of PAC, MC, and PAC + MC made a substantial impact on the cultivars (Table 2). Under the PAC treatment, Muganli 57 exhibited a pronounced dominance in several yield components. The cultivar produced a significantly higher number of capsules (167.3) compared to Paloma’s 43.7. Similarly, Muganli 57 had more branches, with 7.8 compared to Paloma’s 3.3. The stem length of the first capsule was also longer in Muganli 57 at 77.3 cm, while Paloma lagged at 62.6 cm. In terms of plant height, Muganli 57 reached 190.6 cm, towering over Paloma’s height of 132.3 cm. A stark difference was observed in seed yield, with Muganli 57 producing 40.9 g/plant, dwarfing Paloma’s yield of 1.7 g/plant. However, both cultivars were relatively close in 100 seed weight, with Muganli 57 at 0.3 g and Paloma at 0.2 g.
For the MC treatment, Muganli 57 continued to outperform. It produced 149.5 capsules, more than double Paloma’s 63.0. Branching was also superior in Muganli 57 with 7.0 branches, compared to Paloma’s 4.5. The stem lengths to the first capsule were closer between the two cultivars, with Muganli 57 at 72.1 cm and Paloma at 68.1 cm. In terms of height, Muganli 57 achieved 179.8 cm, while Paloma reached 140.3 cm. The seed yield for Muganli 57 was 23.5 g/plant, significantly higher than Paloma’s 2.7 g/plant. Both cultivars had similar 100 seed weights, with Muganli 57 at 0.3 g and Paloma at 0.2 g.
When treated with the combined Paclobutrazol and Mepiquat Chloride (PAC + MC), Muganli 57 maintained its superior performance. It yielded 129.6 capsules, compared to Paloma’s 71.0. Muganli 57 had 6.6 branches, while Paloma had 5.0. The stem lengths to the first capsule were relatively close, with Muganli 57 measuring 76.0 cm and Paloma at 69.1 cm. In terms of plant height, Muganli 57 stood at 166.5 cm, taller than Paloma’s 141.0 cm. The seed yield difference remained notable, with Muganli 57 producing 19.8 g/plant and Paloma yielding only 3.1 g/plant. However, there was a more pronounced difference in the 100 seed weight, with Muganli 57 weighing in at 0.3 g and Paloma at 0.1 g. Totally, across all treatments, Muganli 57 consistently demonstrated superior yield and yield components compared to the Paloma cultivar. The effects of the growth regulators were evident, with each treatment showing distinct impacts on the two cultivars, but the overarching trend of Muganli 57’s dominance remained consistent.
Effect of Doses of Applications on Agronomic Traits
In this study, three different doses of the growth regulators (PAC, MC, and PAC + MC) were applied and compared with control conditions (Table 2). Results showed that there were significant differences among doses of the studied traits in the paclobutrazol application. For the number of branches, the control dose (A0) exhibited the highest value of 8.2, which was significantly different from the other doses. The stem length of the first capsule was the longest for the A02 and A03 doses, both showing values above 74 cm. In terms of seed yield, the A03 dose showed the highest yield at 29.7 g/plant, while the A01 and A02 doses had similar yields of 19.6 and 20.4 g/plant, respectively. The 100 seed weight remained relatively consistent across all doses.
The control dose (A0) for mepiquat chloride treatment had the highest number of branches at 8.2. The stem length to the first capsule was the longest for the A03 dose at 83.8 cm, while the A01 and A02 doses had shorter lengths of 61.7 and 69.8 cm, respectively. The 100 seed weight was highest for the A03 dose at 0.4 g, with the A01 and A02 doses both showing weights of 0.2 and 0.3 g, respectively. For the combined treatment, in terms of the number of branches, the control dose (B0) exhibited the highest value at 8.2. The stem length to the first capsule was the longest for the B02 dose at 82.3 cm, followed closely by the B03 dose at 79.8 cm. The B01 and B0 doses had shorter stem lengths of 68.0 and 65.1 cm, respectively. The 100 seed weight remained relatively consistent across all doses.
The results indicate that the application of Paclobutrazol, Mepiquat chloride, and their combined treatment had varying effects on the yield and yield components of sesame, depending on the dose. The control doses generally exhibited superior performance in terms of the number of branches, while specific doses within each treatment showed advantages in other yield components.
Effect of Growth Regulators on Oil Content and Fatty Acid Composition
A significant variation in both oil content and fatty acid compositions was found among all the applications (Table 3). The results indicate that the application of PAC, MC, and PAC + MC treatment had distinct effects on the oil content and fatty acid composition of sesame. While control treatment resulted in the highest oil content, PAC treatment led to higher contents of oleic and stearic acids. The combined treatment showed intermediate effects for most fatty acids. The application of PAC resulted in the highest oil content of 49.4% among the growth regulators. The MC and PAC + MC treatments yielded slightly lower oil contents of 48.6% and 48.2%, respectively. The MC treatment and control exhibited the highest palmitic acid content at 9.5%, followed by the combined P + MC treatment at 9.4%. The PAC treatment had the lowest content at 9.3%. The PAC treatment showed the highest stearic acid content at 4.8%, while both the MC and PAC + MC treatments had similar contents of 4.5%. The PAC treatment led to the highest oleic acid content at 43.4%. The PAC + MC treatment had a slightly lower content at 41.0%, and the MC and control treatment had the lowest at 40.8%. Both the MC and PAC + MC treatments had similar linoleic acid contents of 44.2% and 44.1%, respectively, which were higher than the PAC treatment’s content of 41.6%. The MC treatment had the highest linolenic acid content at 0.4%, while both the PAC and PAC + MC treatments had similar lower contents at 0.3%.
Effect of Growth Regulators on Sesame Cultivars for Oil Traits
The study aimed to evaluate the effects of Paclobutrazol, Mepiquat chloride, and their combined application on the oil content and fatty acid composition of two sesame cultivars, Muganli-57 and Paloma. The results indicate that the application of Paclobutrazol, Mepiquat chloride, and their combined treatment had distinct effects on the oil content and fatty acid composition of the two sesame cultivars. Muganli-57 generally exhibited higher oil and fatty acid contents than Paloma across the treatments. By studying the paclobutrazol treatment, it can be seen that for the cultivar Muganli-57, the oil content was 53.0%, which was significantly higher than Paloma’s oil content of 45.7%. Muganli-57 exhibited a higher palmitic acid content (9.5%) compared to Paloma (9.0%). Conversely, Paloma had a slightly higher stearic acid content (4.9%) than Muganli-57 (4.7%). Both cultivars showed similar oleic acid contents, with Paloma having a slightly higher value (43.9%) than Muganli-57 (42.9%). For linoleic acid, Muganli-57 had a higher content (42.0%) than Paloma (41.2%). Paloma exhibited a higher linolenic acid content (0.3%) compared to Muganli-57 (0.2%). Both cultivars had similar arachidic acid contents.
