
Abstract
During summer, native anadromous Arctic char (Salvelinus alpinus) and the alien species pink salmon (Oncorhynchus gorbuscha) now coexist in marine environments in Svalbard, following the introduction of the latter in the Barents Region. To investigate potential dietary competition between these two salmonid species, stomach contents from Arctic char (n = 301) and pink salmon (n = 28) were sampled from different areas within the archipelago. The most important prey in terms of biomass for both salmonid species were amphipods; Themisto libellula (B = 26.0%) for Arctic char and Onisimus litoralis (B = 35.0%) for pink salmon. Pianka’s niche overlap revealed that dietary overlap between the two species was moderately high (Oobs = 0.59); both species had strong associations with intertidal invertebrates in areas where direct comparisons were possible (Kongsfjorden/Krossfjorden). However, both salmonid species did also eat some fish, with Arctic char consuming more offshore pelagic fish, while the small number of fish eaten by pink salmon were primarily coastal demersal fish species. Arctic char was a more generalist feeder, while pink salmon was more of a dietary specialist. Furthermore, the diet composition of the Arctic char consisted of 32.9% Atlantic prey while the pink salmon, surprisingly, ate only Arctic species, likely due to their tightly coastal feeding habits. Even though the sample size for pink salmon was low, this study contributes new insights into salmonid diets in Svalbard and the potential for introduced species to compete with native Arctic endemics, particularly in the expected warmer Arctic of the future.
Similar content being viewed by others


Introduction
The High Arctic Svalbard Archipelago is situated in the northern part of the Barents Sea (Fig. 1). Relative to other areas at the same latitude, the climate in Svalbard is mild, in large part because of heat transported by the West Spitsbergen Current (WSC), an extension of the North Atlantic Current (Beszczynska-Möller et al. 2012; Smedsrud et al. 2013). The freshwater systems in Svalbard cover no more than 400 km2 (0.6%) of its land area and are characterized by low precipitation, low primary production, low biodiversity, long seasonal ice cover, and cold water (von Quillfeldt and Øseth 2016). Arctic char (Salvelinus alpinus) is the only limnic vertebrate known to reproduce successfully in Svalbard (Gulseth and Nilssen 2000; Johnsen et al. 2021). This salmonid fish has a circumpolar distribution in the Arctic (Johnson 1980). It is the northernmost freshwater fish species, with a natural range stretching from Ireland and the British Isles to Svalbard and Severnaya Zemlya on the Eurasian Continent, and from Maine to Ellesmere Island on the North American Continent, (Johnson 1980; Ferguson et al. 2019; Svenning et al. 2020; Chernova et al. 2021). Arctic char usually occupies cold lakes, often as the sole fish species (Klemetsen et al. 2003; Ferguson et al. 2019), but there are also purely riverine populations (Jensen and Rikardsen 2008). This species exhibits remarkable life cycle variability in different populations; many populations are stationary in one waterbody and exhibit polymorphic body sizes with a large and a small (dwarf) morphotype (Hindar and Jonsson 1993; Tallman et al. 1996), while in other populations, some individuals perform seasonal migrations to the sea (i.e., anadromy).

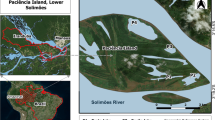
Map of the Svalbard Archipelago, identifying locations (and areas) where Arctic char (Salvelinus alpinus) and pink salmon (Oncorhynchus gorbuscha) were collected for dietary studies during 2015–2018
Anadromy in Arctic char is complex and takes place only in populations in the northern part of the species range. Seaward migrations are undertaken by both juvenile and mature fish (Rikardsen and Elliott 2000; Klemetsen et al. 2003). Stationary and anadromous Arctic char exist sympatrically and belong to the same gene pool; individuals may even shift from one form to the other during their lifetime (Nordeng 1983; Hindar et al. 1986). In Svalbard, Arctic char occupy some 100–150 lakes, but only about 20 of these are home to anadromous Arctic char (Svenning 2010). Stationary fish are mainly small individuals that feed on insects and zooplankton. These fish do not get longer than 15 cm. However, some stationary fish shift to cannibalism and can grow to be 70 cm long (Gullestad 1975; Gulseth and Nilssen 2001; Johnsen et al. 2021). In lakes connected to the sea, some individuals become anadromous and can grow to large size (Svenning and Borgstrøm 1995; Gulseth and Nilssen 2001; Svenning et al. 2007). In Svalbard, Arctic char normally migrate to the sea as soon as the rivers thaw in late spring/early summer and return to the lakes in the late summer or autumn (Gulseth and Nilssen 2000; Svenning et al. 2020). In recent years, there have been early (April–May) migrations to the sea, followed by early returns, although small Arctic char have been caught at sea as late as October (Christensen, unpubl. data). Svalbard Arctic char undertake their first migration when they are 4–8 years old, i.e., 3–4 years before they spawn for the first time (Dahl 1926; Gullestad 1975; Gulseth and Nilssen 2001). During the relatively short period spent at sea, anadromous Arctic char grow rapidly (Gulseth and Nilssen 2001; Klemetsen et al. 2003). However, migration comes with extreme risk, return rates range between ~ 33% for the smallest fish (< 20 cm) and ~ 70% for the largest fish (> 35 cm; Gulseth and Nilssen 2000). Mortality at sea is not well studied for Svalbard Arctic char but it is attributed to predation by seals or piscivorous fish, e.g., Atlantic cod (Gadus morhua) or not being able to survive the winter at sea after failing to migrate upstream before rivers freeze/dry up (Gulseth and Nilssen 2000; Svenning and Gullestad 2002). In addition, local citizens in Svalbard actively fish for anadromous Arctic char with gillnets and rods, which has reduced some local Arctic char populations (Svenning et al. 2020; Johnsen et al. 2021).
