
Abstract
The abundance and distribution of demersal fishes rely on larvae successfully settling from the pelagic environment to a benthic habitat and their subsequent survival. With high mortality rates during this life stage, settling to a habitat that maximizes survival is critical. However, relationships between settlement choices and subsequent survival are poorly understood and may vary among species with different habitat preferences. To test this, we focused on five congeneric (Pomacentrus) damselfish species that are known to differ in their habitat choices and explored whether habitat associations at settlement influenced survival. Newly settled individuals were tagged and monitored daily for two weeks to estimate natural mortality rates. Morphological attributes of fish and characteristics of settlement habitats, including depth, rugosity, benthic substrata, and local fish assemblages, were used to predict mortality. We found that some species displayed stronger associations with specific benthic substrata at settlement, but contrary to expectations, these selected habitat characteristics were relatively weak predictors of survivorship. Our survival analysis revealed that the best predictors of survivorship were rugosity (P. adelus and P. amboinensis) and two morphological traits, body depth and ocellus size (P. chrysurus and P. adelus). Interestingly, we found that P. moluccensis that settled in areas of high coverage of mounding coral experienced increased mortality. Of the remaining substrata, analysis showed that instead of associating with habitat characteristics enhancing survivorship, individuals tended not to associate with habitats characteristics that increased mortality (e.g., Turbinaria and sand). This study highlights the species-specific drivers of early post-settlement mortality in coral reef fishes.
Similar content being viewed by others


Introduction
Settlement from the planktonic environment ends the larval stage for most marine fishes and establishes them into their juvenile habitats. Ontogenetic transitions to new habitats are a window of vulnerability, as newly settled juveniles are exposed to a wide range of predators for the first time (Caley et al. 1996; Holbrook and Schmitt 2003). This period is particularly important for coral reef fishes, where settlement is followed by high levels of juvenile mortality, creating a life-history bottleneck that can shape reef populations and communities (Doherty et al. 2004; Almany and Webster 2006). However, the outcomes of predator–prey interactions can depend on a myriad of factors including prey boldness and escape performance (Fuiman et al. 2010; McCormick et al. 2018), size and condition of larvae (Booth and Beretta 2004; Hoey and McCormick 2004), camouflage (Mihalitsis et al. 2024), competition for refuge (Holbrook and Schmitt 2002; Bonin et al. 2009), predator density (Webster 2002; Holbrook and Schmitt 2003) as well as environmental factors including the availability of live coral or preferred habitat (Coker et al. 2012; Majoris et al. 2018), rugosity and structure of coral (Beukers and Jones 1997; Fontoura et al. 2020; Fakan et al. 2023). The relative importance of these factors vary among fish species, depending on their habitat and species associations at settlement (Webster 2002). As settlement rates and early post-settlement survival are considered to be primary factors regulating population structures for coral reef fishes (Jones 1990; Doherty and Fowler 1994; Doherty et al. 2004), choices made by individuals at settlement can have population level consequences (Komyakova et al. 2019).
Coral reefs are biologically and structurally heterogeneous environments composed of a patchwork of microhabitats and habitat selection at settlement is well-established for reef fishes (Sale et al. 1984; Tolimieri 1995; Caley et al. 1996; Booth and Wellington 1998). With settlement occurring during the night, larval fishes primarily use audio cues to navigate and locate reefs (Kingsford et al. 2002) and olfactory cues of both local fishes and substrate for microhabitat selection (Lecchini et al. 2005; Coppock et al. 2020). This microhabitat selection process can differ markedly among species. For example, in cafeteria choice experiments, settlement-stage Pomacentrus moluccensis consistently selected to settle on live coral and habitats containing conspecifics (either juveniles or adults), while P. chrysurus non-selectively settled onto rubble, live or dead coral and avoided habitats with conspecifics (Öhman et al. 1998). Furthermore, naturally settling pomacentrids have been shown to avoid experimental patches emanating olfactory cues of resident predators, suggesting a strong and innate olfactory recognition of predators (Vail and McCormick 2011). In natural settings, however, the optimal habitats may not always be available with settlement patterns likely representing a compromise between locally available resources, selection, and avoidance preferences.
Selecting for a specific habitat or narrow scope of habitats is an ecological strategy (MacNally 1995) that provides these specialist species improved fitness or survivorship and therefore a competitive edge over more generalist species when using their preferred habitats (MacNally 1995; Caley and Munday 2003). For example, the neon goby (Elacatinus lori) ends their larval phase by selectively settling on the outer wall of sponges before recruiting into the sponge tube where they spend much of their adult life (D’Aloia et al. 2011; Majoris et al. 2018). While E. lori are known to settle on various sponges, they preferentially settle on yellow sponges (Aplysina fistularis) over brown sponges (Agelas conifera) and this selection preference has been shown to significantly increase their survivorship in their host sponge (Majoris et al. 2018). Conversely, habitat generalist species have the ability to adapt to a wide range of environmental conditions and resources; thus, they can thrive in diverse habitats (MacNally 1995). As habitat generalists, their associations with benthic characteristics at settlement should be broad and less focused (Wilson et al. 2008), and the factors potentially affecting their survivorship are likely to be more varied. Clearly, for species that have strong associations with particular habitat at settlement, the availability of the required habitat will be a major determinant of their subsequent distribution patterns (Holbrook et al. 2000), while this will not necessarily be the case for less specialized species. Settlement into suboptimal habitats can be detrimental to growth and physical condition (Feary et al. 2009), yet much less in known about how preferences during settlement may affect survivorship (Booth and Beretta 1994). Therefore, a better understanding of the environmental factors which promote survivorship is needed as these choices may strongly influence future population structures.
The capacity to perceive social contexts and subsequently make decisions about whether to settle within an existing assemblage of residents is crucial for both fitness and survival. For habitat specialists, olfactory cues of conspecifics can presumably indicate that the habitat is fitting (Lecchini et al. 2005); however for generalists, these cues may suggest higher density and/or competition and therefore may avoid such cues (Coppock et al. 2016). Joining aggregations of conspecifics may be costly, particularly for subordinates as adults aggression can influence settlement sites (Ben-Tzvi et al. 2009) and higher densities can increasing the likelihood of density-dependent mortality (Shima 2001). Therefore, deciding to settle with or avoid conspecifics may be critical determinate of survivorship.
