
Abstract
Current knowledge of the origins and routes of introduction of both tetraploid and hexaploid free-threshing wheats (FTWs) from western Asia into Europe remains imprecise. Archaeobotanical distinction of ploidy level is often dependent on sparsely recovered rachis segments, while more specific identification of cultivars within ploidy groups using morphological characteristics of either grain or chaff is generally considered unreliable. This study offers a complementary approach by using geometric morphometric (GMM) analysis of grain shape to assess taxonomic and variety level distinctions in archaeobotanical FTW remains. Two substantial and well-preserved assemblages of Neolithic FTW grains from 7th millennium bce Çatalhöyük (central Anatolia, Turkey) and 6th millennium Kouphovouno (Peloponnese, Greece) were analysed with the aim of shedding light on the naked wheats cultivated by these early agricultural communities. While chaff remains from Çatalhöyük indicate a hexaploid FTW crop, no such diagnostic remains were recovered from Kouphovouno. GMM analysis of the archaeobotanical grains has corroborated early cultivation of hexaploid naked wheat at Çatalhöyük, whilst revealing that a tetraploid wheat, most closely resembling ‘unimproved’ durum wheat, was likely grown at Kouphovouno. The Kouphovouno findings are contextualised within existing theories of a Mediterranean route for introduction of tetraploid FTWs from western Asia to south-western Europe. At both sites, an assessment of the role of FTWs over time suggests changes in the cultural value attributed to these crops and/or the sustainability of the agricultural regimes they were associated with.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Types of free-threshing wheat and their identification in the archaeobotanical record
The ears of free-threshing wheats (FTWs) have tough rachises and thin glumes, from which ‘naked’ grains are released upon threshing. In contrast, glumed (hulled) wheats have tough glumes and, when threshed, break up into spikelets, which require further processing to release the grain (Fig. 1; Hillman 1984). FTWs are by far the most commercially significant group today, with Triticum aestivum L. ssp. aestivum (bread wheat) accounting for as much as 90–95% of the global wheat market and T. turgidum L. ssp. durum (hard or durum wheat) primarily used for pasta, the second-highest produced (Giraldo et al. 2019).

Schematic representation of crop processing sequences for glume and free-threshing wheats, adapted from an illustration by Kathryn Killackey
Durum and bread wheats represent two ploidy groups, respectively, the tetraploid turgidum group and the hexaploid aestivum group. Tetraploid wheats have four sets of chromosomes (BBAA) while hexaploid wheats have six (BBAADD). The latter evolved from tetraploid wheats via polyploidisation, the result of intergeneric hybridisation with the wild grass Aegilops (Fig. 2). Allopolyploidy (having chromosomes from two or more taxa) is linked to greater adaptive potential, as genomes previously adapted to different environments converge within a single organism, and this has doubtless played a role in the near-global predominance of hexaploid bread wheat today (Dubcovsky and Dvorak 2007).

Schematic diagram of the evolution of durum, bread and club wheats, adapted from Pont et al. (2019)
Within these ploidy groups is a range of accepted sub-species of FTW with distinct distributions, culinary properties and growing requirements (Table 1). We use here the widely adopted taxonomy of van Slageren (1994), but alternative taxonomies consider these wheat types sufficiently distinct to categorise them as different species (Goncharov 2011). Beyond the accepted (sub)species, a much wider array of named varieties with different morphological and adaptive characteristics are documented in recent history (Percival 1921). Yet, despite this diversity, charred remains in archaeobotanical contexts are often identified with limited precision as ‘free-threshing’, ‘turgidum-aestivum’ or ‘aestivo-compactum’ types. It is possible to distinguish between tetraploid and hexaploid FTW remains based on diagnostic criteria of the rachis segments, although it is noted that variations in rachis morphology along the ear can complicate such identification (Hillman et al. 1996; Hillman 2001). Furthermore, due to the greater susceptibility of rachis segments to destruction during charring (Boardman and Jones 1990) and an apparent tendency for FTWs to be processed in bulk immediately after harvesting and away from domestic charring contexts (Jones 1987), these diagnostic elements are often not preserved in the archaeobotanical record, or present only in small quantities.
The charred grain of FTW is more commonly recovered than the chaff, but has proven problematic to identify reliably to ploidy level. While possible distinguishing criteria have been noted, such as the depth of the ventral furrow relative to the height of the grain, the same sources acknowledge a lack of consistency in these characteristics (for example, Moffett 1987; Hillman et al. 1996). Analysis of ancient DNA extracted from grains presents an alternative approach (for example, Schlumbaum et al. 1998; Fernández et al. 2013; Bilgic et al. 2016) but issues of preservation and contamination (as well as cost) inhibit widespread application. As regards distinguishing wheat remains within ploidy levels to (sub)species level, there is an even less established basis for separation using either grain or rachis segment characters (Jones 1998; Hillman et al. 1996, p. 204).
Overall, there are significant limitations to traditional morphological approaches when identifying archaeobotanical FTW remains. Geometric morphometric (GMM) analysis of grain shape is therefore offered here as an approach that complements and extends traditional morphological identification of grain (beyond ‘free-threshing wheat’) and rachis (beyond ploidy level). The application of GMM to archaeobotanical research has flourished in recent years and there is much scope for its further development and application in the field (Portillo et al. 2020; Evin et al. 2022), to which this study seeks to contribute.
Early archaeobotanical evidence for free-threshing wheats, potential origins and histories
The earliest cultivated wheats were the glume wheats Triticum monococcum (einkorn) and T. dicoccum (emmer), domesticated forms of which emerged in the ‘Fertile Crescent’ of south-west Asia from the early Pre-Pottery Neolithic B, around 8700−8200 cal bce (Arranz-Otaegui et al. 2016). Genetic sequencing suggests that the earliest FTWs were tetraploids which evolved from emmer wheat, with hexaploid wheats subsequently originating from hybridisation between a tetraploid FTW and the wild grass Aegilops tauschii Coss. or a lineage close to it (Fig. 2; Dvorak et al. 2012; Pont et al. 2019). It has been proposed that this tetraploid FTW was a durum (Pont et al. 2019), although parentage by a now-extinct ancestral form has also been suggested (Levy and Feldman 2022). It has also been proposed that the offspring of this hybridisation would have initially been hulled (Dvorak et al. 2012), but this form has yet to be identified archaeobotanically. The earliest recorded FTW remains have little chronological separation from the earliest domesticated glume wheats, with free-threshing examples of (as yet) unknown ploidy level identified from the second half of the 9th millennium bce at Aşıklı Höyük, Turkey (Ergun et al. 2018) and the late 9th to early 8th millennium bce Tell Aswad, Syria (van Zeist and Bakker-Heeres 1982; Zohary et al. 2012, p. 45).
It has been proposed that, based on the main distribution area of Aegilops tauschii (syn. A. squarrosa) (Zohary 1969; van Zeist 1976) and links with specific gene pools (Dvorak et al. 1998), hexaploid wheat could only have developed after tetraploid FTW spread to Transcaucasia and Caspian Iran in the 6th millennium bce. However, hexaploid FTW is reported from as early as the late 9th to mid 8th millennium cal bce at Cafer Höyük, south-east Turkey (de Moulins 1993, 1997; radiocarbon dates corrected after The Platform for Neolithic Radiocarbon Dates, Ex Oriente 2022) and from the late 8th to early 7th millennium cal bce at Çatalhöyük, south-central Turkey (Bogaard et al. 2013, 2017, 2021). Tetraploid and hexaploid FTW remains are also recorded from 8th millennium cal bce Abu Hureyra, Syria (de Moulins 1997; Ex Oriente 2022), the first half of the 7th millennium cal bce at El-Kowm, Syria (de Moulins 1997; Borrell et al. 2011) and from the mid to late 8th millennium cal bce at Can Hasan III, near Çatalhöyük (Hillman 1972, 1978; Fairbairn et al. 2020). Notably, at Can Hasan III tetraploid rachis is only found in the latest 2 phases of occupation, while hexaploid rachis is present throughout (Hillman 1978, p. 168); however recent radiocarbon dating has shown the high potential for redeposition of material within this sequence (Fairbairn et al. 2020, p. 347). In line with this evidence, Giles and Brown (2006) propose an origin of hexaploid FTW in southeast Turkey or northern Syria, arguing that the genetic evidence theoretically supports a scenario where ‘peripheral’ communities of Aegilops tauschii first came into contact with tetraploid FTW in these regions, with a second hexaploidisation possibly occurring in Iran.
