
Abstract
In a landscape with patchily distributed forage resources, an animal’s distribution may reflect the distribution of the resources if the population is forage-limited in time or space. This may be particularly explicit in climatically extreme and seasonally variable environments, notably alpine and polar environments during winter. Sustainable management considers the amount of available alpine ground lichen in winter as a predictor of carrying capacity for the last remaining populations of wild European Mountain reindeer Rangifer tarandus tarandus in alpine environments in South Norway. Ground lichen growth is constrained to elevated, wind-blown, snow-free ridges. However, wild Mountain reindeer also persist in areas providing little lichen resources. Our alternative functional hypothesis is that the critical resource is the amount of snow-free feeding areas during winter, which provide direct access to forage, likely to be energy-profitable regardless of the type of vegetation. We sampled animal-borne videos from wild Mountain reindeer during maximum accumulation of snow in late winter/spring, in three contrasting areas providing variable amounts of ground lichen and alternative vegetation resources. Direct observations of reindeer foraging via videos document an active selection and strong preference for feeding on snow-free patches, regardless of type of vegetation. Active ‘cratering’ in snow was not observed. In contrast, walking behaviours occurred much more frequently across snow-covered areas. Remote sensing data and analyses corroborated these observations. In the sustainable management of wild Mountain reindeer amounts of vegetated snow-free areas is an important functional predictor of winter carrying capacity.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Animals living in fluctuating and extreme, changing environments may adapt via plasticity to cope with environmental variability (West-Eberhard 2003; Higginson et al. 2012; Boutin and Lane 2014) or genetically by counter-gradient evolution (Quinby et al. 2020). Behavioural plasticity is important for an organism’s potential to adapt in rapidly changing environments (Snell-Rood 2013; Groothuis and Taborsky 2015). Behavioural choices of individual animals influence their growth, survival and reproduction, and therefore the ecology of populations (Sutherland 1996; Clark and Mangel 2000; DeAngelis 2018). Available foraging resources in space and time and corresponding animal foraging behaviours, determine forage intake and cost, i.e., net energy gain. Optimal foraging theory posits that foraging behaviours optimize net energy gain (MacArthur and Pianka 1966; Pyke 1984; Perry and Pianka 1997). Therefore, distribution of an organism between resource sites may be expected to match the distribution of the resources (Ideal Free Distribution model) (Fretwell and Lucas 1970; Sutherland 1983; Křivan et al. 2008). This pattern may, however, be modified by organism discrimination constraints, competitive interactions, competitive asymmetries, and travel between sites (Abrahams 1986; Kennedy and Gray 1993; Calsbeek and Sinervo 2002), or simply haphazard choices.
Northern ecosystems are variable environments in space and time, typically providing patchy and sometimes lush foraging habitats in the summer growth season, whereas the long winter is often the critical ‘season of discontent’. Animal populations may be limited by available forage resources, for example in ungulates, with mortality often aggravated by predation (e.g., Sæther and Andersen 1990; Patterson and Power 2002; Beschta et al. 2014). The cold-adapted northern reindeer Rangifer tarandus is the most widely distributed mammalian herbivore in the extreme high-latitude alpine and sub-polar environments (Pruitt 1959; Skogland 1983, 1984). The usually low production and patchy distribution of high quality vegetation resources in these ecosystems, dictates the typical foraging strategies exhibited by reindeer; they graze extensively and are almost constantly in motion, with some of the longest ungulate migrations known (Johnson et al. 2001; Falldorf et al. 2014; Joly et al. 2019). During winter, foraging resources are even more patchy and limited, depending on snow and ice cover. For the last remaining wild Mountain reindeer Rangifer tarandus tarandus in Western Europe, living in South Norway, availability of food resources in winter, notably lichen, is considered the factor limiting reindeer populations (Gaare and Skogland 1975, 1980; Skogland 1984). Therefore, as established in the current environmental quality standard for wild reindeer (Kjørstad et al. 2017) populations in Norway are (apart from their status, habitat, and amount of human disturbance) managed primarily by available lichen resources. This may, however, be a question of availability rather than necessity. As noted by Bergerud (1996): ‘The hypothesis that winter lichen supplies determine abundance and set the carrying capacity is rejected. Lichens are not necessary food for caribou.’ Indeed, wild reindeer also persist in areas in Norway which provide limited lichen coverage (Kastdalen 2011), but where snow-free areas with other types of vegetation are accessible, for example on south-west facing, sun-exposed steep slopes.
In alpine and sub-polar environments, vegetation patterns integrate snow conditions across space and time, reflecting the ‘normal’ snow situation (Dahl 1956; Hesjedal 1975; Odland and Munkejord 2008). This normal situation largely depends on prevailing wind directions, blowing snow away from exposed areas into terrain leesides. Thus, annual snow distribution patterns, including snow-free or low snow-depth patches, tend to be relatively stable (Sturm and Wagner 2010). Snow cover is, in addition to temperature (which is also affected by snow cover), the main physical factor forming plant communities in alpine areas, as it decouples alpine plant life from atmospheric conditions (Körner and Hiltbrunner 2021). The slow-growing ground lichens are favoured on snow-free or low snow-depth low-alpine landscape patches (Dahl 1956; Gjaerevoll 1956). It is their physiological adaptation, such as an extremely high frost tolerance while still maintaining photosynthetic capacity (Dahl 1954; Kappen and Lange 1972), which allows lichens even to dominate within those areas where rigorous environmental conditions weaken the competition by vascular plants (Lechowicz and Adams 1974). As such, ground lichens prevail within exposed, wind-blown and, thus, snow-free habitats, which are—due to the lack of an insulating snow cover—characterized by an extended period of deeply frozen soil (Odland and Munkejord 2008; Odland et al. 2018; Löffler and Pape 2020).
However, we propose it may not be lichen resources per se that are crucial for population winter carrying capacity in wild Mountain reindeer, but rather the availability of more or less snow-free areas providing easily accessible forage. Indeed, it is a common plant adaptation in alpine regions to store nutrients and organic materials closer to the growing points above ground, e.g., to keep (dead) leaves during winter for insulation, or to preform (flowering) buds already during late summer and autumn for earlier sprout and flowering in spring the next year (Wielgolaski and Goodall 1997). As such, snow-free habitats, irrespective of their lichen cover, likely provide energy-efficient foraging habitats. This hypothesis was indirectly supported by remote sensing data (Romtveit et al. 2021), suggesting that wild Mountain reindeer were located on or close to snow-free patches much more frequently than expected from a ‘random walk’. Therefore, we predict direct observations of reindeer foraging behaviours will document that Mountain reindeer preferably feed on snow-free patches, and primarily walk when on snow. Unfortunately, systematic and extensive direct field observations are impractical to implement. An alternative for direct observations is to fit wild reindeer with video cameras (Newmaster et al. 2013; Thompson et al. 2015). Here we use direct observations from animal-borne remote videos across extended time in late winter with maximum snow accumulation (March–April) and across three areas with contrasting amounts of lichen resources, to test the hypotheses that (1) wild Mountain reindeer primarily forage on snow-free patches during winter, regardless of available vegetation, and (2) primarily move across snow.