For mepiquat chloride treatment, Muganli-57 had a higher oil content (52.7%) compared to Paloma (44.5%). Palmitic acid content was higher in Muganli-57 (9.9%) than in Paloma (9.0%). Paloma exhibited a slightly higher stearic acid content (4.7%) compared to Muganli-57 (4.4%). Oleic acid content was slightly higher in Paloma (41.4%) than in Muganli-57 (40.1%). Linoleic acid content was higher in Muganli-57 (44.8%) compared to Paloma (43.6%). Paloma had a higher linolenic acid content (0.4%) than Muganli-57 (0.2%). Arachidic acid content was higher in Paloma (0.5%) compared to Muganli-57 (0.4%).
For the combined treatment, Muganli-57 exhibited a higher oil content (52.6%) than Paloma (43.8%). Palmitic acid content was higher in Muganli-57 (9.8%) compared to Paloma (9.0%). Paloma had a slightly higher stearic acid content (4.7%) than Muganli-57 (4.3%). Oleic acid content was slightly higher in Paloma (41.3%) compared to Muganli-57 (40.6%). Linoleic acid content was higher in Muganli-57 (44.3%) than in Paloma (43.9%). Paloma exhibited a higher linolenic acid content (0.3%) compared to Muganli-57 (0.2%). Both cultivars had similar arachidic acid contents.
Effect of Doses of Applications on Fatty Acid Composition
The results elucidate that the application of different doses of paclobutrazol, mepiquat chloride, and their combination distinctly influences the oil content and fatty acid composition in sesame (Table 4). The control groups (A0 and B0) generally exhibited higher oil and fatty acid contents across the treatments, providing a nuanced understanding of how these growth regulators can modify the oil profile of sesame under varied doses. For the PAC treatment, the control dose (A0) exhibited the highest oil content (51.3%), followed by A03 (49.2%), A01 (48.8%), and A02 (48.2%). Palmitic acid content was highest in A03 and A0 (both 9.5%), followed by A01 (9.3%) and A02 (8.8%). Stearic acid content was highest in A02 (5.3%), followed by A01 (5.0%), A03 (4.7%), and A0 (4.4%). Oleic acid content was highest in A02 (46.3%), followed by A03 (42.3%), A01 (44.0%), and A0 (40.9%). Linoleic acid content was highest in A0 (44.7%), followed by A03 (42.5%), A01 (40.6%), and A02 (38.5%). Linolenic acid content was highest in A02 (0.4%), followed by A01 (0.3%), A0 (0.2%), and A03 (0.2%). Arachidic acid content was highest in A02 (0.5%), followed by A01 (0.4%), A03 (0.4%), and A0 (0.4%).
For the Mepiquat chloride treatment, the control dose (A0) showed the highest oil content (51.3%), followed by A03 (48.7%), A01 (48.0%), and A02 (46.6%). Palmitic acid content was highest in A01 (9.6%), followed by A03 (9.6%), A0 (9.5%), and A02 (9.2%). Stearic acid content was highest in A01 (4.7%), followed by A02 and A03 (both 4.5%), and A0 (4.4%). Oleic acid content was highest in A01 and A0 (both 41.0%), followed by A03 (40.8%), and A02 (40.3%). Linoleic acid content was highest in A02 (45.0%), followed by A03 and A0 (both 44.2%), and A01 (43.6%). Linolenic acid content was highest in A01 (0.4%), followed by A02, A03, and A0 (all 0.3%). Arachidic acid content was highest in A01 (0.5%), followed by A02 and A03 (both 0.4), and A0 (0.4).
For the combined treatment, the control dose (B0) exhibited the highest oil content (51.2%), followed by B01 (48.1%), B03 (47.7%), and B02 (45.9%). Palmitic acid content was highest in B0 (9.5%), followed by B01 (9.4%), B02 (9.5%), and B03 (9.2%). Stearic acid content was highest in B03 (4.7%), followed by B01 (4.6%), B02 (4.4%), and B0 (4.4%). Oleic acid content was highest in B01 (41.6%), followed by B0 (41.1%), B03 (40.7%), and B02 (40.5%). Linoleic acid content was highest in B02 (44.7%), followed by B03 (44.4%), B01 (43.9%), and B0 (44.0%). Linolenic acid content was highest in B01 and B02 (both 0.3%), followed by B03 and B0 (both 0.2%). Arachidic acid content was similar across all doses.
Correlation Analysis of the Studied Traits
Our comprehensive correlation analysis elucidated significant relationships among the morphological and biochemical traits across the studied genotypes subjected to various treatment conditions (Fig. 2). The analysis brought to light several noteworthy associations, highlighting the intricate interplay between plant structure, yield, and biochemical composition. A paramount finding was the strong positive correlation between the number of capsules and several key plant traits. Specifically, the number of branches (r = 0.726), plant height (r = 0.895), and seed yield (r = 0.889) were all positively correlated with the number of capsules. This correlation underscores the critical role of capsule count as a pivotal indicator of plant health, vigor, and productivity, suggesting that plants exhibiting a higher capsule count generally manifest increased branching, taller stature, and enhanced yield. Further, seed yield demonstrated significant correlations with a suite of traits. It was positively correlated with the number of branches (r = 0.599), plant height (r = 0.845), and the 1000 seed weight (r = 0.680). Additionally, a noteworthy correlation was observed between seed yield and oil content (r = 0.735), indicating that plants with higher yields tend to also have superior oil production. This relationship is particularly salient for breeding and cultivation strategies aimed at maximizing oil yield alongside seed production.