For species showing strong settlement preferences for particular environmental characteristics (e.g., Holbrook et al. 2000; Bonin 2012; Pratchett et al. 2012; Coker et al. 2015), it is reasonable to expect that habitat selection would be beneficial. However, little is known about how habitat associations during settlement may affect the survivorship of reef fishes. To address this gap, we conducted an extensive field survey to determine whether the biological and physical factors of microhabitats used at settlement promote survival. Firstly, we quantified a suite of environmental and habitat factors of microhabitats on which we observed newly settled damselfish individuals, including rugosity, depth, microhabitat composition and local fish community. We also measured for each fish different morphological traits recognized as important drivers of survival, namely standard length, body depth, and ocellus size. We then followed individuals through time to estimate natural mortality rates. This allowed us to establish whether the microhabitats on which fish settled were also associated with differential survivorship. We selected five common congeneric damselfish species that are known to vary in habitat use (Öhman et al. 1998; Hata and Ceccarelli 2016; Pratchett et al. 2016): P. adelus, P. amboinensis, P. chrysurus, P. moluccensis and P. wardi. We hypothesize that some species would show stronger associations for particular habitat characteristics and for this selection to provide improved survivorship, while other species would have weaker habitat associations, and that the survivorship of these habitat generalists would be more influenced by non-habitat related traits.
Methodology
Study species
Five common site-attached congeneric damselfish species (Pomacentrus, Pomacentridae) that differ in their habitat use were selected for the study (Pratchett et al. 2016): (1) the lemon damsel, P. moluccensis, is an obligate coral-dwelling damselfish often found on colonies of branching coral; (2) the ambon damselfish, P. amboinensis, is a facultative coral-dwelling species associated with a variety of substrata including live and dead coral; (3) the obscure damselfish, P. adelus, often associated with dead coral and rubble and pavement; (4) the white-tailed damselfish, P. chrysurus, commonly inhabiting algal-covered rubble areas; (5) the ward’s damselfish, P. wardi, inhabiting the reef edge or reef top associated with pavement, rubble and soft coral.
Field protocol
Habitat associations at settlement and early post-settlement survivorship of the five species were quantified across two shallow sites on the leeward (i.e., western) aspect of Lizard Island (14.6680°S, 145.4638°E), a high continental island in the northern Great Barrier Reef (GBR). Each site was 15 × 25 m, and flagging tape was attached to the substrata at 5 m intervals to create fifteen 5 × 5 m cells. Using these cells, key benthic features were mapped. Starting seven days prior to the new moon in October and November 2021, which coincides with the period of peak settlement for reef fishes on the northern GBR (Meekan et al. 1993), each site was systematically searched daily (08:00–13:00) for any newly settled fishes of the five target species. Each newly settled individual was captured using a dilute clove oil solution and hand-nets, transferred to a small plastic bag where it was laterally photographed against grid paper (0.5 cm grid) and tagged with an individually identifiable fluorescent elastomer tag (Northwest Marine Industries Inc.) using a 27-gauge needle (see Fig S1). When found in groups or near previously tagged individuals, each fish was uniquely tagged. Elastomer tags have been shown to not impact growth or survivorship of newly settled pomacentrids (Hoey and McCormick 2006). Fish were left in the plastic bag at the exact site of capture for 10 min (to allow full recovery), after which they were released at the site of capture. All tagged fish were observed to begin feeding within 30 s of release. The site of capture was recorded on a site map, marked with a numbered tag and small piece of flagging tape attached to the substratum (see Fig S1). Morphological characteristics (i.e., standard length, body depth, and ocellus area) of each individual were subsequently measured using ImageJ from the lateral photographs of each fish. Standard length and body depth were highly correlated (R2 = 0.83), therefore body depth was used in all further analysis, as it is considered a major morphological constraint which may restrict prey consumption (Mihalitsis and Bellwood 2017).
While an individual fish was recovering from the tagging process, a 0.75 × 0.75 m quadrat centered on the exact site of capture was placed on the substratum and a photograph taken from a height of ~ 1 m above the substratum. The photo-quadrats were analyzed using photoQuad (Trygonis and Sini 2012), recording the substratum under 100 stratified random points within each quadrat. The substratum was categorized as pavement (smoothed reef structure), rubble, sand, macroalgae, hard coral, soft coral, or other (e.g., clams, sponge). Macroalgae were identified to genus and hard and soft corals identified to genus and later grouped into broad growth forms (i.e., coarse branching, fine branching, mounding and soft coral) for analysis. The rugosity of the substratum within the quadrat was estimated as the ratio of a chain (link size 1.5 cm) that followed the reef contour to the linear distance across the centre of the quadrat (0.75 m) and again perpendicular to the first measure (McCormick 1994); then, the mean of both measures were used. The time and water depth of each quadrat was recorded, and depth standardized using tide charts. To estimate baseline benthic composition and compare across species settlement association, an additional 70 control quadrats were haphazardly photographed, and benthic characteristics measured in the same manner (see Table S1). All non-cryptic fishes in the immediate vicinity (within a 1 m radius) of the quadrat were identified to species, their total length (TL) estimated to the nearest cm. Surveys were conducted at each quadrat after the tagging of the focal recruit and each survey was conducted only once. From these surveys, individual species and life stages were pooled to create fish community variables (i.e., potential predators, conspecific juveniles and adults, congeneric juveniles and adults, and other territorial damselfishes). The potential predator group consisted of the non-transient predators Pseudochromis fucus, Thalassoma lunare, T. hardwicke, T. nigrofasciatum, and Synodus dermatogenys. Conspecific juveniles were defined as being ≤ 2 cm total length (TL), while adults were ≥ 3 cm TL. Territorial adults consisted of other noticeably aggressive pomacentrid species from the genera Stegastes and Dischistodus. Together, these physical and biological microhabitat characteristics are known to correlate with distribution patterns of newly settled reef fish (McCormick and Hoey 2006).
To quantify survivorship, each site was thoroughly searched daily (15:00–17:00) for 14 days, and the presence or absence of tagged fish was recorded. When an individual appeared to be missing, a thorough search was conducted across an area of ~ 2 m radius from where the fish was initially tagged. If the tagged individual was not found over three consecutive days, it was classified as deceased.
