
Abstract
Albatrosses are renowned for their high philopatry, which has been proposed as a major barrier to their dispersal and hybridization with other albatross species. Except for shy-type albatross species from New Zealand, examples of colonization of new breeding sites and interbreeding with closely related species are rare. During the austral summer of 2022, while conducting land-based monitoring of resident Black-browed (Thalassarche melanophris) and Grey-headed (T. chrysostoma) albatross populations on the Diego Ramírez Islands, Chile (56 S), we documented the first (and southernmost) breeding record of a nesting pair of White-capped Albatross (T. steadi), a species endemic to the Auckland Islands, New Zealand (~ 7000 km away). The species identity was confirmed through molecular analysis. This discovery underscores the importance of Diego Ramírez and its surrounding waters as a hotspot for global albatross conservation.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Albatrosses are large, long-lived seabirds with wide foraging and migratory distributions. They typically breed on remote islands or in continental areas that are difficult to access (Chown et al. 1998; Tickell 2000). Albatrosses are highly philopatric, even among other seabird and waterbird species (Coulson 2016). They might also experience genetic barriers, as has become evident in birds at some colonies located in close proximity to one another (Danckwerts et al. 2021).
There are very few published accounts of albatrosses exploring alternative breeding sites and attempting to breed outside of their known range. There are only four known cases of Salvin’s Albatross (Thalassarche salvini) breeding at the Crozet Archipelago–Île des Pingouins–which lies approximately 7850 km from its nearest known colony, the Snares Islands, south of New Zealand, despite surveillance over decadal time scales (Jouventin 1990). Furthermore, a single-banded male Laysan Albatross (Phoebastria immutabilis) from Kaena Point in Oahu Island, Hawaii, was found on Natividad Island in the Mexican Pacific, approximately 4390 km from its breeding site (Albores-Barajas et al. 2020).
Here, we describe the first confirmed breeding of a conspecific pair of the White–capped Albatross (T. steadi) at the subantarctic Diego Ramírez Islands, Chile (hereafter, Diego Ramírez), which hold approximately 155,000 pairs of Black-browed albatrosses (T. melanophris) in six breeding sites and approximately 18,000 pairs of Grey-headed albatrosses (T. chrysostoma) (Robertson et al. 2014, 2017). Until now, the White-capped Albatross was considered to be endemic to the southern waters of New Zealand at around 50ºS (Agreement on the Conservation of Albatrosses and Petrels 2011).
Materials and methods
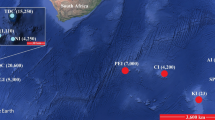
Diego Ramírez (56º31′17’’S; 68º42′53’’W) lies in the northern section of the Drake Passage, approximately 112 km southwest of Cape Horn, southern Chile. This archipelago includes two main islands (Bartolomé 0.93 km2, Gonzalo 0.49 km2), and several adjacent islets (Fig. 1). Between 25 and 27 February 2022, we monitored breeding albatross activity on Gonzalo Island by conducting 14 transects (each 20 m wide, totaling 6.9 km).

The Diego Ramírez Islands (56°S), Chile. Aerial view of its two main islands: Bartolomé in the background (0.93 km2) and Gonzalo in the foreground (0.49 km2). Picture adapted from Robertson et al. (2007)
During these surveys, we recorded the identity of albatross species, breeding stage (e.g., incubation, guard, post-guard), chicks’ weight (measured with a Pesola spring scale, accuracy: 50 g), and tarsus length (measured with a Vernier caliper, accuracy: ± 0.05 mm). In addition, we gathered baseline information on habitat characteristics, such as cardinal orientation, tussock grass coverage and height, and terrain slope. Subsequently, we deployed six camera stations for an 8-month period (March–October 2022), positioned at distances ranging from 1.5 to 2.0 m from focal nests. These cameras recorded adult attendance during the post-guard stage from 18 March to 20 May 2022.
