
Abstract
Pollen diagrams covering the Bronze Age to Roman period from northeast Northumberland are scarce. We present a 14C-dated pollen record from a peat-filled forest hollow in Chillingham Wild Cattle Park, northeast Northumberland, that spans the Iron Age. For the first time for this part of Northumberland, fungal spores are also analysed to investigate whether clearances took place in the context of crop cultivation or pastoralism. The pollen diagram shows significant woodland cover during the Late Bronze Age, with small-scale landscape openness potentially representing crop cultivation and animal grazing under a tree canopy. A local clearance event, with arboreal pollen falling to 13.2%, is radiocarbon dated to the late Early to Middle Iron Age, likely coinciding with the construction and occupancy of the nearby Ros Castle hillfort. A significant rise in dung fungal spores and the presence of Plantago and other open vegetation taxa indicate animal husbandry played an important role in this clearance phase. A subsequent large decrease in dung fungi and a small rise in arboreal pollen may reflect a degree of abandonment of the area after the climate cooled. Peat formation ceased in the late Middle to Late Iron Age as the hollow filled up. Limited renewed peat growth occurred in the last century or so, after peat shrinkage due to drying and consequent oxidation provided the possibility for renewed peat formation within the hollow.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Northumberland, in northeast England, is rich in hillforts, other settlements and the remains of field systems, but to date Iron Age vegetation changes are poorly documented. Complete forest clearance does not seem to have taken place before the Iron Age, earlier clearance events being small-scale and of short duration (Turner 1968; Mannion 1978 (across the border in Scotland); Davies and Turner 1979; Moyle 1980; Macklin et al. 1991; Hewitt et al. 1996). Existing pollen diagrams from the area cover the Lateglacial to the Bronze Age (e.g. Akeld Steads/Milfield Basin, Borek 1975, Tipping 1998; Bradford Kaims and Longlee Moor, Bartley 1966, undated; Embleton Bog, Dixon et al. 2015; Wooler Water, Clapperton et al. 1971, dated in Tipping 1992; Din Moss, Roxburghshire (across the border in Scotland); Hibbert and Switsur 1976). Other diagrams cover the later part of the Holocene but have not been radiocarbon dated (e.g. Broad Moss, Davies and Turner 1979; Linton Loch, Mannion 1978; Coom Rigg Moss, Chapman 1964; Edlingham, Moyle 1980; Trickley Wood, Turner 1968). A radiocarbon dated profile is available from Wether Hill, but this mainly covers the Roman period (Davies and Dixon 2007).
The core from Camp Hill Moss (Davies and Turner 1979) was for many years the only dated core from northeast Northumberland that spans the Bronze Age to the Roman period. However, the crucial date of 640 bp for the first extensive clearance in the sequence at Camp Hill Moss, at a depth of 52.5–54 cm, is problematic, and Davies and Turner (1979) suggest this more likely represents a Roman or early Medieval clearance. Tipping (1992) reinterprets the diagram as containing a hiatus, possibly due to Medieval peat cutting. The study also predates the advances in accelerator mass spectrometry (AMS) dating from the 1980s onwards. Davies and Turner (1979) correlated the clearance phase at Camp Hill Moss with the closely similar, but undated, record at Broad Moss. This phase at Broad Moss was dated by Passmore and Stevenson (2012) in a new core to 2,230 ± 70 bp (2,360–2,150 cal bp).
The first extensive clearance at 175 cm depth in a core from Ford Moss is estimated to date from ca. 2,170 bp (Davis and Passmore 2012), but this is extrapolated upwards from the most recent radiocarbon date of 2,570 ± 40 bp (2,760–2,500 cal bp) at 217 cm. Two thin peat beds underneath and near the base of a gravel fill on a terrace of the River Breamish near Powburn were dated by radiocarbon to 2,743 and 2,356 cal bp, and 1,723, 2,334 and 1,946 cal bp, respectively (Tipping 1992), with clearance taking place between these two records. Further south, a dated soil profile from Callaly Moor (Macklin et al. 1991) may contain Iron Age pollen assemblages. Further north, an intensive clearance phase in the extensive raised mire of Dogden Moss in the Scottish Borders, which started at 2,520 cal bp, shows that from a more regional perspective, clearance started in the Early to Middle Iron Age (Dumayne-Peaty 1999).
Inferences about the relative importance of crop cultivation and animal husbandry in the area have so far been based exclusively on pollen evidence, in particular the pollen of cereals and arable weeds such as Polygonum, Papaver rhoeas-type, Brassicaceae, Caryophyllaceae and Amaranthaceae (Chenopodiaceae) as evidence for crop cultivation (Davies and Turner 1979; Moyle 1980; Davies and Dixon 2007; Dixon et al. 2015). A predominantly pastoral system is deduced from the absence of or low values for such taxa, as well as the presence of herbs indicative of disturbance, such as Plantago lanceolata, Ranunculus acris-type, Rumex acetosa, Cichorium intybus-type and Succisa pratensis (Davies and Dixon 2007).
Over the past decades, counts of coprophilous fungal spores from sedimentary sequences have increasingly been used as indicators of herbivore presence, and, therefore, potential impacts of animal husbandry (e.g. van Geel 2006 and articles in that volume; Haas 2010 and articles in that volume; Perrotti and van Asperen 2019; Gauthier and Jouffroy-Bapicot 2021; van Asperen et al. 2021). No dung fungal spores have so far been studied in cores from northeast Northumberland (although there is a single record from the area near Hadrian’s Wall; Yeloff et al. 2007). Here we present a 14C-dated pollen and fungal spore record from Chillingham Wild Cattle Park, northeast Northumberland, that covers the Iron Age, to clarify the timing of the first extensive clearance events in the area, and to investigate whether this clearance was mainly due to crop cultivation or to pastoralism.
Study site
Chillingham Wild Cattle Park, Northumberland (Fig. 1), located between the Cheviot Hills to the west and the coastal plains on the east, was a typical deer park until 1750. The flatter, lower ground was covered by the ‘Great Wood of Chillingham’ (Hall 2013). Open wood pasture surrounded it on the hillsides. The wood was felled around 1754. In the late 1700s the estate steward removed the ditches, walls and hedges, and dug out all the stumps of the Great Wood. The lower ground was converted into pasture, whilst new oak and beech woods were planted on the hillsides (Hall 2013). The ash and alder trees on the wetter ground (now known as ‘The Allers’) were left to regrow from the stumps. The park was developed from 1799 to accommodate the cattle. It was made to look ‘natural’ but includes some non-native trees. The hillside woodlands were felled between 1914 and 1918, at which time they predominantly consisted of oak with some conifers also present (S. Hall, pers. comm., February 2023). Plantations consisting mainly of pine were planted on the hillside in the 1950 and 1960s.

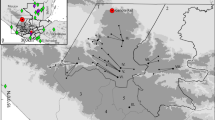
Location of core CC14 (1) and modern samples (2 and 3) in Chillingham Wild Cattle Park. Green areas represent woodland; contour lines are at 5 m intervals. Inset map: overview map of northern England and Scotland; a rectangle indicates the location of Chillingham Wild Cattle Park. The maps include public sector information and Ordnance Survey data licensed under the Open Government Licence v3.0. © Crown copyright and database rights 2012, 2019 Ordnance Survey 100,025,252
A feral herd of Chillingham cattle has occupied the park since at least 1646 (Hall 2007, 2013). The herd was probably founded by the gathering together of attractive cattle from husbanded herds sometime between 1225 and 1646, when the Park became a hunting park (Hall 1988). At present, the herd consists of about 100 animals, which are roaming freely in the park and do not receive any veterinary intervention. Between 1914 and 2007 there were several hundred sheep in the park (S. Hall, pers. comm., February 2023). The park is also frequented by fallow and roe deer, badgers, foxes and a variety of smaller mammals.
Although there are not many Roman remains, the Roman road known as the Devil’s Causeway ran through the nearby Valley of the Till. The (unexcavated) Iron Age hillfort of Ros Castle lies on the edge of the Park (Blood 1995), and many other hillforts and remains of settlements are present in the surrounding area (Jobey 1965; Cunliffe 2005), the nearest being at Old Bewick. The area has many cairnfields (Jobey 1968; Davies and Turner 1979; Macklin et al. 1991), the nearest being on Millstone Hill, Willie Law, Camp Hill and Chatton Sandyfords just east of the Park (Jobey 1968; Davies and Turner 1979). Some of these are funeral cairns, while others relate to field clearance. There are well-preserved remains of cultivation (Topping 1989, 1990/1991; Hewitt et al. 1996; Frodsham 2004).
