
Abstract
Continuous sedimentary lake records covering the Last Glacial Maximum (LGM) are very rare in Europe. Here, we conducted additional palynological analyses to connect two separate radiocarbon-dated pollen sequences of Lago della Costa (AP2 and AP1) to one complete vegetation history record of the Euganean Hills (“Colli Euganei”, northern Italy) covering the past ca. 32,000 years. Previous studies on autochthonous organic lake sediments revealed that during the Last Glacial Maximum (LGM) Lago della Costa likely acted as a refugium of temperate woody taxa. Based on the new data we present the first continuous record of the vegetation history of the Euganean Hills during the past 32,000 years. Specifically, we investigate the early establishment and mass expansion of deciduous Quercus at the refugial site at ca. 16,800–16,600 cal bp. In the oak forests other temperate taxa (e.g. Corylus avellana, Abies alba, Ulmus, Fraxinus excelsior, Vitis, Hedera, Tilia, Acer, Fagus sylvatica) also established at around this time, i.e. 2,000 years prior to the onset of the Bølling/Allerød interstadial (about 14,700–12,800 cal bp). The comparison of our data with other well-dated records from the river Po catchment and refugial sites elsewhere in Italy and southern Europe suggests expansions of boreal and nemoral forests over wide areas prior to the Bølling/Allerød interstadial, i.e. 2,000 years before comparable large-scale afforestation north of the Alps or ca. 45°N. Pollen-inferred expansions of temperate tree communities (e.g. oak forests) correspond to a climatic warming as reconstructed from quantitative, pollen-independent evidence (e.g. chironomids) from the Apennines, Northern Italy, Southern Switzerland and the Swiss Plateau, suggesting that pre-Bølling afforestation processes south of the Alps were promoted by increasing (summer) temperatures during the Oldest Dryas.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
In the last years, a growing body of palaeoecological, genetic, and climate modelling literature, suggested a more northerly glacial survival for temperate tree species in Europe than solely in southern Europe (Beug 1975; Fineschi et al. 2002; Willis and van Andel 2004; Birks and Willis 2008; Birks 2019; Guido et al. 2020). Although this issue is controversially debated (Tzedakis et al. 2013) this means that populations may have been closer to their present range-limits than previously estimated, potentially reducing spreading rates across the continent (Feurdean et al. 2013). Conversely, there is abundant palaeoecological evidence that temperate trees survived the LGM in refugia south of the Alps or ca. 45°N (Lang et al. 2023).
The southern slope of the Alps was extensively glaciated during the LGM (Ehlers and Gibbard 2004). Conifer and several temperate tree species survived nearby in the ice-free Po Plain and along the southeastern alpine border, as testified by the biostratigraphic evidence (e.g. Ravazzi et al. 2004; Monegato et al. 2015). Pollen evidence of a glacial macrorefugium for temperate trees on the eastern Po Plain is preserved in the autochthonous lake sediment records from Lago della Costa at the southeastern border of the Euganean Hills (Kaltenrieder et al. 2009). This site is located south of another important site on the Po Plain, Lago di Fimon in the Berici Hills, with a vegetation history spanning back to the LGM and beyond (Pini et al. 2010). Fimon was, however, located much closer to the LGM ice masses (25 vs. 55 km) and thus more exposed to climatic and environmental extremes. A long sequence spanning the last glacial cycle and the late Middle Pleistocene is also available from the lowermost Friulian Plain (Monegato et al. 2007; Fontana et al. 2008; Pini et al. 2009) ca. 120 km north-east of Lago della Costa. Lago di Fimon, similar to the landscape reconstructed in the Friulian Plain, was affected by fluvioglacial aggradation during the LGM. At these more northern and likely cooler sites, conifer trees and shrubs, and, with a low probability, very restricted, mesic and more cold-tolerant Fagus sylvatica and Abies alba persisted during the LGM (e.g. Pini et al. 2009).
Quantitative vegetation-independent palaeotemperature reconstructions show that LGM mean July air temperatures were 10–12 °C lower than today in the south-eastern sector of the Po Plain (Samartin et al. 2016). The chironomid-inferred temperature reconstructions depict that in the plains at the southern edge of the Euganean Hills (Lago della Costa) mean July temperatures were on an average as high as 12–16 °C during 31,000–19,000 cal bp, while in the Euganean Hills they were likely 2–4 °C warmer at elevated sites, i.e. at ca. 14–20 °C (Gubler et al. 2018). The coldest LGM mean July temperatures were around 10–13 °C (Samartin et al. 2016) on the Po Plain and 12–17 °C in the Euganean Hills (Gubler et al. 2018). This particular microclimatic setting in the Euganean Hills likely permitted temperate tree taxa to survive the harshest LGM conditions (Kühl and Moschen 2012; Samartin et al. 2016).
While unglaciated areas in the river Po basin had partly remained forested during the LGM (Ravazzi et al. 2004, 2006; Kaltenrieder et al. 2009; Monegato et al. 2015; Marcolla et al. 2021), in the southern Alps and their forelands Juniperus scrublands (pollen, stomata finds) established ca. 18,000–17,500 cal bp in the formerly glaciated areas (Vescovi et al. 2007). Subsequently, synchronous early afforestation processes occurred over wide areas in the lowlands of southern Switzerland and northern Italy at about 16,500–16,000 cal bp, 2,000–1,500 years prior to the large-scale afforestation of central Europe in response to the Bølling warming at ca. 14,700–14,600 cal bp (Hofstetter et al. 2006; Vescovi et al. 2007; Ravazzi et al. 2014).
Comparison of the pollen-inferred palaeovegetational reconstructions with independent palaeoclimatic series (chironomid records) suggests that early pre-Bølling afforestation processes in the formerly glaciated areas of northern Italy and neighbouring areas were promoted by increasing temperatures, i.e. early summer temperature warming at ca. 16,500–16,000 cal bp; Samartin et al. 2012; Bolland et al. 2020). North of the Alps the pre-Bølling warming likely released only minor expansion of tree Betula stands (Bolland et al. 2020; Rey et al. 2020).
In summary, several studies with independent lines of evidence showed that the Euganean Hills supported a unique LGM refugium in the hills of the Adriatic Po Plain (Lona 1957; Paganelli and Miola 1991; Kaltenrieder et al. 2009, 2010; Samartin et al. 2016; Gubler et al. 2018). Consequently, this area is particularly interesting for understanding the beginning of European postglacial re-afforestation dynamics. In the low-resolution overview study of land core AP2, the early expansion of mixed oak stands is recorded only in the three uppermost pollen samples. In this study, we focus on the period 18,000–14,000 cal bp, which, due to its distribution on different land and lake cores, was so far interrupted or uncovered. In order to connect the land core AP2 to the lake core AP1 and thus establish a continuous record of environmental change for the past 32,000 years, we carried out additional high-resolution pollen analyses in the uppermost sediment section of the land core AP2. This procedure allows us to reconstruct the re-expansion of important temperate tree taxa, specifically the early expansion of deciduous Quercus and the establishment of stands of other temperate taxa such as Abies alba, Corylus avellana, Alnus glutinosa, Fraxinus excelsior, Carpinus betulus, Tilia, Ulmus, Fagus sylvatica, Vitis, Hedera and Acer at ca. 16,800–16,600 cal bp. Furthermore, we refined the pollen determinations, now distinguishing Pinus cembra-type from Pinus sylvestris-type in all pollen samples from core AP1/2 to determine the abundance and quantitative relationship of both Pinus taxa. Our new results are compared with proximal records from south and north of the Alps that were ice-covered during the LGM as well with more distant refugial sites in southern Europe to assess the spatial extent and consistency of the palaeovegetational and palaeoclimatic patterns observed.