Materials and methods
In a case-series and use-versus-available study design (Manly et al. 2002), the two main types of behaviour, feeding and walking, were associated with percentage (10% increments) of snow cover/snow-free patches, as estimated by direct observation from video recordings, representative of visually based foraging during day/twilight hours. At night, resting behaviours are prevalent (Collins and Smith 1989).
Videos and behavioural data
Video data amounted to 2076 scenes (separate video recordings) with different behaviours (number of times exhibited within a scene), from five reindeer individuals across three separate reindeer areas with contrasting vegetation (Hardangervidda, Setesdal-Austhei, Nordfjella), and during an extended time period in late winter/spring (Fig. 1, Tables 1, 2). In Scandinavian mountains, snow cover is typically most extensive in late April (Kohler et al. 2006; Lawrence and Slater 2010). Snow cover (thickness and duration) and snow melt may vary across years, but snow-free areas tend to be similar across winters for topographic and wind reasons (Kohler et al. 2006; Odland et al. 2018), and the corresponding snow melt patterns remain remarkably consistent among microsites (Gjaerevoll 1956; Billings and Bliss 1959). Modelling of snow cover based on remote sensing data (Romtveit et al. 2021), indicated snow melt starting to accelerate during the last week of May in all three areas and in both study years (2012, 2017).

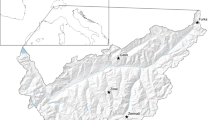
Areas. Wild Mountain reindeer Rangifer tarandus tarandus areas in South Norway (Inset: location in Europe). Green areas constitute the two prioritized larger European wild Mountain reindeer regions. The three areas studied are in the south of the European region (ca. 58° 33′ 21′′–60° 46′33′ N, 6° 43′ 32′′–8° 56′ 54′′ E): 7 = Hardangervidda (8130 km2), 2 = Setesdal Austhei (2400 km2), and 11 = Nordfjella (3000 km2)
Of the studied areas, Hardangervidda generally has considerably more lichen resources than especially Setesdal Austhei (Table 1) (Kastdalen 2011; Falldorf et al. 2014; Odland et al. 2014). The Hardangervidda lichen resources are, however, variable (see SD in Table 1), particularly in the more continental eastern parts, and with more resources in areas disturbed by human infrastructure and therefore less used by wild reindeer. In Setesdal Austhei lichen height and volume was low in all areas studied (Table 1).
The videos used here, are from two female reindeer each on Hardangervidda and Setesdal Austhei (Fig. 1, Table 2), fitted with collared video cameras type Vectronic GPS Plus with a Müvi camera unit (https://www.yumpu.com/en/document/view/5136810/gps-plus-collar-manager-vectronic-aerospace -gmbh) during the late winter/spring periods April–May 2012, and one female reindeer in Nordfjella in 2017 (permits from the Norwegian Food Safety Authority FOTS: ID 15116, license # 19/20935) (for details see Strand et al. 2006, 2011, 2015). Females tend to be more representative of the herd and population demography (Strand 2009; Strand et al. 2015). Case individuals were limited by logistics (access, tagging time window, economy). Using a helicopter, groups of reindeer were identified, and a random female was immobilized with a mix of medetomidine (12 mg) and ketamine (120 mg) in 3 ml arrows fired from a Danarms℗ CO2- air rifle into the animal’s thigh. Blood, hair, and tissue samples were collected before the drop-off collar with video, radio and GPS units was fitted. Temperature, pulse, respiration, and blood O2 were monitored during the operation. Constriction was avoided by fitting collars with four finger-widths (male hands) of slack and the presence of a cotton weak zone.
Based on the video recordings, and previous studies (Gaare et al. 1975; Thomson 1978; Collins and Smith 1989; Mårell 2006), reindeer behaviours were classified into 42 different behavioural categories, of which 30 were observed in the present study (Fig. 2). Duration of each type of behaviour was measured. The 2076 different scenes with visible and audible behaviours amounted to 19 h and 41 min of video material (Table 2). The two dominant behaviours were active feeding (class 11, also considering class 10 Digging/cratering) and walking (combined class 7 Walking-locally, and class 8 Walking-directed) and are both relevant to the present study. Visible snow cover per classified behaviour was estimated (consistently by the same person) from the video shot with the largest local area perspective relevant the classified behaviour, and in 10% increments. Because data tended to be bimodal (either near no snow or near complete snow cover), we also included a 5% and a 95% category. For logistic and local project reasons, preset recording frequency and duration varied between individuals (Table 2), but was mainly 3 min, and with a maximum of 22 scenes in one 3-min-recording. There were, however, some much longer recordings in Setesdal Austhei. To better balance data by individuals, behavioural data from the longer recordings were truncated at 22 ‘scenes’ of 3 min before analyses (Table 2).

Behaviours. Number of times different classified behaviours were observed in recorded videos (n = 2076 scenes) from five wild Mountain reindeer Rangifer tarandus tarandus during maximum snow cover in late winter/spring (April–May) across three wild reindeer areas in South Norway. The dominant behaviours are feeding (10–11) and walking (7–8). Observed behavioural classes were: 1 Only fur visible, 2 Lying/resting, 3 Lying (listening/looking), 4 Lying (ruminating), 5 Standing (listening/looking), 6 Standing (ruminating), 7 Walking (locally), 8 Walking (targeted), 9 Running, 10 Digging, 11 Eating, 13 Fixing her fur, 14 Scratches herself, 22 Shaking head/body, 23 Rising, 24 Eating snow, 25 Drinking, 26 Stretching (sound, shake, yawn), 27 Getting scared, 28 Standing (relaxing), 29 Sudden directional change, 30 Headbutting, 31 Stretching (whole body), 33 Laying down, 34 Black screen, 35 Sweeping antlers, 36 Sneezing, 38 Smelling her calf, 40 Yawning, 41 Heavy breathing, 42 Licking her calf
Resource selection and remote sensing data
Remote sensing data and analyses were used to estimate snow cover and available feeding resources within the areas used by the individual reindeer (used area estimated as Maximum Convex Polygon (MCP) from GPS positions) (Table 3). Snow cover (%) was estimated from cloud-free Sentinel-2 scenes. For the study areas Hardangervidda and Setesdal Austhei, with GPS data from 2012, Sentinel-2 scenes were selected from years where snow depths followed the same pattern as in 2012 measured from the daily snow-depth estimates of the Norwegian Water Resources and Energy Directorate (NVE) snow model in 1 km grid (Saloranta 2012). For the area Nordfjella, with GPS data from 2017, Sentinel-2 data from the same year were used.