Statistical analyses
Settlement associations
The differences among species in their association at settlement with (a) habitats characteristics or (b) fish community characteristics were explored separately using one-way permutational multivariate analyses of variance (PERMANOVA) based on Bray-Curtis distances (n = 10,000 permutations). Bonferroni corrected pairwise comparisons were conducted. Control quadrats were included with habitats characteristics (a), to investigate how specialized species are and for which habitat characteristics. However, control quadrats could not be included in fish community characteristics (b) due to the grouping of conspecific and congeneric species. The nature of differences found between species by each PERMANOVA were visually displayed using a canonical discriminant analysis (CDA) (Seber 1984). Species centroids and their 95% confidence intervals were plotted along the first two canonical axes, with habitat variables as vectors. The nature and strength of their association with each species are shown by their direction and length.
Survivorship
Differences in the survivorship among pomacentrid species were compared using a Cox’s proportional hazard regression analysis (Cox PH) and trends were displayed using a Kaplan–Meier plot. This semi-parametric model describes the relation between the event (mortality), as expressed by the hazard function and a set of covariates (Bradbury et al. 2003). The Cox PH model assumption of propotional hazards between species were visually explored using Schoenfeld residuals and were satisfied. Next, the impact of morphological, environmental and local communities on the survivorship of each species were explored separately by Cox PH regressions. Initially, models were fitted using a maximum of six covariates, using the dredge function in the MuMIn package (Barton 2009) in R (version 4.3.1). Then, model averaging of non-significantly different weighted models (wi ≤ 0.95%), were used to construct the most likely model including significant covariates (P = 0.1). Again, the Cox PH model assumptions of proportional hazards for all significant variables within each species were explored and satisfied using Schoenfeld residuals. For P. adelus, ocellus size exhibited strong but highly variable influence on the model and was therefore included as a categorical predictor (< or ≥ median ocellus size) in the final model. Hazard ratios of the variables selected from the most likely model were visually explored using forest plots, which display the mean hazard ratio (HR) and 95% confidence intervals (CI). HR’s above 1 indicates the covariate is positively associated with the probability of mortality occuring. The effect size, determined by 1-HR (see Bradbury et al. 2003), is denoted as odds ratios (OR). The OR can be interpreted as a percentage of their association with affecting the risk of mortality. For example, a HR of 0.16 is associated with reducing the risk of mortality by 84% (1-HR).
Results
Settlement associations
There was a significant difference in habitat associations among the five damselfish species at settlement (PERMANOVA; F5,384 = 20.45, p < 0.001), with P. moluccensis being associated with more rugose microhabitats and a higher cover of fine-branching corals, while P. amboinensis was associated with deeper and sandier sites and P. chrysurus being associated with a higher cover of rubble (Fig. 1a). In contrast, P. adelus and P. wardi exhibited more generalized associations with rubble, pavement and the macroalga Turbinaria and had a higher degree of spread of habitat use. Pairwise comparisons showed that all species significantly differed (p < 0.05) from each other and controls except, P. adelus and control quadrats and also P. adelus and P. wardi (see Table S2).

Differences among five newly settled pomacentrid species in their association at settlement with a habitat characteristics and b fish community characteristics. Biplots of canonical discriminant analyses show the first two canonical axes with loading scores. Crosses represent species centroids, circles surrounding crosses represent 95% confidence intervals for species. Vectors represent the direction and strength of the original variables to discriminate among species. Pomacentrus adelus (blue), P. amboinensis (purple), P. chrysurus (green), P. moluccensis (red), and P. wardi (orange) and control quadrats (black)
The five damselfish species also differed in their associations with fish community composition (PERMANOVA; F4,315 = 7.62, P < 0.001; Fig. 1b). Pairwise comparisons between all 5 species showed that most species associated with different fish assemblages at settlement (p < 0.05); the only exception being P. amboinensis, that was not significantly different to P. moluccensis or P. chrysurus (Table S3). Relatively low R2 values for most pairwise comparisons of fish community characteristics suggests overlaps in associations of fish community variables, which is likely a response due to the coarse classifications of fish community traits (See Table S3). P. adelus associated with congeneric juveniles and P. wardi with congeneric adults and other territorial adults. P. chrysurus was found to associate with conspecific adults and predators. Both P. moluccensis and P. amboinensis tended to have a higher association with conspecific adults and juveniles than the other species; however, these associations were generally more variable, as indicated by the wider spread of habitat data.
Survivorship
The survival trajectories did not differ among the five pomacentrid species, with 33–52% of individuals remaining after 14 days post-settlement (χ24 = 6.87, P = 0.143; Fig. 2). Overall survival curves were steepest in the first four days, with 75.4% (P. wardi) to 58.7% (P. amboinensis) of individuals remaining after four days.

Survival curves of five pomacentrid species for 14 days post-settlement at Lizard Island, northern Great Barrier Reef. Red: Pomacentrus moluccensis (n = 67); purple: P. amboinensis (n = 63); blue: P. adelus (n = 61); green: P. chrysurus (n = 72); orange: P. wardi (n = 57)
Influence of environmental associations on survivorship
P. moluccensis
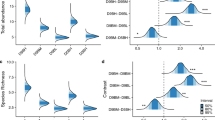
The Cox PH regression analysis for P. moluccensis shows the presence and abundance of congeneric juveniles and the cover of mounding corals and sand significantly improved the fit of the model (χ23 = 13.48, P = 0.004; Figs. 3a, 4). The presence and abundance of local congeneric juveniles (≤ 2 cm) were related with a reduced risk of mortality in P. moluccensis (HR = 0.69, P = 0.008) by 31% (i.e., 1-HR). In contrast, the cover of mounding coral was significantly positively associated with increased mortality risk (HR = 1.03, P = 0.040). Sand tended to increase the mortality risk but was found to be non-significant (HR = 1.04, P = 0.06).