Methods
A 136-cm-long core (CC14) was extracted from the deepest part of a small depression (~ 130 m diameter) in The Allers, a riparian wetland adjacent to a small stream (55.52537, −1.88903; 110 m above Ordnance Datum) on 2 December 2014 using a Russian corer with a diameter of 5 cm. The core was removed in 50 cm segments. Core segments were transferred into lengths of plastic drainpipe, wrapped in cling film, and, once transferred to the laboratory, stored at 4 °C.
Prior to pollen sampling, magnetic susceptibility measurements were taken at intervals of 2 cm with a Bartington Magnetic Susceptibility Meter MS2, using the Surface Scanning Sensor MS2E (Dearing 1999).
Since the core contained wood fragments, seven wood samples (not identified) were selected that on visual inspection were horizontally orientated rather than vertically, and that did not have the appearance of roots. These, along with a bulk peat sample, were submitted for AMS radiocarbon dating to Beta Analytic. The radiocarbon ages were input to a model which estimates the accumulation rate and any changes in it with Bayesian statistics, using the R package “rbacon2” v. 3.0.0 (Blaauw and Christen 2011). Constraints were imposed based on prior information. This produced an age-depth model that describes the accumulation history of the core. Below, calibrated dates are given as ‘cal bp’, and estimated ages from the Bacon model are indicated as such.
Subsamples of 0.5 cm thickness with a sampling resolution of 10 cm to a depth of 110 cm were prepared using standard pollen analytical methods (Stockmarr 1971; Moore et al. 1991), modified to maximise fungal spore recovery (van Asperen et al. 2016). Two Lycopodium tablets (n spores = 37,168, batch no. 177745, Laboratory of Quaternary Biology, Lund University, Sweden) dissolved in 10 mL 10% HCl were added to enable calculation of concentrations (Hicks et al. 1999). The samples were first heated in a 10% KOH solution to promote disaggregation of material and remove soluble organic compounds. Items larger than 125 μm were removed by sieving. Particles < 6 μm were removed with a fine mesh sieve. The samples were then treated with 10% HCl, stained with safranine and mounted in silicone oil using tertiary butyl alcohol. Pollen and fungal spores were identified and counted at 400× magnification until at least 350 Lycopodium spores (Etienne and Jouffroy-Bapicot 2014) and 200 pollen and fern spores had been counted. Pollen was sparse in some samples, and these samples were counted until at least 500 Lycopodium spores had been counted (see below, Perrotti et al. 2022). Taxonomical nomenclature follows Stace (2019) for vascular plants, Bennett (1994) for pollen and Bell (2005), Doveri (2007) and Guarro et al. (2012) for fungal spores. Plantago includes all species of Plantago.
Total pollen (TP) consists of the total of all land pollen, excluding Cyperaceae, indeterminate grains, Sphagnum and ferns, but including Alnus. Whilst Alnus pollen are normally removed from TP for alder carr deposits as their often large numbers would swamp the signal of the other pollen types (Janssen 1959), in this case Alnus pollen were not numerous. Unless otherwise indicated below, percentage values are based on TP as defined above. Sphagnum, fern and fungal spore abundances were expressed as percentages of TP. Concentrations for all taxa were also calculated.
The pollen diagram was created using the R package “rioja” v1.0-5 (Juggins 2020) with R v. 4.0.4 (R Core Team 2021). Pollen zones were defined based on the results of a stratigraphically constrained hierarchical cluster analysis (CONISS) using the Euclidean distance of pollen percentage data for AP and NAP with the R package “vegan” v. 2.6-4 (Oksanen et al. 2022). These were tested against a broken stick model to obtain the number of pollen zones (Bennett 1996).
For comparison, pollen and NPP assemblages from several 6-month collection seasons (October 2014–October 2016) from a pollen trap (CT2; van Asperen et al. 2020) located 15 m from the coring site and not accessible to the cattle are included, as well as from three soil samples taken around the pollen trap (CS2) and three further soil samples taken 15 m to the south-east of the coring site (CS3), near a burn.
Results
Core description
The bottom deposit in the core consists of blueish light-grey sandy clay (Fig. 2). The clay is probably of glacial origin, as the area’s superficial deposits consist of Late Weichselian deposits. Its impermeable nature may have contributed to the subsequent growth of the peat lying above. Between 116 and 125 cm, alternating bands of peat and clay occur. The top 116 cm of the core consisted of highly humified brown peat with larger pieces of wood and other plant macrofossils. Pollen analysis was carried out from the point where peat accumulation became more stable and episodic erosion events seem to have ceased, from 110 cm upwards.

Summary pollen diagram from core CC14, based on percentage TP excluding Cyperaceae and ferns. Magnetic susceptibility values and estimated ages in years bp based on the Bacon model are also shown
Magnetic susceptibility
Magnetic susceptibility values are high between 134 and 116 cm (Fig. 2), reflecting the glacial clay as well as the zone of episodic erosional deposits between 125 and 116 cm, which is also visible in the fluctuating values between 122 and 116 cm. The higher values are likely due to the presence of iron-containing minerals. Magnetic susceptibility then becomes slightly negative between 114 and 18 cm, as is typical for diamagnetic materials such as peat (Mullins 1977; Blundell et al. 2009). There was thus likely little input of inorganic sediment in this part of the sequence. This is followed by slightly positive values between 18 and 2 cm. It is well-known that topsoils in temperate areas show higher values of magnetic susceptibility than subsoils (Le Borgne 1955; Mullins 1977). Furthermore, in the post-industrial revolution, especially post-1945, pollution is registered in peat bogs, even when they are, like Chillingham, located at a significant distance from industrial centres (Oldfield et al. 1978). The slightly higher values may also indicate a small influx of inorganic sediment through sheet flow, although this was not visible in the stratigraphy.
Dating
Radiocarbon dates are presented in Table 1. Three dates are recent: one bulk-peat date based on percent modern carbon (pMC) at 30 cm, and two at 75 and 80 cm. The other five dates all fall in the Iron Age. Given the Iron Age dates are stratigraphically consistent with each other, and a modern date for almost the entire sequence is unlikely due to the highly humified character of the peat, it is likely that the modern dates are due to root disturbance or contamination with modern material. These samples were taken after several years of core storage, in contrast to the Iron Age dates of 2,430 ± 30 bp at 58–60 cm and 2,470 ± 30 bp at 107–109 cm, which were obtained on material sampled from the fresh core. The three modern dates were thus disregarded in the Bacon model. The part of the core between 52 and 109 cm could potentially cover the period between 2,716 and 2,318 cal bp, which covers the Earliest and Early Iron Age according to Cunliffe’s (2005) scheme. A more precise date cannot be given for this section of the core due to the presence of the Iron Age or Hallstatt plateau in the radiocarbon calibration curve. The date of 2,319–2,115 cal bp at 35 cm places this part of the core in the Middle Iron Age. Priors for the Bacon model (Fig. 3) included an age of −64 ± 1 at the top of the core (reflecting the year the core was taken) and an age of 0 ± 20 (ad 1950) at 20 cm as an estimate of the date for the widespread planting of Pinus on the hillsides surrounding the core location. Based on the changes in the pollen and NPP content at 30 cm documented below we specified a hiatus at 32 cm with a maximum age of 2,055 bp (the mean of the calibrated radiocarbon date at 35 cm, plus 60 years for 3 cm of deposition at 20 year/cm). The model estimates a mean age (95% confidence level) of 2,803 bp at 108 cm, with a fairly stable accumulation rate until a mean age of 2,100 bp at 35 cm. The short time period between the calibrated dates of 2,716 and 2,318 cal bp for the section of the core between 109 and 52 cm on the one hand, and 2,115 cal bp at 35 cm on the other, suggests that the core spans a maximum of ~ 700 years of deposition. The small hollow in which the deposit formed seems to have been initially filled not many centuries after the Middle Iron Age. Peat formation then ceased, until some further growth occurred in the last century or so when the peat had shrunk due to drying out and consequent oxidation.