The recorded breeding albatrosses included a white downy chick in the guard–post-guard stage of the breeding cycle, with two adults attending the nest. While these adults exhibited external features (size, bill, plumage coloration) resembling those of the closely related albatross species, the New Zealand White–capped Albatross and the Tasmania Shy Albatross (T. cauta), the varying intensity in bill coloration, including the amount of yellow along the cutting edge, was not a defining feature for distinguishing between these two albatross species (see Tennyson 2020; adult details in Online Resource 1).
To identify the species identification of the new chick (and distinguish it from the White-capped Albatross and the Shy Albatross), we conducted a molecular analysis of a blood sample using PCR amplification of a target 181 pb fragment of Domain I of the mitochondrial control region, as described by Abbott and Double (2003a). Whole genomic DNA was extracted using the PureLink Genomic DNA Mini Kit (Invitrogen, Carlsbad, CA). We used primers GluR7 and TEST F4t to amplify the target fragment, and sequencing was performed at AUSTRAL–omics (www.australomics.cl). Additionally, the sequence was aligned using Vector NTI Software (ThermoFisher Scientific) against sequences of representative species of the shy complex from Genbank (www.ncbi.nlm.nih.gov), including Buller’s Albatross (T. bulleri, accession number DQ029001, DQ029001), Salvin’s Albatross (DQ029007, DQ029007), Chatham Albatross (T. eremita, DQ029005, DQ029006), Shy Albatross (DQ029003, DQ029004), and White-capped Albatross (DQ029009, DQ029010).
Finally, we mapped the known breeding distribution and geographic range of the White-capped Albatross using the rworldmap (South 2011) and ggplot2 (Wickham 2016) packages of R (R Core Team 2022). The geographic range (polygon) was downloaded from The IUCN Red List of Threatened Species (Version 2022–2; https://www.iucnredlist.org/).
Results and discussion
Our survey covered 0.14 km2, representing 30.9% of Gonzalo Island’s total area. Records included dominant breeding Black-browed Albatross and Grey-headed Albatross identified in earlier surveys (Schlatter and Riveros 1987; Robertson et al. 2017). Within the survey area, the former predominantly breeds in unvegetated areas surrounded by tussock grass, while the latter breeds around the margins of Black-browed colonies on steeper terrain, such as cliff faces.
On 26 February 2022, we discovered a group of four active Grey-headed Albatross nests with no adult in attendance, indicating that the chicks were in the post-guard phase of the breeding cycle. Additionally, we encountered one adult albatross of the shy-type sitting on an active nest surrounded by tussock grass measuring 0.9 m in height, among a group of Grey-headed albatrosses (breeding density 0.015 nests m−2; Fig. 2a).

a The White-capped Albatross (Thalassarche steadi) nesting in Diego Ramírez. Brood under parental guard with chick in white down and of smaller size when compared to Grey-headed Albatross nesting in the area. b White-capped Albatross chick close-up standing alone in the early post-guard period. Pictures by Cristóbal Anguita (left) and Carlos Garcés Letelier (right)
Based on a single polymorphism (SNP) at site 121 identified through Domain I mitochondrial analysis, the chick was determined to be a White–capped Albatross (Thalassarche steadi) (Abbott and Double 2003a). The sequence for this individual has been deposited in Genbank under the accession number OR184845. This record marks a new breeding albatross species for this region in the southeast Pacific.
The White-capped Albatross has demonstrated more dispersion than the closely related Shy Albatross from Tasmania, with less population diversity and high fidelity to nearby waters, such as the Bass Strait (Abbott and Double 2003b). It is part of the shy-type albatross complex, identified by recent genetic diversification and exploratory behavior (Phillips et al. 2018).
Miskelly et al. (2020) described the breeding timing of the White-capped Albatross in the Auckland Islands, indicating that laying typically occurs by the end of November, with hatching in the second half of January. Consequently, the chick at Diego Ramírez was approximately 40 days old, with a body weight of 3.650 kg and a tarsus length of 8.11 cm on 16 March 2022 (Fig. 2b). In contrast, the Grey-headed Albatross chicks at Diego Ramírez were 5 months old, with a mean weight of 4.920 kg ± 0.14 S.E., and a tarsus length of 9.22 cm ± 0.07 S.E. (n = 20).