Forest plots of time-dependent Cox proportional hazard regression analysis for Pomacentrus moluccensis, P. amboinensis, P. adelus, P. chrysurus (a-d). Each cube in the forest plot represents the hazard ratio surrounded by its 95% confidence interval and hazard ratios above 1 indicate that the covariate is positively associated with the probability of mortality occuring. Effect size calculated as 1-HR, is denoted (OR). Note: none of the variables considered was related to the survivorship of Pomacentrus wardi

Logistic regressions showing relationships of habitat, fish community and morphological variables selected by the Cox proportional hazard regression models that were found to influence the survivorship of four pomacentrid species: Pomacentrus moluccensis, P. amboinensis, P. adelus, and P. chrysurus. Note: significance notation derived from the Cox proportional hazard regression models and none of the variables considered was related to the survivorship of Pomacentrus wardi
P. amboinensis
The addition of rugosity, congeneric adults and Turbinaria coverage significantly improved the final model (χ23 = 12.57, P = 0.005) for P. amboinensis (Figs. 3b, 4). Higher rugosity significantly reduced risk of mortality by 85% (HR = 0.15, P = 0.010). The abundance of local congeneric adults and Turbinaria were both associated with increasing the probability of mortality by 24 and 27%, respectively (HR = 1.24, P = 0.010; HR = 1.27, P = 0.040).
P. adelus
Rugosity, territorial adults and ocellus size (< median) significantly improve the survivorship model (χ23 = 14.64, P = 0.002) for P. adelus (Figs. 3c, 4). Rugosity was negatively associated with mortality (HR = 0.16, P = 0.016) and improved the model fit by 84%. Having an ocellus smaller than the median (0.025 mm2) significantly increased the risk of mortality (HR = 2.19, P = 0.029) by 119% compared to individuals with an ocellus greater than median (Fig. 4). Territorial adults tended to increase the mortality risk but was found to be non-significant (HR = 1.43, P = 0.15).
P. chrysurus
Only body depth and predator abundance improved the survival model for P. chrysurus (χ22 = 13.26, P = 0.0013; Fig. 3d). Having a shallower body depth was found to significantly increase mortality risk (HR = 0.0013, P = 0.003) by 99%. Increases in the abundance of local predators tended to increase mortality for P. chrysurus; however, this was found to be non-significant (HR = 1.16, P = 0.076) (Fig. 4).
P. wardi
For P. wardi, no variables were found to significantly improve the survival model. The relative importance of the measured variable’s ability to explain the time-dependent mortality rate of P. wardi was considerably lower than the other Pomacentrus species examined. The relative importance of the best predictor (congeneric adults) was ~ 40%, less than half of other species’ best predictors (See Supplementary Fig. S2).
Discussion
Overview
Identifying the drivers of survival following settlement is critical to understand and predict population structures for coral reef fishes (Booth and Beretta 1994, 2021; Doherty and Fowler 1994). Here, we provide a detailed assessment the factors that affect early post-settlement survival of five congeneric coral reef damselfishes. Our study confirms that the drivers of mortality in juvenile coral reef fishes are species specific and include a range of habitat, fish community and morphological characteristics. Contrary to expectation, we found only a weak relationship between the habitat associations at settlement and the factors that influenced subsequent survival. For example, P. moluccensis associated with areas of increased rugosity and cover of fine branching coral at settlement, P. amboinensis associated with deeper sites with higher cover of sand, and P. chrysurus associated with areas of higher rubble cover, yet none of these characteristics was important in predicting the survivorship in the respective species over the following 14 days. Interestingly, most of the habitat characteristics which were identified as important for survivorship were characteristics which showed a negative association at settlement. For example, relative to other species P. amboinensis had a negative association with the macroalga Turbinaria at settlement and those individuals which settled into areas with higher cover of Turbinaria had increased mortality. Similarly, P. moluccensis had a negative association with sand at settlement, and increasing levels of sand tended to increase mortality, albeit non-significantly. The one exception was that P. moluccensis associated with mounding corals at settlement, which was found to increase their risk of mortality over the next 14 days.
Our hypothesis that survivorship in generalist species would be broadly impacted by a range of habitat, fish communities and morphological characteristics was generally supported. Survivorship models for P. adelus selected three variables with each belonging to a habitat characteristic, fish community feature or morphological trait. While no variables were found to significantly improve the survival model for P. wardi, the six most important traits included three habitat characteristics, two fish community traits and one morphological (See Fig S2), broadly supporting that survivorship would be impacted by various traits. The lack of determination and clarity in this model may indicate that survival for this generalist species was particularly context dependent.
Habitat associations and their implications on survivorship
Coral reefs are heterogeneous environments composed of various microhabitat characteristics that may determine which species can thrive in that microhabitat. The current study illustrates clear differences in settlement associations among the five pomacentrid species examined, with three species (P. moluccensis, P. amboinensis, and P. chrysurus) associating for particular habitat characteristics, while P. adelus and P. wardi were more variable in their habitat associations (i.e., generalists). Our results of habitat associations align with previous works suggesting that P. moluccensis is associated with various types of hard coral at settlement (Ticzon et al. 2012; Pratchett et al. 2016) and P. chrysurus are predominantly found on rubble habitat (Low 1971). In contrast, P. amboinensis is considered a facultative coral specialist (Pratchett et al. 2012); however, its association with various types of hard coral were similar to the average coverage of the corals found in the control quadrats (see Table S1), suggesting little to no association with hard corals. While P. amboinensis was observed to associate with live coral, our data suggest that settling at the base of the reef in areas with higher cover of sand may be the primary habitat consideration for this species. Consistent with this hypothesis, Ticzon et al. (2012) found that juvenile P. amboinensis predominately associated with sand, using it more than any other species they investigated.
As mortality is a strong selective pressure on recently settled reef fish, it is expected that selection for a particular microhabitat would confer some fitness advantage (Tolimieri 1995; Majoris et al. 2018). However, our results do not necessarily support this hypothesis, as most analyses did not identify the habitat characteristics chosen at settlement as important drivers of mortality. High rugosity or relatively high cover of fine branching coral were positively associated with settlement of P. moluccensis, however, neither were identified as important predictors of survivorship. In an earlier study using experimental mesocosms, the survivorship of P. moluccensis has been shown to increase when sheltering in coral with more rugosity (Beukers and Jones 1997). It may be that the relationship between rugosity and survival follows a logistic-like curve, such that having a certain level of rugosity provides a benefit, but beyond a specific threshold, this benefit reaches a plateau. In the current study, P. moluccensis may already be selecting for rugosity beyond the point of receiving a benefit, while P. amboinensis and P. adelus appear to be settling in the middle of their curve, such that those with above average rugosity benefited while those with below average rugosity suffer increased mortality. Conversely, P. chrysurus selectively settled in relatively lower rugosity microhabitats which was found not to relate to their survivorship, suggesting the importance of rugosity or level required is likely species specific, depending on the species ecological needs. As in some cases, increased rugosity may not always benefit prey (Rilov et al. 2007; Fakan et al. 2023).