For April the estimates of snow cover were derived from the same reference median date, since snow conditions were relatively stable with snow melt starting in late April according to the NVE data. For May, snow cover were estimated from scenes centered on three time periods: 1–15, 16–25 and after 25th. The snow cover estimates, with a 20 × 20 m resolution, (Table 3), were derived from a local snow-cover model based on independent data from World-View 2 and 3 with acquisitions in April and May (30 and 40 cm pixel size, 4 subscenes of 25 km2 sampling within Hardangervidda). The local model was developed following the approach in Gascoin et al. (2020).
Cover of green vegetation (%), lichen coverage (%), and barren ground (%) (Table 3) were estimated earlier from a Landsat 5 scene (Kastdalen 2011). The Normalized Difference Vegetation Index (NDVI) is an indicator of vegetation greenness which was used to indicate general differences in vegetation cover across the three areas (Table 3). NDVI was estimated with Google Earth Engine (Gorelick et al. 2017) as the magnitude of a five year harmonic model using Sentinel-2 images (Derwin et al. 2020). Remote sensing data were also used to explore resource selection patterns and compare with direct video observations of feeding and walking behaviours. Because distances between resources were relatively short, we used Step Selection Analysis (SSA) (Thurfjell et al. 2014; Fieberg et al. 2021). It compares environmental attributes of observed steps (the linear segment between two consecutive observations of position) with alternative random steps taken from the same starting point. For each GPS position observed, we modelled resource availability by randomly choosing step angle from a von Mises distribution and step length from a gamma-distribution (Signer et al. 2019). The estimated resource availability was then compared with the next GPS position used by the reindeer (the selected spot), and analysed with a used:available relation of 1:15 (Avgar et al. 2016). All points on water and outside the MCP, neither used by reindeer, were removed. Data were centered and standardized (Schielzeth 2010) before analysed in a conditional regression using the clogit function in the R survival package (Therneau and Grambsch 2000), where each used GPS point and associated availability points constituted a stratum in the model formula. The environmental variables used to explore potentially influencing reindeer movement to snow-free areas, were (Table 4): snow cover (%), slope (10 m grid), relative topographic position, i.e., elevation of GPS position relative to mean elevation within a small 50 m radius landscape and a larger 500 m radius (10 m grid) (Lindsay 2014; Newman et al. 2018), number of days to peak phenology estimated from the phase component of the harmonic model, as a proxy for early development of vegetation in snow-free areas, magnitude of NDVI, and solar radiation from the period March 15th to May 15th. In addition, the behavioural variable step length, the distance between each GPS position (log meters), was included in the model. The individual variance inflation factors (VIF’s) were all less than 1.7.
Statistical analyses were implemented in R v. 4.2 (R Core Team 2022), emphasizing model simplicity (Murtaugh 2007; Kass et al. 2016).
Results
Behaviours
Across the in total 2076 scenes from direct observations, the two dominant behaviours were feeding (30% of all recorded behaviours, 47% of time) and walking (32 and 17%) (Fig. 2). Local walking constituted 21% of recorded behaviours, but only 8% of time, and directed walking only 11% of recorded behaviours, and 9% of recorded time. Observations of direct feeding constituted virtually all feeding time, whereas digging/cratering associated with feeding was barely observed, less than 1% of the time (2 min 31 s). Moreover, digging was almost exclusively associated with ‘edge’-digging, i.e., into the snow edges of snow-free patches. None of the video borne female reindeer exhibited ‘classic’ cratering in snow-covered areas. The closest was one scene where the reindeer used their muzzle to shovel off new snow from raised rocks to feed on the underlying rock surface lichen (Hardangervidda, ID 33, scene 51, May 4, 2012).
Neither feeding, nor walking was a ‘random walk’ relative to snow conditions (Fig. 3) (χ2 homogeneity test, χ212 = 1677.1 and χ211 3063.7, P < 0.0001). There was a strong association between feeding behaviours and no or low snow cover, and a corresponding, but opposite association between walking and high snow cover (χ2 association test, χ24 = 276.41, P < 0.0001). The reindeer chose to walk to bare patches, rather than to dig in snowy patches. Video feeding scenes were almost exclusively associated with no or little snow cover, with 79% of the scenes with no or snow cover less than or equal to 5% (Fig. 3, top). Walking was typically associated with very high or complete snow cover (Fig. 3, bottom). However, the strong association between feeding and snow-free patches, also implied that at substantial number of walking scenes were associated with snow-free areas (33.5% with less than or equal to 5% snow cover, Fig. 3, bottom). A typical pattern from the longer videos in the Setesdal Austhei area, was alternating scenes of feeding on snow-free patches, then walking across snow to feed on the next snow-free patch. For example, one of the longest continuous scenes of alternating feeding and walking (ID 50 in Setesdal Austhei, Table 2) lasted 19 min 51 s, with feeding exclusively on snow-free (less than 5%) patches (18 scenes, 13 min 53 s), and walking across snow (more than 95%) to the next patch (18 scenes, 5 min 24 s). The only interruptions were three short scenes of ‘standing’ (i.e., to listen/watch out; 3 scenes, 34 s) (Fig. 4).

Snow. Frequency of feeding relative to snow cover (a) and of walking relative to snow cover (b)

Individuals. Frequency of feeding (a) and walking (b) behaviours across the five individuals. The individuals exhibited similar behavioural patterns, but individual 231 Nordfjella deviated somewhat with less walking. Individual 31 contributed with only 9 scenes, all feeding
Across wild reindeer areas, the relative frequencies of feeding and walking behaviours were virtually identical in the Hardangervidda and Setesdal Austhei areas (79 and 119 resp. 486 and 492 scenes) (Fig. 5). The reindeer in Nordfjella, however, exhibited a lower frequency of walking (51 feeding and 23 walking scenes). Based upon the elevation derived from GPS positions, individuals utilized the area from the sub-alpine birch forest and upwards, well into the mid-alpine belt. Due to the general decrease of the elevational vegetation belts towards the coast, this corresponds to 895–1641 (mean 1384) meters above sea level for Nordfjella, 936–1545 (mean 1280) meters above sea level for Hardangervidda, and 655–1356 (mean 929) meters above sea level for Setesdal Austhei. Remote sensing data indicated high snow cover in all three areas (Table 3) until mid-May, thereafter, snow melt progressed rapidly, and depending on elevation (Table 3). These results also indicate substantial differences in availability of green vegetation, both in summer and winter, and in lichen coverage within the areas utilized by the studied reindeer. Especially Setesdal Austhei provides considerably more green vegetation and less lichen, almost absent, compared to Hardangervidda (Table 3).