While information regarding the reproductive phenology of the White-capped Albatross remains scarce, closely related species such as the Shy Albatross in Tasmania exhibit a breeding period of eight months, from laying in September to fledging chicks in April (Hedd 1999; Baker 2016). This suggests that the White-capped Albatross nest recorded at Diego Ramírez was laid in November, hatched in January, entered a post-guarding period until April, and fledglings began departing in June.
The frequency of adult attendance in the post-guard phase of the White-capped Albatross was recorded at 63 visits. These visits ranged from zero to two each day until 10 April, when three visits were noted (Fig. 3a). Adults typically arrived between 6 am and 6 pm, with the most common arrival time at 9 am (recorded 17 times, representing 27% of visits; Fig. 3b). In the advanced post-guard phase (early May 2022), adults exhibited a wider schedule of visit times, with more frequent noon visits (Fig. 3b). The time spent at the nest included feeding and preening of the chick, with a minimum stay of approximately 10 min and an extraordinary maximum stay of 960 min (approximately 16 h) by the end of the post-guard phase. The latter included the adult resting at one side of the nest after the usual preening and feeding of the chick (Fig. 3c).

Parental attendance of the White-capped Albatross at Diego Ramírez. The period covered post-guard between 18 March and 20 May 2022 when both adults are foraging at sea at the same time. Above: daily frequency of visits by adults. Middle: schedule of adult arrival (sunrise ~ 6:20 am, sunset ~ 8:11 pm; UTC-3), and Below: time duration of single adult visit per day
Parental attendance by both parents was recorded until 20 May. A month later, at midday on 26 June 2022, the White-capped Albatross chick fledged from Diego Ramírez (a video sequence of parental attendance is provided in Online Resource 2). This fledging occurred two months earlier than in New Zealand (mid-August; Robertson 1985). However, the departure date from Diego Ramírez is more consistent with the intended fledging period of the White-capped Albatross on the opposite side of the South Pacific, now believed to occur between June and July (Agreement on the Conservation of Albatrosses and Petrels 2011).
Seventy-nine days after its departure (end of June 2022), the nest of the White-capped Albatross fledging was occupied by a pair of adult Grey-headed albatrosses. However, these were displaced during five visits of an adult White-capped Albatross to the same nest. This White–capped Albatross remained at the nest until our last visit on 23 October 2022 and occasionally exhibited signs of courtship behavior typical for Thalassarche albatrosses, as described by Tickell (2000). Furthermore, during this last visit, we identified the presence of a second male approximately 60 m away, vocalizing and displaying courtship behavior on the nest, consistent with behavior observed before November, when laying is expected at Diego Ramírez (see Online Resource 3).
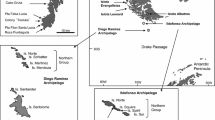
The White-capped Albatross is endemic to New Zealand, with breeding grounds on three groups of islands (Auckland ~ 99.9%, Antipodes < 0.1%, and Chatham < 0.1%), totaling approximately 97,000 breeding pairs (Agreement on the Conservation of Albatrosses and Petrels 2011; Fig. 4). Outside of this range, there is only one recorded instance of White-capped Albatross breeding activity: an individual pairing and raising a chick with a Black-browed Albatross couple at Bird Island, South Georgia (Phalan et al. 2004; Phillips et al. 2018; Fig. 4).

The known breeding distribution of the White-capped Albatross in the Southern Ocean. This albatross species is concentrated in the southwest Pacific including records on conspecific pairs and heterospecific courtship displays in islands of the region and the Auckland Islands as its main breeding location. Out of this endemism range, the White capped was recorded in a mixed-species pair in South Georgia or with suspected breeding in the Prince Edward Island. In orange, the distribution range of the White-capped Albatross now is expanded to the southeast Pacific through its conspecific breeding recorded in this study
Other potential instances of breeding exploration by the White-capped Albatross outside of its regular range include a single adult displaying with an adult Salvin’s Albatross on Toru Islet, Snares Islands off southern New Zealand, as well as a probable White–capped Albatross incubating with an unknown partner at Prince Edward Island in the southwest Indian Ocean (Miskelly 1984; Ryan et al. 2009; Fig. 4). Thus, the couple at Diego Ramírez represents the southernmost record and the first conspecific pair recorded for this albatross species, approximately 7100 km away from its main breeding colony at Disappointment Island in the Auckland Group of New Zealand waters (Fig. 4).