Analysis of trait interrelationships through correlation analysis
The analysis also revealed that oil content was significantly correlated with the fatty acid profile of the seeds. A notable positive correlation with palmitic acid content (r = 0.739) and a negative correlation with stearic acid (r = -0.563) were observed. This suggests that variations in oil content attributable to different treatments may concurrently influence the fatty acid composition of the oil. The positive correlation between oil content and the 1000 seed weight (r = 0.724) further implies that interventions that augment seed size could potentially enhance oil yield. Intriguing patterns emerged from the correlation analysis of fatty acids. Palmitic acid displayed a positive correlation with oil content (r = 0.739) and an inverse relationship with stearic acid (r = -0.638), hinting at a biochemical trade-off in the synthesis of these fatty acids. Moreover, a pronounced inverse correlation between oleic acid and linoleic acid (r = -0.979) was detected, underscoring the competitive biosynthesis pathways in oilseed plants where the augmentation of one fatty acid often occurs at the detriment of another. Linolenic acid exhibited a notable negative correlation with both the number of capsules (r = -0.667) and seed yield (r = -0.605), suggesting that higher concentrations of linolenic acid might be linked to reduced productivity under certain conditions. Conversely, arachidic acid was positively correlated with stearic acid (r = 0.871) and negatively with oil content (r = -0.548), indicating its prevalence in scenarios less conducive to oil accumulation. These correlations elucidate the complex relationships governing plant morphology, yield, and oil composition, offering valuable insights for the optimization of agronomic practices. By leveraging these findings, it may be possible to tailor treatment conditions to enhance desired plant traits, including yield, oil content, and specific fatty acid compositions, thereby informing targeted breeding and cultivation strategies.
Cluster Analysis of the Studied Treatments
The cluster analysis of different treatments and their impact on various traits of Paloma and Muganli57 genotypes revealed significant insights into optimizing plant growth conditions for both productivity and quality (Fig. 3). The analysis delineated four distinct clusters based on the treatments applied (PAC, MC, PAC + MC, Ctrl) and doses (0.1, 0.2, 0.3, and control), alongside measured traits including the number of capsules, number of branches, stem length to the first capsule, plant height, seed yield, 1000 seed weight, oil content, and fatty acid composition. Cluster 1 comprised treatments with PAC and MC at lower doses (0.1 and 0.2). This cluster was characterized by moderate values across the traits of interest, with an oil content of around 44%. The plants within this cluster demonstrated a balanced fatty acid composition, indicative of good quality, albeit not the highest productivity levels. Cluster 2 encompassed a broader range of treatments, including PAC, MC, their combinations at varying doses, and control plants. This cluster exhibited a wide diversity in plant traits, with seed yield showing considerable variation. Interestingly, the oil content in this cluster tended to be lower than that observed in Cluster 1, particularly for the PAC + MC 0.2 treatment. The presence of control treatments within this cluster suggested that untreated plants could perform comparably to treated ones in specific contexts, both in terms of productivity and quality. Cluster 3 primarily consisted of plants treated with MC and PAC + MC at higher doses, alongside some control samples. This cluster was notable for its generally higher seed yield, particularly in specimens treated with PAC_0.3 and MC, pointing towards enhanced productivity. Furthermore, the oil content was markedly higher in this cluster, especially in treatments like PAC + MC_0.1 and MC alone, suggesting that these treatments could bolster quality alongside productivity. Cluster 4, although the smallest, was distinguished by a single treatment of PAC at 0.2. This cluster boasted the highest seed yield among all the groups, albeit with slightly lower oil content compared to the highest values observed in other clusters. This finding underscored a focus on productivity, with the substantial oil content also indicating good quality.

Comprehensive cluster analysis of treatment effects on key variables
Principal Component Analysis (PCA) of the Studied Treatments
Principal component analysis (PCA) was performed to explore the impact of different growth regulators and their concentrations on two genotypes, ‘Muganli57’ and ‘Paloma,’ across multiple phenotypic traits (Fig. 4). The PCA revealed that the first four principal components (PCs) cumulatively explained 88.443% of the total variance within the dataset, highlighting the significant patterns of trait variation under different treatment conditions. Specifically, PC1 accounted for 54.339% of the variance, emphasizing its dominant role in capturing the effects of the treatments on the phenotypic traits. PC2 contributed to 20.003%, PC3 to 7.962%, and PC4 to 6.1393% of the variance, respectively. The distribution of treatments across the PCs underscored the differential response of the ‘Paloma’ and ‘Muganli57’ genotypes to the applied treatments. For instance, ‘Paloma’ treated with Paclobutrazol at different concentrations (0.1, 0.2, and 0.3 CC in 1 L) demonstrated a pronounced shift along PC1 and PC2, indicating a significant alteration in traits associated with these components. Similarly, the ‘Muganli57’ genotype exhibited a distinct pattern, particularly in response to Paclobutrazol and Pix treatments, suggesting genotype-specific responses to growth regulation. Trait loadings on the principal components revealed the contributions of various phenotypic traits to the observed variance. The number of capsules, plant height, seed yield, and oil content were among the traits that were heavily loaded on PC1, indicating their significant influence on the differentiation observed under different treatment conditions. The loadings on PC2, PC3, and PC4 further highlighted the complex interplay of traits such as stem length to the first capsule, 1000 seed weight, and fatty acid composition in shaping the overall phenotype under various treatments. Further analysis of PCA scores for each treatment combination across genotypes highlighted how specific treatments influenced the traits associated with PCs. This facilitated a nuanced evaluation of genotype performance under varying treatment conditions. Comparison between the two genotypes, Muganlı and Paloma, revealed distinct responses to the treatments. Muganlı consistently exhibited positive scores on PC1 for treatments such as PAC at 0.3 CC in 1 L, and MC at lower doses, and their combinations, indicating superior performance in traits pivotal for productivity and quality. Conversely, Paloma demonstrated negative scores with PAC treatments, suggesting reduced performance in key traits. However, this genotype might outperform in alternative specific conditions not directly captured by PC1. The analysis of treatment doses, specifically Paclobutrazol (PAC), Pix (MC), and their combinations, illuminated the variable effectiveness of these applications contingent upon the genotype. For Muganlı, a higher dose of PAC (0.3 CC in 1 L) and lower doses in combinations (e.g., PAC + MC at 0.1 CC in 1 L) were identified as optimal, suggesting these conditions favorably balance growth regulation with productivity. In contrast, Paloma appeared to benefit more from control conditions or lower treatment doses, as indicated by better scores on PC2, hinting at a genotype-specific sensitivity to the intensity of applied treatments. Based on the comprehensive PCA, Muganlı emerged as the more resilient genotype across a spectrum of treatments, consistently showing positive associations with productivity and quality traits on PC1. Accordingly, for Muganlı, PAC applied at 0.3 CC in 1 L or combination with MC at 0.1 CC in 1 L is recommended to harness the best performance. On the other hand, Paloma, as suggested by PCA score trends, may achieve optimal outcomes under control conditions or with lower doses of treatments, indicating a nuanced sensitivity to the treatment regimen. This detailed PCA-based assessment provides a strategic guide for selecting genotype and treatment combinations that optimize productivity and quality, offering valuable insights for targeted agricultural practices.