Cultivation of FTW then spread east and westwards, reaching China by the 3rd millennium bce (Liu et al. 2016) and Britain by the early 4th millennium bce (Jones and Rowley-Conwy 2007; Carruthers 2019). A cache of grains found in an aceramic Neolithic level at Knossos, Crete, indicates that FTW reached the Mediterranean region by the earlier 7th millennium bce (Sarpaki 2013; Douka et al. 2017). However, while identified as bread wheat in the original report (Helbæk 1968, cited in Sarpaki 2013), the ploidy level of these remains has not been conclusively demonstrated (Sarpaki 2013). Elsewhere in the region, tetraploid rachis remains are recorded for the later 6th millennium bce Cardial lakeshore settlements of La Marmotta, central Italy and La Draga, Catalonia (Jacomet 2007, p. 246; Rottoli and Pessina 2007; Antolín 2016). The earliest written evidence for Europe suggests that, certainly by Classical Antiquity (8th century bce to 5th century CE), tetraploid durum and to a lesser extent Triticum turgidum (rivet wheat) were the predominant wheat crops grown in the Mediterranean (Jasny 1944).
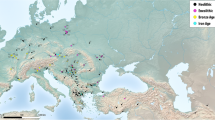
Historically, it was believed that the FTW of Neolithic Europe was a compact form of hexaploid wheat equated with modern-day Triticum compactum (club wheat) (Percival 1921). However, studies of exceptionally well preserved whole ears by Maier (1996) demonstrated cultivation of tetraploid wheats at the early fourth millennium bce lake shore settlement of Hornstaad-Hörnle IA, south-west Germany. The author therefore proposed a Mediterranean route of introduction for tetraploid wheats to south-west Europe, reaching the northern foothills of the Alps ~ 4000 bce. This theory was developed further by Kirleis and Fischer (2014), who defined two agricultural traditions using archaeobotanical data from central and northern Europe. The first relates to predominantly hexaploid FTW cultivation by the late Linearbandkeramik culture, associated with the spread of Neolithic culture from the east through the Balkans and Carpathian basin. The second, predominantly tetraploid, tradition was associated with early Neolithic groups in the northern Alpine foothills, arriving from the Mediterranean and southern France. This tradition subsequently diffused north via contacts between the Michelsberg and Trichterbecher cultures of northern Germany and southern Scandinavia. A review of (published) identified rachis segments would appear to support the theory of a more northerly hexaploid and more southerly tetraploid route into western Europe (Fig. 3).

Distribution map of identified free threshing wheat rachis segments from Neolithic sites in Europe and western Asia. Arrows show potential introduction routes of hexaploid and tetraploid FTW. Site names and references as in ESM 1; Çatalhöyük and Kouphovouno, archaeological sites included in this GMM study
Overall, while the understanding of the origins and introduction routes of FTWs across western Asia and into Europe is improving, much remains unclear. It is notable that, despite our expectation that the evolution of tetraploid FTWs predated hexaploid FTWs, there is a lack of securely identified archaeobotanical evidence to confirm this. The morphological ambiguity reported for certain early FTW rachis finds as from Aşıklı Höyük (Ergun, personal communication) and Bulgaria (Marinova 2007, p. 102) may relate to transitional stages in the evolutionary development of FTW that are yet to be defined. While our knowledge may be improved by the recovery of further archaeobotanical data (or reviewing of existing material), geometric morphometrics (GMM) may also help to refine this picture.
The archaeobotanical material
To assess the potential of GMM in relation to early FTW cultivation, material was selected from 2 sites with unusually substantial and well-preserved assemblages of Neolithic FTW grains, Çatalhöyük in central Anatolia and Kouphovouno in southern Greece (Fig. 3). Çatalhöyük represents a notably early assemblage of FTW in the region where it potentially first evolved, while Kouphovouno is situated along the key Mediterranean route through which the spread of tetraploid FTW is hypothesised (Fig. 3; Maier 1996; Kirleis and Fischer 2014).
Çatalhöyük
Çatalhöyük is the source of one of the largest and most well-studied archaeobotanical assemblages relating to an early farming community in western Asia. The tell site is one of a number of densely occupied ‘megasites’ that emerged in the near East during the latter half of the Neolithic (Der and Issavi 2017). The inhabitants used a broad spectrum of crops and other food plants throughout the occupation sequence (Bogaard et al. 2017, 2021).
The grains examined from Çatalhöyük derive from the Neolithic East Mound (7100− 5950 cal bce), mostly from the Middle phase (6700 − 6500 cal bce) and the majority are preserved by charring in situ (ESM 2). While initial studies suggested the potential presence of tetraploid FTW (Fairbairn et al. 2002), subsequent work identified the rachis segments as hexaploid (Fig. 4; Bogaard et al. 2013, 2017). This identification was supported by ancient DNA (aDNA) analysis of the grains which demonstrated a sequence similar to modern hexaploid wheats (Bilgic et al. 2016). While the proportion of FTW in relation to other cereals does not follow a clear increase with time at Çatalhöyük, there is a shift towards free-threshing cereals, especially naked Hordeum (barley), between early and late phases, perhaps indicating an increasing preference over time for crops requiring less labour-intensive processing (Bogaard et al. 2017, 2021). However, in the Late-Final Neolithic East Mound and Chalcolithic West Mound phases we see a more even balance between hulled and free-threshing cereals (Bogaard et al. 2021).

Charred hexaploid free threshing wheat rachis from Middle Neolithic layers at Çatalhöyük. a rachis segments from context 11,923, illustrated by Kathryn Killackey; scale bar, 2.5 mm. b photograph of rachis segments from context 6151; scale bar, 1 mm
Palaeoenvironmental studies suggest a landscape in which small low energy channels flowed across the alluvial plain, creating a mosaic of both dry and wetland environments within the site catchment (Ayala et al. 2021, 2022; Hodder 2021; Veropoulidou 2021). Isotopic analysis indicates that wheat was generally grown under better watered conditions than barley (Wallace et al. 2015) but that growing conditions were highly variable even within the storage deposits of individual burnt buildings (Vaiglova 2016; Stroud et al. 2021). Analysis of the arable weed flora supports this ‘mosaic’ of varied growing environments, but also identified a single FTW deposit associated with a flora indicative of particularly high input management, comparable to modern plots which are regularly irrigated, tilled, weeded and manured (Green et al. 2018).
Overall, FTW was a staple crop of the Neolithic East Mound settlement, the importance of which may have increased over time as free-threshing cereals were increasingly favoured. While grown under a range of conditions, it seems likely that FTW would have been sown in better-watered areas of the landscape, at least in comparison to barley and, at least in some circumstances, was considered worth the investment of high input management.
Kouphovouno
Kouphovouno is a tell site located in the southern Peloponnese, on the outskirts of modern Sparta, Greece. While occupied intermittently for much of the prehistoric period, the analysed remains date to the Middle and Late Neolithic settlement (~ 5800–5000 bce), an area extending over approximately 4–5 ha (Cavanagh et al. 2017). Archaeobotanically, southern Greece is relatively poorly represented for this period (Livarda 2014), and the site therefore presents a valuable opportunity to study plant husbandry in the region.