The marine diet of Arctic char in Svalbard has been the subject of several studies. Pelagic, intertidal, and demersal fish as well as crustaceans were reported to be prey for char by Dahl (1926). More recent studies report similar results, with some variation; the char diet includes capelin (Mallotus villosus), juvenile Cottidae and Liparidae, Mysis oculata, Gammarus spp. and euphausiids—mainly Thysanoessa spp. (Skogstad and Skogstad 2006; Ebne 2009; Bergane 2018). Euphausiids appear to be especially important for small Arctic char (< 40 cm), while fish becomes more prevalent in larger individuals (Ebne 2009; Bergane 2018).
Pink salmon (Oncorhynchus gorbuscha) is a native Pacific salmonid species that has a wide distribution around the North Pacific Ocean (Heard 1991; Hard et al. 1996). From 1956 until the end of the century, fertilized eggs and pink salmon fry were released in rivers in northwest Russia, which has led to pink salmon occurring in both Russian and mainland Norwegian rivers (Gordeeva et al. 2015; Niemelä et al. 2016; Sandlund et al. 2019), where it spawns successfully in some rivers in both countries (Hesthagen and Sandlund 2015; Mo et al. 2018). Unlike the iteroparous Arctic char, pink salmon are semelparous and spend most of their life at sea, having a life cycle of only two-years. Fry hatch during spring from eggs laid the previous year and migrate downstream to the sea almost as soon as they have emerged from the riverbed (Quinn 2018). Pink salmon remain at sea for one winter and return to spawn in rivers during summer/fall and die shortly thereafter (Heard 1991; Sandlund et al. 2019). Pink salmon was first caught in Svalbard in 1961 and now occurs regularly in the archipelago, both in freshwater and in the sea (Gullestad 1968; Witkowski and Głowacki 2010; Gjelland and Sandlund 2012).
Currently, there is no threat of pink salmon reproducing successfully in Svalbard rivers since they freeze or dry up during fall and winter. However, this might change in the future with continued warming of the climate and increasing levels of precipitation. Their presence in freshwater systems suggests that they are already attempting to spawn, and pink salmon have been documented to aggressively protect their spawning redds (Mo et al. 2018; Sandlund et al. 2019). This could potentially affect Arctic char. A more pressing issue is potential dietary competition between the species when foraging at sea. Pink salmon in its natural range feed on many of the same prey types as Arctic char, e.g., euphausiids, amphipods, and small fish, but also on pteropods, squid, and copepods (Hard et al. 1996; Kaeriyama et al. 2000, 2004), which is also true for pink salmon off the coast of mainland Norway (Diaz Pauli et al. 2023). During years with large spawning stocks of pink salmon in the Pacific, zooplankton biomass can be significantly reduced, which affects zooplankton consumption by competing salmonids (Shiomoto et al. 1997; Ruggerone and Nielsen 2004). High abundance of pink salmon has been shown to have negative effects on recruitment, growth, and survival in sockeye salmon (Oncorhynchus nerka) in the North Pacific (Ruggerone and Connors 2015).