BACON-derived radiocarbon age-depth model for core CC14. In the main graph, the blue markers represent the calibrated radiocarbon constraints and the green markers the priors derived from the recent known history of the Park vegetation; the vertical dotted black line indicates the position of the postulated hiatus; the dotted red line with grey shading indicates the best age-model solution based on the mean age for each depth with a 95% confidence interval. The three upper graphs show, from left to right, the stability of the Markov Chain Monte Carlo iterations and the prior (green line) and posterior (grey shading) accumulation rate, memory and hiatus size
Pollen zones
The broken stick model indicates two pollen zones can be recognized. CONISS indicates the main transition in pollen assemblages is between the samples at 60 and 70 cm (Fig. 5). However, the statistical model is weakened by the relatively small number of samples and large number of variables (taxa). Furthermore, the model assumes a stratigraphically continuous relationship, which we know in this core is broken by a hiatus in accumulation at 32 cm. Based on the dating (Fig. 3), concentration (Fig. 4), pollen and NPP evidence (Fig. 5), and the assumed location of the hiatus, we identify another transition around 30 cm.

Pollen and selected NPPs concentration diagram, core CC14; total pollen includes Cyperaceae

Pollen and NPP diagram with CONISS clusters from core CC14, based on %TP excluding Cyperaceae and ferns (NB Cyperaceae based on %TP including Cyperaceae, excluding ferns)
Zone CC14-A (110–70 cm)
The percentage of arboreal pollen (AP) in the samples from 110 to 70 cm (45–56%; Fig. 2) is around the 50% threshold often quoted for woodland presence near an upland site (Birks 1988) or relatively open canopy woodlands (Tinsley and Smith 1974). It is therefore likely that stands of trees, and probably areas of woodland (cf. Bunting 2002) were present on or near the site. These values, and the lack of disturbance indicators, except for low Plantago values, indicate that the vegetation was largely stable during this period. Woodland stands consisted mainly of Alnus (9–24%), Betula (3–20%), Corylus-type (2–11%) and Quercus (2–18%). Lower values are recorded for Pinus (0–5%), Fraxinus (0–7%) and Ulmus (0–3%). The high values for Alnus reflect trees near to the coring site growing in wet local conditions. However, open areas were also present, reflected in the values for grassland species (Poaceae, 20–43%). The small reduction in AP values at 100 cm may represent a limited and temporary opening up of the landscape. Betula and Ericales (20%) reach their highest values at 80 cm, whilst Alnus and Poaceae are at their lowest, indicating a small-scale disturbance, possibly reflecting a slight drying out of the area. However, patches of woodland persisted, since AP remains at 54% and anthropogenic disturbance indicators such as Asteraceae remain at low levels. The concomitant peak in dung fungi (Figs. 4 and 5) may reflect an increased use of the woodland for animal husbandry.
Zone CC14-B (70–30 cm)
Between 70 and 60 cm AP falls from 51 to 27%, and from 50 cm upwards does not recover its former abundance (Fig. 2). The local area seems to have become increasingly wet, with Cyperaceae values rising from 20% TP (TP incl. Cyperaceae) at 70 cm to 36% TP (TP incl. Cyperaceae) at 60 cm, remaining high with a peak of 60% TP (TP incl. Cyperaceae) at 30 cm (Fig. 5). At the same time, Poaceae increases to a peak of 77% TP (excl. Cyperaceae) at 30 cm. Plantago, an indicator of pastoral activities (Behre 1981), is present in most samples in the sequence, but is more common between 60 and 30 cm (0–4%). Asteraceae are also more abundant (2–13%). A slight increase of AP to 22% at 40 cm could indicate a small degree of forest regeneration.
Zone CC14-C (30–0 cm)
Although there is no clear change in the character of the peat itself, there are a number of changes in the pollen assemblage around 30–20 cm. TP concentrations are clearly higher above this depth than below it (Fig. 4). Magnetic susceptibility is higher above 20 cm (Fig. 2). The small peak in Quercus at 20 cm is followed by an increase in Pinus and Picea, reflecting recent vegetation changes on the slopes around the park and the present-day regional presence of plantations (Fig. 5). The trees that grow in The Allers at present (Fraxinus, Salix and to a lesser extent Alnus), as well as the lesser clubmoss Selaginella, increase from 20 cm upwards.
Pastoral indicators Plantago (2–5%) and Asteraceae (6–7%) are present. Interestingly, Potentilla-type pollen occurred in the top 10 cm of the core (1–2%), which may be due to the presence of sheep in the Park in the last 100 years, as sheep grazing creates conditions in which Potentilla erecta thrives (Moore et al. 1986). A small number of spheroidal carbonaceous particles (SCPs) is present in the top two samples. SCPs are a by-product of high-temperature combustion of fossil fuels, and are first recorded in Britain in the 19th century, with a peak in the mid-20th century (Swindles 2010; Swindles et al. 2015). Their presence further corroborates the young age of these sediments. If there was inwash of reworked sediment from elsewhere, we would expect the proportion of indeterminate grains to be higher in the top 20–30 cm of the core, as well as a change in the character of the deposit. Since this is not the case, it seems likely that the small hollow had filled up and deposition ceased. Renewed deposition took place when the peat had shrunk due to repeated drying out and consequent oxidation. The lowering of the ground surface through shrinkage likely created space for the subsequent recent sediment accumulation and provides a rationale for the hiatus in the chronology.
Cereals
No cereal-type pollen (defined as Poaceae pollen with a diameter > 40 μm and a protuberant annulus with a diameter > 10 μm; Andersen 1979; Dickson 1988; Joly et al. 2007) were recorded. Cereals are self-pollinating, with the exception of the wind-pollinated Secale, which was introduced to Britain in the late Iron Age or Roman period, although there are sparse earlier records from southern England and Wales (Chambers and Jones 1984). The absence of cereal-type pollen may indicate there was no cereal cultivation in the immediate vicinity of the core which, especially in the wet conditions evidenced in the upper part of the core, is to be expected. As cereals are self-pollinating and hence do not disperse widely, cereals could have been cultivated nearby without being encountered in the core, given the low overall pollen count (range: 59–240; average: 130). However, other, more widely dispersing indicators of arable agriculture are largely absent.
Non-pollen palynomorphs
High concentrations for dung fungal spores are regarded as reflecting the presence of herbivores (van Asperen et al. 2021). In Chillingham Wild Cattle Park, current dung fungal spore influx correlates primarily to winter grazing, whilst summer grazing is less visible in the dung fungal spore record (van Asperen et al. 2020). A number of coprophilous fungal spore types (van Asperen et al. 2021) were encountered, including Hypocopra-type (BRN-8), Podospora-type (HdV-368), Sordaria-type (HdV-55A), Sporormiella-type (HdV-113) and Trichodelitschia-type (HdV-546) (Fig. 5). These are here combined into a total dung fungal spore percentage (Figs. 2, 4 and 5). Expressed as %TP, dung fungi are relatively common throughout the sequence, above the 2% threshold that is often quoted as representing a background level of deposition (Gill et al. 2013; but see Perrotti and van Asperen 2019) in all samples. Dung fungi are particularly abundant from a first peak of 37% at 80 cm, coinciding with the drying out of the area (see above), and remain high to 50 cm, with values of 29–51%. Dung fungal diversity is also highest in this part of the sequence. Between 40 cm and the top of the core, dung fungal spore counts are comparatively lower at 13–21%, but still significantly above the background level.
Quantification of fungal spores as %TP is sensitive to fluctuations in pollen accumulation rates (Perrotti and van Asperen 2019). For this reason, dung fungal spore concentrations can provide additional evidence of herbivore presence. Due to the variation in total pollen concentrations (Fig. 4), dung fungal spore concentrations show a different pattern from dung fungal abundance expressed as %TP. The pattern of low values for samples from 110 to 90 cm and high values for samples from 80 to 50 cm is maintained in the concentration data. Values rise significantly above background levels at 80 cm and initially remain high whilst the environment became wetter. There is still significant tree cover when dung fungal spore values started to rise, but the peak at 80 cm may represent grazing under a tree canopy.
The dung fungal concentrations from 40−0 cm show a different pattern to the percentage data. After a clear drop at 40 cm, dung fungal spores increase again, reaching their highest concentrations in the entire core at 10 cm. This later peak cannot be attributed solely to preservation, since other fungal spores show a range of patterns of variation (Fig. 4). Furthermore, the conditions that adversely affect pollen preservation do not necessarily affect the preservation of fungal spores to the same extent (Perrotti and van Asperen 2019). Error margins on estimated dung fungal spore concentrations are higher at low overall spore counts, and at low tracer (Lycopodium) counts (Perrotti et al. 2022). Since we obtained high Lycopodium counts on samples poor in fungal spores, this should reduce the uncertainty associated with calculating concentrations. Given the recent historical and current high density of grazers in the park, the high concentrations in near-surface samples are not surprising and support the use of these palynomorphs as a grazing indicator.