While hulled barley was the most ubiquitous crop at the site, FTW was also well represented. The majority of FTW remains came from 10 grain-rich units interpreted as in situ charring of grain stores in domestic contexts. The largest proportion was associated with ‘Structure CIV’, a burnt building dating to the Middle Neolithic period. The assemblage is generally poor in chaff and no rachis internodes which could be identified as either tetraploid or hexaploid wheat were found. Previous archaeobotanical findings suggest a geographical divide in wheat preference during the Neolithic in Greece, with einkorn more common in the north, while emmer and FTW were predominant in the south (Livarda 2014). However, at Kouphovouno, einkorn appears to replace FTW as the major wheat type during the Late Neolithic.
Analysis of weed seeds associated with crops indicates that arable plots were long established, autumn sown and tended towards high soil fertility, suggesting a high input regime, as mentioned above. Isotopic analysis indicates that FTW was preferentially manured in comparison to hulled barley and einkorn, with pulse crops also grown under high-input regimes (Vaiglova et al. 2014, 2020). Comparison with faunal isotopic values suggests that FTW was reserved exclusively for human consumption (Vaiglova et al. 2014). These findings represent a system in which crops for human consumption, including FTW and pulses, were intensively managed under rotation in plots close to settlements, while potential fodder crops such as barley were grown more extensively further away (Vaiglova et al. 2014). As settlement size decreased by the Late Neolithic, emphasis apparently shifted towards the less intensively managed einkorn. This seems to have occurred in the context of reduced cooperation between households, as evidenced in the reduction of communal spaces and changes in pottery styles (Mee et al. 2014; Renard and Cavanagh 2017).
Materials and methods
Approach
In order to gain insight into the types of free threshing wheats cultivated by early farming communities at Çatalhöyük and Kouphovouno, elliptic Fourier analysis was done on modern and archaeological grain specimens. The first stage of analysis sought to establish the degree to which different types of modern wheat could be distinguished by the shape of the grain, while the second stage compared archaeological specimens with this modern material.
Sampling and photography
Modern reference material was obtained from gene banks (Table 2). Modern Triticum aestivum (bread wheat), T. compactum (club wheat), T. durum (durum wheat) and T. turgidum (rivet wheat) were selected for the main study as crops grown in Europe and western Asia with potentially prehistoric provenance.
A more limited examination was also made of T. turgidum ssp. carthlicum (‘Persian’ wheat) (Table 1). This is believed to result from hybridisation between a form of T. aestivum (proposed as ssp. carthlicoides) with domesticated T. dicoccum (emmer) (Kuckuck 1979; Haque et al. 2011; Oliveira et al. 2020). ‘Persian’ wheat is distinct in being genetically tetraploid, but shares features of rachis morphology with hexaploid wheats (Hillman 2001). When a single accession of T. carthlicum was compared with modern and archaeological material, the charred grains were noted to be longer and narrower than any of the study material, and of quite distinct proportions overall. Therefore while initial examination would not suggest this to be a morphological match for the material studied here, it would merit further study given its particular history and morphology.
The selection of material for this study prioritised availability of whole ears as well as close geographical origin to the study sites where possible, except T. turgidum (rivet wheat) because of limited material. This resulted in a combination of accessions classified as ‘landraces’ and a smaller number of modern ‘advanced’ or ‘improved’ cultivars. The grains were sampled equally from the base, middle and top of each ear to account for variation within the ear (nine grains per ear), with three ears sampled per accession. Archaeological material was assessed for inclusion on the basis of good preservation, while grains with significant distortion or damage were excluded. Examples of severe distortion from charring include the exudation of endosperm to form protrusions and cavities within the grain (Charles et al. 2015). Grains showing mechanical damage or with minimal survival of the outer pericarp were also excluded.
The grains were photographed using a Nikon SMZ25 stereo microscope with an apochromatic optical system and Pixelink M12BC-CYL microscopy camera. Initially grains were photographed mostly at 10× in dorsal, lateral and polar views (Fig. 5). However, once it was established that the polar view was of limited value in the analysis of wheat grains (Roushannafas et al. 2022), only dorsal and lateral views were taken.

Dorsal, lateral and polar views of a free threshing wheat grain with alignment points shown as +
Charring
Charring of modern reference material was done to replicate well-preserved archaeological material using parameters established in Charles et al. (2015). The grains were wrapped in foil and placed in beakers of sand to reduce oxygen availability and then charred in a preheated oven at 230 °C for 6 h. The temperature in both the oven and beakers was monitored using a data logger.
Digitisation
The outlines of the grains were traced using AutoCAD 2020. Each outline was represented by 40–80 individually placed reference points. While the number of points should ideally be higher and less variable, previous experimentation using more points found that this had a minimal effect on reclassification (Roushannafas et al. 2022). Elliptic Fourier analysis, as used here, is advantageous in not requiring points to be evenly spaced (Bonhomme et al. 2014). Alignment points were added to minimise differences in orientation between the grains (Fig. 5) and the coordinates of each outline were then extracted.
Analysis
Elliptic Fourier analysis of grain shapes was used (Giardina and Kuhl 1977; Kuhl and Giardina 1982), a type of outline analysis appropriate to ‘continuous’ shapes (Bonhomme et al. 2014). In this approach, outlines are decomposed into Fourier Series or harmonic sums of trigonometric functions weighted with harmonic coefficients (Bonhomme et al. 2014). The analysis used the R v. 3.6.2 environment and the packages geomorph v. 3.6.3 and Momocs v. 1.2.9 toolkit for 2D morphometrics, the latter developed by Vincent Bonhomme based on functions published by Claude (2008). The data from the grains were imported into Momocs as coordinates and then scaled by centroid size in order to emphasise shape over dimension. Fourier transforms were calculated for the outlines of each grain individually and then combined (Bonhomme et al. 2017). Eight harmonics were retained for each view, gathering at least 99% of the total harmonic power.
The results were mainly obtained with linear discriminant analysis (LDA) with leave-one-out cross-validation. This was used to test the degree to which grains could be correctly reclassified to type based on their shapes, scoring each known specimen (‘actual’) against the group it most closely resembled according to the model (‘classified’). LDA was also used to reclassify unknown archaeobotanical grains into the group(s) from known modern material with the closest morphological affinity. When plotted, the LDA visually represents the relationships between different morphological types, with similar groups plotting more closely together, and anomalous types occupying more peripheral positions, according to their relationship to the main groups.
Repeatability and measurement error
The repeatability of the measuring method was assessed after Evin et al. (2020), in which five grains from the same ear were each photographed in dorsal and lateral views and digitised in five separate sessions. Procrustes ANOVA regression for shape variables on the PCA scores for these outlines produced a non-significant value (≥ 0.05) when a particular session was used as the dependent variable, and a significant value when specimen identity was used as the dependent variable. This suggested that when the same grain was photographed, digitised and aligned many times, these different measuring sessions did not produce significantly different outlines, and that any variation between different sessions was less significant than the variation between the specimens.
Measurement error was calculated using the results of ANOVA after the equation defined in Claude (2008, pp 65–66) as ‘the ratio of the within-measurement component of variance on the sum of the within- and among-measurement component’:
Using this equation, the measurement error for the dorsal view was calculated at 4.38% and for the lateral view at 5.94%.
Results
Stage 1: modern reference material study
The first stage of analysis sought to establish whether different ploidy levels and (sub)species of FTW could be reliably separated by their grain morphology. Charred modern specimens of bread wheat (hereafter ‘aestivum’), club wheat (‘compactum’), durum wheat (‘durum’) and rivet wheat (‘turgidum’) were analysed (Table 2).