During recent decades, both the volume and the temperature of Atlantic Water (AW) transported to the Barents Sea has increased markedly (Lind et al. 2018; Pörtner et al. 2019). This has resulted in large reductions in sea ice extent and volume (Tverberg et al. 2014; Lind et al. 2018) and “Atlantification” of the marine ecosystem (Renaud et al. 2012; Fossheim et al. 2015; Weydmann-Zwolicka et al. 2021). These system changes are likely to impact the diet of anadromous char and potentially also alter predator pressure with increases in the presence of piscivorous Atlantic gadids, e.g., Atlantic cod, haddock (Melanogrammus aeglefinus), and saithe (Pollachius virens; Renaud et al. 2012). The local harbor seal (Phoca vitulina) population is also increasing in number and expanding their distribution within the archipelago (Bengtsson et al. 2021). Furthermore, Atlantification of the ecosystem might increase competition with the alien species pink salmon, especially if it successfully spawns in the archipelago.
The purpose of this study was to increase knowledge regarding the marine diet of Arctic char and pink salmon in Svalbard, and to investigate potential dietary overlap between these species. In addition, community structure of the salmonids in terms of size, sex and maturity were analyzed. Such knowledge will be needed by environmental authorities in the years to come to manage salmonids in a new climate regime.
Materials and methods
Study area
Arctic char and pink salmon were collected between mid-July and mid-August from 2015 to 2018 at six different coastal areas in Svalbard: (1) inner Isfjorden; (2) outer Isfjorden; (3) Forlandsundet; (4), Kongsfjorden/Krossfjorden; (5) North Spitsbergen; and 6) Wahlenbergfjorden (Fig. 1). Isfjorden, Kongsfjorden/Krossfjorden and Forlandsundet are all greatly influenced by AW brought up the west coast of Spitsbergen by the WSC (Nilsen et al. 2016; Tverberg et al. 2019; Skogseth et al. 2020). North Spitsbergen is also influenced by AW from the WSC, but usually has a more pronounced surface layer of Arctic Water (ArW) due to its proximity to the Arctic Ocean (Cokelet et al. 2008; Geoffroy et al. 2017). Wahlenbergfjorden is mixed, with influences from both AW and ArW from Hinlopen Strait in its outer parts, while the inner parts of the fjord are characterized by locally produced meltwater from glaciers and sea ice (Bartels et al. 2018; Menze et al. 2020).
Field and laboratory methods
Fish were caught using 27 m long and 1.8 m high gillnets with mixed mesh size ranging from 35 to 63 mm (knot to knot). The nets were attached to the shore and stretched through the littoral and sublittoral zones. Immediately after capture, fish were killed and fork length to the closest mm and body mass to the closest g were measured. Sex and reproductive status (juvenile, mature spawning in the year of capture or mature not spawning in the year of capture) were determined based on the state of the gonads (Dahl 1917). Mature fish that would not spawn this year had undeveloped gonads, but well-developed blood vessels to the gonads. Mature females that would not spawn this year additionally had a mix of residual eggs and undeveloped eggs. Otoliths were removed and put in 70% ethanol mixed with glycerol and stomachs were taken out and stored at − 20 °C until analyzed.
In the lab, otoliths were put on a black Petri dish with a few drops of glycerol and age was determined by counting opaque zones under a stereo microscope (Barber and McFarlane 1987). Stomachs were thawed in lukewarm water and opened with a pair of scissors and their contents were washed with tap water over a sieve with a 0.5 mm2 mesh size. All remaining hard parts were placed in a Petri dish and all elements were identified to the lowest possible taxonomic level using identification guides by Klekowski and Węsławski (1992) and Vassilenko and Petryashov (2009). When possible, the size of prey organisms or identifying body parts (otolith, carapax) were measured. Biomass of consumed prey was reconstructed by using data on size-weight relationships of relevant organisms collected along the west coast of Svalbard (1990–2000) by the Institute of Oceanology of the Polish Academy of Sciences (IO PAN).
Statistical analysis
All statistical analyses were performed with R version 4.1.2 in RStudio (R Core Team 2021), and significance level was set at α < 0.05. Potential differences in length and age between juveniles, sexually mature females and sexually mature males of Arctic char were tested using Kruskal–Wallis rank-sum tests (Hollander et al. 2013), followed by Dunn’s test of multiple comparisons (Dunn 1964), using Holm’s method for adjusted p values (Holm 1979). A Student’s T test was used to test difference in length between males and females of pink salmon. Assumptions of homogeneity of variance and normal distribution were tested using Levene’s test (Levene 1960) and Shapiro–Wilk’s normality test (Shapiro and Wilk 1965). Length was used instead of body mass to compare sizes between the groups to avoid the influence of body condition, which might be more affected by the timing of descent/ascent to/from sea.