Since 2019, Diego Ramírez has been designated as a Marine Protected Area and a seabird refugium, as it is free of invasive mammals, unlike the Cape Horn islands, which are affected by at least 11 mammalian species (Schüttler et al. 2019). In contrast, the Auckland Islands have been inhabited by sheep, rats, rabbits, feral cats, and pigs, among others (Falla 1965), resulting in unmeasured impacts on the White-capped Albatross through the destruction of breeding sites (Flux 2002). At sea, despite conservation measures currently in place in their natal waters (Fisheries New Zealand 2020), at least 8,000 White-capped albatrosses are killed annually in longline and trawl fisheries (Baker et al. 2007, 2023). Chilean domestic fisheries still overlap with the feeding grounds of breeding and non-breeding albatrosses, including trawl fisheries, resulting in annual potential seabird fatalities with an estimated mean of 66,500 individuals when cryptic multipliers are added to observer-collected data (Richard and Adasme 2019).
Given these threats, Chilean albatross colonies require ongoing, long-term monitoring of species composition and status to inform conservation efforts effectively. Forty-five years ago, Diego Ramírez was virtually unknown in the global albatross scene (Tickell 1976). Just four years later, in the Austral summer of 1980–1981, Diego Ramírez was identified as an important hotspot for subantarctic seabird diversity, supported by the absence of detrimental invasive rodents (Schlatter and Riveros 1987).
Diego Ramírez is one of six known albatross colonies distributed between 51º–56ºS, with five located in the open ocean and one occurring in the protected waters of the Chilean channels (Lawton et al. 2003; Robertson et al. 2007). Notably, given the focus of this paper, a single juvenile or subadult Salvin’s Albatross has been recorded resting near breeding Black-browed albatrosses and Rockhopper Penguins (Eudyptes chrysocome) at Diego Ramírez (Arata 2003).
In addition, signs of breeding activity by new albatross species have been suspected from aerial surveys at other Chilean colonies, such as two potential nesting Salvin’s albatrosses at Islotes Evangelistas, approximately 52ºS (Robertson et al. 2017). However, the last aerial survey of this location took place as long ago as October 2014.
It is hoped that our land-based survey mentioned here can serve as a starting point for updating the diversity and status of albatrosses in colonies distributed in the exposed waters off Chile, to recognize the presence of new albatross species, and to study their natural history, recruitment rates, and other demographic parameters at their natal sites.
References
Abbott CL, Double MC (2003a) Phylogeography of shy and white-capped albatrosses inferred from mitochondrial DNA sequences: implications for population history and taxonomy. Mol Ecol 12:2747–2758. https://doi.org/10.1046/j.1365-294X.2003.01944.x
Abbott CL, Double MC (2003b) Genetic structure, conservation genetics and evidence of speciation by range expansion in shy and white–capped albatrosses. Mol Ecol 12:2953–2962. https://doi.org/10.1046/j.1365-294X.2003.01980.x
Agreement on the conservation of albatrosses and petrels (2011) ACAP species assessment: White-capped Albatross Thalassarche steadi. https://acap.aq/