An LDA of all 324 modern grains correctly reclassified 89.8% of grains to ploidy level and 86.1% to (sub)species, when using the dorsal and lateral views (Tables 3 and 4). When plotted, the greatest overlap is observed between aestivum and compactum when using the first two linear discriminant functions (LD1 and 2), but these can be seen to separate along the third discriminant function (LD3) (Fig. 6). When accession was used as a discriminating factor, the reclassification rate was only slightly lower at 82.1% (Table 3), suggesting that the model is sensitive to varietal differences. When all ‘improved/advanced’ cultivars were removed from the model dataset, correct reclassification by taxon remained similar, at 85.6% (Table 3). The results suggest that at the group, if not individual, level, there are predictable morphological differences between grains of different taxonomic status and that these distinctions are preserved despite distortion from charring.

Linear discriminant analysis (LDA) results from comparing charred modern free threshing wheat (sub)species based on grain shape. Tcom T. compactum; Taes T. aestivum; Tdur T. durum; Tturg T. turgidum. Inset shows separation along third discriminant function LD3 when viewed as a 3D plot
Stage 2a: comparison between sub-species and archaeobotanical material
The next stage of analysis compared archaeobotanical grains from Kouphovouno and Çatalhöyük with the modern reference material. Eighty well preserved grains from each site were selected from a range of deposits (ESM 2). Archaeological grains were compared with both the full dataset and ‘unimproved’ subset, with the strongest association observed with the grains from the latter group. When plotted against the unimproved varieties there is significant overlap between the Çatalhöyük and both the compactum and aestivum grains (Fig. 7), and between the Kouphovouno material and the durum group (Fig. 8).

Linear discriminant analysis comparing Çatalhöyük (‘Catal’) material with modern landraces or traditional varieties. Taes T. aestivum; Tcom T. compactum; Tdur durum; Tturg T. turgidum

Linear discriminant analysis comparing Kouphovouno (‘Kouph’) material with modern landraces or traditional varieties. Taes T. aestivum; Tcom T. compactum; Tdur T. durum; Tturg T. turgidum
When the archaeological grains were entered as unknowns for reclassification against unimproved varieties, 82.6% of Çatalhöyük grains classified as either aestivum or compactum and 15.0% as durum (Table 5). From this we may infer that the predominant free threshing wheat there was a form most closely resembling modern hexaploid wheats. Since, based on the Stage 1 results, we would expect at least 14% of the grains to be classified incorrectly, we cannot say whether grains classified as durum represent a minority crop or merely atypical grains. The same analysis on the Kouphovouno material classified 81.3% as durum and 17.5% as either aestivum or compactum (Table 5). We may infer therefore that the main FTW crop resembled a tetraploid wheat most similar to durum, although we cannot rule out a hexaploid wheat having been present. Pairwise comparisons of mean shapes (Fig. 9) reinforced results of the LDA, with the Çatalhöyük material bearing the strongest resemblance to hexaploid wheat (particularly aestivum) and the Kouphovouno grains to durum. Procrustes ANOVA analysis of PCA scores found significant differences (p = 0.001) in shape between the Çatalhöyük and Kouphovouno assemblages in both dorsal and lateral views.

Pairwise comparisons of mean shapes of grains including archaeological material, in dorsal and lateral views. Catal, Çatalhöyük; Kouph, Kouphovouno; Taes T. aestivum; Tcom T. compactum; Tdur T. durum: Tturg T. turgidum
Stage 2b: comparisons of varieties within sub-species with archaeological material
Having established a potentially closer association between the archaeological material and traditional varieties of modern wheat, the next stage was to look more closely at variation within ploidy and (sub)species in comparison to the archaeological material.
The Çatalhöyük material was compared with five aestivum and three compactum accessions from Europe and western Asia (Table 2). An LDA of these eight modern accessions correctly classified 95.8% of grains to (sub)species. However, when the archaeological grains were classified against these modern accessions they showed no clear association with either (sub)species or accession (Table 6). It is notable that accessions which had been collected furthest away from the archaeological sites in terms of latitude (England and Serbia) appear to be the least similar to the Çatalhöyük material, in which higher proportions of grains classified as traditional varieties from central and western Asia. Similarly, when the Kouphovouno grains were compared with modern durum accessions, the highest proportions of grains were classified as ‘traditional’ accessions from North Macedonia, and to a lesser extent Cyprus, while the grains least resembled ‘improved’ varieties from Turkey and France (Table 7).
These results suggest a relationship between geographical origin, improvement status and grain shape, although further work would be needed to elaborate this. We cannot draw firm conclusions on the basis of this fairly limited range, particularly without more detailed collection data. Nevertheless, it is possible that regional variations in grain shape have persisted from the earliest stages of farming to the present day. Even if this suggestion remains tentative, our results show the importance of comparing archaeological grains with modern reference material from geographically close locations, and these ideally being traditional landraces.
The suggestion of geographical variation naturally gives rise to the question of how the growing environment might influence grain shape. GMM analysis of grains grown under different agricultural regimes is currently underway to address this. Preliminary results from analysis of a single accession of Sham4 bread wheat grown in crop trials at Tal Jebeen near Aleppo, Syria (Table 2; Fraser et al. 2011; Wallace et al. 2013), suggest that variations in size and shape corresponding to different manuring and irrigation regimes are similar in scale to natural variations occurring along the ear and within individual spikelets (ESM 3; Roushannafas 2022). Within-ear variation does, however, seem to be influenced by growing condition, in that under more stressed conditions, ears produce fewer grains overall and per spikelet. Further research may be able to disentangle these interrelated factors, but results so far do not indicate that these morphological differences are more distinctive than those seen at the (sub)species or varietal level.
Discussion
Potential for identification of ploidy and (sub)species using geometric morphometrics
The first stage of analysis assessed whether outline analysis could be used to distinguish charred FTW grains to ploidy and (sub)species level. The modern reference material study (Table 3) correctly assigned 85.6–86.1% of charred modern grains to (sub)species and 89.8% to ploidy level, a slightly lower rate than the 96.3% obtained in a parallel study of glume wheat grains (Roushannafas et al. 2022). However, in practice, our ability to distinguish between the two hexaploid FTW forms (aestivum and compactum) proved more complex, particularly when comparing them to archaeological material. The relatively high reclassification rates obtained for the modern reference material when accession was used as a discriminating factor (82.1%) does suggest that the model is sensitive to differences at the level of variety. It is therefore possible that adding more (landrace) accessions to the modern reference dataset would provide the model with enough information to establish a more consistent morphological separation of aestivum from compactum wheats. This is also suggested by the fact that the eight hexaploid accessions analysed in Stage 2b could be correctly reclassified to (sub)species at 95.8%.
It was also observed that, at the individual level, a grain may stray quite far from the ‘normal’ shape range for its type, and such analysis should therefore be interpreted in terms of the overall assemblage. A small number of grains classifying or plotting against a particular (sub)species does not constitute unambiguous evidence that this taxon was cultivated. The proposed reclassification rates are also based on the assumption that the grain being classified belongs to one of the represented taxonomic categories, and that the comparative material covers the morphological range of those (sub)species reasonably well, of which we cannot be certain. Such limitations are not unique to the GMM approach. Comparative models can only be improved by increasing the range and relevance of the comparative material used. However, while increasing the size of a physical reference collection may improve the accuracy of identifications, cross-referencing additional specimens can add significantly to the workload of a researcher. The advantage of a GMM approach is that specimens can be compared simultaneously and a dataset built up cumulatively without overwhelming an individual researcher’s capacity; particularly so if GMM specialists take an open science approach which facilitates the re-use of gathered data (data and scripts for the present article can be found at https://github.com/TinaRoushan).