Winter foraging. Still images from video (ID 11411/50, scene 41, Setesdal Austhei) indicating preference for snow-free foraging areas consistent across many more Mountain reindeer Rangifer tarandus tarandus in addition to the video-tagged individuals. The female continuously walked across 100% snow cover to feed on 0–5% snow cover patches. In this image, we see how the other animals have the same behaviour, as the female approaches the snow-free area
Resource selection
Analyses of resource selection based on remote sensing data corroborated the behavioural data analyses from video scenes, indicating a preference for feeding on elevated patches with little snow cover (Table 4), but also reflected differences among areas, and individual variation, which tends to weaken potential patterns across areas, e.g., step length. However, step length was less than expected in Setesdal Austhei in May (Table 4), possibly reflecting the rapidly increasing amount of snow-free patches particularly in that area in May (Table 3). All reindeer consistently avoided areas with snow, except one individual in Setesdal Austhei, which also moved less than the other individuals (Table 4). Reindeer also consistently avoided (steeply) sloping areas, and were rarely observed in locations with slopes exceeding 30° (Slope in Table 4). Tagged reindeer were almost never tracked in areas with more than 30° slope. Within these local and relatively small-hilly landscapes, the reindeer preferred to be on the ridges (Topo50m in Table 4), and this preference for elevated, and presumably less snow-covered areas, was seen also on a larger 500 m landscape scale in April (Topo500m in Table 4). This tendency was, however, weakest in Setesdal Austhei and as spring progressed, as they tended to use more the gentle south-west facing slopes with higher solar radiation and earlier developed vegetation (DayPheno in Table 4). The magnitude of the NDVI appeared to be more important in the high-elevation Nordfjella area with its more extensive barren areas. Solar radiation was important in April but weakened as snow melt progressed in May. For one reindeer in Setesdal Austhei and one in Hardangervidda, the importance of solar radiation combined with early developed vegetation, suggest they were foraging more in the hillsides.
Discussion
Direct observations via animal-borne videos document that wild Mountain reindeer, when having a choice, exhibit a strong preference for snow-free habitat patches in winter, at least during maximum snow accumulations and start of snow melt in late winter/spring, i.e., April–May. To the extent that reindeer were observed digging, this only occurred along the edges of snow-free patches. Digging in snow-covered areas, i.e., cratering, was not observed. Cratering comes at an energetic cost (Fancy and White 1985) which may be avoided in alpine areas which alternatively provide snow-free foraging habitat. Also considering the high energetic efficiency of locomotion in wild reindeer (Fancy and White 1987), but clearly depending on snow conditions, the consistent selection of snow-free feeding patches appears to be a behavioural adaptation to the alpine environments providing such snow-free habitats. The benefit of this optimal foraging strategy will, however, likely depend on the amount and spatial distribution of snow cover and such snow-free patches. Reports of cratering are, to our knowledge, primarily from areas with more continuous and loose snow cover. The cratering alternative depends on snow conditions (LaPerriere and Lent 1977; Skogland 1978; Fancy and White 1985), to reach plant cover. Reindeer try to reduce energetic cost when digging for food in winter by avoiding areas with deep or hard snow (Skogland 1984). Cratering also has a limited potential net energy gain considering the small vegetation area uncovered, again depending on snow conditions (LaPerriere and Lent 1977; Helle 1984; Kumpula et al. 2004). Snow-free patches likely offer larger and easily available forage. Previous studies of reindeer have also noted a preference for elevated ridge areas and the correlated ground lichen forage in winter/spring (e.g., White and Trudell 1980; Swanson and Barker 1992; Gaare 1997; Mårell and Edenius 2006; Pape and Löffler 2015).
Although the number of tagged individuals was limited by logistics, we note that results were consistent with predictions—across individuals, albeit with some individual variation, across stratified contrasting areas, and across time. Moreover, in the videos many more reindeer, not included in the analyses, showed the preference for snow-free foraging areas (Fig. 5). Our direct observational data are from the late winter/early spring period. This period may be particularly important for evaluating carrying capacities based on lichen resources (Skogland 1985; Gates et al. 1986; Nellemann 1996). It exhibits maximum snow accumulation and starting snow-melt, a period also typically providing more dense and packed snow (Tucker et al. 1991). This will favour easy reindeer locomotion, but not cratering behaviours. If we had observations from early winter with less and more loose snow cover, we may have observed more cratering.
Based on our direct observation results, we argue that amount and spatial distribution of snow-free (and low snow-depth) patches is an important functionally-based predictor of winter carrying capacity for wild Mountain reindeer, because it directly reflects forage availability at low energetic cost. That does not mean that reindeer would be unable to graze without such patches, e.g., when compared to reindeer who do crater throughout winter, e.g., in semi-domesticated reindeer in lowland areas (Kumpula et al. 2004). But that would come at an energetic and population carrying capacity cost, e.g., in mountains with deeper and more packed snow. Lichen resources generally considered the favoured and critically important reindeer winter forage (reviewed in e.g., Bergerud 1996; Heggberget et al. 2002; Joly et al. 2009) and is the key indicator of winter carrying capacity in the sustainable management of wild Mountain reindeer in Norway (Kjørstad et al. 2017; Rolandsen et al. 2022). We suggest this is a correlation, rather than a direct cause-and-effect. As indicated by Bergerud (1996), lichens are not a necessary food for reindeer, and winter lichens supplies do not necessarily set the carrying capacity for reindeer, if alternative forage is available. It is well documented that reindeer populations may be sustained also in areas not providing lichens (Leader-Williams et al. 1981; Bergerud 1996; Heggberget et al. 2002; Hansen et al. 2009). Wild reindeer may eat a mixture of lichens, mosses and vascular plants such as shrubs and graminoids also in winter (Skogland 1984; Mathiesen et al. 2000; Heggberget et al. 2002; Storeheier et al. 2002), but depending on availability (e.g. Johnson et al. 2001; Bremset Hansen et al. 2009). We suggest forage availability may be best predicted via amounts of vegetated snow-free patches in alpine areas, and not restricted to lichen resources only. Snow-free patches also have the considerable benefit of being estimable via remote sensing data (Romtveit et al. 2021).