Though the overall rugosity was not important for P. moluccensis, it may be that the finer structural components of coral are important. For example, we found that the cover of mounding corals in the immediate vicinity of newly settled P. moluccensis was positively related to the probability of mortality. The lack of holes and reduced sheltering capacity of mounding coral (Aston et al. 2022) likely increases the exposure of small-bodied and newly settled fish to predation (Almany 2004). These areas with high cover of mounding corals did not necessarily translate to areas of lower rugosity, and may be related to the scale and nature of the rugosity provided (e.g., fine-scale structure within the branches of branching corals versus the vertical relief provided by mounding corals). While these likely underestimated the true rugosity, our results suggest that the finer scale refuge is important for survival. Consistent with this conclusion, Hixon and Beets (1993) found that Chromis cyanea had improved early post-settlement survival on experimental reef patches with small holes compared to large holes or no holes.
The habitat preferences of coral reef fish vary, and due to the high heterogeneity of reefs, some individuals inevitably settle in suboptimal habitats. Our data reveal that when assessing the relative influence of microhabitat characteristics on survival, insights from negatively associated habitat characteristics outweigh those from associated habitat characteristics. As fish traverse their home ranges, they are likely to use these suboptimal habitats. Thus, it is essential to consider all habitat characteristics surrounding their settlement site at a scale relevant to their home ranges. While data on the home ranges and space use of coral reef fishes, especially recently recruited juveniles, are limited, some studies suggest that adult P. moluccensis have home ranges of < 2m2 (Streit et al. 2021), with juveniles expected to have even smaller ranges (Welsh et al. 2013). Anecdotal evidence by Booth and Beretta (1994) reports observing nine uniquely recognizable recruits which were never found more than 30 cm away from their initial location during a 30-day census period. This suggesting that home ranges of recruits may be relatively small and comparable to the quadrats used in the current study. Although when examining habitat use, previous studies have primarily focused on single point substratum assessments (e.g., McCormick and Hoey 2004; Wilson et al. 2008; Bonin 2012; Pratchett et al. 2012) for exploring habitat selection and habitat selectivity measures (see Manly et al. 2002). However, these methods may miss broader habitat characteristics that influence fitness traits. For instance, P. amboinensis showed negative associations with the macroalga Turbinaria, significantly impacting survivorship, while P. moluccensis displayed a negative association with sand, which tended to increase their risk of mortality. Therefore, assessing the relative abundance of microhabitat characteristics and home ranges the fish interact with, rather than focusing solely on single substratum types, may lead to a more comprehensive understanding of the environmental factors influencing various fitness traits.
Habitat associations during the early life stage of coral reef fish may offer fitness benefits beyond their impact on early post-settlement survival. In the current study, species that associated with particular habitat characteristics received no survival benefits from such characteristics. However, they likely derive benefits in terms of faster growth from increased access to their preferred food sources. For instance, P. moluccensis is commonly found residing in corals high above the reef framework, positioning them close to plankton, which constitutes their primary diet (Pratchett et al. 2001). Similarly, unconsolidated substrata (e.g., sand and rubble) can foster higher densities of invertebrates (Wen et al. 2013), which serve as a vital nutrient source for P. amboinensis (Debenay et al. 2011). Additionally, P. chrysurus is known to defend rubble-dominated territories, offering them access to the turf matrix that forms a significant part of their diet (Low 1971). The increased availability of these food resources may enhance growth rates and potentially reduce predation risk, thereby contributing to overall fitness improvement.
Fish associations and their implications on survivorship
Grouping with conspecifics is an important trait to predict fitness outcomes in reef fishes (Booth 1995; McCormick and Meekan 2007). The current study highlights that species associations with characteristics of the fish community at settlement were indeed important, even though its importance was weaker than their associations with habitat characteristics. Our results show that P. adelus, P. chrysurus and P. wardi tended to settle in areas with higher abundances of congeneric species, while both P. amboinensis and P. moluccensis tended to settle in areas with higher abundances of conspecifics. These results align with previous works detailing grouping behaviors in both P. amboinensis and P. moluccensis (Öhman et al. 1998).
The formation of groups may be an ecological trade-off between survival and growth or may be due to preferential settlement. Grouping can have fitness impacts and is known to enhance survivorship in recently settled reef fishes in some circumstances (Booth 1995; McCormick and Meekan 2007). Conversely, grouping can negatively impact the growth rates for recently settled reef fish (Jones 1990; Booth 1995). For example, Booth (1995) showed that the time to reach maturity increased with group size, suggesting that living in smaller groups or living solitarily would allow an individual to more swiftly enter into the adult population. In the current study, various associations with fish communities were found to impact the survivorship of each species. Congeneric adults significantly increased the risk of mortality in recently settled P. amboinensis. Also, the abundance of adult territorial damselfishes (i.e., Stegastes and Dischistodus) similarly reduced the survivorship of P. adelus, another territorial or farming damselfish. Although newly settled farming damselfish are unlikely to be directly competing for the same resources, adult damselfish may see them as future competitors. Alternatively, settling near other damselfish species has been shown to enhance early post-settlement survival (McCormick and Hoey 2004) and may be an attempt to reduce conspecific group size, which can reduce growth rates (Booth 1995). Interestingly, we found a positive relationship between congeneric juveniles and the survivorship of P. moluccensis. The additional prey items nearby may be reducing the predation pressure on P. moluccensis causing increased survivorship (Ferrari et al. 2020). Additionally, as densities increase, the optimal habitats will become saturated forcing newer recruits to suboptimal habitats. Our data suggest that early mortality rates are impacted by resident populations and probably in a density-dependent manner.