Unraveling the multidimensional impact of treatments using principal component analysis (PCA)
Best Dose and Treatment Recommendations
For maximizing productivity, the PAC treatment at 0.3 (Cluster 3) and 0.2 (Cluster 4) emerged as the optimal strategies, showcasing the highest seed yields (Fig. 5). When considering quality, as evidenced by oil content and fatty acid composition, MC treatments (Cluster 3) and lower doses of PAC + MC (also in Cluster 3) were found to enhance quality without significantly compromising productivity. For those seeking a balanced choice between productivity and quality, the PAC + MC combination at 0.1 (Cluster 3) presented itself as an excellent option, offering a harmonious balance between these two critical objectives. In summary, the selection of the best dose and treatment hinges on specific goals: PAC at higher doses is preferable for productivity, while MC and PAC + MC at moderate doses are recommended for achieving a balance of productivity and quality. There is a slight preference for the PAC + MC combination at 0.1 for those aiming to strike an optimal balance between these objectives.

Optimal dose and treatment recommendations for enhanced sesame plants performance
Discussion
Sesame is a valuable oilseed crop with significant economic importance in many developing countries. Research aimed at enhancing seed yield and quality is crucial (Myint et al. 2020). The application of growth regulators, such as Paclobutrazol (PAC) and Mepiquat chloride (MC), and their combined use has been a topic of interest in plant physiology, primarily due to their potential to modify plant growth and development (Bañón et al. 2023). While numerous studies have highlighted the efficacy of triazoles, particularly paclobutrazol, as beneficial anti-gibberellic triazoles (Peng et al. 2014; Kuai et al. 2015; Kamran et al. 2018), limited research exists on the impact of different concentrations and application techniques of growth regulators on sesame’s physiological growth and oil content (Mehmood et al. 2021). Also, our study extends this body of research by exploring the nuanced impact of these growth regulators, both individually and in combination, on sesame’s physiological growth and oil content, an area that has received limited attention thus far.
In this study, growth regulator application increased the number of capsules per plant. Specifically, the PAC + MC (at B01) and paclobutrazol applications (at A01) yielded the most capsules. It seems that the two growth regulators had a synergistic impact on reproductive capability, as the number of capsules increased with the combination treatment (PAC + MC). This finding underscores the potential of a tailored approach in the application of growth regulators to enhance sesame productivity beyond traditional practices. Paclobutrazol’s effect in reducing gibberellin production leads to decreased internode elongation and increased lateral branching (Zhang et al. 2022). On the other hand, Mepiquat chloride, which inhibits cell division and elongation, likely enhanced the number of capsules by favoring a denser development pattern, thereby improving reproductive efficiency (Basra 2000). Interestingly, our findings also highlight the complexity of growth regulator interactions within the plant system, as evidenced by the nuanced effects on seed yield. The stem length to the first capsule, an essential trait for harvesting efficiency, was the longest in the combined treatment. This balance suggests that optimizing growth regulator combinations is a key strategy in improving not only yield but also ease of harvest, addressing both agronomic and operational challenges in sesame cultivation. Another possible explanation is that the levels of MC and PAC are optimally balanced (Rademacher 2015). However, it’s intriguing to note that while the combined treatment showed benefits in terms of branching and capsule formation, it did not translate to a significant increase in plant height. This observation reflects the intricate balance between the dwarfing effect of MC and the branching promotion by PAC, offering insights into their potential roles in sculpting plant architecture for enhanced yield without compromising plant stability and harvestability. One possible explanation is that the dwarfing impact of MC balanced out the pro-lateral growth effects of PAC, the two regulators functioning in opposition to one another (Rajiv et al. 2018). Seed yield, a critical determinant of the economic value of a crop, was highest in the PAC treatment. This outcome suggests that the direct application of Paclobutrazol, by influencing the plant’s hormonal balance, notably enhances its reproductive efficiency, a critical insight for sesame breeders and growers aiming to maximize yield. Although branching and capsule development are crucial, this also implies that Paclobutrazol alone may have a greater impact on sesame seed output than other physiological processes (Arnold and Dongyun 2003). The process may include increased allocation of resources to reproductive structures in the presence of PAC, potentially affecting the balance of endogenous hormones such as gibberellins and cytokinins (Zhu et al. 2004). Our results align with Mehmood et al. (2021), who found that paclobutrazol led to a higher seed yield compared to the control. Consistent with Mehmood et al. (2021), our study also found that increasing paclobutrazol doses positively influenced seed yield. The higher 100 seed weight observed in the MC treatment among growth regulators might be a manifestation of the growth regulator’s influence on cell division and elongation (Wang et al. 2022a). Such findings are pivotal, indicating that MC’s role extends beyond plant stature regulation to potentially influencing seed development processes, thereby impacting seed quality and market value. By retarding rapid cell division, MC allows for more extended nutrient accumulation in individual seeds, leading to heavier seeds (Severino and Auld 2013). This is consistent with previous studies where MC has been shown to enhance seed filling and increase seed weight in other crops. Contrary to our findings, Mehmood et al. (2021) reported increased seed weight after paclobutrazol application. These differences may result from genetic and environmental variances. Similarly, Li et al. (1993) found that paclobutrazol application post-soybean flowering increased 100-seed weight. Our results demonstrated that the physiological responses of sesame to Paclobutrazol and Mepiquat chloride, both individually and in combination, offer a complex interplay of growth and developmental processes. While the combined application shows promise in enhancing certain agronomic traits, the individual application of Paclobutrazol seems more beneficial for seed yield. These findings underscore the importance of understanding the nuanced effects of growth regulators on plant physiology to optimize their application for improved agronomic outcomes.