Materials and methods
Study site
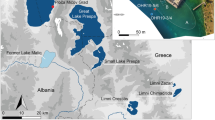
The study site, Lago della Costa (7 m a.s.l.; 45°16’13”N, 11°44’35”E), is a small lake (3 ha) in the regional park ‘Parco Regionale dei Colli Euganei’ close to Arquà Petrarca at the southeastern border of the Euganean Hills (Fig. 1). Due to the progressive removal of mud from the lake for therapeutic and cosmetic purposes, it was challenging to find an undisturbed coring spot in the lake. In collaboration with the mining company (Società Fango Lago Costa d’Arqua s.r.l.), who had access to the excavation history and a wealth of historical bathymetry maps (e.g. Stegagno 1901; Corpo reale delle miniere, Padova 1930), we were able to find a drilling area in the lake that was never used and thus included undisturbed organic lake sediments. The older sediments used for the period prior to ca. 16,700 cal bp were drilled on land. Further details on the coring campaigns in 2001 are available in Kaltenrieder et al. (2009, 2010).

Maps (A, B) showing the location of the study site in northeastern Italy; A: overview of the region around the Euganean Hills; B: Euganean Hills with the study site, Lago della Costa; C: photograph with Lago della Costa and Monte Ricco (to the south)
The lake has no surface inlet but several subaqueous thermal springs, originating from ancient rainwater of the Alps (Small Dolomites, Pasubio, Recoaro and the Vicentino-Trentini plateaus; Piccoli et al. 1976, 1981). The thermal water is related to a geothermal gradient from the Alps to the Euganean Hills, unrelated to any local underground heat sources (Cantaluppi et al. 2014). Radiometric studies on rocks attributed the Euganean eruptive complex an Oligocene age (35 million years) in its most recent terms. The volcanic activity is therefore far too old to be a source of heat or any other volcanic feature such as old carbon delivery to the air (Borsi et al. 1969; Piccoli et al. 1976; Soter 2011; Cantaluppi et al. 2014). We can thus exclude any influence of carbon degasification on our radiocarbon age measurements. In addition to these subaqueous thermal springs, Lago della Costa was until modern times fed by freshwater input from precipitation, creeks and springs, which are now used for agricultural purposes (REGPARK 2015). At the southern border of the lake, there is a small artificial outlet.
The temperature gradients today are broad enough to permit both, warm-loving evergreen Mediterranean (e.g. Q. ilex woodlands) and mesophilous communities (e.g. F. sylvatica stands) to occur in the Euganean Hills, whereas both are absent from the adjacent low-lying Po Plain today, likely because of climatically extreme and disturbed local conditions (e.g. frost, drought). Consequently, this hilly area is representing a main centre of past and recent floristic biodiversity (Pignatti 1982). For further details on the environmental conditions of Lago della Costa site (geology, pedology, recent climate and vegetation) see Kaltenrieder et al. (2009, 2010), Samartin et al. (2016) and Gubler et al. (2018).
Lithology, pollen analysis and zonation
Full material and methodological descriptions of coring, lithology (including loss on ignition), pollen, macrofossils, ostracods, chironomids, microscopic and macroscopic charcoal analyses are provided in Kaltenrieder et al. (2009, 2010) and Samartin et al. (2016). For this study, 12 additional pollen samples were taken at the surface of the AP2 sediment land core to better resolve the time of the postglacial re-establishment of deciduous oak forests. Oak pollen was differentiated between the two pollen types Quercus deciduous (Q. pubescens, Q. petraea, Q. robur) and Q. ilex-type containing evergreen Quercus species. In addition, in all 219 pollen samples of AP1/2, we conducted new determinations of P. cembra-type and P. sylvestris-type, based on the presence of verrucae on the ventral surface of the grain body, on the shape of sacci (more elongated than and not as rounded as in P. sylvestris-type) and on the presence of a slightly undulating outer layer on the proximal surface of the grain body towards the sacci (Klaus 1972).
The pollen diagrams were drawn using TILIA 2.0.4 and TiliaGraph 2.0.4. The results are presented as TgView 2.0.2 pollen diagrams (Grimm 1992–2005) subdivided into local pollen assemblage zones (LPAZ) by using the zonation method of optimal partitioning with sum-of-squares (Birks and Gordon 1985), identifying the statistically significant LPAZ limits with the broken-stick approach (Bennett 1996).
Merging the sediment cores on the basis of their chronology
Lago della Costa consists of two parallel cores which have been published in two separate articles: a sediment core retrieved in the present lake (AP1; Kaltenrieder et al. 2010) spanning back to the Late-glacial and a land core taken close to the today’s shore (AP2; Kaltenrieder et al. 2009) covering the early Late-glacial, LGM and beyond to ca. 32,000 cal bp.
Only terrestrial plant macrofossils were used for dating (Table 1). The 14C ages were converted to calibrated ages (years cal bp) using the program CALIB 8.2 (Stuiver and Reimer 1993) with the IntCal20 calibration curve (Reimer et al. 2020). Considering the uncertainties of the radiocarbon dates there is an overlap (and thus no gap) between the two cores AP1 and AP2. Specifically, the oldest 14C age from the lake core AP1 (14,270 ± 70 uncal bp), 6 cm above the sediment bottom, provides an estimated basal sediment age of ca. 17,340 cal bp, while the youngest age from the land core AP2 (14,086 ± 100 uncal bp) at 125 cm below sediment surface provides an age of ca. 17,130 cal bp (Table 1). This implies an age overlap between AP1 and AP2 of ca. 200 years according to the calibrated age medians. Therefore, two new age-depth models were calculated for AP1 and AP2 with the program clam using a smooth spline (Blaauw 2010). According to the median of the interpolated values, the depth of 1,279 cm at AP1 chronologically corresponded to a depth of 52 cm in the sediment core AP2. Consequently, the two pollen sequences were merged at the sediment depth of 1,279 cm in AP1 and at the depth of 52 cm in AP2 to build the new complete core AP1/2. This procedure resulted in one chronology on one continuous composite depth, which was constructed by adding 1,227 cm to the original sample depths of AP2 (starting at 52 cm, i.e. a new depth of 1,279 cm). Above 52 cm, topmost material of AP2 was partly removed before coring because we found traces of disturbance and reworking (ploughland). Hence, an original depth of AP2 corresponding to e.g. 90 cm has now a new artificial depth of 1,317 cm. After eliminating the chronological overlap and assigning the new depths to AP2, we built a common age-depth model for the composite cores AP1/2 (Fig. 2). After exploring linear, spline, GAM and Bayesian age-depth modelling, we decided to apply clam using a smooth spline (Blaauw 2010; Blaauw and Heegaard 2012). Moreover, a 95% probability distribution envelope was added using the GAM model by Heegaard et al. (2005). The age-depth model is based on linear interpolation of the R-Stat values of calibrated 14C ages (Heegaard et al. 2005).