Concentrations of Glomus-type (HdV-207), an endomycorrhizal fungus that forms chlamydospores, the presence of which in pollen samples are taken as indicative of soil erosion (van Geel et al. 1989, 2003), are high in the bottom part of the core (81% at 110 cm; Fig. 4), in accordance with the disturbed character of the peat and clay. At 20 cm there is a peak in Glomus-type (111%). Although the present-day open parkland may be susceptible to erosion during high rainfall events, the magnetic susceptibility values and the character of the peat itself do not reflect substantial inwash of mineral sediments from elsewhere (see above). These spores may therefore be autochthonous and associated with the roots of the present-day peat-forming vegetation (Kołaczek et al. 2013).
Pollen concentrations
Total pollen concentrations (Fig. 4) are generally higher in the top 30 cm of the core and lower in the deeper samples. However, the lowest pollen concentrations are found in the middle of the core in pollen zone CC14-B (see below). The preservation of pollen deteriorates from the top to the bottom of the core. This was to be expected as the peat has a highly humified character. It is probable that the peat was subject to (periodical/seasonal) drying out after the cessation of deposition, leading to oxidation and hence loss and deterioration of pollen. Alder wood peat is known to generally contain poorly preserved pollen (Barthelmes et al. 2006) particularly in conditions where sediment accumulation rates are very low (Waller and Schofield 2007). However, the dip in pollen concentrations in pollen zone CC14-B at 60 cm could be due to variation in accumulation rate. The Relevant Source Area of Pollen (RSAP) for the canopy-forming and field-layer vegetation for small depressions in woodlands are generally estimated as 50–100 m and < 10 m, respectively (Bradshaw 1981; Sugita 1994; Calcote 1995; Bunting et al. 2005). The inferences from this pollen diagram thus likely reflect the local environment rather than a regional pollen influx (Jacobson and Bradshaw 1981; Barthelmes et al. 2006).
Pollen trap and soil samples
There is a clear contrast within the modern samples between the pollen trap (CT2) and CS2 soil samples on the one hand, and the CS3 soil samples on the other (Fig. 6a, b). The CT/CS2 samples come from an exclosure located at the centre of a small stand of trees, whereas the CS3 samples come from a grazed area on the edge of the wooded area. The CT/CS2 samples have much higher AP values (60–72%) than the CS3 samples (26–38%). Fraxinus (21–67%), as well as Selaginella (0–29%), are relatively common in the CT/CS2 samples, whilst Alnus (0–23%), Quercus (1–6%) and Betula (0–3%) are also present. Plantago (0–1%) values are low in the exclosure samples. In contrast, the CS3 samples are dominated by Poaceae (47–61%), with small amounts of Alnus (0–3%), Betula (0–2%) and Quercus (1–4%) pollen. Plantago (1–3%) is somewhat more common. However, Pinus is more common in the soil samples (15–36%) than in the pollen trap samples (2–11%). Dung fungal values vary between the samples without a clear pattern (0–6%).

a Summary and b detailed percentage pollen data from pollen traps (CT2; S = summer, W = winter, the number referring to the years 2014–2016) and soil samples (CS2 in an exclosure and CS3 on the edge of the woodland), based on %TP excluding Cyperaceae and ferns; Cyperaceae values are based on %TP including Cyperaceae
Discussion
The pollen diagram from The Allers in Chillingham Wild Cattle Park presented here shows a similar woodland composition to the woodland assemblages previously documented for the Neolithic to Iron Age at other sites in northeast Northumberland, with Alnus, Betula, Corylus-type, and Quercus as the main taxa (Davies and Turner 1979). This is characteristic for the mixed oak woodlands of the area, which are reported to have contained larger proportions of Betula and Corylus than further south (Turner 1981). At Chillingham, AP values are initially around 50%, indicating a significant degree of woodland cover (Tinsley and Smith 1974; Birks 1988; Bunting 2002). Throughout the Neolithic and Bronze Age, small clearances are evidenced in pollen diagrams from Northumberland. However, in these diagrams, AP values remain around 35–60% (e.g. Chapman 1964; Turner 1968; Borek 1975; Mannion 1978; Davies and Turner 1979; Moyle 1980; Macklin et al. 1991; Hewitt et al. 1996). Agricultural indicators such as Plantago, Polygonum and Rumex appear in these clearance phases.
At Chillingham, the slight drop in AP and the presence of Plantago at 100 cm in the CC14 core (estimated age 2,731 cal bp in the Bacon model) may represent a similar Late Bronze-Early Iron Age local small-scale clearance. There is no evidence for the local cultivation of cereals, likely due to the relatively wet character of the site. However, this does not preclude the presence of crop cultivation in drier parts of the region. Whilst Plantago is present, dung fungi, though they represent a mostly local signal, are not abundant. Thus, these small clearances could potentially relate to arable or mixed farming. The NPP assemblages provide evidence of a more sustained shift in land use and increase in animal presence from 80 cm upwards (estimated age 2,569 cal bp in the Bacon model) that may represent an opening up of the tree canopy to give wood pasture, taking advantage of a slight drying out of the area. Thus, expanding the palynological analysis to include NPPs has the potential to identify human activities that cannot be inferred directly from the pollen assemblages.
Across Northumberland, AP values fall to much lower levels during later clearance phases, but there are only a few sites in northeast Northumberland where the first intensive woodland clearance phase is well-dated (Davies and Turner 1979; Dumayne-Peaty 1999; Passmore and Stevenson 2012).
At Callaly Moor, about 18 km south of Chillingham, a soil complex (unit CM-E) exposed in a stream-cut section was dated to 2,540 bp at its base (Macklin et al. 1991). The sediments in which the soil developed were thus likely deposited during the Iron Age. Whilst cereal-type pollen is fairly common in the underlying Bronze Age alluvial layers, AP values, comprising Alnus, Betula, Corylus-type and Quercus, are around 40%. These increase somewhat towards the bottom of the soil complex, then fall in the soil complex as heath spreads over the area. Cereal-type pollen remain present in small numbers, although the area is reported to have been abandoned during the Iron Age (Macklin et al. 1991).
Two thin peat beds were found underneath and near the base of a gravel fill on a terrace of the River Breamish near Powburn, about 10 km southeast of Chillingham. The peat bed of Sample 2, Locality 3 is dated to 2,743 and 2,356 cal bp, and its pollen evidences a damp woodland, but a peak in Plantago occurs in the middle of the deposit (Tipping 1992). A second peat bed (Sample 3, Locality 2) has three dates of 1,723, 2,334 and 1,946 cal bp. By this time, the landscape had opened up, with high values for Poaceae, Plantago and Rumex.
The pollen diagrams from Camp Hill Moss (~ 0.5 km east of the eastern boundary of Chillingham Wild Cattle Park) and Broad Moss (~ 10 km southwest of Chillingham) show many similarities (Davies and Turner 1979). A period of repeated small Early Bronze Age clearances is dated to between 3,510 ± 70 and 3,110 ± 80 bp at Camp Hill Moss. AP values remain around 70%, with Alnus, Corylus-type, Betula and Quercus as the main tree taxa, but Poaceae increase and Plantago and Rumex are recorded. This is followed by forest regeneration, with Betula playing a greater role than before. Further small-scale clearances are recorded between 2,670 ± 70 and 640 ± 80 bp at Camp Hill Moss. AP values drop to around 60% and Poaceae and Plantago values are higher than in the earlier clearance phase. A few cereal-type pollen appear in the record. Full clearance only takes place above this level. Davies and Turner (1979) consider the date to be erroneous, based on inconsistencies between the peat accumulation rate and climatic conditions during this period. They regard 640 ± 80 bp as an unlikely late date for full clearance, leaving open the possibility that full clearance took place during the later Iron Age or Roman period. Tipping (1992) reinterprets the diagram as containing a hiatus, possibly due to Medieval peat cutting. Thus, it is not clear at what date the first full clearance took place at Camp Hill Moss.