Snow cover (thickness and duration) is considered the most important ecological factor in alpine areas, affecting soil temperature and moisture, and duration of the growing season, which in turn control plant distribution in the alpine environment (Burns et al. 1982; Williams et al. 1998). The distribution of ground lichen in alpine areas correlates strongly with availability of snow-free habitats, i.e. typically exposed, wind-blown patches on elevated ridges experiencing an extended period of (severe) frost within the soil (Odland and Munkejord 2008; Odland et al. 2018; Löffler and Pape 2020). In the one area studied here (Setesdal Austhei), lichen forage resources may be limited, suggesting low winter carrying capacity. However, the area still provides snow-free habitats, mostly exposed tops and south-west facing, sun-exposed steep slopes, dominated by vascular plants (e.g., Betula nana, Empetrum nigrum, Vaccinium vitis-idaea, Juncus trifidus, Carex bigelowii), which are grazed by reindeer. This may also provide for a more optimal winter diet for reindeer.
A high correlation between lichen and snow-free patches ensures that a focus on lichen as key winter forage resources will work well much of the time. But it will be inherently biased, in alpine and polar areas, underestimating the importance of snow-free patches providing alternative forage. We propose such patches may be of particular benefit to reindeer in winter. Lichens are rich in carbohydrates, but low in nitrogen and minerals (Nieminen and Heiskari 1989; Storeheier et al. 2002). Reindeer have several physiological adaptations to a low-protein and low-mineral winter diet (McEwan and Whitehead 1970; Hyvärinen et al. 1977; Staaland et al. 1986), when feeding on lichen. Nevertheless, reindeer must take nitrogen and minerals from their body reserves to digest lichen, and lose body mass (Finstad and Kielland 2011), maybe as much as 6.3 g body protein per 1000 g dry lichen (Jacobsen and Skjenneberg 1975). Rognmo et al. (1983) observed higher neonatal calf mortality in captive female reindeer fed lichen only, but ad libitum, in late pregnancy (28% vs. 7% in control). In a functional response, reindeer recycle the lighter nitrogen in the waste product urea and use it in place of dietary protein, and reindeer more depleted in nitrogen reserves may compensate by increasing consumption of high-protein green buds and catkins during late winter/spring (Finstad and Kielland 2011). On an anecdotal basis, Eastern Russian Chukchi reindeer herders working in Alaska, well aware of reindeer’s strong preference for human urine (urea), would openly urinate as an efficient measure to collect the herd (from Jackson’s diary, in Andersen 2011). In an adaptive context, it may therefore seem a bit puzzling that reindeer appear to favour the carbohydrate-rich lichen (Danell et al. 1994; Heggberget et al. 2002; Vistnes and Nellemann 2008; Hansen et al. 2010), suggesting that factors other than nutrition influence diet choice, e.g., palatability. A non-optimal diet, especially with respect to proteins, has also been reported for the snow-free season for woodland caribou, based on animal-borne video recordings (Thompson et al. 2015). In contrast to lichens, new growth of vascular plants like graminoids, dwarf shrubs, and Betula sp. have high levels of crude protein and minerals (Storeheier et al. 2003; Ophof et al. 2013), and as an adaptation to the extremely short growing season in alpine and polar regions, it is more economic for plants to store more nutrients and organic materials (in protected buds) closer to the growing points above ground, often as green leaves during winter (Wielgolaski and Goodall 1997), providing accessible winter forage. More vascular plants forage would therefore be of great benefit to reindeer particularly during winter, providing a better nutritionally-balanced diet (Heggberget et al. 2002). Warenberg (1982) found that many plants growing in snow-poor habitats, may have green buds in winter. The preformation of leaf and flower buds in the previous season (i.e., autumn), which then provide fodder resources for reindeer, is a common feature of alpine plants to adapt to the short growing season (Körner 2021).
A potential limitation of our study could be that other herd members (e.g., large males breaking trail at the front of the herd) had cratered to create some small no/low snow patches prior to the arrival of the tagged females with camera. We consider this unlikely. It was never observed, and any cratering animals would have been in front of the tagged females. Also, cratered patches would have been small, which the observed patches were generally not, as illustrated by the photos in Fig. 5.
In conclusion, direct observations via animal-borne video indicate that snow-free and low snow-depth forage patches are strongly selected by wild Mountain reindeer in late winter/spring. Such patches are often dominated by ground lichen, apparently preferred by reindeer, but snow-free patches may also be dominated by vascular plants. They may be particularly important in the reindeer winter diet, contributing protein and minerals. Amount of lichen, traditionally considered important for winter carrying capacity in alpine reindeer areas, tend to correlate with amount of snow-free forage patches, and may therefore in part substitute as a predictor of winter carrying capacity. However, it will be biased, underestimating the importance of other forage resources, when available on snow-free patches. Therefore, amounts of vegetated snow-free patches, e.g., as indicated by a relatively high NDVI index, likely will be a better predictor of winter carrying capacity of wild Mountain reindeer areas, a key consideration in sustainable management.
Data availability
Dryad (https://doi.org/10.5061/dryad.9s4mw6mmr). GPS positions may be viewed at: https://www.dyreposisjoner.no/Account/Login?ReturnUrl=%2F. Developed and maintained by Norwegian Institute of Nature Research (NINA). All data are also archived and publicly available at the Norwegian Wild Reindeer Centre, Skinnarbu, 3660 Rjukan, Norway. If required, we agree to deposit data in other public repositories.