Age-depth model for composite Lago della Costa (AP1/2). Black dots represent calibrated ages of plant macrofossils with 2σ error bars (IntCal20, Reimer et al. 2020). Connecting black line shows the modelled chronology (linear interpolation between smoothed (spline) values; Clam 2.2, Blaauw 2010). Blue envelope represents the 95% confidence interval (GAM, Heegaard et al. 2005)
The chronology of the new combined pollen record (total of 15 radiocarbon dates; Table 1) is spanning the last ca. 32,000 cal bp. In the new age-depth model (Fig. 2) and in the composite pollen diagram (Fig. 3) the original date at 14,270 ± 70 uncal bp, which anchors the basal age of AP1 at 1,290–1,298 cm depth, is not shown anymore, because belonging to an AP1 depth that was replaced by AP2 depths. The biostratigraphy of the sites was not considered directly when merging the cores, however, the chronological integration is in good agreement with the pollen patterns in AP1 and AP2 (ESM). For instance the Late-glacial Quercus mass expansion occurs at the top of the AP2 core (ca. 80–60 cm in original AP2 depth, now ca. 1,307–1,287 cm in new artificial depth), but is already accomplished at the bottom of the AP1 core (1,280–1,300 cm).

Pollen percentage (selected types) and microscopic charcoal influx (particles/cm2/yr) diagram of combined AP1/2. Empty curves show 10x exaggerations
Results and interpretation
The new composite pollen diagram is subdivided into 10 statistically significant local pollen assemblage zones (LPAZ), PAP12-1 to PAP12-10 (Fig. 3). The new zonation emphasizes the main traits of the vegetation history and landscape development and shows a highly dynamic transitional phase at 16,900–16,000 cal bp (Fig. 4). A detailed description of the vegetation history and dynamics at Lago della Costa during the last 32,000 years has been presented in Kaltenrieder et al. (2009, 2010). For the results, we focus on this transition for which new samples were analysed, for the vegetation history of the other periods during the past 32,000 years we refer to the original publications (Kaltenrieder et al. 2009, 2010).

Pollen percentage (selected types) diagram of combined AP1/2 for the high-resolution section of the early Late-glacial period. Empty curves show 10x exaggerations
Vegetation conditions after termination I and vegetation change at 16,900–16,000 cal bp
After the LGM, at Termination I (PAP12-5 and PAP12-6; 18,500–17,300 cal bp), minerogenic sediment deposition increased at the site, perhaps as a consequence of increased water discharge which caused increasing soil erosion. Subsequently, during PAP12-7 (17,300–16,700 cal bp) autochthonous sedimentation re-established (silty-marl to marl, silty and calcareous gyttja) leading to high pollen quality and concentrations. Our pollen data suggest the presence of isolated tree stands (Larix decidua, P. sylvestris-type, Quercus deciduous, C. betulus, F. sylvatica, Ulmus) in the hills, surrounded by steppic environments (e.g. relatively high pollen percentages of Artemisia, Chenopodiaceae, Poaceae and Juniperus) of the plain. The microscopic charcoal record shows that fire activity during the early Late-glacial was low.
The additional pollen samples taken from AP2 depict a gradual expansion of temperate trees at 16,900–16,600 cal bp (Fig. 4). Around 16,900 cal bp Corylus avellana, Quercus deciduous, Ulmus, F. sylvatica, A. glutinosa-type, and C. betulus reached their empirical limit (i.e. continuous curves), suggesting the establishment of first local populations at the lake (Lang 1994; Birks and Tinner 2016). At ca. 16,800 cal bp Abies and at 16,700 cal bp Vitis followed. At the same time pollen of Quercus, C. avellana, A. glutinosa-type, F. excelsior-type, Tilia and Ulmus peaked, suggesting first considerable expansions of these temperate trees in the Lago della Costa area. Insect-pollinated Hedera and Acer show regular pollen occurrences after ca. 16,650 cal bp, pointing to the establishment of these arboreal plants in the close proximity of the lake. Similarly, evergreen broadleaved Q. ilex-type reached its absolute pollen limit (first occurrence) at ca. 16,650 cal bp. Interestingly, during this period also boreal taxa such as Larix decidua and Pinus cembra expanded, pointing to the establishment of nemoboreal forests within ca. 200 years. Forests expanded at the expenses of steppic vegetation, as shown by the decline of Artemisia and Chenopodiaceae (Fig. 4). However, high NAP values show that open land, primarily grasslands (Poaceae) remained important. By 16,600–16,000 cal bp temperate trees became dominant in the forests and boreal taxa as P. sylvestris-type, P. cembra-type and Betula declined. The zonation shows that the major vegetation break occurred around 16,700 cal bp (significant zone boundary PAP12-7 to PAP12-8). After ca. 16,700 cal bp Late-glacial and early to Mid Holocene vegetation remained rather stable for millennia (Figs. 3 and 4). Microscopic charcoal influx shows that the expansion of temperate trees was connected to an increase of fire activity, perhaps in response to warmer conditions and/or increasing biomass and thus fuel availability.
Discussion
Vegetation and climatic situation at the end of the last glaciation
During the last glaciation including the Last Glacial Maximum (LGM), the Euganean Hills and surrounding Po Plain, as well as the Adriatic alluvial Plain were unglaciated (e.g. Monegato et al. 2007, 2015). The glacio-eustatic sea lowstand along the Italian coast set the whole northern Adriatic shelf under continental conditions. The alluvial plain of the river Po extended to about 300 km south of the present Venetian coastal area (e.g. Lambeck et al. 2002). During the LGM aggradation phase, glaciofluvial sediments were deposited and extremely harsh climatic conditions prevailed in the Plain, which strongly affected vegetation. Pollen sequences indicate that at that time, the eastern Po Plain and the enlarged Adriatic alluvial Plain were situated in a cold, dry environment with a dominance of steppes with Poaceae, Artemisia and Chenopodiaceae, accompanied by spots of parklands with cold-adapted boreal elements such as P. cembra, P. sylvestris, L. decidua, Juniperus, Picea abies and tree Betula (Bortolami et al. 1977; Lowe et al. 1996; Combourieu-Nebout et al. 1998; Ravazzi et al. 2004; Monegato et al. 2007; Marcolla et al. 2021). As tree Betula taxa do not tolerate prolonged summer drought and drying cold winds (Atkinson 1992), its occurrence points to more moderate habitat conditions, perhaps in topographically moist depressions. Although the LGM was generally characterized by very dry conditions, precipitation patterns over the study region were variable. A moisture gradient may have occurred regionally, with orographic rainout triggered by southern air circulation (Pini et al. 2010; Luetscher et al. 2015), increasing moisture availability from the Adriatic alluvial Plain to the mountain fringes. Such moisture variability may explain the presence of a forest belt of boreal trees extended over the southern and southeastern alpine forelands during the LGM where moisture availability was sufficient (Pini et al. 2010; Ravazzi et al. 2014; Monegato et al. 2015). However, the very cold and dry conditions excluded temperate broad-leaved trees with relatively high summer temperature requirements, even where moisture availability was sufficient to let boreal forests grow in the Po Plain.
A rare presence of temperate (e.g. deciduous Quercus) and mesophilous (e.g. Fagus) taxa is depicted in the pollen records of organic layers of two new long sedimentary successions of continental and shallow marine deposits from the adjacent Venetian plain, less than 20 km away from Lago della Costa (Marcolla et al. 2021), characterized by the abundance of taxa of cold and dry climate (e.g. Pinus). The authors explain this presence of pollen of temperate trees by transport from the near refugial site Euganean Hills, where our data suggest the survival of isolated temperate tree stands (Fig. 3, Kaltenrieder et al. 2009, 2010; Samartin et al. 2016; Gubler et al. 2018).