Although predators are known to be important regulators of coral reef fish communities (Webster 2002; Feeney et al. 2012; Chivers et al. 2019), the abundance of local predators (Pseudochromis, Thalassoma and Synodus) was only associated with the survival rates of one of the five species, P. chrysurus (albeit non-significantly). This suggests that the mortality rates are not related to the predator densities recorded using visual surveys at the scale of the current study (Connell 1996). This could be a result of local predators maintaining a larger home range than 1 m perimeter surrounding the quadrat, resulting in underestimates of local predator and their activity from abundance. Additionally, a primary source of the mortality may be related to transient predators and/or cryptic predators (Hoey and McCormick 2004; Mihalitsis et al. 2022) that may not be adequately quantified using visual surveys of specific sites. Also, due to the timing of the surveys (08:00–13:00), crepuscular or nocturnal fishes which may represent important predators may have been similarly underrepresented. While various components of the fish community impacted the survivorship of the studied species based on the species, it is clear that local communities do influence the survivorship at this life stage.
Morphological implications on survivorship
While habitat and fish communities were found to be important predictors of survival, morphological traits of individuals were found to be particularly informative. Although only two morphological variables (ocellus size and body depth) were used in the final survival analysis (of the total of 18 variables), these variables scored in the top 6 variables for relative importance for survival in three of the five species (Fig. S2), and for two of the five species they were the best predictors of survivorship. Interestingly, the morphological traits tended to be more important for species which are known territorial algae farmers (Hata and Ceccarelli 2016). Our data suggest that in microhabitats with more aggressive residents, either size or dominance marking of larger ocellus may are providing survival benefits.
Although ocelli are hypothesized to confer some survival benefit (McPhail 1977), few studies have explicitly demonstrated such benefits. We found the relative ocellus size to be the primary predictor of survival in P. adelus. The responsiveness of relative ocellus size to predator cues found in a recent study (McCormick et al. 2019), also emphasizes their likely importance to survival. Interestingly, the ocellus was not found to be important predictors of survival in P. amboinensis or P. chrysurus. Indeed, while ocelli can provide certain advantages to some species in terms of predator deterrence and prey confusion, they may not be universally favored in all environments. Planktivores (e.g., P. moluccensis), for instance, are rarely found to possess ocelli (Hemingson et al. 2021), indicating that the benefits of ocelli in reducing predation risk may not apply universally across all ecological niches. Thus, ocelli may not serve as a broad and universally effective strategy for countering predation risks in all species and environments.
Body depth was also found to be a strong predictor in survival for P. chrysurus. Body depth is an important morphological trait which can enhance prey’s escape responses (Domenici et al. 2008) and restrict predation by gape-limited predators (Mihalitsis and Bellwood 2017). Together, our results support the long standing belief that bigger is better for coral reef fishes recruits (Vigliola and Meekan 2002), as larger individuals of P. chrysurus had higher survival rates. In contrast to its importance, we only found size to predict survivorship of one species. This disparity may stem from the influence of local predators on P. chrysurus (although non-significant), which are smaller than transient predators and, consequently, more limited by gape size.
In summary, our study revealed contrasting habitat selection behaviors among different fish species during settlement, with some displaying specific preferences for certain microhabitat characteristics, while others exhibited a more generalized approach. Furthermore, we identified several environmental and morphological traits associated with the survival of recently settled fishes. Intriguingly, species that associated with particular microhabitats showed higher survival rates when they were not associated with specific habitat characteristics. While we initially anticipated that microhabitat characteristics promoting settlement would also enhance survival, the predictive power of such associations was generally weak within the scope of our investigation. Nonetheless, selecting specific habitat characteristics during settlement may yield longer-term fitness benefits, such as improved growth and faster maturation. This study underscores the diverse ways in which various habitat characteristics can influence populations.
References
Almany GR (2004) Does increased habitat complexity reduce predation and competition in coral reef fish assemblages? Oikos 106:275–284
Almany GR, Webster MS (2006) The predation gauntlet: early post-settlement mortality in reef. Fishes 25:19–22
Aston EA, Duce S, Hoey AS, Ferrari R (2022) A protocol for extracting structural metrics from 3D reconstructions of corals. Front Mar Sci 9:1–14
Barton K (2009) MuMIn : multi-model inference. R package version 1. 0. 0. http://r-forge.r-project.org/projects/mumin/
Ben-Tzvi O, Kiflawi M, Polak O, Abelson A (2009) The effect of adult aggression on habitat selection by settlers of two coral-dwelling damselfishes. PLoS ONE 4:e5511
Beukers JS, Jones GP (1997) Habitat complexity modifies the impact of piscivores on a coral reef fish population. Oecologia 114:50–59
Bonin MC (2012) Specializing on vulnerable habitat: Acropora selectivity among damselfish recruits and the risk of bleaching-induced habitat loss. Coral Reefs 31:287–297
Bonin MC, Srinivasan M, Almany GR, Jones GP (2009) Interactive effects of interspecific competition and microhabitat on early post-settlement survival in a coral reef fish. Coral Reefs 28:265–274
Booth DJ (1995) Juvenile groups in a coral-reef damselfish: density-dependent effects on individual fitness and population demography. Ecology 76:91–106