References
Abrahams MV (1986) Patch choice under perceptual constraints: a cause for departures from an ideal free distribution. Behav Ecol Sociobiol 19:409–415
Andersen R (2011) Redningsmenn og lykkejegere—Norske pionerer i Alaska. H. Aschehoug & Co (W. Nygaard), Oslo
Avgar T, Potts JR, Lewis MA, Boyce MS (2016) Integrated step selection analysis: bridging the gap between resource selection and animal movement. Methods Ecol Evol 7:619–630. https://doi.org/10.1111/2041-210X.12528
Bergerud A (1996) Evolving perspectives on caribou population dynamics, have we got it right yet? Rangifer 16:95–116
Beschta RL, Eisenberg C, Laundré JW, Ripple WJ, Rooney TP (2014) Predation risk, elk, and aspen: comment. Ecology 95:2669–2671. https://doi.org/10.1890/13-1624.1
Billings WD, Bliss L (1959) An alpine snowbank environment and its effects on vegetation, plant development, and productivity. Ecology 40:388–397
Boutin S, Lane JE (2014) Climate change and mammals: evolutionary versus plastic responses. Evol Appl 7:29–41
Bremset Hansen B, Herfindal I, Aanes R, Sæther BE, Henriksen S (2009) Functional response in habitat selection and the tradeoffs between foraging niche components in a large herbivore. Oikos 118:859–872
Burns SF, Tonkin PJ, Thorn C (1982) Soil-geomorphic models and the spatial distribution and development of alpine soils. In: Space and time in geomorphology: Binghamton geomorphology symposium, 1982
Calsbeek R, Sinervo B (2002) An experimental test of the ideal despotic distribution. J Anim Ecol 71:513–523
Clark CW, Mangel M (2000) Dynamic state variable models in ecology: methods and applications. Oxford University Press, Oxford
Collins WB, Smith TS (1989) Twenty-four hour behaviour patterns and budgets of free-ranging reindeer in winter. Rangifer 9:2–8
Dahl E (1954) Lichens. Bot Rev 20:463–476
Dahl E (1956) Rondane: Mountain vegetation in South Norway and its relation to the environment. Skrifter Norske Videnskapsakademi i Oslo, Matematisk-Naturvidenskapelig Klasse 3:1–374
Danell K, Utsi PM, Palo TR, Eriksson O (1994) Food plant selection by reindeer during winter in relation to plant quality. Ecography 17:153–158
DeAngelis DL (2018) Individual-based models and approaches in ecology: populations, communities and ecosystems. CRC Press, Boca Raton
Derwin JM et al (2020) Estimating tree canopy cover using harmonic regression coefficients derived from multitemporal Landsat data. Int J Appl Earth Obs Geoinf 86:101985. https://doi.org/10.1016/j.jag.2019.101985
Falldorf T, Strand O, Panzacchi M, Tømmervik H (2014) Estimating lichen volume and reindeer winter pasture quality from Landsat imagery. Remote Sens Environ 140:573–579
Fancy SG, White RG (1985) Energy expenditures by caribou while cratering in snow. J Wildl Manage 49:987–993
Fancy S, White R (1987) Energy expenditures for locomotion by barren-ground caribou. Can J Zool 65:122–128
Fieberg J, Signer J, Smith B, Avgar T (2021) A ‘how to’guide for interpreting parameters in habitat-selection analyses. J Anim Ecol 90:1027–1043
Finstad GL, Kielland K (2011) Landscape variation in the diet and productivity of reindeer in Alaska based on stable isotope analyses. Arct Antarct Alp Res 43:543–554
Fretwell SD, Lucas HL (1970) On territorial behavior and other factors influencing habitat distribution in birds. Acta Biotheor 19:16–36
Gascoin S et al (2020) Estimating fractional snow cover in open terrain from sentinel-2 using the normalized difference snow index. Remote Sens 12:2904
Gates C, Adamczewski J, Mulders R (1986) Population dynamics, winter ecology and social organization of Coats Island caribou. Arctic. https://doi.org/10.14430/arctic2077
Gjaerevoll O (1956) The plant communities of the Scandinavian alpine snow-beds. Det Kgl Norske Videnskabers Selskabs Skrifter 1:405
Gorelick N, Hancher M, Dixon M, Ilyushchenko S, Thau D, Moore R (2017) Google Earth Engine: Planetary-scale geospatial analysis for everyone. Remote Sens Environ 202:18–27. https://doi.org/10.1016/j.rse.2017.06.031
Groothuis TG, Taborsky B (2015) Introducing biological realism into the study of developmental plasticity in behaviour. Front Zool 12:1–14
Gaare E (1997) A hypothesis to explain lichen-Rangifer dynamic relationships. Rangifer 17:3–7
Gaare E, Skogland T (1975) Wild reindeer food habits and range use at Hardangervidda. In: Wielgolaski FE (ed) Eological studies 17 Fennoscandian tundra ecosystems part 2. Springer, Berlin, pp 195–205
Gaare E, Skogland T (1980) Lichen-reindeer interaction studied in a simple case model. In: Reimers E, Gaare E, Skjenneberg S (eds) Proceedings of 2nd international reindeer/Caribou Symposium, Røros, Norway, 1979. Direktoratet for vilt og ferskvannsfisk, Trondheim, pp 47–56
Gaare E, Thomson B, Kjos-Hanssen O (1975) Reindeer activity on hardangervidda. In: Fennoscandian tundra ecosystems. Springer, Berlin, pp 206–215
Hansen BB, Aanes R, Herfindal I, Saether BE, Henriksen S (2009) Winter habitat-space use in a large arctic herbivore facing contrasting forage abundance. Polar Biol 32:971–984. https://doi.org/10.1007/s00300-009-0597-2
Hansen BB, Aanes R, Saether BE (2010) Feeding-crater selection by high-arctic reindeer facing ice-blocked pastures. Can J Zool-Revue Canadienne De Zoologie 88:170–177. https://doi.org/10.1139/z09-130
Haviarová P (2019) Estimation of lichen volume in Setesdal Austhei and Hardangervidda and its relation to microtopographical gradients and distance to human structures Master thesis University of South-Eastern Norway. University of South-Eastern Norway
Heggberget TM, Gaare E, Ball JP (2002) Reindeer (Rangifer tarandus) and climate change: importance of winter forage. Rangifer 22:13–31
Helle T (1984) Foraging behaviour of the semi-domestic reindeer (Rangifer tarandus L.) in relation to snow in Finnish Lapland, vol 19. Kevo Subarctic Research Station, Kevo Subarctic Reserach Institute, University of Turku
Hesjedal O (1975) Vegetation mapping at hardangervidda. In: Wielgolasky FE (ed) Fennoscandian tundra ecosystems. Part 1: Plants and microorganisms. Ecological studies, vol 16. Springer, Berlin
Higginson AD, Fawcett TW, Trimmer PC, McNamara JM, Houston AI (2012) Generalized optimal risk allocation: foraging and antipredator behavior in a fluctuating environment. Am Nat 180:589–603. https://doi.org/10.1086/667885
Hyvärinen H, Helle T, Nieminen M, Väyrynen P, Väyrynen R (1977) The influence of nutrition and seasonal conditions on mineral status in the reindeer. Can J Zool 55:648–655