During the LGM, valley glaciers of the Alps remained only 30 km away from the Berici Hills (Castiglioni 1940), a relict karstic plateau ca. 20 km north of the Euganean Hills. In contrast to the Euganean Hills their slopes are everywhere quite gentle and straight, with a maximum elevation of 444 m a.s.l. The basin of Lake Fimon is the most important valley dissecting the plateau to the north, open to southward winds from the Alps and thus fully exposed to cold air outbursts from the former ice masses. These geographical and geomorphological conditions did not allow the growth of temperate broadleaved trees with high summer temperature requirements during the LGM. Indeed, in the sedimentary record of Lake Fimon, pollen of temperate trees is almost absent since 30,000 cal bp. Only scattered stands with coniferous Pinus-Larix and some Picea occurred during the LGM period (Wick 1996; Pini et al. 2009, 2010).
Environmental and climatic conditions south of the Alps between 19,000 and 17,000 cal bp
Unambiguous evidence exists that the collapse of the ice sheet in the lowlands south and north of the Alps occurred within 1,000–1,500 years, starting not later than 19,300 cal bp, at the end of the LGM (Rey et al. 2020). Fluvial incision of melt water led to the formation of valleys in the Po Plain, which were up to 14 m deep and 0.5–2 km wide (Mozzi et al. 2010, 2013; Fontana et al. 2014). Moreover, glacier melting at the foothills of the southern and southeastern Alps led to increased surface water discharge and slope instability inducing the aggradation of the rivers Adige, Brenta, Piave and Tagliamento in the eastern Po Plain (Fontana et al. 2008). High sedimentation rates prior to 18,000 cal bp are also documented in pre-alpine lake sediments (e.g. at Lago di Origlio, Tinner et al. 1999; Vescovi et al. 2007) that became ice-free at the end of the LGM around 19,000 cal bp (Rey et al. 2020). The huge loss of ice masses and the fast retreat of glaciers were likely controlled by increasing summer insolation (Berger and Loutre 1991) as well as constantly rising CO2 and CH4 concentrations in the atmosphere (Lourantou et al. 2010). Between 19,000 and 17,000 cal bp, increasing temperatures are registered at many study sites worldwide according to the rising northern hemisphere insolation values (He et al. 2013).
South of the Alps and close to the LGM refugia in the eastern Po Plain, scrublands of heliophilous Juniperus spread into the steppic tundra on formerly glaciated areas above 400 m a.s.l. and stabilized the soils, promoting the subsequent afforestation processes (Tinner et al. 1999; Vescovi et al. 2007). For instance, high Juniperus pollen values and stomata suggest the presence of juniper stands at 18,000–17,000 cal bp at Lago di Origlio at 416 m a.s.l., ca. 230 km to the west of our study site (Figs. 5 and 6). The early expansion of Juniperus scrublands or woodlands was most probably the result of climate warming (+ 2.5–3 °C) at the onset of the Late-glacial around ca. 18,800 cal bp as recorded in chironomid-inferred mean July air temperature reconstructions (Samartin et al. 2016; Finsinger et al. 2019).

Comparison of percentages of tree pollen and arboreal pollen showing the afforestation at different study sites south and north of the Alps with an independent climate proxy: Lago della Costa (blue, this study), Annone (green, Wick 1996), Origlio (red, Tinner et al. 1999), Balladrum (purple, Hofstetter et al. 2006), Burgäschisee (brown, Rey et al. 2017); reconstructed July air temperatures at sea level based on chironomid data from Lago della Costa (magenta, Samartin et al. 2016), Lago di Origlio (light-blue, Samartin et al. 2012), and from the northern Apennines, Italy, green and orange: Lago Gemini and Lago Verdarolo (Samartin et al. 2017)

Comparison of selected pollen taxa from different study sites south and north of the Alps with an independent climate proxy. Selected pollen data from Lago della Costa (this study), Annone (Wick 1996), Origlio (Tinner et al. 1999), Balladrum (Hofstetter et al. 2006), Burgäschisee (Rey et al. 2017). Pollen diagrams show percentages of Pinus cembra-type (purple), Juniperus (brown), Betula (blue), B. nana (cyan; only at site Burgäschisee); deciduous Quercus (red), Corylus avellana (green); for reconstructed July air temperatures see Fig. 5
Early expansion of mixed oak forests at the Euganean Hills and contemporaneous re-afforestation processes south of the Alps
After early vegetation responses to Termination I warming, major vegetation reorganizations occurred around 17,000–16,000 cal bp. Our new pollen data (Fig. 4) show a moderate expansion of forest patches with cold-adapted and continental P. cembra and L. decidua at around 16,900 cal bp. It was followed by the initial expansion of populations of deciduous Quercus and other temperate trees at ca. 16,800 cal bp. Already at around 16,700 cal bp, Quercus deciduous had reached conspicuous values of 15% and C. avellana of 6%. The vegetation shift ended at ca. 16,600–16,000 cal bp, when temperate trees became dominant in the forests and boreal taxa such as P. sylvestris, P. cembra and Betula declined. Pollen and macrofossil records show that at around 16,500–16,000 cal bp several tree species were spreading at other sites (Vescovi et al. 2007; Ravazzi et al. 2014). Pollen evidence from the Brianza area (Lago di Annone, 226 m a.s.l., Wick 1996) north-west of Lago della Costa suggests the establishment of open forests with Betula and P. sylvestris with some P. cembra. In contrast, in the Swiss Ticino area ca. 30 km to the west of the Italian Brianza area, radiocarbon dated pollen and macrofossil records suggest that P. cembra expanded massively at about 400–500 m a.s.l. at ca. 16,500–16,000 cal bp (Tinner et al. 1999; Hofstetter et al. 2006; Vescovi et al. 2007). Apparently, P. cembra preferred the siliceous soils of the Ticino area. Tree Betula taxa were also part of the afforestation at the referred sites at 16,500–16,000 cal bp in the northern Po Plain basin including the southern pre-Alps (Figs. 4 and 5). During this first afforestation phase with P. cembra, L. decidua, Betula, P. sylvestris, J. communis and P. abies prior to the Bølling, treeline in the region was ca. at 800 m a.s.l. (Tinner and Vescovi 2007; Lang et al. 2023; Fig. 7). If the chronological uncertainties of several hundred years are considered (Fig. 2), the afforestation dynamics observed at Lago della Costa at 16,800–16,600 cal bp do not differ significantly in time from those observed at the other sites south of the Alps at 16,500–16,000 cal bp. Indeed according to the GAM-model the 2σ uncertainty (for both depth and age uncertainties) around 16,800–16,600 cal bp reaches ca. ±700 years.

Schematic illustration showing the development of vegetation belts south of the Alps, in northern Italy and southern Switzerland. During the Last Glacial Maximum (LGM), fragmented forests and scattered tree stands existed in the lowlands, dominated by boreal species, with rare occurrences of temperate species in sheltered habitats as the Euganean Hills. Forests expanded during the Late-glacial in response to warming, the treeline reached ca. 800 m a.s.l. at 16,000 cal bp and ca. 1,800 m at 14,000–13,000 cal bp. At low elevations, steppic communities and parklands persisted in the Po Plain, possibly because of dry conditions and river disturbance. Further warming at the onset of the Holocene allowed a further expansion of forests, which reached modern treeline elevations above 2,000 m a.s.l. Following Beug (1975), Tinner and Vescovi (2007), Kaltenrieder et al. (2009, 2010) and Lang et al. (2023)
Contemporaneous early afforestation processes in the formerly unglaciated and glaciated areas of northern Italy and southern Switzerland (Fig. 7) were likely triggered by increasing (summer) temperatures (Samartin et al. 2016). For comparison with Lago della Costa, chironomid-inferred temperature reconstructions from Lago di Origlio were corrected to the altitude of Lago della Costa (7 m a.s.l.) assuming an altitudinal lapse rate of 0.6 °C/100 m (Livingstone and Lotter 1998). At ~ 17,500–17,000 cal bp, both chironomid-inferred reconstructions from Lago di Origlio and Lago della Costa show mean July temperature estimates around 12.5 °C (Fig. 5), suggesting rather cool conditions at sea level. At Lago di Origlio mean July temperatures were around 10 °C, locating the site close to treeline. Subsequently, at ca. 16,500–16,000 cal bp, summer temperatures became ca. 2 °C warmer south of the Alps (Samartin et al. 2012). During this period, mean July temperatures may have increased to ca. 12 °C at Lago di Origlio and 14.5 °C around Lago della Costa, explaining the different afforestation processes with dense boreal P. cembra forests establishing at Lago di Origlio and open mixed temperate Quercus forests around Lago della Costa. Besides warmer (summer) conditions, the expansion of forest stands was facilitated by increasing rainfall, as suggested by oxygen isotope values from speleothems (Frisia et al. 2005).