Booth DJ, Beretta GA (1994) Seasonal recruitment, habitat associations and survival of pomacentrid reef fish in the US Virgin Islands. Coral Reefs 13:81–89
Booth DJ, Beretta GA (2004) Influence of recruit condition on food competition and predation risk in a coral reef fish. Oecologia 140:289–294
Booth DJ, Beretta GA (2021) Long-term demographics of a coral-reef fish: growth, survival and abundance at several spatial scales. Coral Reefs 40:1257–1266
Booth DJ, Wellington G (1998) Settlement preferences in coral-reef fishes: effects on patterns of adult and juvenile distributions, individual fitness and population structure. Austral Ecol 23:274–279
Bradbury MJ, Clark TG, Love SB, Altman DG (2003) Survival analysis part II: multivariate data analysis—an introduction to concepts and methods. Br J Cancer 89:431–436
Caley JM, Munday PL (2003) Growth trades off with habitat specialization. Proc R Soc B Biol Sci 270:175–177
Caley JM, Carr MH, Hixon MA, Hughes TP, Jones GP, Menge BA (1996) Recruitment and the local dynamics of open marine populations. Annu Rev Ecol Syst 27:477–500
Chivers DP, McCormick MI, Fakan EP, Barry RP, Edmiston JW, Ferrari MCO (2019) Coral degradation alters predator odour signatures and influences prey learning and survival. Proc R Soc B Biol Sci 286:20190562
Coker DJ, Graham NAJ, Pratchett MS (2012) Interactive effects of live coral and structural complexity on the recruitment of reef fishes. Coral Reefs 31:919–927
Coker DJ, Hoey AS, Wilson SK, Depczynski M, Graham NAJ, Hobbs JPA, Holmes TH, Pratchett MS (2015) Habitat selectivity and reliance on live corals for Indo-Pacific hawkfishes (family: Cirrhitidae). PLoS ONE 10:1–17
Connell SD (1996) Variations in mortality of a coral reef fish: Links with predator abundance. Mar Biol 126:347–352
Coppock AG, Gardiner NM, Jones GP (2016) Sniffing out the competition? Juvenile coral reef damselfishes use chemical cues to distinguish the presence of conspecific and heterospecific aggregations. Behav Process 125:43–50
Coppock AG, González-Murcia SO, Srinivasan M, Gardiner NM, Jones GP (2020) Different responses of coral and rubble-dwelling coral reef damselfishes (Family: Pomacentridae) to chemosensory cues from coral reef microhabitats. Mar Biol 167:1–11
D’Aloia CC, Majoris JE, Buston PM (2011) Predictors of the distribution and abundance of a tube sponge and its resident goby. Coral Reefs 30:777–786
Debenay JP, Sigura A, Lou JJ (2011) Foraminifera in the diet of coral reef fish from the lagoon of New Caledonia: Predation, digestion, dispersion. Rev Micropaleontol 54:87–103
Doherty PJ, Fowler A (1994) Demographic consequences of variable recruitment to coral reef fish populations: a congeneric comparison of two damselfishes. Bull Mar Sci 54:297–313
Doherty PJ, Dufour V, Galzin R, Hixon MA, Meekan MG, Planes S (2004) High mortality during settlement is a population bottleneck for a tropical surgeonfish. Ecology 85:2422–2428
Domenici P, Turesson H, Brodersen J, Brönmark C (2008) Predator-induced morphology enhances escape locomotion in crucian carp. Proc R Soc B Biol Sci 275:195–201
Fakan EP, Allan BJM, Illing B, Hoey AS, McCormick MI (2023) Habitat complexity and predator odours impact on the stress response and antipredation behaviour in coral reef fish. PLoS ONE 18:e0286570
Feary DA, McCormick MI, Jones GP (2009) Growth of reef fishes in response to live coral cover. J Exp Mar Bio Ecol 373:45–49
Feeney WE, Lönnstedt OM, Bosiger Y, Martin J, Jones GP, Rowe RJ, McCormick MI (2012) High rate of prey consumption in a small predatory fish on coral reefs. Coral Reefs 31:909–918
Ferrari MCO, McCormick MI, Fakan EP, Barry RP, Chivers DP (2020) The fading of fear effects due to coral degradation is modulated by community composition. Funct Ecol 34:2120–2130
Fontoura L, Zawada KJA, D’agata S, Álvarez-Noriega M, Baird AH, Boutros N, Dornelas M, Luiz OJ, Madin J, Maina JM, Pizarro O, Torres-Pulliza D, Woods RM, Madin EMP (2020) Climate-driven shift in coral morphological structure predicts decline of juvenile reef fishes. Glob Chang Biol 26:557–567
Fuiman LA, Meekan MG, McCormick MI (2010) Maladaptive behavior reinforces a recruitment bottleneck in newly settled fishes. Oecologia 164:99–108
Hata H, Ceccarelli DM (2016) Farming behaviour of territorial damselfishes. Biol Damselfishes 160:122–152
Hemingson CR, Siqueira AC, Cowman PF, Bellwood DR (2021) Drivers of eyespot evolution in coral reef fishes. Evolution (n y) 75:903–914
Hixon MA, Beets JP (1993) Predation, prey refuges, and the structure of coral-reef fish assemblages. Ecol Monogr 63:77–101
Hoey AS, McCormick MI (2004) Selective predation for low body condition at the larval-juvenile transition of a coral reef fish. Oecologia 139:23–29
Hoey AS, McCormick MI (2006) Effects of subcutaneous fluore scent tags on the growth and survival of a newly settled coral reef fish, Pomacentrus amboinensis (Pomacentridae). In: Proceedinas 10th International coral reef symposium, pp 420–424
Holbrook SJ, Forrester GE, Schmitt RJ (2000) Spatial patterns in abundance of a damselfish reflect availability of suitable habitat. Oecologia 122:109–120
Holbrook SJ, Schmitt RJ (2002) Competition for shelter space causes density-dependent predation mortality in damselfishes. Ecology 83:2855–2868
Holbrook SJ, Schmitt RJ (2003) Spatial and temporal variation in mortality of newly settled damselfish: patterns, causes and co-variation with settlement. Oecologia 135:532–541
Jones GP (1990) The importance of recruitment to the dynamics of a coral reef fish population. Ecology 71:1691–1698
Kingsford MJ, Leis JM, Shanks A, Lindeman KC, Morgan SG, Pineda J (2002) Sensory environments. Larval Abilit Local Self-Recruit 70:309–340
Komyakova V, Munday PL, Jones GP (2019) Comparative analysis of habitat use and ontogenetic habitat-shifts among coral reef damselfishes. Environ Biol Fishes 102:1201–1218
Lecchini D, Shima J, Banaigs B, Galzin R (2005) Larval sensory abilities and mechanisms of habitat selection of a coral reef fish during settlement. Oecologia 143:326–334
Low RM (1971) Interspecific territoriality in a pomacentrid reef fish, pomacentrus flavicauda whitley. Ecology 52:648–654
MacNally RC (1995) Ecological versatility and community ecology. Cambridge University Press, Cambridge