The differential response of the two sesame cultivars, Muganli 57 and Paloma, to the application of growth regulators underscores the intricate interplay between genotype and external growth modulators in determining plant performance. Muganli 57 consistently outperformed other varieties in terms of yield and yield components, indicating genetic advantages that make it more responsive to the growth regulators used (Johnson and Pettigrew 2006). Paclobutrazol, known for its role in inhibiting gibberellin biosynthesis, can lead to reduced internode elongation and enhanced branching (Desta and Amare 2021). Given that Muganli 57 had the most capsules and branches when treated with PAC, this might be the reason why it was so dominant. The cultivar’s inherent genetic composition appears to enhance its responsiveness to the branching-promoting effects of PAC, resulting in an increased number of capsules and, consequently, a higher seed yield. On the other hand, the reduced response of Paloma suggests a lower sensitivity to gibberellin inhibition, which correlates with a decreased number of branches and capsules. Mepiquat chloride’s primary action is to retard cell division and elongation, which can lead to more compact growth and potentially higher seed weight (Rajiv et al. 2018). While both cultivars exhibited a response to MC treatment, the variance in their responses can be linked to their distinct growth habits (Rabiei et al. 2018). Given its less vigorous nature, Paloma likely derives less benefit from the compacting effect of MC, resulting in fewer capsules and branches. The combined application of Paclobutrazol and Mepiquat chloride (PAC + MC) aimed to harness the benefits of both growth regulators. While the combined treatment did enhance certain agronomic traits, the overarching trend of Muganli 57’s dominance remained unchanged. This suggests that while external growth modulators can influence plant performance, the inherent genetic makeup of a cultivar plays a pivotal role in determining its final agronomic outcome (White and Hoogenboom 2003). This study showed that the differential response of the two sesame cultivars to growth regulators highlights the importance of genotype selection in agronomic practices. While growth regulators can optimize certain agronomic traits, the choice of cultivar remains paramount. Muganli 57, with its consistent superior performance across treatments, emerges as a promising candidate for sesame cultivation, especially when growth regulators are part of the agronomic strategy.
The use of growth regulators like Paclobutrazol (PAC) and Mepiquat chloride (MC) in our study underscored the nuanced interplay between chemical application and plant response, a theme that is consistent with broader agricultural research. As documented by Bañón (et al. 2023), the efficacy of these agents in modulating plant growth and optimizing yield is well-established, yet their dose-dependent effects necessitate a refined approach to application. Our findings contribute to this ongoing discourse, emphasizing the critical importance of dose optimization to maximize the benefits of growth regulators in agriculture. This underscores the broader implication that, while growth regulators hold substantial promise for yield enhancement, their application must be meticulously calibrated to achieve desired outcomes. In this study, we evaluated the impact of different doses of PAC, MC, and their combination (PAC + MC) on the agronomic traits of sesame. Paclobutrazol is known to inhibit gibberellin synthesis, which can lead to reduced stem elongation and enhanced branching (Desta and Amare 2021). In our study, the control dose of PAC, with no application, exhibited the highest branching, suggesting that in the absence of PAC, the plant’s natural growth habit is to produce more branches. However, as the concentration of P increased, particularly in the A03 dose (600 mg/L), there was a noticeable increase in seed yield. This indicates that under higher PAC concentrations, despite producing fewer branches, the plant reallocates its resources to reproductive growth, resulting in enhanced seed production (Anwar et al. 2017). Interestingly, the doses A02 and A03 showed the greatest stem length to the first capsule, demonstrating that while PAC generally decreases overall plant height, it does not reduce the length to the first reproductive node. Mepiquat chloride is known to retard cell elongation, leading to more compact plants (Tung et al. 2020). The controlled dose (A0) for MC, similar to PAC, showed the highest branching. This indicates that without MC, plants are inclined to develop more branches. However, as the concentration of MC increased, particularly in the A03 dose (600 mg/L), there was a noticeable increase in the stem length of the first capsule. This observation suggests that MC, while lowering overall plant height, promotes the extension of the initial stem segment before the onset of branching (Kamran et al. 2018). The highest 100 seed weight in the A03 dose suggests that higher concentrations of MC might lead to denser seeds, possibly due to its impact on cell division and elongation (Tung et al. 2019). The combination of PAC and MC aimed to harness the benefits of both regulators. The control dose (B0) showed the highest branching, consistent with the individual applications of PAC and MC. However, the combined doses, particularly B02 (200 mg P + 200 mg MC/L), showed the longest stem length to the first capsule. These findings imply a synergistic interaction between the growth-retarding effect of MC and the gibberellin-inhibiting action of PAC, resulting in increased stem elongation before branching occurs (Howard 2012). Our results showed that the dose-dependent response of sesame to PAC, MC, and their combination underscores the importance of precise dosing in agronomic practices (Qin et al. 2020). While growth regulators can enhance certain yield components, their effects can be dose-dependent, and finding the optimal dose is crucial for maximizing yield (Kamran et al. 2018). The observed variations in responses also highlight the complex interplay between vegetative and reproductive growth in plants and the role of growth regulators in modulating this balance (Bartoli et al. 2013).
The application of growth regulators, specifically Paclobutrazol (PAC) and Mepiquat chloride (MC), and their combined use (PAC + MC) has demonstrated a significant impact on the oil content and fatty acid composition of sesame, as evidenced by the results presented in Table 3. The physiological and biochemical pathways influenced by these growth regulators have led to variations in the metabolic allocation towards oil biosynthesis and fatty acid composition, which are critical aspects of the nutritional and industrial value of sesame oil (Murphy 2014). Mepiquat chloride (MC) and the combined treatment (PAC + MC) resulted in slightly lower oil contents, indicating that the physiological effects of MC, such as reduced cell elongation, might not favor lipid accumulation to the same extent as PAC (Wang et al. 2022b). PAC treatment led to the highest stearic and oleic acid contents, indicating a potential enhancement in the biosynthesis or conservation of these fatty acids. The observed patterns reflect PAC’s influence on plant metabolism and development, which may alter the activity of enzymes responsible for the desaturation and elongation of fatty acids (Whitaker and Wang 1987). MC treatment led to higher content linoleic and linolenic acids, which are polyunsaturated fatty acids compared to PAC and PAC + MC. It appears that MC preferentially supports the desaturation pathways of fatty acids, a process possibly tied to its role in adjusting plant growth and development, with implications for membrane fluidity and function (Wang et al. 2022a). The presence of higher arachidic acid content in plants treated with PAC and MC individually, as opposed to the combined treatment, implies that the separate applications of these growth regulators may specifically promote the synthesis or conservation of this saturated fatty acid through distinct metabolic or regulatory pathways (Kim et al. 2011).