The well-dated Monticchio record in southern Italy (Watts et al. 1996; Allen et al. 2000, 2002) provides the opportunity to check if a similar oak forest expansion occurred at other Italian refugial sites under a comparable climatic setting. Today the two lakes Costa and Monticchio experience similar vegetation (spanning from mesomediterranean Quercus ilex to oromediterranean F. sylvatica in the catchment) and climatic conditions (ca. 21–23 °C mean July, 2–3 °C mean January and 700–900 mm annual precipitation; Watts et al. 1996; Tinner et al. 2013; Gubler et al. 2018). Lago della Costa is at the upper temperature range and thus slightly warmer, while Monticchio is at the lower end and thus cooler. Because this climatic similarity results from the altitudinal and latitudinal positions of the two sites, we expect that it was also present in the past. The pollen data suggest that at Monticchio, at ca. 500–1,000 m a.s.l., deciduous Quercus gradually displaced pine stands and open steppe tundra after ca. 16,500–16,000 cal bp (Allen et al. 2000, 2002). The establishment of first considerable oak stands (1–5% Quercus pollen) occurred at ca. 16,000–15,000 cal bp, while oak forests (> 10–20%) established only after ca. 14,500–14,000 cal bp. Fagus sylvatica also established stands at 16,000–15,000 cal bp at Monticchio, but in contrast to deciduous Quercus it did not spread at 14,500–14,000 cal bp, as was the case at Lago della Costa. These similar vegetational dynamics at the two sites may have resulted from two warming pulses, a minor at around 16,500 and a major at 14,700 cal bp (Samartin et al. 2012). Cooler climatic conditions at Monticchio may explain why the deciduous Quercus peak was delayed, if compared to Lago della Costa.
Similar Late-glacial forest dynamics occurred in the today’s mesomediterranean belt in central Italy. The palynological study of the lacustrine sediments from Lagaccione, a maar-lake in central Italy, has provided a palaeoenvironmental record for the last 100,000 years (Magri 1999). The pollen data show that at Lagaccione, at 355 m a.s.l., deciduous Quercus (together with Betula) started to increase to 2–5% at ca. 16,700–16,200 cal bp (13,615 ± 200 years bp, conv. uncal), marking the establishment of first deciduous oak stands. Oak forests (> 45%) established only after ca. 14,000–13,800 cal bp. Similarly to Monticchio, F. sylvatica also established first stands (more or less continuous pollen curve), but did not spread afterwards. In conclusion, as in Monticchio, it is difficult to determine precisely the early oak establishment and expansion in Lagaccione, because the temporal resolution of the pollen diagrams is much coarser than at Lago della Costa.
Further to the east, refugial sites in southeastern, peri-adriatic Europe such as Ioannina (Lawson et al. 2004), Prespa (Panagiotopoulos et al. 2013), and Tenaghi Philippon (Wijmstra 1969) also experienced early spreads of boreal and/or temperate tree taxa ca. 2,000 years prior to the Bølling/Allerød interstadial, which confirms that the early Late-glacial temperate tree stand dynamics reconstructed at our site in the Euganean Hills are not exceptional, neither temporally, nor spatially.
Contemporaneous pre-Bølling climate warming and vegetation shifts north of the Alps
Post-LGM warming at 16,500–16,000 cal bp also had impacts on vegetation dynamics north of the Alps (Rey et al. 2017, 2020; Bolland et al. 2020). For instance at Burgäschisee and Moossee, two lakes on the Swiss Plateau, steppic herbaceous tundra shifted to shrub tundra with first tree birch stands around 16,100 cal bp (Rey et al. 2017, 2020; Bolland et al. 2020 and Fig. 6). The latter vegetational re-organization which chronologically approximately corresponds to the mixed oak forest expansion at Lago della Costa, is well documented at several lowland sites north of the Alps (e.g. Welten 1982; Lotter 1999; Rey et al. 2020). Recent chironomid-inferred July temperature estimates for Burgäschisee (Bolland et al. 2020) confirm the inferences from the Po Plain basin (Samartin et al. 2012) that climate warming around 16,500–16,000 cal bp may have triggered early vegetation reorganizations. However, north of the Alps the warming around 16,000 cal bp was less pronounced and reached about 1 °C to values of ca. 10 °C, thus locating the lowlands of the Swiss Plateau at treeline (Bolland et al. 2020). Taken together, these results from the lowlands north of the Alps suggest an earlier Late-glacial temperature increase in southwest central Europe than expected, although with a less pronounced warming than the onset of the Bølling-Allerød interstadial. Warming at ca. 16,500–16,000 cal bp promoted early boreal and temperate afforestation processes in the formerly glaciated lowlands south of the Alps, while north of the Alps the shift promoted a moderate expansion of isolated treeline Betula stands.
Factors for the fast vegetation responses after climate warming in the early Late-glacial and perspectives for the future
Vegetation responses to early Late-glacial warming occurred under conditions of low competition and negligible human impact, with no apparent inertia, suggesting dynamic equilibrium with climate (Birks and Ammann 2000; Tinner and Kaltenrieder 2005). No major lags of population establishments imply a very high sensitivity and adjustment capacity of plant communities to climatic and environmental changes at centennial time scales. In the case of Lago della Costa no spatial spreading of species was needed, given that the site acted as a refugium of temperate species during the harsh ice age conditions. However, the records from the formerly glaciated southern pre-Alps and Swiss Plateau show that vegetation was able to respond rapidly also in areas that were unvegetated during the Ice Age. Rapid vegetation responses imply high rates of spread, which can be partly explained by the very efficient distribution mechanisms of plants (winged fruits transported by wind or palatable seeds transported by birds over very long distances (Firbas 1949; Tinner and Lotter 2006).
Facilitation might have been another important factor for the rapid establishment of tree taxa into new areas after the last ice age. Most plant species form mutualistic associations with mycorrhizal fungi in a symbiosis established immediately after seed germination. Mycorrhiza play a critical role in the capture of nutrients from the soil and therefore in plant nutrition. Several studies with oak and birch (Shaw 1974; Miles and Kinnaird 1979) suggest that especially on nutrient-poor sites mycorrhizal infection seems to be important for seedling survival of these tree species. Correia et al. (2019) provide the first evidence that forest birds can co-disperse viable seeds and viable mycorrhizal fungal diaspores, postulating a mechanism for the establishment of mycorrhizal associations after seed dispersal and for the colonisation of new sites by mycorrhiza fungi over long distances. Thus, from birds dispersed seeds and mycorrhizal fungi act as facilitating symbiotic partners into particularly suitable germination sites (Wenny 2001; Traveset et al. 2014). Also in future, efficient transport mechanisms of diaspores may contribute to maintain vegetation in equilibrium with climate under global warming conditions, herewith reducing the risk of losses of plant species as a consequence of insufficient dispersal mechanisms, enabling them to reach areas with suitable climatic conditions.