Jacobsen E, Skjenneberg S (1975) Some results from feeding experiments with reindeer. In: Luick JR, Lent PC, Klein DR, White RG (eds) Proceedings of First International Reindeer and Caribou Symposium 9–11 August 1972. Biol Pap Univ Alaska, Fairbanks, pp 95–107
Johnson CJ, Parker KL, Heard DC (2001) Foraging across a variable landscape: behavioral decisions made by woodland caribou at multiple spatial scales. Oecologia 127:590–602
Joly K, Jandt RR, Klein DR (2009) Decrease of lichens in Arctic ecosystems: the role of wildfire, caribou, reindeer, competition and climate in north-western Alaska. Polar Res 28:433–442
Joly K et al (2019) Longest terrestrial migrations and movements around the world. Sci Rep 9:1–10
Kappen L, Lange OL (1972) Die Kälteresistenz einiger Makrolichenen. Flora 161:1–29
Kass RE, Caffo BS, Davidian M, Meng X-L, Yu B, Reid N (2016) Ten simple rules for effective statistical practice. PLoS Comput Biol 12(6):e1004961. https://doi.org/10.1371/journal.pcbi.1004961
Kastdalen L (2011) Lavkart Setesdal/Ryfylkeheiene og Setesdal Austhei—metodeutvikling og validering av kart, vol 7. Direktoratet for Naturforvaltning, Trondheim
Kennedy M, Gray RD (1993) Can ecological theory predict the distribution of foraging animals? A critical analysis of experiments on the ideal free distribution. Oikos 68:158–166
Kjørstad M et al (2017) Miljøkvalitetsnorm for villrein. Forslag fra en ekspertgruppe, vol 1400. Norwegian Institute for Nature Research, Trondheim
Kohler J, Brandt O, Johansson M, Callaghan T (2006) A long-term Arctic snow depth record from Abisko, northern Sweden, 1913–2004. Polar Res 25:91–113
Křivan V, Cressman R, Schneider C (2008) The ideal free distribution: a review and synthesis of the game-theoretic perspective. Theor Popul Biol 73:403–425
Kumpula J, Lefrere SC, Nieminen M (2004) The use of woodland lichen pasture by reindeer in winter with easy snow conditions. Arctic 57:273–278
Körner C (2021) Alpine plant life: functional plant ecology of high mountain ecosystems. Springer, Cham
Körner C, Hiltbrunner E (2021) Why is the alpine flora comparatively robust against climatic warming? Diversity 13:383
LaPerriere AJ, Lent PC (1977) Caribou feeding sites in relation to snow characteristics in northeastern Alaska. Arctic. https://doi.org/10.14430/arctic2690
Lawrence DM, Slater AG (2010) The contribution of snow condition trends to future ground climate. Clim Dyn 34:969–981
Leader-Williams N, Scott TA, Pratt R (1981) Forage selection by introduced reindeer on South Georgia, and its consequences for the flora. J Appl Ecol 18:83–106
Lechowicz MJ, Adams MS (1974) Ecology of Cladonia lichens. II. Comparative physiological ecology of C. mitis, C. rangiferina, and C. uncialis. Can J Bot 52:411–422
Lindsay J (2014) The whitebox geospatial analysis tools project and open-access GIS. In: Proceedings of the GIS research UK 22nd annual conference, The University of Glasgow, 2014. pp 16–18
Löffler J, Pape R (2020) Thermal niche predictors of alpine plant species. Ecology 101:e02891
MacArthur RH, Pianka ER (1966) On optimal use of a patchy environment. Am Nat 100:603–609
Manly B, McDonald L, Thomas DL, McDonald TL, Erickson WP (2002) Resource selection by animals: statistical design and analysis for field studies, 2nd edn. Kluwer Academic Publishers, Dordrecht
Mathiesen SD, Sormo W, Haga OE, Norbert HJ, Utsi THA, Tyler NJC (2000) The oral anatomy of Arctic ruminants: coping with seasonal changes. J Zool 251:119–128. https://doi.org/10.1111/j.1469-7998.2000.tb00597.x
McEwan E, Whitehead P (1970) Seasonal changes in the energy and nitrogen intake in reindeer and caribou. Can J Zool 48:905–913
Murtaugh PA (2007) Simplicity and complexity in ecological data analysis. Ecology 88:56–62
Mårell A (2006) Summer feeding behaviour of reindeer—a hierarchical approach, Acta Universitatis Agriculturae Sueciae vol 56, Agricultural University of Sweden (SLU), Umeå, pp 1652–6880
Mårell A, Edenius L (2006) Spatial heterogeneity and hierarchical feeding habitat selection by reindeer Arctic. Antarct Alp Res 38:413–420
Nellemann C (1996) Terrain selection by reindeer in late winter in central Norway. Arctic. https://doi.org/10.14430/arctic1209
Newman D, Lindsay J, Cockburn J (2018) Evaluating metrics of local topographic position for multiscale geomorphometric analysis. Geomorphology 312:40–50
Newmaster SG et al (2013) Examination of two new technologies to assess the diet of woodland caribou: video recorders attached to collars and DNA barcoding. Can J for Res 43:897–900
Nieminen M, Heiskari U (1989) Diets of freely grazing and captive reindeer during summer and winter. Rangifer 9:17–34
Odland A, Munkejord HK (2008) Plants as indicators of snow layer duration in southern Norwegian mountains. Ecol Indic 8:57–68. https://doi.org/10.1016/j.ecolind.2006.12.005
Odland A, Sandvik SM, Bjerketvedt DK, Myrvold LM (2014) Estimation of lichen biomass with emphasis on reindeer winter pastures at Hardangervidda, S Norway. Rangifer 34:95–110
Odland A, Sundstøl SA, Bjerketvedt DK (2018) Alpine lichen-dominated heaths: ecology, effects of reindeer grazing, and climate change. A review. Oecol Mont 27:30–50
Ophof A, Oldeboer K, Kumpula J (2013) Intake and chemical composition of winter and spring forage plants consumed by semi-domesticated reindeer (Rangifer tarandus tarandus) in Northern Finland. Anim Feed Sci Technol 185:190–195
Pape R, Löffler J (2015) Ecological dynamics in habitat selection of reindeer: an interplay of spatial scale, time, and individual animal’s choice. Polar Biol 38:1891–1903
Patterson BR, Power VA (2002) Contributions of forage competition, harvest, and climate fluctuation to changes in population growth of northern white-tailed deer. Oecologia 130:62–71
Perry G, Pianka ER (1997) Animal foraging: past, present and future. Trends Ecol Evol 12:360–364
Pruitt WO (1959) Snow as a factor in the winter ecology of the barren ground caribou (Rangifer arcticus). Arctic 12:158–179
Pyke GH (1984) Optimal foraging theory: a critical review. Annu Rev Ecol Syst 15:523–575
Quinby BM, Belk MC, Creighton JC (2020) Behavioral constraints on local adaptation and counter-gradient variation: implications for climate change. Ecol Evol 10:6688–6701
R Core Team (2022) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. May 15 2022. https://www.R-project.org
Rognmo A, Markussen KA, Jacobsen E, Grav HJ, Blix AS (1983) Effects of improved nutrition in pregnant reindeer on milk quality, calf birth weight, growth, and mortality. Rangifer 3:10–18