The application of growth regulators, specifically Paclobutrazol (PAC) and Mepiquat chloride (MC), and their combined use (P + MC), has shown to exert differential effects on the oil content and fatty acid composition of sesame cultivars, Muganli-57 and Paloma. The physiological and biochemical responses of these cultivars to the growth regulators provide insights into the intricate interplay between plant genetics and external growth modulation. Muganli-57 consistently exhibited a higher oil content across all treatments compared to Paloma. The genetic composition of Muganli-57 appears to predispose it towards higher lipid accumulation, likely due to more efficient lipid biosynthesis pathways or a superior allocation of resources to oil production (Kumar et al. 2022). The differential response of the two cultivars to growth regulators underscores the importance of genotype-environment interactions in determining crop quality traits (Mecha et al. 2022). With PAC treatment, the increased oil content observed in Muganli-57 could be due to its enhanced response to gibberellin inhibition, which reallocates metabolic resources from vegetative growth towards reproductive development and lipid accumulation (Chandra and Roychoudhury 2020). The differences in fatty acid composition between the two cultivars, notably in palmitic, stearic, and linoleic acids, indicate that Paclobutrazol regulates the enzymes involved in fatty acid biosynthesis and modification differently in Muganli-57 compared to Paloma (Yang et al. 2023). The impact of Mepiquat chloride on reducing cell elongation likely results in a more compact plant structure, which could affect lipid biosynthesis pathways distinctly between the two cultivars (Kentelky et al. 2021). The elevated oleic acid content in Paloma implies that MC’s influence on the desaturation pathways may be more significant in this cultivar than in Muganli-57. The combined application of PAC and MC seemed to bring about an intermediate response in the two cultivars. While Muganli-57 still maintained its dominance in terms of oil content, the differences in fatty acid composition became more nuanced. The combined application of PAC and MC is posited to trigger a more complex physiological response, affecting multiple pathways involved in fatty acid synthesis and modification, suggesting an intricate interplay between these growth regulators and plant metabolic processes (Kim et al. 2018). The differential response of Muganli-57 and Paloma to growth regulator treatments highlights the importance of cultivar selection in sesame cultivation (Pathak et al. 2014). While growth regulators can modulate plant growth and development, the inherent genetic potential of the cultivar plays a pivotal role in determining the final oil content and fatty acid profile (Subedi et al. 2020). For sesame breeders and growers aiming to enhance oil quality, a combined approach of selecting high-yielding cultivars and optimizing growth regulator applications might be the most effective strategy (Tripathy et al. 2019; Myint et al. 2020).
The application of growth regulators, specifically Paclobutrazol (PAC) and Mepiquat chloride (MC), and their combined use (PAC + MC), has shown to exert differential effects on the oil content and fatty acid composition of sesame, contingent upon the dosage applied. The physiological and biochemical responses of sesame to these growth regulators provide insights into the intricate interplay between external growth modulation and lipid metabolism (Mehmood et al. 2021). The control group had the highest oil content, while paclobutrazol led to a slight decrease. In contrast, mepiquat chloride treatment resulted in the lowest oil content. The combined PAC + MC treatment showed intermediate effects. Our findings align with Kim et al. (2018), who observed similar trends in oil content with growth regulator applications. Significant variations in fatty acids were observed across applications and doses (Table 4). Compared to the control group, saturated fatty acid levels increased, but the PAC + MC application reduced palmitic acid levels and increased stearic acid levels. Unsaturated fatty acids, such as oleic acid, linoleic acid, linolenic acid, and arachidic acid, showed slight increases, similar to the control group. This diverges from Kim (2018), who found reduced levels of saturated fatty acids relative to unsaturated ones with paclobutrazol treatment. Similarly, the mepiquate chloride application reduced the overall saturated fatty acid content in the cotton seed (Sawan et al. 2007).
Our study contributed to the growing evidence on the strategic use of growth regulators in oilseed crops, highlighting the promise of Paclobutrazol and Mepiquat chloride in modulating not just growth, but also reproductive outcomes and seed quality in sesame. Moreover, the differential cultivar responses observed further emphasize the importance of cultivar-specific management practices to fully leverage the benefits of growth regulator applications in sesame cultivation.
Conclusion
Our study provided critical insights into the nuanced roles of Paclobutrazol (PAC) and Mepiquat chloride (MC), both individually and in combination, on sesame cultivation, a topic that has been underexplored in agricultural research. The differential response of sesame to varying doses of growth regulators underscores the importance of dose optimization in influencing oil content and fatty acid composition. While growth regulators can modulate lipid metabolism, the dose-specific responses highlight the need for a balanced approach. This nuanced understanding challenges the conventional one-size-fits-all approach to the use of growth regulators in crop management, advocating for a more tailored strategy that considers the specific needs and responses of the crop.
Excessive application might not always lead to enhanced oil quality, and in some cases, might even be counterproductive. The detailed analyses affirm Muganli57 as the superior genotype for achieving high productivity and quality under specified treatments. Specifically, PAC at 0.3 CC in 1 L and the PAC + MC combination at 0.1 CC in 1 L are recommended to optimize these outcomes. These findings not only contribute to the body of knowledge on sesame agronomy but also provide a practical framework for farmers and agricultural practitioners to improve yield and oil quality through precise growth regulator management.
Moreover, our research highlights the potential of integrating genetic selection with agronomic practices to enhance crop performance. The identification of Muganli57 as a particularly responsive genotype to growth regulator treatments underscores the importance of selecting cultivars that are well-suited to specific agronomic interventions. These findings provide a strategic guide for selecting genotype and treatment combinations that enhance agricultural productivity and quality, offering valuable insights for targeted crop management practices.
In conclusion, the application of growth regulators offers a promising avenue to modulate the oil content and fatty acid composition of sesame. However, understanding the dose-specific responses is crucial to harness the full potential of these external modulators. Moving forward, our study called for further research into the interactive effects of growth regulators and genotypes under different environmental conditions to fully realize the benefits of these practices in sustainable agriculture. By doing so, we can develop more resilient and productive cropping systems that meet the growing demands for agricultural products.