Data availability
Sequence AP1: Pollen dataset: https://data.neotomadb.org/58184; Charcoal dataset: https://data.neotomadb.org/58185.
Sequence AP2: Pollen dataset: https://data.neotomadb.org/58184; Charcoal dataset: https://data.neotomadb.org/58185.
Change history
09 May 2024
The word ‘suggesting’ is updated to ‘suggested’ in the second line of the Introduction section.
References
Allen JRM, Watts WA, Huntley B (2000) Weichselian palynostratigraphy, palaeovegetation and palaeoenvironment; the record from Lago Grande Di Monticchio, southern Italy. Quat Int 73–74:91–110
Allen JRM, Watts WA, McGee E, Huntley B (2002) Holocene environmental variability – the record from Lago Grande Di Monticchio, Italy. Quat Int 88:69–80
Atkinson MD (1992) Betula pendula Roth (B. verrucosa Ehrh.) and B. pubescens Ehrh. J Ecol 80:837–870
Bennett KD (1996) Determination of the number of zones in a biostratigraphical sequence. New Phytol 132:155–170
Berger A, Loutre MF (1991) Insolation values for the climate of the last 10 million years. Quat Sci Rev 10:297–317
Beug H-J (1975) Changes of climate and vegetation belts in the mountains of Mediterranean Europe during the Holocene. Bull Geol 19:101–110
Birks HJB (2019) Contributions of Quaternary botany to modern ecology and biogeography. Plant Ecol Divers 12:189–385. https://doi.org/10.1080/17550874.2019.1646831
Birks HH, Ammann B (2000) Two terrestrial records of rapid climatic change during the glacial–Holocene transition (14,000–9,000 calendar years B.P.) from Europe. Proc Natl Acad Sci USA 97:1390–1394
Birks HJB, Gordon AD (1985) Numerical methods in Quaternary Pollen Analysis. Academic, London
Birks HJB, Tinner W (2016) European tree dynamics and invasions during the Quaternary. In: Krumm F, Vítková L (eds) Introduced tree species in European forests: opportunities and challenges. European Forest Institute, Joensuu, pp 22–43
Birks HJB, Willis KJ (2008) Alpines, trees, and refugia in Europe. Plant Ecol Divers 1:147–160. https://doi.org/10.1080/17550870802349146
Blaauw M (2010) Methods and code for ‘classical’ age-modelling of radiocarbon sequences. Quat Geochronol 5:512–518. https://doi.org/10.1016/j.quageo.2010.01.002
Blaauw M, Heegaard E (2012) Estimation of age-depth relationships. In: Birks HJB, Lotter AF, Juggins S, Smol JP (eds) Tracking environmental change using lake sediments. data handling and numerical techniques, vol 5. Springer, Dordrecht, pp 379–413
Bolland A, Rey F, Gobet E, Tinner W, Heiri O (2020) Summer temperature development 18,000–14,000 cal. BP recorded by a new chironomid record from Burgäschisee, Swiss Plateau. Quat Sci Rev 243:106484. https://doi.org/10.1016/j.quascirev.2020.106484
Borsi S, Ferrara G, Piccoli G (1969) Determinazioni col metodo K/Ar dell’età delle eruzioni euganee (Determinations with the K/Ar method of the age of the Euganean eruptions). Rendiconti della Società Italiana di Mineralogia e Petrologia 25:27–34
Bortolami GC, Fontes JC, Markgraf V, Saliege JF (1977) Land, sea and climate in the northern Adriatic region during late Pleistocene and Holocene. Palaeogeogr Palaeoclimatol Palaeoecol 21:139–156
Cantaluppi C, Fasson A, Ceccotto F, Cianchi A, Degetto S (2014) Radionuclides Concentration in Water and Mud of Euganean Thermal District. Int J Environ Res 8:237–248. https://doi.org/10.22059/ijer.2014.713
Castiglioni GB (1940) Carta Della Massima Espansione Glaciale (scala 1:1.200.000). In: Dainelli G (ed) Atlante Fisico-Economico d’Italia. Consociazione Turistica Italiana, Milano
Combourieu-Nebout N, Paterne M, Turon J-L, Siani G (1998) A high resolution record of the last deglaciation in the Central Mediterranean Sea: palaeovegetation and palaeohydrological evolution. Quat Sci Rev 17:303–317. https://doi.org/10.1016/S0277-3791(97)00039-5
Correia M, Heleno R, da Silva LP, Costa JM, Rodríguez-Echeverría S (2019) First evidence for the joint dispersal of mycorrhizal fungi and plant diaspores by birds. New Phytol 222:1054–1060. https://doi.org/10.1111/nph.15571
Ehlers J, Gibbard PL (eds) (2004) Quaternary glaciations – extent and chronology. Part I: Europe. Elsevier, Amsterdam
Feurdean A, Bhagwat SA, Willis KJ, Birks HJB, Lischke H, Hickler T (2013) Tree migration-rates: narrowing the gap between inferred post-glacial rates and projected rates. PLoS ONE 8:e71797. https://doi.org/10.1371/journal.pone.0071797
Fineschi S, Taurchini D, Grossoni P, Petit RJ, Vendramin GG (2002) Chloroplast DNA variation of white oaks in Italy. Ecol Manage 156:103–114
Finsinger W, Schwörer C, Heiri O et al (2019) Fire on ice and frozen trees? Inappropriate radiocarbon dating lead to unrealistic reconstructions. New Phytol 222:657–662
Firbas F (ed) (1949) Spät- und nacheiszeitliche Waldgeschichte Mitteleuropas nördlich Der Alpen, vol 1. Allgemeine Waldgeschichte. Fischer, Jena
Fontana A, Mozzi P, Bondesan A (2008) Alluvial megafans in the Venetian–Friulian Plain (north-eastern Italy): evidence of sedimentary and erosive phases during late pleistocene and Holocene. Quat Int 189:71–90. https://doi.org/10.1016/j.quaint.2007.08.044
Fontana A, Mozzi P, Marchetti M (2014) Alluvial fans and megafans along the southern side of the alps. Sediment Geol 301:150–171. https://doi.org/10.1016/j.sedgeo.2013.09.003
Frisia S, Borsato A, Spötl C, Villa IM, Cucchi F (2005) Climate variability in the SE alps of Italy over the past 17 000 years reconstructed from a stalagmite record. Boreas 34:445–455. https://doi.org/10.1080/03009480500231336
Grimm E (1992–2005) Tilia version 2.0.2 and TiliaGraph 1.12. Illinois State Museum, Research and Collection Centre, Springfield
Gubler M, Henne PD, Schwörer C, Boltshauser-Kaltenrieder P, Lotter AF, Brönnimann S, Tinner W (2018) Microclimatic gradients provide evidence for a glacial refugium for temperate trees in a sheltered hilly landscape of Northern Italy. J Biogeogr 45(2):564–2575. https://doi.org/10.1111/jbi.13426
Guido MA, Molinari C, Moneta V et al (2020) Climate and vegetation dynamics of the Northern Apennines (Italy) during the Late Pleistocene and Holocene. Quat Sci Rev 231:106206
He F, Shakun JD, Clark PU, Carlson AE, Liu Z, Otto-Bliesner BL, Kutzbach JE (2013) Northern Hemisphere forcing of Southern Hemisphere climate during the last deglaciation. Nature 494(7435):81–85. https://doi.org/10.1038/nature11822
Heegaard E, Birks HJB, Telford RJ (2005) Relationships between calibrated ages and depth in stratigraphical sequences: an estimation procedure by mixed-effect regression. Holocene 15:612–618. https://doi.org/10.1191/0959683605hl836rr