Rolandsen CM et al (2022) Klassifisering av de ti nasjonale villreinområdene etter kvalitetsnorm for villrein. Første klassifisering—2022, vol 2126. Norsk Institutt for Naturforskning, Trondheim
Romtveit L et al (2021) Optimal foraging by a large ungulate in an extreme environment: wild mountain reindeer select snow-free feeding habitats in winter. Ecol Evol 11:10409–10420
Saloranta T (2012) Simulating snow maps for norway: description and statistical evaluation of the seNorge snow model. Cryosphere 6:1323–1337
Schielzeth H (2010) Simple means to improve the interpretability of regression coefficients. Methods Ecol Evol 1:103–113
Signer J, Fieberg J, Avgar T (2019) Animal movement tools (amt): R package for managing tracking data and conducting habitat selection analyses. Ecol Evol 9:880–890
Skogland T (1978) Characteristics of snow cover and its relationship to wild mountain reindeer (Rangifer tarandus-tarandus) feeding stratergies. Arct Alp Res 10:569–579. https://doi.org/10.2307/1550680
Skogland T (1983) The effects of density dependent resource limitation on size of wild reindeer. Oecologia 60:156–168. https://doi.org/10.1007/BF00379517
Skogland T (1984) Wild reindeer foraging-niche organization. Holarct Ecol 7:345–379
Skogland T (1985) The effects of density-dependent resource limitations on the demography of wild reindeer. J Anim Ecol 54:359–374
Snell-Rood EC (2013) An overview of the evolutionary causes and consequences of behavioural plasticity. Anim Behav 85:1004–1011
Storeheier PV, Mathiesen SD, Tyler NJC, Olsen MA (2002) Nutritive value of Terricolous lichens for reindeer in winter. Lichenologist 34:247–257. https://doi.org/10.1006/lich.2002.0394
Storeheier PV, Mathiesen SD, Tyler NJC, Schjelderup I, Olsen MA (2003) Utilization of nitrogen- and mineral-rich vascular forage plants by reindeer in winter (vol 139, pg 151, 2002). J Agric Sci 140:128–128. https://doi.org/10.1017/s002185960300995x
Strand O (2009) Villreinens arealbruk i et landskapsøkologisk perspektiv. In: Sørensen R, Ydse H, Rusten E, Skorem J (eds) Villrein og ferdsel. Seminat Hjerkinn 21–22 oktober 2009., vol NVS Rapport 2/2009 Norsk Villreinsenter Nord, Hjerkinn, Norsk Villreinsenter Nord, Hjerkinn, pp 15–17
Strand O, Bevanger K, Falldorf T (2006) Reinens bruk av Hardangervidda. Sluttrapport fra Rv7-prosjektet. Norsk Institutt for Naturforskning, Trondheim
Strand O, Panzacchi M, Jordhøy P, Moorter BV, Andersen R, Bay LA (2011) Villreinens bruk av Setesdalsheiene. Sluttrapport fra GPS-merkeprosjektet 2006–2010, vol 694. Norwegian Institute for Nature Research (NINA), Trondheim
Strand O, Jordhøy P, Panzacchi M, Van Moorter B (2015) Veger og villrein. Oppsummering–overvåking av Rv7 over Hardangervidda, vol 1121. Norwegian Insitute for Nature Reserach (NINA), Trondheim
Sturm M, Wagner AM (2010) Using repeated patterns in snow distribution modeling: an Arctic example. Water Resour Res. https://doi.org/10.1029/2010WR009434
Staaland H, Hove K, White R (1986) Mineral absorption in relation to nutritional ecology of reindeer. Rangifer 6:279–287
Sutherland WJ (1983) Aggregation and theideal free’distribution. J Anim Ecol 52:821–828
Sutherland WJ (1996) From individual behaviour to population ecology, vol 11. Oxford University Press, Oxford
Swanson J, Barker M (1992) Assessment of Alaska reindeer populations and range conditions. Rangifer 12:33–43
Sæther B-E, Andersen R (1990) Resource limitation in a generalist herbivore, the moose Alces alces: ecological constraints on behavioural decisions. Can J Zool 68:993–999
Therneau TM, Grambsch PM (2000) The cox model. In: Modeling survival data: extending the cox model. Springer, New York, pp 39–77
Thompson ID, Wiebe PA, Mallon E, Rodgers AR, Fryxell JM, Baker JA, Reid D (2015) Factors influencing the seasonal diet selection by woodland caribou (Rangifer tarandus tarandus) in boreal forests in Ontario. Can J Zool 93:87–98
Thomson BR (1978) Behaviour of wild reindeer in Norway. University of Edinburgh, Edinburgh
Thurfjell H, Ciuti S, Boyce MS (2014) Applications of step-selection functions in ecology and conservation. Mov Ecol 2:4. https://doi.org/10.1186/2051-3933-2-4
Tucker B, Mahoney S, Greene B, Menchenton E, Russell L (1991) The influence of snow depth and hardness on winter habitat selection by caribou on the southwest coast of Newfoundland. Rangifer 11:160–163
Vistnes II, Nellemann C (2008) Reindeer winter grazing in alpine tundra: impacts on ridge community composition in Norway. Arct Antarct Alp Res 40:215–224. https://doi.org/10.1657/1523-0430(07-001)[vistnes]2.0.co;2
Warenberg K (1982) Reindeer forage plants in the early grazing season. Growth and nutritional content in relation to climatic conditions. Acta Phytogeogr Suec 70:71
West-Eberhard MJ (2003) Developmental plasticity and evolution. Oxford University Press, Oxford
White RG, Trudell J (1980) Habitat preference and forage consumption by reindeer and caribou near Atkasook, Alaska. Arct Alp Res 12:511–529
Wielgolaski FE, Goodall DW (1997) Polar and alpine tundra. Elsevier, Amsterdam
Williams MW, Brooks PD, Seastedt T (1998) Nitrogen and carbon soil dynamics in response to climate change in a high-elevation ecosystem in the Rocky Mountains, USA. Arct Alp Res 30:26–30
Acknowledgements
We are grateful to Roy Andersen for his services in tagging reindeer, and to Lena Romtveit and Anders Mossing for help with photos, maps and discussions. The Norwegian Wild Reindeer Centre, Skinnarbu, provided financial support for part of this work.
Funding
Open access funding provided by University Of South-Eastern Norway.
Author information
Authors and Affiliations
Contributions
OS and JH conceived and designed research. MH, OS, LK, DKB, AO and JH conducted experiments/field work. MH, LK, DKB, AO, RP and JH contributed new reagents or analytical tools. MH, LK and JH analyzed data. MH and JH wrote the manuscript with input from all authors. All authors read and approved the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
The use of reindeer in this study was approved by the Norwegian Food Safety Authority FOTS: ID 15116, license # 19/20935 and complied with institutional and national guidelines.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Holtan, M., Strand, O., Kastdalen, L. et al. Wild Mountain reindeer Rangifer tarandus tarandus winter foraging: snow-free areas a key resource for feeding. Polar Biol 46, 1321–1334 (2023). https://doi.org/10.1007/s00300-023-03204-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-023-03204-x