The contribution of different prey types to the diet of Arctic char and pink salmon was explored at the population level by calculating the frequency of occurrence (FOi; the percentage of all individuals that had consumed prey type i), relative frequency (Ni; the percentage of prey type i to the total number of prey) and relative percentage of (reconstructed) biomass (Bi; the percentage biomass of prey type i to the total prey biomass; Hyslop 1980; Chipps and Garvey 2007).
Dietary patterns of Arctic char and pink salmon were explored by grouping prey in ecologically relevant classes; pelagic invertebrates, demersal invertebrates, intertidal invertebrates, pelagic fish, demersal fish and “other”. The “other” class consisted of prey which could not be identified to taxonomic groups lower than to order, e.g., fragments of fish and invertebrates, and hence were excluded from further analysis along with individual Arctic char and pink salmon which only had prey items of this class in their stomachs. Relative importance of prey and feeding strategy (generalized vs specialized) were evaluated visually by plotting FO against B of the different prey classes in a modified “Costello plot” (Costello 1990; Amundsen et al. 1996).
Prey associated with AW (Thysanoessa inermis, Themisto abyssorum, capelin and Atlantic herring (Clupea harrengus)) were grouped together as “Atlantic prey” and the rest were classified as “Arctic prey” when comparing the contribution of prey associated with AW vs prey associated with ArW to diet composition.
Dietary overlap of co-occuring Arctic char and pink salmon populations, based on reconstructed biomasses of the five different prey classes described above, was estimated by calculating Pianka’s niche overlap index (Oobs) and comparing it with a null expectation (Oexp) using the ra3 algorithm (1000 replications) in the EcoSimR package in R (Pianka 1974; Gotelli et al. 2015). Levels of overlap were set to low (< 0.4), intermediate (0.4–0.6) and high (> 0.6; Grossman 1986).
To visualize differences in diet composition between sampling areas and the two salmonid fishes, non-metric multidimensional scaling (NMDS) was applied with the R package vegan (Oksanen et al. 2013), using Euclidean distance and center log ratio transformed biomass proportions (0:1) of the five prey classes at the individual predator scale, accounting for zeroes by replacing them with pseudo-counts of 0.001 (Sisk-Hackworth and Kelley 2020). Ellipses showing 95% confidence interval of the mean position for each group (area/species) were drawn to visualize potential overlap. To test if diet composition differed between species/areas, permutational multivariate analysis of variance (PERMANOVA; Anderson 2001) was used with 1000 permutations to test if group centroids differed significantly from each other, followed by pairwise comparisons with Holm’s adjusted p values. Assumption of multivariate homogeneity of groups dispersions was tested prior to performing PERMANOVA (Anderson 2006). Due to patchiness in the data—e.g., some areas were not sampled during certain years and some sex/maturity classes were missing from some years/areas—all individuals, regardless of year of sampling and sex/maturity class were pooled for each area when comparing diet composition of Arctic char between areas.
Results
In total, 301 Arctic char and 28 pink salmon were collected in Svalbard between 2015 and 2018. 272 char and 17 pink salmon had identifiable prey items in their stomachs (Table 1). Most Arctic char were caught during 2017 (n = 103) and 2018 (n = 116), in North Spitsbergen (n = 94) and Forlandsundet (n = 73, Table 1). Most pink salmon were caught in 2017 in Kongsfjorden/Krossfjorden (n = 21). Among the Arctic char, 41 (13.6%) were sexually mature males, 80 (26.6%) juvenile males, 71 (23.6%) sexually mature females, 108 (35.9%) juvenile females and 1 (0.3%) juvenile of undetermined sex. Of the sexually mature Arctic char, only 7 of the males and 1 of the females had gonadal development suggesting that they would breed in the season of capture. The mean length and body mass of the Arctic char were 45.3 cm (range 22.5–72.0 cm) and 1160 g (range 110–4900 g, and the mean age was 11 years (range 5–20 years, Table 2). Eight of the Arctic char could not be successfully aged. Juvenile, mature female and mature male Arctic char overlapped in terms of age/length (Fig. 2), however there were significant differences between the groups (Kruskal–Wallis test: age, H2 = 16.73, p = 0.0002; length, H2 = 21.80, p < 0.0001). Pairwise testing revealed that juveniles were on average significantly younger (10.3 ± 2.91 years, n = 182) and shorter (43.6 ± 9.5 cm, n = 189) than both mature females (age, 11.6 ± 3.09 years, n = 71, padj = 0.01; length, 47.6 ± 6.3 cm, n = 71, padj = 0.002) and mature males (age, 12.0 ± 2.73 years, n = 40, padj = 0.001; length, 49.7 ± 7.09 cm, n = 41, padj = 0.0003). Arctic char from North Spitsbergen were longer than fish at all other locations, and Arctic char from Outer Isfjorden were on average shorter (Online Resource 1). The proportion of invertebrates (both demersal and intertidal) decreased and the proportion of fish increased in the diet of Arctic char with increasing length of the predator (Online Resource 2). No sex differences were apparent (Online Resource 3).