Archaeological results
The identification of two morphologically distinct groups of free threshing wheat from the two archaeological sites is relevant to our understanding of the early origins and subsequent history of FTW. At Çatalhöyük, chaff evidence identified the presence of hexaploid FTW (Bogaard 2016). Results here would seem to confirm that a hexaploid form was the dominant FTW crop (although we cannot rule out the minor presence of a tetraploid wheat). Analysis established a strong morphological similarity of the archaeobotanical material to modern-day examples from the aestivo-compactum group, particularly traditional varieties from central and western Asia. Despite initial analysis suggesting that modern bread and club wheat can be separated by grain shape, the Çatalhöyük grains could not be clearly identified as either aestivum or compactum, although a higher proportion was classified as the former. This result could be interpreted in a number of ways:
-
1.
There is a diversity of forms or cultivars among the Çatalhöyük FTW grains
-
2.
The Çatalhöyük grains are of a type with morphology that is distinct from or intermediate between bread and club wheat.
-
3.
The method is not sufficiently sensitive to distinguish between bread and club wheat when comparing archaeological grains which have been preserved under variable conditions with ones charred in controlled modern experiments.
-
4.
The model requires more data to distinguish between bread and club wheat when making comparisons with archaeological material.
With a sequence beginning in the late 8th millennium bce, Çatalhöyük is one of several sites with finds of hexaploid FTW from a period in which early records for the tetraploid form remain elusive. These findings demonstrate the apparently rapid evolution and adoption of the hexaploid form and the need for further investigation into the mechanisms of this process.
From Kouphovouno no chaff was found by which the FTW remains could be identified to ploidy level. Results show that the 6th millennium bce Kouphovouno grains most closely resemble tetraploid durum wheat, particularly traditional varieties originating from North Macedonia and Cyprus (Table 7). The Kouphovouno results contribute to the theory of early FTW cultivation in the Mediterranean mainly of tetraploid forms. This was most likely to have been a durum-like wheat suited to warm and relatively dry conditions which was, certainly by the period of Classical Antiquity, ‘the greatly predominant naked wheat’ of the region (Jasny 1944, p 27). This tradition survived into the present day, and the region has remained a centre of diversity for durum wheats (Soriano et al. 2018). The same tradition diffused through southern France to arrive in the lake shore settlements around the Alps by ~ 4000 bce (Maier 1996), while hexaploid wheats spread from western Asia to the north and west through the Balkans and Carpathian basin (Kirleis and Fischer 2014).
Research suggests that durum needs a deep rich soil (Percival 1921; Hirzel and Matus 2013) and isotopic evidence indicates that at Kouphovouno the FTW crops were preferentially manured in comparison to barley and einkorn (Vaiglova et al. 2014, 2020). While we cannot be certain of the ecological requirements of past crops, it is possible that this distinction in practice reflects the understanding of the farmers about the needs of their various crops and/or their prioritisation of FTW as a crop grown exclusively for human consumption (Vaiglova et al. 2014, 2023). Preferential manuring of tetraploid FTW over barley has also been observed at the Neolithic circum-Alpine settlement of Sipplingen (Styring et al. 2016). Conversely, at Neolithic Halai (central Greece), isotopic results for several crops including FTW indicate cultivation under a range of conditions, which was interpreted as an overall risk management strategy in response to the relatively drier climatic conditions than those at Kouphovouno (Vaiglova et al. 2023). It is likely that the intensive management of FTW at Kouphovouno was related to the particular value of the crop, and also to the environmental context, the ecological requirements of the crop and available resources.
Hexaploid FTW at Çatalhöyük was apparently grown under more variable conditions, also probably reflecting risk management in a semi-arid landscape (Bogaard et al. 2017), and potentially the result of households cultivating plots of different sizes and various distances from the main settlement in a ‘radial wedge’ model (Bogaard et al. 2021), with smaller, more intensely cultivated plots situated closer to the settlement. It is possible that from an early stage, hexaploid FTW possessed greater adaptive potential than tetraploid wheat from its higher ploidy status (Dubcovsky and Dvorak 2007), allowing it to be grown under a wider range of conditions. In particular storage contexts FTW had clearly been grown under high input regimes (Green et al. 2018) and was generally better watered than barley (Wallace et al. 2015), suggesting that, at least in some cases, the crop was considered worth the relatively high labour investment.
At both sites we see a change in the final phases towards increased cultivation of hulled cereals. At Çatalhöyük the Final Neolithic and Chalcolithic West Mound phases have increasing proportions of hulled barley and glume wheats, and at Kouphovouno einkorn became the main wheat crop in the Late Neolithic as the settlement contracted. A similar pattern is observed in the final (Horgen, ca. 3300–2800 cal bce) cultural phase at Hornstaad-Hörnle, southwest Germany, where δ15N values and relative abundance of FTW tetraploid wheat in comparison to emmer decrease (Styring et al. 2016). This suggests a shift in the balance between the cultural value of the crop versus the burden of its requirements. It is possible that the high input regime associated with FTW, at least at Kouphovouno, became difficult to sustain. A shift towards glume wheats may also relate to the fact that their ‘hulledness’ provides better resistance to pest or fungal attack, both in the field and in storage, as compared to FTWs (Nesbitt and Samuel 1996). The progression of bread and durum wheats towards pre-eminence over other (hulled) wheat forms was not, apparently, a linear process.
Conclusions
This study established that it is possible to identify charred modern free-threshing wheat grains to ploidy and (sub)species using geometric morphometric analysis of their outlines in dorsal and lateral views. Separation of modern Triticum durum (durum), T. turgidum (rivet), T. compactum (club) and T. aestivum (bread) wheat grains was successful to a high level of reclassification. Applying these distinctions to archaeological material was broadly successful, although distinction within the hexaploid group proved less clear than when comparing modern grains. Whether this is because the archaeological material is morphologically diverse or simply due to limitations of the methodology is unclear. Success in classification according to accession was relatively high, suggesting sensitivity of the model to variation within modern sub-species. It is therefore possible that morphological variation within the hexaploid group could be clarified by expanding modern reference datasets. Overall, the results suggest that there is significant potential for GMM analysis to refine taxonomic classifications of archaeobotanical grains at the assemblage (if not individual) level.
Analysis of charred archaeobotanical material from Kouphovouno and Çatalhöyük revealed two distinct morphological types of grains relating to tetraploid and hexaploid FTW forms respectively. At Çatalhöyük, remains from the 7th millennium bce most closely resembled modern grains from the hexaploid aestivum-compactum group, reinforcing evidence from previous chaff analysis. Further confirmation of the hexaploid identification highlights the dearth of early evidence for the tetraploid form which should logically have preceded it, and the need to elucidate the apparently relatively short period in which hexaploid wheat evolved.
At Kouphovouno, where diagnostic chaff remains were absent, results suggest cultivation of a tetraploid wheat closely resembling durum. Archaeobotanical evidence of the earliest cultivation of tetraploid FTW remains limited and the Kouphovouno material represents a significant addition to evidence for a Neolithic tradition of predominantly tetraploid FTW cultivation which spread west along the Mediterranean coast. At both sites, a revival in cultivation of hulled wheat forms is evidenced in later phases, suggesting a change in the balance between the cultural value of FTW and the desire or ability to meet its growing and/or storage requirements. The current widespread preference for FTWs was not, therefore, a foregone conclusion; we do not see early farming societies progressing steadily towards the crops which we now consider to be ‘optimal’, but rather making contextually-situated decisions about which crops to grow according to various factors.
At both sites, comparisons between modern and archaeological FTW grains suggested that archaeological material was more similar to ‘traditional’ varieties from regions closest to the site of study. These findings suggest the importance of careful selection of comparative material for morphological studies of archaeobotanical material, with geographical region and improvement status (traditional or not) potentially highly significant. The role played by the growing environment is less clear, with preliminary results suggesting a possible link between growth stress, grain production and grain morphology, which influences the grain shape less distinctly than taxon or variety.