Albores-Barajas YV, Soldatini C, Bambini G, Favilli E (2020) One swallow does not make a summer, but could a laysan albatross pair make a colony at natividad Island, Mexico? Oryx 54:13–14
Arata J (2003) New record of Salvin’s albatross (Thalassarche salvini) at the Diego Ramirez Islands, Chile. Notornis 50:169–171
Baker GB, Double MC, Gales R et al (2007) A global assessment of the impact of fisheries-related mortality on shy and white-capped albatrosses: conservation implications. Biol Conserv 137:319–333. https://doi.org/10.1016/j.biocon.2007.02.012
Baker GB, Jensz K, Cunningham R et al (2022) (2023) Population assessment of White-capped albatrosses Thalassarche steadi in New Zealand. Emu. https://doi.org/10.1080/01584197.2022.2161915
Baker GB (2016) Demography of shy and white-capped albatrosses: conservation implications. University of Tasmania. PhD Thesis. https://doi.org/10.25959/23239178.v1
Bartle JA (1991) Incidental capture of seabirds in the New Zealand subantarctic squid trawl fishery, 1990. Bird Conserv Int 1:351–359. https://doi.org/10.1017/S0959270900000708
Chown SL, Gaston KJ, Williams PH (1998) Global patterns in species richness of pelagic seabirds: the procellariiformes. Ecography 21:342–350. https://doi.org/10.1111/j.1600-0587.1998.tb00399.x
Coulson JC (2016) A review of philopatry in seabirds and comparisons with other waterbird species. Waterbirds 39:229–326. https://doi.org/10.1675/063.039.0302
Danckwerts DK, Humeau L, Pinet P et al (2021) Extreme philopatry and genetic diversification at unprecedented scales in a seabird. Sci Rep 11:6834. https://doi.org/10.1038/s41598-021-86406-9
Falla RA (1965) Birds and mammals of the subantarctic islands. Proc NZ Ecol Soc 12:63–68
Fisheries New Zealand (2020) national plan of action-seabirds 2020. reducing the incidental mortality of seabirds in fisheries. 21. https://www.mpi.govt.nz/
Flux IA (2002) New Zealand white-capped mollymawk (Diomedea cauta steadi) chicks eaten by pigs (Sus scrofa). Notornis 49:175–176
Hedd A (1999) Foraging ecology of shy albatrosses Thalassarche cauta breeding in Australia: implications for interactions with longline fisheries. university Of Tasmania
Jouventin P (1990) Shy albatrosses diomedea cauta salvini breeding on Penguin Island, crozet archipelago, Indian Ocean. Ibis 132:126–127
Lawton K, Robertson G, Valencia J et al (2003) The status of black-browed albatrosses Thalassarche melanophrys at diego de almagro Island, Chile. Ibis 145:502–505. https://doi.org/10.1046/j.1474-919X.2003.00186.x
Miskelly CM (1984) Birds of the western chain, snares Islands, 1983–84. Notornis 31:209–223
Miskelly CM, Elliott GP, Parker GC et al (2020) Birds of the Auckland Islands, New Zealand subantarctic. Notornis 67:59–151
Phalan B, Phillips RA, Double MC (2004) A White-capped albatross, Thalassarche [cauta] steadi, at South Georgia: first confirmed record in the south-western Atlantic. Emu 104:359–361. https://doi.org/10.1071/MU03057
Phillips RA, Cooper J, Burg TM (2018) Breeding-site vagrancy and hybridization in albatrosses. Ibis 160:907–913. https://doi.org/10.1111/ibi.12622
R Core Team (2022) R: A language and environment for statistical computing. R foundation for statistical computing. https://www.R-project.org/.