A similar clearance phase was recorded by Davies and Turner (1979) at Broad Moss, with a clear decrease in AP, increase in Poaceae, and peak in Plantago. Cereal-type pollen are only sparsely present. However, this clearance phase was initially only dated by correlation with the Camp Hill Moss clearance phase. Passmore and Stevenson (2012) cored the same location and dated a phase of major semi-permanent forest clearance to 2,360–2,150 cal bp, with significant decreases in the values for all tree types, as well as Corylus. However, even though this clearance phase occurs at the same depth as in Davies and Turner’s (1979) diagram, there are some differences, possibly due to the fact that they included local taxa (Cyperaceae and Ericales) in the pollen sum (Passmore and Stevenson 2012), whereas Davies and Turner (1979) did not. There is no clear increase in Poaceae and increases in pastoral indicators are small. Nevertheless, this is the closest dated core to that of Chillingham that provides evidence of clearance in the Middle Iron Age.
The first extensive clearance at Ford Moss (~ 15 km northwest of Chillingham) at 175 cm is estimated to date to ca. 2,170 bp (Davis and Passmore 2012). Although woodland cover had been reducing for some time, at this point the landscape becomes largely open. Many agricultural weeds are present, as well as small amounts of cereal-type pollen.
At Chillingham, by 2,403 cal bp (as estimated in the Bacon model at 60 cm), AP values had dropped to 27%, reaching a minimum of 13% at 50 cm (estimated age 2,315 cal bp), indicating the landscape was open in character. This provides evidence for significant local clearance during the late Early to Middle Iron Age, likely coinciding with the construction and occupancy of the nearby Ros Castle hillfort and predating any Roman influence in the region. Given the small size of the sedimentary basin, the concurrent rising frequency of Cyperaceae pollen suggests the local area became wetter, potentially as a result of reduced evapotranspiration that followed the removal of trees. However, as the values for Poaceae, as well as other open land taxa such as Plantago and Asteraceae also increase, the reduction in AP is unlikely to be due to increased wetness only. In contrast to the earlier, smaller-scale clearances, dung fungal spore values also increase substantially at this point, remaining high for most of the remainder of the sequence. This indicates that animal husbandry played a more important role in this clearance phase. The large decrease in dung fungi at 40 cm, concomitant with a small rise in AP, dated to 2,173 cal bp in the Bacon model, likewise indicates a decreasing impact of husbandry on the local landscape and limited woodland regeneration. Cunliffe (2005) documents a degree of abandonment of the uplands around this time, potentially influenced by a cooling climate from 2,250 bp onwards (as documented by Chiverrell et al. 2008; Blundell and Langdon 2023).
Further north, the extensive raised mire of Dogden Moss, about 45 km northwest of Chillingham, high percentages of Corylus-type pollen indicate that the local woodland had already been impacted by human land use, with pastoral indicators also present (Dumayne-Peaty 1999). However, the landscape remained wooded until the start of a rapid and intensive clearance phase that is dated to 2,520 cal bp. AP drops from 30 to 40 to 25% (or from 80 to 40% if Corylus-type and Salix are included). Avena-Triticum-type pollen and agricultural weeds provide evidence that arable agriculture was practiced, and grazing indicators continue to be present.
Taking a wider geographical view, evidence from some cores west and southwest of the Cheviot Hills indicates the landscape opened up during the Bronze Age. Drowning Flow became dominated by heather during the Bronze Age (Moores 1998). Increases in Pteridium from the early Iron Age onwards are interpreted as over-burning or overgrazing of heather moorland, while Plantago and cereal-type pollen are present from ca. 1,905–1,705 bp. Nearby Bloody Moss shows a similar change to heather moorland during the Bronze Age, followed by an increase of Poaceae, Plantago and the presence of cereal-type pollen (Moores 1998). Chronological control is imprecise, with the peak in agricultural indicators occurring halfway (ca. 350 cm) between a date of 3,235–2,850 cal bp at 421–428 cm and 1,375–960 cal bp at 221–228 cm.
More localised deposits from river valleys west of the Cheviot Hills evidence a different range of human activities. At Snabdaugh Farm, Terrace 5, near the confluence of the River North Tyne with the Chirdon and Tarset Burns, the landscape seems to have been substantially cleared before 2,710–2,160 cal bp, with high values for Poaceae and agricultural herbs (Moores 1998; Moores et al. 1999). Cereal-type pollen are also present. Two terraces of the River Rede near Brownchesters Farm provide evidence for an open landscape, with pollen assemblages from a core through Terrace 6 showing that Poaceae become dominant from 3,840 to 3,470 cal bp, with Plantago lanceolata and cereal-type pollen present (Moores 1998). However, there is no sedimentary record for the Iron Age from this site (Moores et al. 1999).
At other sites west of the Cheviot Hills, full clearance seems to have taken place at a later date, although there is uncertainty about the timing and whether this was a single event. At Yetholm Loch, tree pollen remains high until a band of coarse inwash sediments, tentatively dated to 2,200–1,700 cal bp disturbs the organic sediments (Tipping 1992). Above this layer, a fully cleared landscape is represented, but the date of the clearance is uncertain. Cereal-type pollen occur from ca. 2,250 bp at Sourhope, along with agricultural herbs (Tipping 1992). At Steng Moss, limited woodland openings took place around 2,586 ± 45 and 2,528 ± 35 bp, possibly reflecting shifting agriculture (Davies and Turner 1979). From 1,970 ± 60 bp there is a substantial rise in herbaceous pollen and cereals were grown.
In the area near Hadrian’s Wall, particularly in the middle section between Carlisle and Hexham, a number of pollen cores from nearby peat bogs and small lakes have been studied. Several analyses of cores from the area around Housesteads Roman Fort give somewhat contradictory results. The blanket mire of Sells Burn became dominated by heather around 2,995–2,775 cal bp (Moores 1998). There are sparse records of Plantago, whilst cereal-type pollen appear much higher up in the sequence. The small lake of Crag Lough records a brief clearance event at 480–490 cm with cereal-type pollen and agricultural weeds, dated to ca. 2,550 bp based on a radiocarbon date of 2,360–2,150 bp at 446–454 cm, with further and more lasting clearance around the time of the radiocarbon date (Dark 2005). More widespread clearance took place around 2,150 bp, although locally trees remained abundant. In contrast, a few kilometers further west, at Fozy Moss a sudden clearance event in a previously largely forested landscape with only small clearances is dated to around 1,825 bp (Dumayne 1993; Dumayne and Barber 1994). Likewise, in an undated core from Muckle Moss, the first major clearance is likely associated with the Roman occupation of the area (Pearson 1960).
Fellend Moss, near Haltwhistle, shows little evidence of land use until the Roman era, with the start of the main clearance event radiocarbon dated to 1,948 ± 45 bp (Davies and Turner 1979). At Butterburn Flow, 10 km further north, the forest slowly starts to open up from 2,250 cal bp (Yeloff et al. 2007). Dung fungi, and particularly Sordaria-type, are present from this point, along with peaks in Plantago, leading Yeloff et al. (2007) to suggest heavy grazing took place in the region. Cereal-type pollen and agricultural weeds are continuously present at low values. The main clearance takes place between 2,040 and 1,900 cal bp. At Walton Moss, northeast of Carlisle, the first but intense forest clearance, accompanied by cereal-type pollen and agricultural weeds, is dated to ca. 1,960 bp (Dumayne and Barber 1994).
There is strong evidence of a hiatus in deposition in the CC14 core around 30 cm. The assemblages recovered from the top 30 cm of the core are closely similar to the soil sample assemblages from the CS3 location (Fig. 6a, b), which is located on the edge of The Allers. AP values are similar (28% in the top sample of the core and 26–38% in the CS3 soil samples), with relatively low values for Alnus and Quercus compared with the higher values in the CT/CS samples. Poaceae and Plantago values are similar in the core and the CS3 samples. However, the relatively high values for Selaginella in the top of the core are more similar to the values in the CT/CS samples. The core assemblages show Fraxinus values (0–6%) that are intermediate between the two locations, in accordance with its position between them.