References
Antolín F (2016) Local, intensive and diverse?: early farmers and plant economy in the North-East of the Iberian Peninsula (5,500–2,300 cal bc). Barkhuis Publishing, Groningen
Arranz-Otaegui A, Colledge S, Zapata L et al (2016) Regional diversity on the timing for the initial appearance of cereal cultivation and domestication in southwest Asia. Proc Natl Acad Sci USA 113:14001–14006. https://doi.org/10.1073/pnas.1612797113
Ayala G, Wainwright J, Lloyd JM et al (2021) Disentangling the palaeoenvironmental reconstructions of Çatalhöyük. In: Hodder I (ed) Peopling the landscape of Çatalhöyük: reports from the 2009–2017 seasons. British Institute at Ankara, London, pp 31–46
Ayala G, Bogaard A, Charles M, Wainwright J (2022) Resilience and adaptation of agricultural practice in neolithic çatalhöyük. Turk World Archaeol. https://doi.org/10.1080/00438243.2022.2125058
Bettge AD (2009) Club wheat: Functionally, the best sub-class and sub-species in soft wheat (webcast). https://www.cerealsgrains.org/publications/plexus/cfwplexus/library/webcasts/Pages/ABettge.aspx. Accessed 1 Nov 2021
Bilgic H, Hakki EE, Pandey A et al (2016) Ancient DNA from 8400 year-old Çatalhöyük wheat: implications for the origin of neolithic agriculture. PLoS ONE 11:e0151974. https://doi.org/10.1371/journal.pone.0151974
Boardman S, Jones G (1990) Experiments on the effects of charring on cereal plant components. J Archaeol Sci 17:1–11. https://doi.org/10.1016/0305-4403(90)90012-T
Bogaard A (2016) Archaeobotany: the wheat and the chaff. Nat Plants 2:16079. https://doi.org/10.1038/nplants.2016.79
Bogaard A, Charles M, Livarda A et al (2013) The archaeobotany of mid-later occupation levels at Çatalhöyük. In: Hodder I et al (eds) Humans and landscapes of Çatalhöyük: reports from the 2000–2008 seasons. Cotsen Institute of Archaeology at UCLA, Los Angeles, pp 93–128
Bogaard A, Filipović D, Fairbairn A et al (2017) Agricultural innovation and resilience in a long-lived early farming community: the 1,500-year sequence at neolithic to early Chalcolithic Çatalhöyük, central Anatolia. Anatol Stud 67:1–28. https://doi.org/10.1017/S0066154617000072
Bogaard A, Charles M, Filipovic D et al (2021) The archaeobotany of Çatalhöyük: results from 2009–2017 excavations and final synthesis. In: Hodder I (ed) Peopling the landscape of Çatalhöyük: reports from the 2009–2017 seasons. British Institute at Ankara, London, pp 91–123
Bonhomme V, Picq S, Gaucherel C, Claude J (2014) Momocs: outline analysis using R. J Stat Softw 56:1–24. https://doi.org/10.18637/jss.v056.i13
Bonhomme V, Forster E, Wallace M et al (2017) Identification of inter- and intra-species variation in cereal grains through geometric morphometric analysis, and its resilience under experimental charring. J Archaeol Sci 86:60–67. https://doi.org/10.1016/j.jas.2017.09.010
Borrell F, Boëda É, Molist M et al (2011) New evidence concerning the neolithisation process of the central syrian desert: the neolithic complex of Mamarrul Nasr. Paléorient 37:35–46
Carruthers W (2019) Charred, waterlogged and mineralised plant remains. In: Rady J, Holman J (eds) Beneath the seamark: 6,000 years of an island‘s history: archaeological investigations at ‘Thanet Earth’, kent 2007–2012, vol 2. Canterbury Archaeological Trust, Canterbury
Cavanagh W, Renard J, Bogaard A et al (2017) Farming strategies at Kouphovouno, Lakonia, in the MN-LN periods. In: Sarris A, Kalogiropoulou E, Kalayci T, Karimali L (eds) Communities, landscapes and interaction in Neolithic Greece. Proceedings of the International Conference, Rethymno. International Monographs in Prehistory, Ann Arbor, pp 281–291, 29–30 May 2015
Charles M, Forster E, Wallace M, Jones G (2015) Nor ever lightning char thy grain”1: establishing archaeologically relevant charring conditions and their effect on glume wheat grain morphology. STAR: Sci Technol Archaeol Res 1:1–6. https://doi.org/10.1179/2054892315Y.0000000008
Claude J (2008) Morphometrics with R. Springer, New York
De Moulins D (1993) Les restes de plantes carbonisées de Cafer Höyük. In: Cauvin J (ed) Cahiers de l’Euphrate, vol 7. Editions Recherche sur les Civilisations, Paris, pp 191–234
De Moulins D (1997) Agricultural changes at euphrates and steppe sites in the mid-8th to the 6th millennium B.C. (BAR International series 683) british archaeological reports, Oxford
Der L, Issavi J (2017) The urban quandary and the ‘mega-site’ from the Çatalhöyük perspective. J World Prehist 30:189–206. https://doi.org/10.1007/s10963-017-9103-x
Douka K, Efstratiou N, Hald MM et al (2017) Dating knossos and the arrival of the earliest neolithic in the southern Aegean. Antiquity 91:304–321. https://doi.org/10.15184/aqy.2017.29
Dubcovsky J, Dvorak J (2007) Genome plasticity a key factor in the success of polyploid wheat under domestication. Science. https://doi.org/10.1126/science.1143986.
Dvorak J, Luo M-C, Yang Z-L, Zhang H-B (1998) The structure of the Aegilops tauschii genepool and the evolution of hexaploid wheat. Theor Appl Genet 97:657–670. https://doi.org/10.1007/s001220050942
Dvorak J, Deal KR, Luo M-C et al (2012) The origin of spelt and free-threshing hexaploid wheat. J Hered 103:426–441. https://doi.org/10.1093/jhered/esr152
Ergun M, Tengberg M, Willcox G, Douché C (2018) Plants of Aşıklı Höyük and changes through time: first archaeobotanical results from the 2010-14 excavation seasons. In: Özbaşaran M, Duru G, Stiner M (eds) The early settlement at Aşıklı Höyük: essays in honor of Ufuk Esin. Ege Yayınları, Istanbul, pp 191–217
Evin A, Bonhomme V, Claude J (2020) Optimizing digitalization effort in morphometrics. Biol Methods Protoc 5:bpaa023. https://doi.org/10.1093/biomethods/bpaa023
Evin A, Bouby L, Bonhomme V et al (2022) Archaeophenomics of ancient domestic plants and animals using geometric morphometrics: a review. Peer Commun J 2:e27. https://doi.org/10.24072/pcjournal.126
Ex Oriente (2022) PPND (The platform for neolithic radiocarbon dates). https://www.exoriente.org/associated_projects/ppnd.php. Accessed 8 June 2022
Fairbairn A, Asouti E, Near J, Martinoli D (2002) Macro-botanical evidence for plant use at Neolithic Çatalhöyük south-central Anatolia, Turkey. Veget Hist Archaeobot 11:41–54. https://doi.org/10.1007/s003340200005
Fairbairn A, Jacobsson P, Baird D et al (2020) Settlement change on the western Konya Plain: refining Neolithic and Chalcolithic chronologies at Canhasan, Turkey. Antiquity 94:342–360. https://doi.org/10.15184/aqy.2020.13
Fernández E, Thaw S, Brown TA et al (2013) DNA analysis in charred grains of naked wheat from several archaeological sites in Spain. J Archaeol Sci 40:659–670. https://doi.org/10.1016/j.jas.2012.07.014
Fraser RA, Bogaard A, Heaton T et al (2011) Manuring and stable nitrogen isotope ratios in cereals and pulses: towards a new archaeobotanical approach to the inference of land use and dietary practices. J Archaeol Sci. https://doi.org/10.1016/j.jas.2011.06.024
Giardina CR, Kuhl FP (1977) Accuracy of curve approximation by harmonically related vectors with elliptical loci. Comput Graph Image Process 6:277–285. https://doi.org/10.1016/S0146-664X(77)80029-4