Hofstetter S, Tinner W, Valsecchi V, Carraro G, Conedera M (2006) Lateglacial and Holocene vegetation history in the Insubrian Southern Alps—new indications from a small-scale site. Veget Hist Archaeobot 15:87–98. https://doi.org/10.1007/s00334-005-0005-y
Kaltenrieder P, Belis CA, Hofstetter S, Ammann B, Ravazzi C, Tinner W (2009) Environmental and climatic conditions at a potential glacial refugial site of tree species near the Southern Alpine glaciers. New insights from multiproxy sedimentary studies at Lago Della Costa (Euganean Hills, Northeastern Italy). Quat Sci Rev 28:2647–2662. https://doi.org/10.1016/j.quascirev.2009.05.025
Kaltenrieder P, Procacci G, Vannière B, Tinner W (2010) Vegetation and fire history of the Euganean Hills (Colli Euganei) as recorded by Lateglacial and Holocene sedimentary series from Lago della Costa (northeastern Italy). Holocene 20:679–695. https://doi.org/10.1177/0959683609358911
Klaus W (1972) Saccusdifferenzierungen an Pollenkörnern ostalpiner Pinus-Arten. Oesterr Bot Z 120:93–116. https://doi.org/10.1007/BF01373261
Kühl N, Moschen R (2012) A combined pollen and δ18OSphagnum record of mid-Holocene climate variability from Dürres Maar (Eifel, Germany). Holocene 22:1075–1085. https://doi.org/10.1177/0959683612441838
Lambeck K, Yokoyama Y, Purcell T (2002) Into and out of the Last Glacial Maximum: sea-level change during Oxygen Isotope Stages 3 and 2. Quat Sci Rev 21:343–360. https://doi.org/10.1016/S0277-3791(01)00071-3
Lang G (ed) (1994) Quartäre Vegetationsgeschichte Europas: Methoden Und Ergebnisse. Fischer, Jena
Lang G, Ammann B, Behre K-E, Tinner W (eds) (2023) Quaternary Vegetation dynamics of Europe. Haupt, Bern
Lawson I, Frogley M, Bryant C, Preece R, Tzedakis P (2004) The Lateglacial and Holocene environmental history of the Ioannina basin, north-west Greece. Quat Sci Rev 23:1599–1625
Livingstone DM, Lotter AF (1998) The relationship between air and water temperatures in lakes of the Swiss Plateau: a case study with palaeolimnological implications. J Paleolimnol 19:181–198. https://doi.org/10.1023/A:1007904817619
Lona F (1957) I depositi lacustri Euganei: Archivio Paleontologico Del Tardoglaciale E Del periodo postglaciale. Mem Biogeogr Adr 5:3–11
Lotter AF (1999) Late-glacial and Holocene vegetation history and dynamics as shown by pollen and plant macrofossil analyses in annually laminated sediments from Soppensee, central Switzerland. Veget Hist Archaeobot 8:165–184. https://doi.org/10.1007/BF02342718
Lourantou A, Lavrič JV, Köhler P et al (2010) Constraint of the CO2 rise by new atmospheric carbon isotopic measurements during the last deglaciation. Glob Biogeochem Cycles 24:GB2015. https://doi.org/10.1029/2009GB003545
Lowe JJ, Accorsi CA, Bandini Mazzanti M et al (1996) Pollen stratigraphy of sediment sequences from lakes Albano and Nemi (near Rome) and from the central Adriatic, spanning the interval from oxygen isotope stage 2 to the present day. In: Guilizzoni P, Oldfield F (eds) Palaeoenvironmental Analysis of Italian Crater Lake and Adriatic sediments. Memorie dell’Istituto Italiano Di Idrobiologia 55. Istituto Italiano di Idrobiologia, Verbania-Pallanza, pp 71–98
Luetscher M, Boch R, Sodemann H et al (2015) North Atlantic storm track changes during the Last Glacial Maximum recorded by Alpine speleothems. Nat Commun 6:6344. https://doi.org/10.1038/ncomms7344
Magri D (1999) Late Quaternary vegetation history at Lagaccione near Lago Di Bolsena (central Italy). Rev Palaeobot Palynol 106:171–208
Marcolla A, Miola A, Mozzi P, Monegato G, Asioli A, Pini R, Stefani C (2021) Middle Pleistocene to Holocene palaeoenvironmental evolution of the south-eastern Alpine foreland basin from multi-proxy analysis. Quat Sci Rev 259:106908. https://doi.org/10.1016/j.quascirev.2021.106908
Miles J, Kinnaird JW (1979) The establishment and regeneration of birch, juniper and Scots pine in the Scottish Highlands. Scott for 33:102–119
Monegato G, Ravazzi C, Donegana M, Pini R, Calderoni G, Wick L (2007) Evidence of a two-fold glacial advance during the last glacial maximum in the Tagliamento end moraine system (eastern Alps). Quat Res 68:284–302. https://doi.org/10.1016/j.yqres.2007.07.002
Monegato G, Ravazzi C, Culiberg M et al (2015) Sedimentary evolution and persistence of open forests between the south-eastern Alpine fringe and the Northern Dinarides during the Last Glacial Maximum. Palaeogeogr Palaeoclimatol Palaeoecol 436:23–40. https://doi.org/10.1016/j.palaeo.2015.06.025
Mozzi P, Piovan S, Rossato S, Cucato M, Abbà T, Fontana A (2010) Palaeohydrography and early settlements in Padua (Italy). Il Quaternario (Ital J Quat Sci) 23:387–400
Mozzi P, Ferrarese F, Fontana A (2013) Integrating digital elevation models and stratigraphic data for the reconstruction of the post-LGM unconformity in the Brenta alluvial megafan (North-Eastern Italy). Alp Mediterr Quat 26:41–54
Paganelli A, Miola A (1991) Chestnut (Castanea sativa Mill.) as an indigenous species in northern Italy. Il Quaternario 4:99–106
Panagiotopoulos K, Aufgebauer A, Schäbitz F, Wagner B (2013) Vegetation and climate history of the Lake Prespa region since the Lateglacial. Quat Int 293:157–169
Piccoli G, Bellati R, Binotti C et al (1976) Il Sistema Idrothermale Euganeo-Berico E La Geologia dei Colli Euganei. Mem Ist Geol Min Univ Padova 30:266 una carta geologica e tavola di pro-fili geologica 1:25000, Padova
Piccoli G, Sedea R, Bellati R et al (1981) Note illustrative della carta geologica dei Colli Euganei alla scala 1:25000. Mem Ist Geol Univ Padova 34:523–566 una carta geologica 1:25000, Padova
Pignatti S (1982) Flora d’Italia, Vols 1–3, 1st edn. Edagricole, Bologna
Pini R, Ravazzi C, Donegana M (2009) Pollen stratigraphy, vegetation and climate history of the last 215 ka in the Azzano Decimo core (plain of Friuli, north-eastern Italy). Quat Sci Rev 28:1268–1290. https://doi.org/10.1016/j.quascirev.2008.12.017
Pini R, Ravazzi C, Reimer PJ (2010) The vegetation and climate history of the last glacial cycle in a new pollen record from Lake Fimon (southern Alpine Foreland, N-Italy). Quat Sci Rev 29:3:115–3137. https://doi.org/10.1016/j.quascirev.2010.06.040
Ravazzi C, Orombelli G, Tanzi G, CLIMEX group (2004) An outline of the flora and vegetation of Adriatic basin (Northern Italy and eastern side of the Apennine) during the last glacial Maximum. In: Antonioli F, Vai GB (eds) Litho-paleoenvironmental maps of Italy during the last two climatic extremes. Explanatory notes. Museo geologico Giovanni Capellini, Bologna, pp 15–20