It is notable that Alnus percentages are low in the top 30 cm of the core and in most surface samples, even though the area is known to have been a patch of mixed ash and alder woodland for the past several centuries. The woodland was last coppiced around 1754 (Hall 2013). Due to heavy grazing pressure, there was effectively no regeneration of the coppiced stools until 1991, when the trees were protected (S. Hall, pers. comm., June 2023). Some regeneration has taken place since then. Coppiced alder protected from grazing tends to produce abundant catkins 5–6 years after coppicing (Waller et al. 2012). However, the older trees are at present senescent and do not produce many catkins, leading to low relative abundance in the surface samples, although occasional bursts of pollen production may take place in spring as evidenced in the pollen trap samples. Aged alder woodland may therefore be invisible in pollen records, with pollen sporadically dispersed from regenerating stems rather than the trees themselves. The presence of Pinus and small numbers of Picea is not surprising. Coniferous plantations have been planted in the region since the 1950s, and these taxa are prolific pollen producers whose pollen disperse over long distances.
It is striking that in the modern samples from Chillingham, AP drops off strongly at the edge of the woodland. This ‘woodland’ is better characterised as a small stand of trees with a diameter of ca. 150 m. Whilst pollen deposition in the centre of the canopy is dominated by tree pollen, the dominance of the trees in the pollen record does not extend far beyond the edge of the stand of trees, and can even be seen to drop off in the top of the core, which is located between the centre of the stand of trees and its edge. In samples taken from a small basin such as The Allers, smaller stands of trees present in the wider landscape may be invisible if the core is located outside of such stands. Caution should be exerted regarding the spatial scope of interpretation from such sites.
Conclusions
Whilst covering a limited temporal range, the new palaeoecological record presented here significantly increases our understanding of Iron Age forest clearance in northeast Northumberland. Forest cover and composition in the Late Bronze Age at Chillingham are similar to those recorded from other sites in the region. Although the locality was likely too wet for cereal cultivation, combining the presence of ruderal herbs associated with agriculture with the dung fungal spore record for small clearances during this period enables us to differentiate between clearance activities likely aimed at crop cultivation vs. animal husbandry, possibly with grazing under a tree canopy in a wood pasture.
The first widespread clearance event at Chillingham, with arboreal pollen falling to 13%, is dated to the late Early to Middle Iron Age, around 2,403 cal bp, with a woodland cover minimum around 2,315 cal bp. This likely coincides with the occupation of the nearby Ros Castle hillfort. A significant rise in dung fungal spores and the presence of Plantago and other open vegetation taxa indicate animal husbandry played an important role in this clearance phase, whilst the absence of cereal-type pollen may indicate cereal crops were not grown in the near vicinity. This somewhat post-dates the regional record at Dogden Moss, where full clearance, during which cereal-type pollen was recorded, dates to 2,520 cal bp (Dumayne-Peaty 1999). However, this record is located at some distance from Chillingham, across a major landscape boundary, the river Tweed. Moreover, the Chillingham clearance is similar in age to major clearance phases in geographically closer sites such as Powburn (predating 2,334 cal bp; Tipping 1992) and Broad Moss (2,360–2,150 cal bp; Passmore and Stevenson 2012), where there are pastoral indicators but cereal-type pollen are not recorded. At Chillingham, a subsequent large decrease in dung fungi and a small rise in AP around 2,173 cal bp may reflect a degree of abandonment of the uplands after the climate cooled from 2,250 bp onwards.
West of the Cheviot Hills, full clearance with cereal-type pollen present and/or the spread of heath largely predates the Iron Age (Drowning Flow, Bloody Moss, Snabdaugh Farm; Moores 1998; Moores et al. 1999). Where the main clearance phase is later (ca. 2,150–1,750 bp), there is clear evidence for arable agriculture (Yetholm Loch, Sourholm; Tipping 1992; Steng Moss; Davies and Turner 1979; Ford Moss may be similar; Davis and Passmore 2012).
The area near Hadrian’s Wall shows similar diversity, with pre-Iron Age woodland opening and the spread of heather recorded at Sells Burn (Moores 1998). Early to Middle Iron Age clearance took place at Crag Lough (Dark 2005) and Butterburn Flow (Yeloff et al. 2007) with evidence for both arable and pastoral agriculture, whilst other sites record first clearances in the latest Iron Age or early Roman period (Walton Moss; Dumayne and Barber 1994; Fellend Moss; Davies and Turner 1979; Fozy Moss; Dumayne 1993; Dumayne and Barber 1994).
Overall, it seems that full forest clearance took place at different times in different parts of Northumberland, with an emphasis on pastoral agriculture in some areas such as northeast Northumberland, and arable agriculture in other areas. Local records from pollen sites with small catchment areas are thus important for building a wider picture of the variation in land use across landscapes.
Peat deposition in the forest hollow ceased shortly before the arrival of Roman influence in the region, to resume only in the last century or so when we can trace the establishment of oak woods on the slopes surrounding the Park, and later coniferous plantations in the region. The open character of the Park and the influence of grazing animals is also clearly represented in this part of the core. Trees growing locally (in particular Alnus) are not well represented. The present alder woodland is senescent, indicating that aged alder woodland may be invisible in pollen sequences.
The core and its associated radiocarbon dates thus show that locally the main large-scale clearance event pre-dates the Roman period. Furthermore, including NPPs in the analysis allows us to elucidate human activities that cannot be inferred from the pollen assemblages alone.
Data availability
The microfossil data will be made available through the European Pollen Database.
References
Andersen ST (1979) Identification of wild grass and cereal pollen. Dan Geol Unders Årbog 1978:69–92
Barthelmes A, Prager A, Joosten H (2006) Palaeoecological analysis of Alnus wood peats with special attention to non-pollen palynomorphs. Rev Palaeobot Palynol 141:33–51. https://doi.org/10.1016/j.revpalbo.2006.04.002
Bartley DD (1966) Pollen analysis of some lake deposits near Bamburgh in Northumberland. New Phytol 65:141–156. https://doi.org/10.1111/j.1469-8137.1966.tb06347.x
Behre K-E (1981) The interpretation of anthropogenic indicators in pollen diagrams. Pollen Spores 23:225–245
Bell A (2005) An illustrated guide to the coprophilous ascomycetes of Australia. Fungal Biodiversity Centre, Utrecht
Bennett KD (1994) Annotated catalogue of pollen and pteridophyte spore types of the British Isles. Department of Plant Sciences, Cambridge
Bennett KD (1996) Determination of the number of zones in a biostratigraphical sequence. New Phytol 132:155–170. https://doi.org/10.1111/j.1469-8137.1996.tb04521.x
Birks HJB (1988) Long-term ecological change in the British uplands. In: Usher MB, Thompson DBA (eds) Ecological change in the uplands. Blackwell Scientific Publications, Oxford, pp 37–56
Blaauw M, Christen JA (2011) Flexible paleoclimate age-depth models using an autoregressive gamma process. Bayesian Anal 6:457–474. https://doi.org/10.1214/11-BA618
Blood K (1995) Ros castle iron age hillfort. Chillingham North Archaeol 12:35–38
Blundell A, Dearing JA, Boyle JF, Hannam JA (2009) Controlling factors for the spatial variability of soil magnetic susceptibility across England and Wales. Earth-Sci Rev 95:158–188. https://doi.org/10.1016/j.earscirev.2009.05.001