Giles RJ, Brown TA (2006) GluDy allele variations in Aegilops tauschii and Triticum aestivum: implications for the origins of hexaploid wheats. Theor Appl Genet. https://doi.org/10.1007/s00122-006-0259-5
Giraldo P, Benavente E, Manzano-Agugliaro F, Gimenez E (2019) Worldwide research trends on wheat and barley: a bibliometric comparative analysis. Agronomy 9:352. https://doi.org/10.3390/agronomy9070352
Goncharov NP (2011) Genus Triticum L. taxonomy: the present and the future. Plant Syst Evol 295:1–11. https://doi.org/10.1007/s00606-011-0480-9
Grausgruber H, Oberforster M, Ghambashidze G, Ruckenbauer P (2005) Yield and agronomic traits of Khorasan wheat (Triticum turanicum Jakubz). Field Crops Res 91:319–327. https://doi.org/10.1016/j.fcr.2004.08.001
Green L, Charles M, Bogaard A (2018) Exploring the agroecology of neolithic Çatalhöyük, central Anatolia: an archaeobotanical approach to agricultural intensity based on functional ecological analysis of arable weed flora. Paléorient 44:29–44
Haque MA, Takayama A, Watanabe N, Kuboyama T (2011) Cytological and genetic mapping of the gene for four-awned phenotype in Triticum carthlicum nevski. Genet Resour Crop Evol. https://doi.org/10.1007/s10722-010-9644-7
Helbæk H (1968) Knossos wheat. Unpublished manuscript, second draft
Hillman GC (1972) Plant remains [In french DH, excavations at can Hasan III 1969–1970. In: Higgs ES (ed) Papers in economic prehistory: studies by members and associates of the british academy major research project in the early history of agriculture. Cambridge University Press, Cambridge, pp 182–188
Hillman GC (1978) On the origins of domestic rye—Secale cereale: the finds from aceramic can Hasan III in Turkey. Anatol Stud 28:157–174. https://doi.org/10.2307/3642748
Hillman GC (1984) Interpretation of archaeological plant remains: the application of ethnographic models from Turkey. In: van Zeist W, Casparie WA (eds) Plants and ancient man: studies in palaeoethnobotany. Proceedings of the Sixth Symposium of the International Work Group for Palaeoethnobotany, Groningen, 30 May. Rotterdam, pp 1–41, 3 June 1983
Hillman GC (2001) Archaeology, percival, and the problems of identifying wheat remains. In: Caligari PDS, Brandham PE (eds) Wheat taxonomy: the legacy of John Percival, vol 3. Academic Press, London, pp 27–36
Hillman GC, Mason S, de Moulins D, Nesbitt M (1996) Identification of archaeological remains of wheat: the 1992 London workshop. Circaea 12:195–210
Hirzel J, Matus I (2013) Effect of soil depth and increasing fertilization rate on yield and its components of two durum wheat varieties. Chil J Agric Res 73:55–59. https://doi.org/10.4067/S0718-58392013000100008
Hodder I (2021) Changing Çatalhöyük worlds. In: Hodder I (ed) Peopling the landscape of Çatalhöyük: reports from the 2009–2017 seasons. British Institute at Ankara, London, pp 1–29
Jacomet S (2007) Neolithic plant economies in the northern Alpine foreland from 5,500–3,500 cal bc. In: Colledge S, Conolly J (eds) The origins and spread of domestic plants in southwest Asia and Europe. Left Coast Press, Walnut Creek, pp 221–258
Jasny N (1944) The wheats of classical antiquity. Johns Hopkins Press, Baltimore
Jones G (1987) A statistical approach to the archaeological identification of crop processing. J Archaeol Sci 14:311–323. https://doi.org/10.1016/0305-4403(87)90019-7
Jones G (1998) Wheat grain identification–why bother? Environ Archaeol 2:29–34. https://doi.org/10.1179/env.1997.2.1.29
Jones G, Rowley-Conwy P (2007) On the importance of cereal cultivation in the british neolithic. In: Colledge S, Conolly J (eds) The origins and spread of domestic plants in southwest Asia and Europe. Left Coast Press, Walnut Creek, pp 391–419
Kirleis W, Fischer E (2014) Neolithic cultivation of tetraploid free threshing wheat in Denmark and northern Germany: implications for crop diversity and societal dynamics of the funnel beaker culture. Veget Hist Archaeobot 23:81–96. https://doi.org/10.1007/s00334-014-0440-8
Kuckuck H (1979) On the origin of Triticum carthlicum Nevski (Triticum persicum Vav.) [a kind of wheat]. Wheat Inform Service 50:1–5
Kuhl FP, Giardina CR (1982) Elliptic fourier features of a closed contour. Comput Graph Image Process 18:236–258. https://doi.org/10.1016/0146-664X(82)90034-X
Kushnareva KK (1997) The southern Caucasus in prehistory: stages of cultural and socioeconomic development from the eighth to the second millennium B.C. University of Pennsylvania Press, Philadelphia
Letts JB (1999) Smoke-blackened thatch: a unique source of late medieval plant remains from southern England. English Heritage, London
Levy AA, Feldman M (2022) Evolution and origin of bread wheat. Plant Cell. https://doi.org/10.1093/plcell/koac130
Liu X, Lister DL, Zhao Z et al (2016) The virtues of small grain size: potential pathways to a distinguishing feature of asian wheats. Quat Int 426:107–119. https://doi.org/10.1016/j.quaint.2016.02.059
Livarda A (2014) Archaeobotany in Greece. Archaeol Rep 60:106–116. https://doi.org/10.1017/S057060841400012X
Maier U (1996) Morphological studies of free-threshing wheat ears from a neolithic site in southwest Germany, and the history of the naked wheats. Veget Hist Archaebot 5:39–55. https://doi.org/10.1007/BF00189434
Marinova E (2007) Archaeobotanical data from the early neolithic of Bulgaria. In: Colledge S, Conolly J (eds) The origins and spread of domestic plants in southwest Asia and Europe. Left Coast Press, Walnut Creek, pp 93–109
Matsuoka Y (2011) Evolution of polyploid Triticum wheats under cultivation: the role of domestication, natural hybridization and allopolyploid speciation in their diversification. Plant Cell Physiol 52:750–764. https://doi.org/10.1093/pcp/pcr018
Mee C, Cavanagh B, Renard J (2014) The middle-late neolithic transition at Kouphovouno. Annu Br School Athens 109:65–95
Michalcová V, Dušinský R, Sabo M et al (2014) Taxonomical classification and origin of Kamut® wheat. Plant Syst Evol. https://doi.org/10.1007/s00606-014-1001-4
Moffett L (1987) The macro-botanical evidence from late saxon and early medieval stafford. Anc Monum Lab Rep 169:87
Mori N, Ohta S, Chiba H et al (2013) Rediscovery of Indian dwarf wheat (Triticum aestivum L. ssp. sphaerococcum (Perc.) MK.): an ancient crop of the Indian subcontinent. Genet Resour Crop Evol 60:1771–1775
Nesbitt M, Samuel D (1996) From staple crop to extinction? The archaeology and history of the hulled wheats. In: Padulosi S, Hammer K, Heller J (eds) Hulled Wheat: Proceedings of the First International Workshop on Hulled Wheats, 21–22 July 1995, Tuscany, International Plant Genetic Resources Institute (IPGRI), Rome, pp 40–99
Oliveira HR, Jacocks L, Czajkowska BI et al (2020) Multiregional origins of the domesticated tetraploid wheats. PLoS ONE 15:e0227148. https://doi.org/10.1371/journal.pone.0227148
Percival J (1921) The wheat plant: a monograph. Duckworth and Co., London