References
Ahmed J, Qadir G, Ansar M et al (2023) Shattering and yield expression of sesame (Sesamum indicum L) genotypes influenced by paclobutrazol concentration under rainfed conditions of Pothwar. BMC Plant Biol 23:1–13
Anwar S, Kuai J, Khan S et al (2017) Soaking seeds with paclobutrazol enhances winter survival and yield of rapeseed in a rice-rapeseed relay cropping system. Int J Plant Prod 11:491–504
Arnold R, Dongyun X (2003) Environmental and cultural influences on flowering of Eucalyptus dunnii. Aciar Proceedings ACIAR 1998:140–147
Arteca RN (2014) Introduction to horticultural science. Cengage Learning
Ashri A (1998) Sesame breeding. Plant Breed Rev 16:179–228
Bañón D, Ortuño MF, Sánchez-Blanco MJ et al (2023) Effects of Paclobutrazol and Mepiquat Chloride on the physiological, nutritional, and morphological behavior of Potted Icterina Sage under Greenhouse conditions. Agronomy 13(8):2161
Bartoli CG, Casalongué CA, Simontacchi M et al (2013) Interactions between hormone and redox signaling pathways in the control of growth and cross tolerance to stress. Environ Exp Bot 94:73–88
Basra A (2000) Plant growth regulators in agriculture and horticulture: their role and commercial uses. CRC
C MK, SA S (2015) Bioactive lignans from sesame (Sesamum indicum L.): evaluation of their antioxidant and antibacterial effects for food applications. J Food Sci Technol 52:2934–2941
Chandra S, Roychoudhury A (2020) Penconazole, paclobutrazol, and triacontanol in overcoming environmental stress in plants. In: Protective Chemical Agents in the Amelioration of Plant Abiotic Stress: Biochem Mol Persp. pp 510–534
Ciaffi M, Tozzi L, Borghi B et al (1996) Effect of heat shock during grain filling on the gluten protein composition of bread wheat. J Cereal Sci 24:91–100
Desta B, Amare G (2021) Paclobutrazol as a plant growth regulator. Chem Biol Technol Agric 8:1–15
Garces R, Mancha M (1993) One-step lipid extraction and fatty acid methyl esters preparation from fresh plant tissues. Anal Biochem 211:139–143
Ghafoorunissa HS, MVV R (2004) Sesame lignans enhance antioxidant activity of vitamin E in lipid peroxidation systems. Mol Cell Biochem 262:195–202
Howard M (2012) Comparing the non-target effects of DMI fungicides on creeping bentgrass putting greens. repository.lib.ncsu.edu
Institute S (2011) SAS/STAT statistical analysis SystemManual (v.9.3. SAS Institute, Cary,NC
Johnson J, Pettigrew W (2006) Effects of mepiquat pentaborate on cotton cultivars with different maturities. J Cotton Sci 10:128–135
Kamran M, Cui W, Ahmad I et al (2018) Effect of paclobutrazol, a potential growth regulator on stalk mechanical strength, lignin accumulation and its relation with lodging resistance of maize. Plant Growth Regul 84:317–332
Kentelky E, Szekely-Varga Z, Bálint J, Balog A (2021) Enhance growth and flower quality of Chrysanthemum indicum L. with application of plant growth retardants. Horticulture 7:532
Khan MSH, Wagatsuma T, Akhter A, Tawaraya K (2009) Sterol biosynthesis inhibition by paclobutrazol induces greater aluminum (Al) sensitivity in Al-tolerant rice. Amer J Plant Physiol 4:89–99
Kim SK, Lee HD, Choi HJ (2011) Effects of mepiquat chloride and trinexapac-ethyl on oil composition, seed yield and endogenous gibberellins in flax. Kor J Plant Resour 24:696–701
Kim SK, Han CM, Shin JH, Kwon TY (2018) Effects of paclobutrazol and prohexadione-ca on seed yield, and content of oils and gibberellin in flax grown in a greenhouse. Kor J Crop Sci 63:265–271
Koutroubas SD, Damalas CA (2015) Sunflower response to repeated foliar applications of paclobutrazol. Planta Daninha 33:129–135
Koutroubas SD, Damalas CA (2020) Physiology and yield of confection sunflower under different application schemes of Mepiquat chloride. Agriculture 10:15
Kuai J, Yang Y, Sun Y et al (2015) Paclobutrazol increases canola seed yield by enhancing lodging and pod shatter resistance in Brassica napus L. Field Crops Res 180:10–20
Kumar S, Saini R, Suthar P et al (2022) Plant secondary metabolites: Their food and therapeutic importance. In: Plant Secondary Metabolites: Physico-Chemical Properties and Therapeutic Applications. pp 371–413
Langham DR, Wiemers T (2002) Progress in mechanizing sesame in the US through breeding. In: Trends in new crops and new uses. In: Proc Fifth Nat Symp Atlanta GA USA. ASHS Press. pp 157–173
Li PQ, Zhang MF, Chen SK (1993) Physiological effect of paclobutrazol spray on soybean. Oil Crops Chin 2:29–31
Mecha E, Erny GL, Guerreiro AC et al (2022) Metabolomics profile responses to changing environments in a common bean (Phaseolus vulgaris L.) germplasm collection. Food Chem 370:131003
Mehmood MZ, Qadir G, Afzal O et al (2021) Paclobutrazol improves sesame yield by increasing dry matter accumulation and reducing seed shattering under rainfed conditions. Int J Plant Prod 15:337–349
Murphy DJ (2014) Using modern plant breeding to improve the nutritional and technological qualities of oil crops. OCL 21:D607
Myint D, Gilani SA, Kawase M, Watanabe KN (2020) Sustainable sesame (Sesamum indicum L.) production through improved technology: an overview of production, challenges, and opportunities in Myanmar. Sustainability 12:3515
Pathak N, Rai AK, Kumari R et al (2014) Sesame crop: an underexploited oilseed holds tremendous potential for enhanced food value. Agricul Sci 5:519–529
Peng D, Chen X, Yin Y et al (2014) Lodging resistance of winter wheat (Triticum aestivum L.): lignin accumulation and its related enzyme activities due to the application of paclobutrazol or gibberellin acid. Field Crops Res 157:1–7
Prakash M, Ganesan J (2000) Effect of plant growth regulators and micronutrients on certain growth analysis parameters in sesame. Sesame Safflower Newsl 15:48–58
Qin R, Noulas C, Wysocki D et al (2020) Application of plant growth regulators on soft white winter wheat under different nitrogen fertilizer scenarios in irrigated fields. Agriculture 10:305
Qureshi M, Langham DR, Lucas SJ et al (2022) Breeding history for shattering trait in sesame: classic to genomic approach. Mol Biol Rep 49:7185–7194
Rabiei B, Bahador S, Kordrostami M (2018) The expression of monoterpene synthase genes and their respective end products are affected by gibberellic acid in Thymus vulgaris. J Plant Physiol 230:101–108