Blundell AC, Langdon PG (2023) A 3000-year multiproxy palaeoclimate record from Killorn Moss, Stirlingshire, Scotland. Mires Peat 29:06. https://doi.org/10.19189/MaP.2022.OMB.Sc.1836954
Borek MJE (1975) Pollen analysis and vegetational history of the ‘Akeld’ Basin. Durham University, Durham
Bradshaw RHW (1981) Modern pollen-representation factors for woods in south-east England. J Ecol 69:45–70. https://doi.org/10.2307/2259815
Bronk Ramsey C (2009) Bayesian analysis of radiocarbon dates. Radiocarbon 51:337–360. https://doi.org/10.1017/S0033822200033865
Bunting MJ (2002) Detecting woodland remnants in cultural landscapes: modern pollen deposition around small woodlands in Northwest Scotland. Holocene 12:291–301. https://doi.org/10.1191/0959683602hl545rp
Bunting MJ, Armitage R, Binney HA, Waller M (2005) Estimates of ‘relative pollen productivity’ and ‘relevant source area of pollen’ for major tree taxa in two Norfolk (UK) woodlands. Holocene 15:459–465. https://doi.org/10.1191/0959683605hl821rr
Calcote R (1995) Pollen source area and pollen productivity: evidence from forest hollows. J Ecol 83:591–602. https://doi.org/10.2307/2261627
Chambers FM, Jones MK (1984) Antiquity of rye in Britain. Antiquity 58:219–224
Chapman SB (1964) The ecology of Coom Rigg Moss, Northumberland: I. Stratigraphy and present vegetation. J Ecol 52:299–313. https://doi.org/10.2307/2257597
Chiverrell RC, Harvey AM, Hunter SY, Millington J, Richardson NJ (2008) Late Holocene environmental change in the Howgill Fells, Northwest England. Geomorphology 100:41–69. https://doi.org/10.1016/j.geomorph.2007.04.036
Clapperton CM, Durno SE, Squires RH (1971) Evidence for the flandrian history of the Wooler water, Northumberland, provided by pollen analysis. Scott Geogr Mag 87:14–20. https://doi.org/10.1080/00369227108736176
Cunliffe BW (2005) Iron Age communities in Britain. An account of England, Scotland and Wales from the seventh century BC until the roman conquest, 4th edn. Routledge, New York
Dark P (2005) Mid- to late-holocene vegetational and land-use change in the Hadrian’s Wall region: a radiocarbon-dated pollen sequence from Crag Lough, Northumberland, England. J Archaeol Sci 32:601–618. https://doi.org/10.1016/j.jas.2004.11.009
Davies AL, Dixon P (2007) Reading the pastoral landscape: palynological and historical evidence for the impacts of long-term grazing on Wether Hill, Ingram, Northumberland. Landsc Hist 29:35–45. https://doi.org/10.1080/01433768.2007.10594588
Davies G, Turner J (1979) Pollen diagrams from Northumberland. New Phytol 82:783–804. https://doi.org/10.1111/j.1469-8137.1979.tb01673.x
Davis BA, Passmore DG (2012) Pollen analysis and chronology at ford moss. In: Passmore DG, Waddington C (eds) Archaeology and environment in Northumberland. Oxbow Books, Oxford, pp 333–337
Dearing J (1999) Environmental magnetic susceptibility using the Bartington MS2 system, 2nd edn. Bartington Instruments, Oxford
Dickson C (1988) Distinguishing cereal from wild grass pollen: some limitations. Circaea 5:67–71
Dixon G, Gardner T, Gething P, Paterson D, Pedersen K, Tipping R, Young G (2015) Bamburgh research project. Bradford Kaims Wetland Heritage Project, Archaeological Report
Doveri F (2007) Fungi fimicoli Italici. Associazione Micologica Bresadola/Fondazione Centro Studio Micologici dell’A.M.B, Trento
Dumayne L (1993) Invader or native?—vegetation clearance in northern Britain during Romano-British time. Veget Hist Archaeobot 2:29–36. https://doi.org/10.1007/BF00191703
Dumayne L, Barber KE (1994) The impact of the romans on the environment of northern England: pollen data from three sites close to Hadrian’s Wall. Holocene 4:165–173. https://doi.org/10.1177/095968369400400206
Dumayne-Peaty L (1999) Late Holocene human impact on the vegetation of southeastern Scotland: a pollen diagram from Dogden Moss, Berwickshire. Rev Palaeobot Palynol 105:121–141. https://doi.org/10.1016/S0034-6667(98)00072-4
Etienne D, Jouffroy-Bapicot I (2014) Optimal counting limit for fungal spore abundance estimation using Sporormiella as a case study. Veget Hist Archaeobot 23:743–749. https://doi.org/10.1007/s00334-014-0439-1
Frodsham PNK (2004) Archaeology in Northumberland National Park. Council for British archaeology research reports 136. Council for British Archaeology, York
Gauthier E, Jouffroy-Bapicot I (2021) Detecting human impacts: non-pollen palynomorphs as proxies for human impact on the environment. In: Marret F, O’Keefe J, Osterloff P, Pound M, Shumilovskikh L (eds) Applications of non-pollen palynomorphs: from palaeoenvironmental reconstructions to biostratigraphy. Geological Society of London Special Publications, London, pp 233–244
Gill JL, McLauchlan KK, Skibbe AM, Goring S, Zirbel CR, Williams JW (2013) Linking abundances of the dung fungus sporormiella to the density of bison: implications for assessing grazing by megaherbivores in palaeorecords. J Ecol 101(1):125–1136. https://doi.org/10.1111/1365-2745.12130
Guarro J, Gené J, Stchigel AM, Figueras MJ (2012) Atlas of soil ascomycetes. CBS-KNAW Fungal Biodiversity Centre, Utrecht
Haas JN (2010) Fresh insights into the palaeoecological and palaeoclimatological value of Quaternary non-pollen palynomorphs. Veget Hist Archaeobot 19:389. https://doi.org/10.1007/s00334-010-0274-y
Hall SJG (1988) Chillingham Park and its herd of white cattle: relationships between vegetation classes and patterns of range use. J Appl Ecol 25:777–789. https://doi.org/10.2307/2403745
Hall SJG (2007) Chillingham wild cattle park, Northumberland. In: Rotherham ID (ed) The history, ecology and archaeology of medieval parks and parklands. Landscape archaeology and ecology 6. Wildtrack Publishing, Sheffield, pp 53–57
Hall SJG (2013) Integrated conservation of a park and its associated cattle herd: Chillingham Park, Northern England. In: Rotherham ID (ed) Trees, forested landscapes and grazing animals, a European perspective on woodlands and grazed treescapes. Routledge, Abingdon, pp 242–254
Hewitt I, Beckensall S, Gale J, Turner J, Nye S (1996) The excavation of Cairns at Blawearie, Old Bewick, Northumberland. Proc Prehist Soc 62:255–274. https://doi.org/10.1017/S0079497X00002802
Hibbert FA, Switsur VR (1976) Radiocarbon dating of flandrian pollen zones in Wales and Northern England. New Phytol 77:793–807. https://doi.org/10.1111/j.1469-8137.1976.tb04676.x
Hicks S, Tinsley H, Pardoe H, Cundill P (1999) European pollen monitoring programme, supplement to the guidelines. Oulu University Press, Oulu
Hua Q, Barbetti M, Rakowski AZ (2013) Atmospheric radiocarbon for the period 1950–2010. Radiocarbon. https://doi.org/10.2458/azu_js_rc.v55i2.16177.2059-2072
Jacobson GL Jr, Bradshaw RHW (1981) The selection of sites for paleovegetational studies. Quat Res 16:80–96. https://doi.org/10.1016/0033-5894(81)90129-0
Janssen CR (1959) Alnus as a disturbing factor in pollen diagrams. Acta Bot Neerl 8:55–58
Jobey G (1965) Hill forts and settlements in Northumberland. Archaeol Aeliana Ser 4:21–64
Jobey G (1968) Excavations of Cairns at Chatton Sandyford, Northumberland. Archaeol Aeliana Ser 4:5–50
Joly C, Barillé L, Barreau M, Mancheron A, Visset L (2007) Grain and annulus diameter as criteria for distinguishing pollen grains of cereals from wild grasses. Rev Palaeobot Palynol 146:221–233. https://doi.org/10.1016/j.revpalbo.2007.04.003
Juggins S (2020) Rioja: analysis of quaternary science data. https://CRAN.R-project.org/package=rioja
Kołaczek P, Zubek S, Błaszkowski J, Mleczko P, Margielewski W (2013) Erosion or plant succession – how to interpret the presence of arbuscular mycorrhizal fungi (Glomeromycota) spores in pollen profiles collected from mires. Rev Palaeobot Palynol 189:29–37. https://doi.org/10.1016/j.revpalbo.2012.11.006
Le Borgne E (1955) Susceptibilité magnétique anormal de sol superficiel. Ann Geophys 11:399–419
Macklin MG, Passmore DG, Stevenson AC, Cowley DC, Edwards DN, O’Brien CF (1991) Holocene alluviation and land-use change on Callaly Moor, Northumberland, England. J Quat Sci 6:225–232. https://doi.org/10.1002/jqs.3390060305
Mannion AM (1978) Late quaternary deposits from Linton Loch, Southeast Scotland I. Absolute and relative pollen analyses of limnic sediments. J Biogeogr 5:193–206. https://doi.org/10.2307/3038172