Pont C, Leroy T, Seidel M et al (2019) Tracing the ancestry of modern bread wheats. Nat Genet 51:905–911. https://doi.org/10.1038/s41588-019-0393-z
Portillo M, Ball TB, Wallace M et al (2020) Advances in morphometrics in archaeobotany. Environ Archaeol 25:246–256. https://doi.org/10.1080/14614103.2019.1569351
Pruidze L, Maisaia I, Sikharulidze S, Tavartkiladze M (2016) Our daily bread: Georgia the ancient cradle of agriculture. Palitra L Publishing, Tbilisi
Renard J, Cavanagh W (2017) Kouphovouno (Lakonia): Some thoughts about the settlement pattern at the end of the middle Neolithic. In: Sarris A, Kalogiropoulou E, Kalayci T, Karimali L (eds) Communities, landscapes and interaction in Neolithic Greece. Proceedings of the International Conference, Rethymno 29–30 May, 2015. International Monographs in Prehistory, Ann Arbor, pp 149–166
Rogosa E (2016) Restoring heritage grains: the culture, biodiversity, resilience, and cuisine of ancient wheats. Chelsea Green Publishing, White River Junction, Vermont
Rottoli M, Pessina A (2007) Neolithic agriculture in Italy: an update of archaeobotanical data with particular emphasis on northern settlements. In: Colledge S, Conolly J (eds) The origins and spread of domestic plants in southwest Asia and Europe. Left Coast Press, Walnut Creek, pp 141–153
Roushannafas T (2022) ‘To call each thing by its right name’? Exploring the potential of geometric morphometrics to interpret the diversity of archaeobotanical wheat grains. Doctoral Thesis, University of Oxford, Oxford
Roushannafas T, Bogaard A, Charles M (2022) Geometric morphometrics sheds new light on the identification and domestication status of ‘new glume wheat’ at Neolithic Çatalhöyük. J Archaeol Sci 142:105599. https://doi.org/10.1016/j.jas.2022.105599
Sarpaki A (2013) The economy of neolithic Knossos: the archaeobotanical data. In: Efstratiou N, Karetsou A, Ntinou M (eds) The neolithic settlement of knossos in Crete: new evidence for the early occupation of Crete and the Aegean Islands. INSTAP Academic Press, Philadelphia, pp 63–94
Schlumbaum A, Neuhaus J-M, Jacomet S (1998) Coexistence of tetraploid and hexaploid naked wheat in a neolithic lake dwelling of central Europe: evidence from morphology and ancient DNA. J Archaeol Sci 25:1111–1118
Slageren MW (1994) Wild wheats: a monograph of Aegilops L. and Amblyopyrum (Jaub. & Spach) Eig (Poaceae). Wageningen Agricultural University Papers, Wageningen
Soriano JM, Villegas D, Sorrells ME, Royo C (2018) Durum wheat landraces from east and west regions of the Mediterranean Basin are genetically distinct for yield components and phenology. Front Plant Sci 9:80. https://doi.org/10.3389/fpls.2018.00080
Stroud E, Bogaard A, Charles M (2021) A stable isotope and functional weed ecology investigation into chalcolithic cultivation practices in central Anatolia: Çatalhöyük, Çamlıbel Tarlası and Kuruçay. J Archaeol Sci Rep 38:103010. https://doi.org/10.1016/j.jasrep.2021.103010
Styring A, Maier U, Stephan E et al (2016) Cultivation of choice: new insights into farming practices at neolithic lakeshore sites. Antiquity 90:95–110. https://doi.org/10.15184/aqy.2015.192
Szczepanek M, Lemańczyk G, Lamparski R (2020) Ancient wheat species (Triticum sphaerococcum Perc. and T. persicum Vav.) in organic farming: influence of sowing density on agronomic traits, pests and diseases occurrence, and weed infestation. Agriculture. https://doi.org/10.3390/agriculture10110556
Vaiglova P (2016) Neolithic agricultural management in the eastern Mediterranean: new insight from a multi-isotope approach. Doctoral Thesis, University of Oxford, Oxford
Vaiglova P, Bogaard A, Collins M et al (2014) An integrated stable isotope study of plants and animals from Kouphovouno, southern Greece: â new look at neolithic farming. J Archaeol Sci 42:201–215. https://doi.org/10.1016/j.jas.2013.10.023
Vaiglova P, Gardeisen A, Buckley M et al (2020) Further insight into neolithic agricultural management at Kouphovouno, southern Greece: expanding the isotopic approach. Archaeol Anthropol Sci 12:43. https://doi.org/10.1007/s12520-019-00960-y
Vaiglova P, Coleman J, Diffey C et al (2023) Exploring diversity in neolithic agropastoral management in mainland Greece using stable isotope analysis. Environ Archaeol 28:62–85. https://doi.org/10.1080/14614103.2020.1867292
Van Zeist W (1976) On macroscopic traces of food plants in southwestern Asia (with some reference to pollen data). Philos Trans R Soc Lond B 275:27–41. https://doi.org/10.1098/rstb.1976.0068
Van Zeist W, Bakker-Heeres JAH (1982) Archaeobotanical studies in the levant, 1: neolithic sites in the damascus basin. aswad, ghoraifé, ramad. Palaeohistoria 24:165–256
Veropoulidou R (2021) The archaeomalacology of the 2009–2017 excavations at neolithic Çatalhöyük. In: Hodder I (ed) Peopling the landscape of Çatalhöyük: reports from the 2009–2017 seasons. British Institute at Ankara, London, pp 47–71
Wallace M, Jones G, Charles M et al (2013) Stable carbon isotope analysis as a direct means of inferring crop water status and water management practices. World Archaeol 45:388–409. https://doi.org/10.1080/00438243.2013.821671
Wallace MP, Jones G, Charles M et al (2015) Stable carbon isotope evidence for neolithic and bronze age crop water management in the eastern Mediterranean and southwest Asia. PLoS ONE 10:e0127085. https://doi.org/10.1371/journal.pone.0127085
Zhuang P, Ren Q, Li W, Chen G (2011) Genetic diversity of Persian wheat (Triticum turgidum ssp. carthlicum): accessions by EST-SSR markers. Am J Biochem Mol Biol 1:223–230. https://doi.org/10.3923/ajbmb.2011.223.230
Zohary D (1969) The progenitors of wheat and barley in relation to domestication and agriculture dispersal in the old World. In: Ucko PJ, Dimbleby GW (eds) The domestication and exploitation of plants and animals. Duckworth and Co, London, pp 47–66
Zohary D, Hopf M, Weiss E (2012) Domestication of plants in the old world: the origin and spread of domesticated plants in Southwest Asia, Europe, and the Mediterranean Basin, 4th edn. Oxford University Press, Oxford
Zwer PK, Sombrero A, Rickman RW, Klepper B (1995) Club and common wheat yield component and spike development in the pacific northwest. Crop Sci. https://doi.org/10.2135/cropsci1995.0011183X003500060012x
Acknowledgements
Thanks are given to the Arts and Humanities Research Council (AHRC) Open-Oxford-Cambridge Doctoral Training Partnership (OOCDTP) and Merton College, Oxford for making this research possible through their funding. Thanks also to Müge Ergun and Alexander Weide for comments on the manuscript, and to Charlotte Diffey for providing contextual information for the Kouphovouno material.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Communicated by A. Fairbairn.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Roushannafas, T., Bogaard, A. & Charles, M. Geometric morphometric analysis of Neolithic wheat grains: insights into the early development of free-threshing forms. Veget Hist Archaeobot 32, 597–613 (2023). https://doi.org/10.1007/s00334-023-00929-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00334-023-00929-x