Richard Y, Adasme L (2019) Assessment of the risk of trawl and longline fisheries to ACAP-listed seabirds in Chile. Florianópolis
Robertson G, Moreno CA, Lawton K et al (2007) An estimate of the population sizes of black-browed (Thalassarche melanophrys) and grey-headed (T. chrysostoma) albatrosses breeding in the Diego Ramírez Archipelago. Chile Emu 107:239–244. https://doi.org/10.1071/MU07028
Robertson G, Moreno C, Arata JA et al (2014) Black-browed albatross numbers in Chile increase in response to reduced mortality in fisheries. Biol Conserv 169:319–333. https://doi.org/10.1016/j.biocon.2013.12.002
Robertson G, Wienecke B, Suazo CG et al (2017) Continued increase in the number of black-browed albatrosses (Thalassarche melanophris) at Diego Ramírez, Chile. Polar Biol 40:1035–1042. https://doi.org/10.1007/s00300-016-2028-5
Robertson C (1985) Reader’s digest complete book of new zealand birds, 1st edn. reader’s digest services Pty limited
Ryan PG, Jones MGW, Dyer BM et al (2009) Recent population estimates and trends in numbers of albatrosses and giant petrels breeding at the sub-Antarctic prince Edward Islands. Afr J Mar Sci 31:409–417. https://doi.org/10.2989/AJMS.2009.31.3.13.1001
Schlatter RP, Riveros GM (1987) Historia natural del Archipiélago Diego Ramírez, Chile. Serie Científica INACH 47:87–112
Schüttler E, Crego R, Saavedra L et al (2019) New records of invasive mammals from the sub-Antarctic Cape Horn Archipelago. Polar Biol 42:1093–1105
South A (2011) Rworldmap: a new R package for mapping global data. R J. https://doi.org/10.32614/rj-2011-006
Tennyson AJD (2020) Variation in the bill colour of the white-capped mollymawk (Thalassarche cauta steadi). Notornis 67:333–340
Tickell WLN (1976) The distribution of black-browed and grey-headed albatrosses. Emu 76:64–68. https://doi.org/10.1071/MU9760064
Tickell W (2000) Albatrosses. Yale University Press
Wickham H (2016) ggplot2: elegant graphics for data analysis. Springer-Verlag, New York
Acknowledgements
We are grateful for the significant support provided by the Ambassade de France au Chili & L’Institut Français du Chili. We would like to extend special thanks to Claire Boudy for her invaluable assistance throughout this process. We also express our gratitude to Jennifer Torres and Ricardo Rozzi of Fundación Omora and the Cape Horn International Center (CHIC) at the Universidad de Magallanes, Chile. Our appreciation extends to all personnel of the III Naval Zone based in Punta Arenas, as well as the officers and crew members of OPV-83 Marinero Fuentealba (Cmte. CF Luis Bustos González, 2° Cmte. CC Benjamín Paredes Benavides, Subof. Boris Aedo Moya, Sgto. 1° Rodrigo Zavala Salgado, and Sgto. 2° Cristian Queipul Queipul). We also thank the Gobernación Marítima Punta Arenas–Chilean Navy, with special recognition to the lighthouse crew members based on Isla Gonzalo (Sgto. 1° Ansen Moreira Vargas, Sgto. 2° Cristian Villa Montecinos, and Cbo. 1º E. David Osorio Donoso). Special acknowledgments are due to Rachael Alderman, Kris Carlyon, Colin Miskelly, and Sandy Bartle for their valuable contributions in providing descriptions, audio records, and identification tips regarding the shy-type albatrosses from Tasmania and New Zealand colonies. We also extend our thanks to Graham Robertson for sharing the aerial view of the Diego Ramírez Islands and providing key suggestions for improving our manuscript. We express our appreciation to two anonymous reviewers whose valuable input has greatly contributed to the enhancement of this manuscript. Finally, we dedicate this study to the memory of Professor Roberto P. Schlatter, whose pioneering work significantly contributed to our understanding of albatrosses in the Diego Ramírez Islands.
Funding
Open Access funding enabled and organized by Projekt DEAL. Ministère de l’Europe et des Affaires Étrangères, Délégation régionale de coopération pour l’Amérique du Sud, Ambassade de France au Chili & L’Institut Français du Chili, project “The grey-headed and the black-browed albatrosses in the Diego Ramírez Archipelago: Breeding habitat selection and their fragility in the face of a changing climate.” Cape Horn International Center (CHIC), Chile (ANID CHIC-FB210018).
Author information
Authors and Affiliations
Contributions
CGS conceived and designed the study. CGS, CA, and CGL conducted the fieldwork. AM conducted the lab work for genetic identification. CGS drafted the manuscript, while CA developed the mapping and graphs. CA, CGL, AM, and PQ reviewed, contributed to, and approved the final draft of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Supplementary file3 (MP4 173883 KB)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Suazo, C.G., Anguita, C., Letelier, C.G. et al. The white-capped albatross: a new breeding record for the Diego Ramírez Islands, Chile. Polar Biol 47, 771–777 (2024). https://doi.org/10.1007/s00300-024-03269-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-024-03269-2