Moore PD, Evans AT, Chater M (1986) Palynological and stratigraphic evidence for hydrological changes in mires associated with human activity. In: Behre K-E (ed) Anthropogenic indicators in pollen diagrams. Balkema, Rotterdam, pp 209–220
Moore PD, Webb JA, Collinson ME (1991) Pollen analysis, 2nd edn. Blackwell Scientific, London
Moores AJ (1998) Palaeoenvironmental investigations of Holocene landscapes in the north Tyne basin, northern England. PhD thesis (unpublished), University of Newcastle-upon-Tyne
Moores AJ, Passmore DG, Stevenson AC (1999) High resolution palaeochannel records of Holocene Valley floor environments in the North Tyne basin, northern England. In: Brown AG, Quine TA (eds) Fluvial processes and environmental change. Wiley, Chichester, pp 283–310
Moyle DW (1980) Pollen analysis of peat deposits near Edlingham, Northumberland. University of Durham, Durham
Mullins CE (1977) Magnetic susceptibility of the soil and its significance in soil science – a review. Eur J Soil Sci 28:223–246. https://doi.org/10.1111/j.1365-2389.1977.tb02232.x
Oksanen J, Simpson GL, Blanchet FG et al (2022) vegan. Community ecology package. R package Version 2.6–4. https://CRAN.R-project.org/package=vegan
Oldfield F, Thompson R, Barber KE (1978) Changing atmospheric fallout of magnetic particles recorded in recent ombrotrophic peat sections. Science 199:679–680. https://doi.org/10.1126/science.199.4329.679-a
Passmore DG, Stevenson T (2012) Pollen analysis and chronology of broad moss. In: Passmore DG, Waddington C (eds) Archaeology and environment in Northumberland. Oxbow Books, Oxford, pp 338–341
Pearson MC (1960) Muckle Moss, Northumberland: I. Historical. J Ecol 48:647–666. https://doi.org/10.2307/2257340
Perrotti AG, van Asperen E (2019) Dung fungi as a proxy for megaherbivores: opportunities and limitations for archaeological applications. Veget Hist Archaeobot 28:93–104. https://doi.org/10.1007/s00334-018-0686-7
Perrotti AG, Ramiadantsoa T, O’Keefe J, Otaño NN (2022) Uncertainty in coprophilous fungal spore concentration estimates. Front Ecol Evol 10:1086109. https://doi.org/10.3389/fevo.2022.1086109
R Core Team (2021) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Reimer PJ, Austin WEN, Bard E et al (2020) The IntCal20 northern hemisphere radiocarbon age calibration curve (0–55 cal kBP). Radiocarbon 62:725–757. https://doi.org/10.1017/RDC.2020.41
Reimer PJ, Bard E, Bayliss A et al (2013) IntCal13 and Marine13 radiocarbon age calibration curves 0–50,000 years cal BP. Radiocarbon 55:1869–1887. https://doi.org/10.2458/azu_js_rc.55.16947
Stace CA (2019) New flora of the British Isles, 4th edn. C&M Floristics, Middlewood Green
Stockmarr J (1971) Tablets with spores used in absolute pollen analysis. Pollen Spores 13:615–621
Sugita S (1994) Pollen representation of vegetation in quaternary sediments: theory and method in patchy vegetation. J Ecol 82:881–897. https://doi.org/10.2307/2261452
Swindles GT (2010) Dating recent peat profiles using spheroidal carbonaceous particles (SCPs). Mires Peat 7:03
Swindles GT, Watson E, Turner TE, Galloway JM, Hadlari T, Wheeler J, Bacon KL (2015) Spheroidal carbonaceous particles are a defining stratigraphic marker for the Anthropocene. Sci Rep 5:10264. https://doi.org/10.1038/srep10264
Tinsley HM, Smith RT (1974) Surface pollen studies across a woodland/heath transition and their application in the interpretation of pollen diagrams. New Phytol 73:547–565. https://doi.org/10.1111/j.1469-8137.1974.tb02132.x
Tipping R (1992) The determination of cause in the generation of major prehistoric valley fills in the Cheviot Hills, Anglo-Scottish Border. In: Needham S, Macklin MG (eds) Alluvial archaeology in Britain. Oxbow Monograph 27. Oxbow Books, Oxford, pp 111–121
Tipping RM (1998) The chronology of late quaternary fluvial activity in part of the Milfield Basin, Northeast England. Earth Surf Process Landf 23:845–856
Topping P ((1990/1991)) The excavation of an unenclosed settlement, field system and cord rig cultivation at Linhope Burn, Northumberland, 1989. North Archaeol 11:1–42
Topping P (1989) Early cultivation in Northumberland and the Borders. Proc Prehist Soc 55:161–179. https://doi.org/10.1017/S0079497X00005375
Turner CH (1968) Studies on a small post-glacial peat deposit in Northumberland. University of Durham, Durham
Turner J (1981) The iron age. In: Simmons IG, Tooley MJ (eds) The environment in British prehistory. Cornell University Press, Ithaka, pp 250–281
Van Asperen EN, Kirby JR, Hunt CO (2016) The effect of preparation methods on dung fungal spores: implications for recognition of megafaunal populations. Rev Palaeobot Palynol 229:1–8. https://doi.org/10.1016/j.revpalbo.2016.02.004
Van Asperen EN, Kirby JR, Shaw HE (2020) Relating dung fungal spore influx rates to animal density in a temperate environment: implications for palaeoecological studies. Holocene 30:218–232. https://doi.org/10.1177/0959683619875804
Van Asperen EN, Perrotti A, Baker A (2021) Coprophilous fungal spores: non-pollen palynomorphs for the study of past megaherbivores. In: Marret F, O’Keefe J, Osterloff P, Pound M, Shumilovskikh L (eds) Applications of non-pollen palynomorphs: from palaeoenvironmental reconstructions to biostratigraphy. Geological Society of London Special Publications, London, pp 245–267
Van Geel B (2006) Quaternary non-pollen palynomorphs’ deserve our attention! Rev Palaeobot Palynol. https://doi.org/10.1016/j.revpalbo.2006.04.001
Van Geel B, Buurman J, Brinkkemper O, Schelvis J, Aptroot A, van Reenen G, Hakbijl T (2003) Environmental reconstruction of a roman period settlement site in Uitgeest (the Netherlands) with special reference to coprophilous fungi. J Archaeol Sci 30:873–883. https://doi.org/10.1016/S0305-4403(02)00265-0
Van Geel B, Coope GR, Van Der Hammen T (1989) Palaeoecology and stratigraphy of the lateglacial type section at Usselo (the Netherlands). Rev Palaeobot Palynol 60:25–129. https://doi.org/10.1016/0034-6667(89)90072-9
Waller M, Grant MJ, Bunting MJ (2012) Modern pollen studies from coppiced woodlands and their implications for the detection of woodland management in Holocene pollen records. Rev Palaeobot Palynol 187:11–28. https://doi.org/10.1016/j.revpalbo.2012.08.008
Waller MP, Schofield JE (2007) Mid to late holocene vegetation and land use history in the Weald of south-eastern England: multiple pollen profiles from the Rye area. Veget Hist Archaeobot 16:367–384. https://doi.org/10.1007/s00334-006-0042-1
Yeloff D, Charman D, Van Geel B, Mauquoy D (2007) Reconstruction of hydrology, vegetation and past climate change in bogs using fungal microfossils. Rev Palaeobot Palynol 146:102–145. https://doi.org/10.1016/j.revpalbo.2007.03.001
Acknowledgements
This research was funded through a Leverhulme Trust Early Career Fellowship (ECF-2013-517, van Asperen). The authors wish to thank the Chillingham Wild Cattle Association and Park staff who contributed greatly to this study by facilitating the fieldwork. We thank Stephen Hall for providing information about the history of the Park. Hazel Clark supported the magnetic susceptibility measurements. Two anonymous reviewers and the editor, Michael O’Connell, provided useful comments that helped us to improve the manuscript. For the purpose of open access, the corresponding author has applied a Creative Commons Attribution (CC BY) licence to any Author Accepted Manuscript version arising from this submission.
Author information
Authors and Affiliations
Contributions
EA and JK contributed to the study conception and design. Data were collected by EA, analysis was performed by EA, JK and HS. The first draft of the manuscript was written by EA and all authors contributed to the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no conflict of interest to declare that are relevant to the content of this article.
Additional information
Communicated by M. O’Connell.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
van Asperen, E.N., Kirby, J.R. & Shaw, H.E. Multi-proxy evidence for woodland clearance in northeast Northumberland (England) during the Iron Age. Veget Hist Archaeobot (2023). https://doi.org/10.1007/s00334-023-00979-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00334-023-00979-1