Rademacher W (2015) Plant growth regulators: backgrounds and uses in plant production. J Plant Growth Regul 34:845–872
Rajiv G, Jawaharlal M, Subramanian S et al (2018) Effect of plant growth retardants on the growth and flowering of nerium (Nerium oleander L.) Cv. Red Chem Sci Rev Lett 7:875–879
Raza MA, Feng LY, Manaf A et al (2018) Sulphur application increases seed yield and oil content in sesame seeds under rainfed conditions. Field Crops Res 218:51–58
Sawan ZM, Fahmy AH, Yousef SE (2007) Cotton seed yield, seed viability, and seedling vigor as affected by nitrogen, potassium, phosphorus, zinc, and a plant growth retardant. Afr J Plant Sci Biotechnol 1:16–25
Severino LS, Auld DL (2013) A framework for the study of the growth and development of castor plant. Ind Crops Prod 46:25–38
Shah SH, Islam S, Parrey ZA, Mohammad F (2021) Role of exogenously applied plant growth regulators in growth and development of edible oilseed crops under variable environmental conditions: a review. J Soil Sci Plant Nutr 21:3284–3308
Sharafi Y, Majidi MM, Goli SAH, Rashidi F (2015) Oil content and fatty acids composition in Brassica species. Int J Food Prop 18:2145–2154
Subedi U, Jayawardhane KN, Pan X et al (2020) The potential of genome editing for improving seed oil content and fatty acid composition in oilseed crops. Lipids 55:495–512
Suman M, Sangma PD, Meghawal DR, Sahu OP (2017) Effect of plant growth regulators on fruit crops. J Pharmacognosy Phytochem 6:331–337
Syahputra BS, Sinniah UR, Omar SRS, Ismail MR (2013) Changes in gibberellic acid (GA3) content in Oryza sativa due to paclobutrazol treatment. J Food Pharma Sci. https://doi.org/10.14499/jfps
Tesfahun W (2018) A review on: response of crops to paclobutrazol application. Cogent Food Agricul 4:1
Tripathy SK, Kar J, Sahu D (2019) Advances in sesame (Sesamum indicum L.) breeding. In: Advances in Plant Breeding Strategies: Ind Food Crops. pp 577–635
Tung SA, Huang Y, Ali S et al (2019) Mepiquat chloride effects on potassium acquisition and functional leaf physiology as well as lint yield in highly dense late-sown cotton. Ind Crops Prod 129:142–155
Tung SA, Huang Y, Hafeez A et al (2020) Morpho-physiological effects and molecular mode of action of mepiquat chloride application in cotton: a review. J Soil Sci Plant Nutr 20:2073–2086
Ustun R, Yol E, Ikten C et al (2017) Screening, selection and real-time qPCR validation for phytoplasma resistance in sesame (Sesamum indicum L. Euphytica 213:1–9
Uzun B, Çağırgan MI (2006) Comparison of determinate and indeterminate lines of sesame for agronomic traits. Field Crops Res 96:13–18
Uzun B, Arslan Ç, Furat Ş (2008) Variation in fatty acid compositions, oil content and oil yield in a germplasm collection of sesame (Sesamum indicum L. J Amer Oil Chemists’ Soc 85:1135–1142
Vineeth TV, Kumar P, Krishna GK (2016) Bioregulators protected photosynthetic machinery by inducing expression of photorespiratory genes under water stress in chickpea. Photosynthetica 54:234–242
Wang N, Wang X, Qi Q et al (2022a) Analysis of the effects of mepiquat chloride priming on the seedling growth-promoting in cotton under salt stress by multi-omics. Ind Crops Prod 186:115296
Wang X, Zhou Q, Wang X et al (2022b) Mepiquat chloride inhibits soybean growth but improves drought resistance. Front Plant Sci 13:982415
Were BA, Onkware AO, Gudu S et al (2006) Seed oil content and fatty acid composition in East African sesame (Sesamum indicum L.) accessions evaluated over 3 years. Field Crops Res 97:254–260
Whitaker BD, Wang CY (1987) Effect of paclobutrazol and chilling on leaf membrane lipids in cucumber seedlings. Physiol Plant 70:404–411
White JW, Hoogenboom G (2003) Gene-based approaches to crop simulation: past experiences and future opportunities. Agron J 95:52–64
Yang Y, Lu Y, Zheng J et al (2023) Paclobutrazol induces the concurrent accumulation of chrysolaminarin and lipids in the diatom Phaeodactylum tricornutum. J Oceanol Limnol 1–12
Yao Y, You Q, Duan G et al (2020) Quantitative trait loci analysis of seed oil content and composition of wild and cultivated soybean. BMC Plant Biol 20:1–13
Yol E, Uzun B (2012) Geographical patterns of sesame accessions grown under Mediterranean environmental conditions, and establishment of a core collection. Crop Sci 52:2206–2214
Yol E, Ustun R, Golukcu M, Uzun B (2017) Oil content, oil yield and fatty acid profile of groundnut germplasm in Mediterranean climates. J Am Oil Chem Soc 94:804
Yol E, Basak M, Kızıl S et al (2021) A high-density SNP genetic map construction using ddRAD-Seq and mapping of capsule shattering trait in sesame. Front Plant Sci 12:679659
Zhang X, Ding L, Song A et al (2022) Dwarf and robust plant regulates plant height via modulating gibberellin biosynthesis in chrysanthemum. Plant Physiol 190:2484–2500
Zhu LH, Peppel A, Li XY, Welander M (2004) Changes of leaf water potential and endogenous cytokinins in young apple trees treated with or without paclobutrazol under drought conditions. Sci Hortic 99:133–141
Acknowledgements
We appreciate the Scientific Research Projects Coordination Unit of Akdeniz University for continuous support and are grateful to the USDA, ARS Plant Genetic Resources Conservation Unit, Griffin, GA, United States supplying genetic material several times.
Funding
Open access funding provided by the Scientific and Technological Research Council of Türkiye (TÜBİTAK).
Author information
Authors and Affiliations
Contributions
MQ performed the field studies and drafted the manuscript. EY conducted statistical analysis. MK, BU, and EY revised the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors have no relevant financial or nonfinancial interests to disclose.
Additional information
Communicated by Mohammad Irfan.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Qureshi, M., Kordrostami, M., Uzun, B. et al. Modulating Growth and Oil Profile of Sesame (Sesamum indicum L.): Paclobutrazol and Mepiquat Chloride Impacts. J Plant Growth Regul (2024). https://doi.org/10.1007/s00344-024-11317-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00344-024-11317-8