Ravazzi C, Donegana M, Vescovi E et al (2006) A new late-glacial site with Picea abies in the northern Apennine foothills: an exception to the model of glacial refugia of trees. Veget Hist Archaeobot 15:357–371. https://doi.org/10.1007/s00334-006-0055-9
Ravazzi C, Pini R, Badino F, De Amicis M, Londeix L, Reimer PJ (2014) The latest LGM culmination of the Garda Glacier (Italian Alps) and the onset of glacial termination. Age of glacial collapse and vegetation chronosequence. Quat Sci Rev 105:26–47
REGPARK (2015) Regional Parc of Euganean Hills. http://www.collieuganei.it/laghifiumi-zone-umide/lago-della-costa/
Reimer PJ, Austin WEN, Bard E et al (2020) The IntCal20 Northern Hemisphere radiocarbon age calibration curve (0–55 cal kBP). Radiocarbon 62:725–757. https://doi.org/10.1017/RDC.2020.41
Rey F, Gobet E, van Leeuwen JFN et al (2017) Vegetational and agricultural dynamics at Burgäschisee (Swiss Plateau) recorded for 18,700 years by multi-proxy evidence from partly varved sediments. Veget Hist Archaeobot 26:571–586. https://doi.org/10.1007/s00334-017-0635-x
Rey F, Gobet E, Schwörer C, Hafner A, Szidat S, Tinner W (2020) Climate impacts on vegetation and fire dynamics since the last deglaciation at Moossee (Switzerland). Clim Past 16:1347–1367. https://doi.org/10.5194/cp-16-1347-2020
Samartin S, Heiri O, Lotter AF, Tinner W (2012) Climate warming and vegetation response after Heinrich event 1 (16 700–16 000 cal yr BP) in Europe south of the Alps. Clim Past 8:1913–1927. https://doi.org/10.5194/cp-8-1913-2012
Samartin S, Heiri O, Kaltenrieder P, Kühl N, Tinner W (2016) Reconstruction of full glacial environments and summer temperatures from Lago Della Costa, a refugial site in Northern Italy. Quat Sci Rev 143:107–119. https://doi.org/10.1016/j.quascirev.2016.04.005
Samartin S, Heiri O, Joos F, Renssen H, Franke J, Brönnimann S, Tinner W (2017) Warm Mediterranean mid-Holocene summers inferred from fossil midge assemblages. Nat Geosci 10:207–212. https://doi.org/10.1038/ngeo2891
Shaw MW (1974) The reproductive characteristics of oak. In: Morris MG, Perring FH (eds) The British Oak: its history and natural history. Botanical Society of the British Isles, Faringdon, pp 162–181
Soter S (2011) Radiocarbon anomalies from old CO2 in the soil and canopy air. Radiocarbon 53:55–69. https://doi.org/10.1017/S0033822200034354
Stuiver M, Reimer PJ (1993) Extended 14C data base and revised CALIB 3.0 14C age calibration program. Radiocarbon 35:215–230
Tinner W, Kaltenrieder P (2005) Rapid responses of high-mountain vegetation to early Holocene environmental changes in the Swiss Alps. J Ecol 93:936–947
Tinner W, Lotter AF (2006) Holocene expansions of Fagus silvatica and Abies alba in Central Europe: where are we after eight decades of debate? Quat Sci Rev 25:526–549. https://doi.org/10.1016/j.quascirev.2005.03.017
Tinner W, Vescovi E (2007) Ecologia e oscillazioni del limite degli alberi nelle Alpi dal Pleniglaciale al presente. In: Frisia S, Filippi ML, Borsato A (eds) Cambiamenti climatici e ambientali in Trentino: dal passato prospettive per il futuro. Museo Tridentino di Scienze Naturali, Trento, pp 7–15
Tinner W, Hubschmid P, Wehrli M, Ammann B, Conedera M (1999) Long-term forest fire ecology and dynamics in southern Switzerland. J Ecol 87:273–289. https://doi.org/10.1046/j.1365-2745.1999.00346.x
Tinner W, Colombaroli D, Heiri O et al (2013) The past ecology of Abies alba provides new perspectives on future responses of silver fir forests to global warming. Ecol Monogr 83:419–439
Traveset A, Heleno R, Nogales M (2014) The ecology of seed dispersal. In: Gallagher RS (ed) Seeds: the ecology of regeneration in plant communities, 3rd edn. CAB International, Wallingford, pp 62–93
Tzedakis PC, Emerson BC, Hewitt GM (2013) Cryptic or mystic? Glacial tree refugia in northern Europe. Trends Ecol Evol 28:696–704. https://doi.org/10.1016/j.tree.2013.09.001
Vescovi E, Ravazzi C, Arpenti E et al (2007) Interactions between climate and vegetation during the Lateglacial period as recorded by lake and mire sediment archives in Northern Italy and Southern Switzerland. Quat Sci Rev 26:1650–1669. https://doi.org/10.1016/j.quascirev.2007.03.005
Watts WA, Allen JRM, Huntley B (1996) Vegetation history and palaeoclimate of the last glacial period at Lago Grande di Monticchio, Southern Italy. Quat Sci Rev 15:133–153
Welten M (ed) (1982) Vegetationsgeschichtliche Untersuchung in den Westlichen Schweizer Alpen: Bern – Wallis. Denkschriften der Schweizerischen Naturforschenden Gesellschaft 95. Birkhäuser, Basel
Wenny DG (2001) Advantages of seed dispersal: a re-evaluation of directed dispersal. Evol Ecol Res 3:51–74
Wick L (1996) Late-glacial and early-holocene palaeoenvironments in Brianza, N Italy. Il Quaternario 9:653–660
Wijmstra TA (1969) Palynology of the first 30 metres of a 120 m deep section in northern Greece. Acta Bot Neerl 18:511–527
Willis KJ, van Andel TH (2004) Trees or no trees? The environment of central and eastern Europe during the last glaciation. Q Sci Rev 23 (23–24):2,369-2,387
Acknowledgements
We are much obliged to Sig. Martinengo, longtime resident by Lago della Costa, Ing. Drago and Ing. De Benedetti from Soc. Fango Lago Costa d’Arqua s.r.l. for their help in identifying the undisturbed lake coring spot as well as for providing historical and bathymetric information. We thank W. Tanner, W. Finsinger and F. Franchini for help during coring, F. Oberli for her generous laboratory assistance and C. Schwörer for helping in statistics. We are grateful to S. Samartin, F. Rey, E. Vescovi, C. A. Belis, S. Hofstetter, G. Procacci, B. Vannière, C. Ravazzi and B. Ammann for data support, fruitful discussions and co-authoring the former publications at Lago della Costa.
Funding
Open access funding provided by University of Bern
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors have no conflict of interest to declare that are relevant to the content of this article.
Additional information
Communicated by T. Giesecke.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Boltshauser-Kaltenrieder, P., Tinner, W. Early expansion of mixed oak stands at 16,800–16,600 cal bp at a northern Italian glacial refugium in the Euganean Hills. Veget Hist Archaeobot (2024). https://doi.org/10.1007/s00334-024-00997-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00334-024-00997-7