Majoris JE, D’Aloia CC, Francis RK, Buston PM (2018) Differential persistence favors habitat preferences that determine the distribution of a reef fish. Behav Ecol 29:429–439
Manly BFJ, McDonald LL, Thomas DL, McDonald TL, Erickson WP (2002) Resource selection by animals. Kluwer Academic Publishers, New York
McCormick MI (1994) Comparison of field methods for measuring surface topography and their associations with a tropical reef fish assemblage. Mar Ecol Prog Ser 112:87–96
McCormick MI, Hoey AS (2004) Larval growth history determines juvenile growth and survival in a tropical marine fish. Oikos 106:225–242
McCormick MI, Hoey AS (2006) Biological and physical correlates of settlement and survival for a coral reef fish, Pomacentrus amboinensis (Pomacentridae). In: Proceedings of the 10th internation coral reef symposium, pp 425–430
McCormick MI, Meekan MG (2007) Social facilitation of selective mortality. Ecology 88:1562–1570
McCormick MI, Fakan EP, Allan BJM (2018) Behavioural measures determine survivorship within the hierarchy of whole-organism phenotypic traits. Funct Ecol 32:958–969
McCormick MI, Fakan EP, Palaciosotero M (2019) Habitat degradation and predators have independent trait-mediated effects on prey. Sci Rep 9:1–11
McPhail JD (1977) A possible function of the caudal spot in characid fishes. Can J Zool 55:1063–1066
Meekan MG, Milicich MJ, Doherty PJ (1993) Larval production drives temporal patterns of larval supply and recruitment of a coral reef damselfish. Mar Ecol Prog Ser 93:217–225
Mihalitsis M, Bellwood DR (2017) A morphological and functional basis for maximum prey size in piscivorous fishes. PLoS ONE 12:1–19
Mihalitsis M, Bellwood DR, Wainwright PC (2024) Sit and survive: predation avoidance by cryptobenthic coral reef fishes. Mar Biol 171:1–7
Mihalitsis M, Morais RA, Bellwood DR (2022) Small predators dominate fish predation in coral reef communities. PLoS Biol 20:1–14
Öhman M, Munday PL, Jones GP, Caley JM (1998) Settlement strategies and distribution patterns of coral-reef. Fishes 225:219–238
Pratchett MS, Gust N, Goby G, Klanten SO (2001) Consumption of coral propagules represents a significant trophic link between corals and reef fish. Coral Reefs 20:13–17
Pratchett MS, Coker DJ, Jones GP, Munday PL (2012) Specialization in habitat use by coral reef damselfishes and their susceptibility to habitat loss. Ecol Evol 2:2168–2180
Pratchett MS, Hoey AS, Wilson SK, Hobbs JPA, Allen GR (2016) Habitat-use and Specialisation among coral reef damselfishes. Taylor & Francis, New York
Rilov G, Figueira WF, Lyman SJ, Crowder LB (2007) Complex habitats may not always benefit prey: linking visual field with reef fish behavior and distribution. Mar Ecol Prog Ser 329:225–238
Sale PF, Douglas WA, Doherty PJ (1984) Choice of microhabitats by coral reef fishes at settlement. Coral Reefs 3:91–99
Seber GAF (1984) Multivariate observations. Wiley, New York
Shima J (2001) Recruitment of a coral reef fish: Roles of settlement, habitat, and postsettlement losses. Ecology 82:2190–2199
Streit RP, Hemingson CR, Cumming GS, Bellwood DR (2021) How flexible are habitat specialists? Short-term space use in obligate coral-dwelling damselfishes. Rev Fish Biol Fish 31:381–398
Ticzon VS, Mumby PJ, Samaniego BR, Bejarano S, David LT (2012) Microhabitat use of juvenile coral reef fish in Palau. Environ Biol Fishes 95:355–370
Tolimieri N (1995) Effects of microhabitat characteristics on the settlement and recruitment of a coral reef fish at two spatial scales. Oecologia 102:52–63
Trygonis V, Sini M (2012) PhotoQuad: a dedicated seabed image processing software, and a comparative error analysis of four photoquadrat methods. J Exp Mar Bio Ecol 424–425:99–108
Vail AL, McCormick MI (2011) Metamorphosing reef fishes avoid predator scent when choosing a home. Biol Lett 7:921–924
Vigliola L, Meekan MG (2002) Size at hatching and planktonic growth determine post-settlement survivorship of a coral reef fish. Oecologia 131:89–93
Webster MS (2002) Role of predators in the early post-settlement demography of coral-reef fishes. Oecologia 131:52–60
Welsh JQ, Goatley CHR, Bellwood DR (2013) The ontogeny of home ranges: evidence from coral reef fishes. Proc R Soc B Biol Sci 280:1–7
Wen CKC, Pratchett MS, Almany GR, Jones GP (2013) Patterns of recruitment and microhabitat associations for three predatory coral reef fishes on the southern Great Barrier Reef, Australia. Coral Reefs 32:389–398
Wilson SK, Burgess SC, Cheal AJ, Emslie MJ, Fisher R, Miller IR, Polunin NVC, Sweatman HPA (2008) Habitat utilization by coral reef fish: implications for specialists vs. generalists in a changing environment. J Anim Ecol 77:220–228
Acknowledgements
We are beholden to Rhondda Jones for her statistical assistance and knowledge. We are grateful to Maddi Craig, John McCormick, Bella Marrable, Erika Gress and Mila Grinblat who aided in the field and Alexia Dubuc for providing feedback on the manuscript. We also thank the directors and staff of Lizard Island Research Station (a facility of the Australian Museum) for their continuous logistical support.
Funding
Open Access funding enabled and organized by CAUL and its Member Institutions. Funding was provided by the Australian Research Council Centre of Excellence for Coral Reef Studies (ASH), the Lizard Island Reef Research Foundation (Gough Fellowship; EPF), and the Australian Museum’s Lizard Island Research Station (EPF).
Author information
Authors and Affiliations
Contributions
EPF, MIM and ASH conceived the ideas and designed methodology; EPF collected the data, analyzed the data and wrote the initial draft of the manuscript; all co-authors substantially revised the manuscript for submission.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Ethical approval
All research was carried out under approval of the James Cook University animal ethics committee (permit: A2683) and according to the University’s animal ethics guidelines.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Fakan, E.P., McCormick, M.I., Jones, G.P. et al. Habitat and morphological characteristics affect juvenile mortality in five coral reef damselfishes. Coral Reefs 43, 171–183 (2024). https://doi.org/10.1007/s00338-023-02462-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00338-023-02462-9