Length and age of Arctic char (Salvelinus alpinus) grouped as juvenile, mature female and mature male based on gonadal development. Boxes contain 25th, 50th (median line) and 75th percentiles, and whiskers stretch between the minimum and the maximum values (excluding outliers). Outliers (> 1.5 × higher or lower than upper or lower quartile) are represented by dots
All pink salmon were one year old with gonads developed for spawning (Table 2)—nine were males and 19 were females. Average length and body mass of pink salmon were 46.1 cm (range 42.0–52.0 cm) and 1395 g (range 920–2230 g; Table 2), respectively. There was no significant difference in length between male and female pink salmon.
Dietary analyses showed that Arctic char had consumed 34 different, identifiable prey species/genera (hereafter only referred to as prey species) in addition to 11 different prey types which could not be identified lower than to order (Table 3). The most numerous prey species of Arctic char was the pelagic invertebrate Themisto libellula (n = 7424, Ni = 37.2%), which was also the species with the highest Bi (26.0%) and second highest FOi (35.7%; Table 3). Juvenile Cottidae (demersal fish) was the most commonly consumed fish species (n = 1019, Ni = 5.1%) and the prey with the second highest Bi (17.8%) and third highest FOi (30.0%; Table 3). Other important prey species included the pelagic crustacean T. inermis (FOi = 40.7%; Ni = 16.5%; Bi = 14.5%), the intertidal amphipod Gammarus setosus (FOi = 17.1%; Ni = 5.9%; Bi = 10.3%) and the pelagic (fish) capelin (FOi = 13.2%; Ni = 0.4%; Bi = 13.7%; Table 3). The most important demersal invertebrate was M. oculata (FOi = 20.4%; Ni = 8.4%; Bi = 4.4%; Table 3). Pelagic invertebrates were the prey class that made up the largest proportion of the Arctic char diet in terms of biomass (41.5%), followed by pelagic fish and demersal fish which made up 18.9% and 18%, respectively (Fig. 3). The Costello plot for Arctic char revealed that it was a generalist feeder—most prey classes had a FO between 16.8 and 37.5%, and B between 5.1 and 18.9%—with pelagic invertebrates as most important prey class (FO = 61.4%, B = 41.5%; Fig. 4). Eight of the Arctic char had only consumed prey of the “other” class (i.e., prey which could not be identified lower than to order).

Diet composition of Arctic char (Salvelinus alpinus) and pink salmon (Oncorhynchus gorbuscha) in Svalbard (2015–2018) presented as reconstructed biomass (proportions) of five different prey classes

Frequency of occurrence and relative percentage of reconstructed biomass of five different prey classes in the diet of Arctic char (Salvelinus alpinus; n = 272) and pink salmon (Oncorhynchus gorbuscha; n = 17), caught in Svalbard (2015–2018). Illustrations by Milena Sontowska
Pink salmon had consumed 12 different, identifiable prey species in addition to 5 prey types which could not be identified lower than to order (Table 3). The most numerous prey species of pink salmon were juvenile Gammarus spp. (n = 139; Ni = 32.6%) and Onisimus litoralis (n = 138, Ni = 32.4%), which are both intertidal invertebrates (Table 3). O. litoralis was also the species with the highest Bi (35.0%), followed by G. setosus (26.0%), while juvenile Gammarus spp. had the highest FOi (52.9%). Juvenile Cottidae had the second highest FOi (41.1%) and the (shared) third highest Bi (12.7%) together with T. inermis (Table 3). Intertidal invertebrates dominated other prey classes in terms of the proportion of biomass (65.1%; Fig. 3) and FO (76.5%). The Costello plot further revealed that pink salmon specialized on this same prey class (Fig. 4).
Dietary niche overlap between Arctic char and pink salmon was investigated only for fish from Kongsfjorden/Krossfjorden (2017), where there was both temporal and spatial overlap between the two species (Arctic char, n = 15, pink salmon, n = 10). Diet composition of Arctic char in this subset, was dominated by pelagic fish (B = 62.6%) and intertidal invertebrates (B = 34.9%) in terms of biomass. The diet of pink salmon was dominated by intertidal invertebrates (B = 77.5%) and demersal fish (B = 21.9%; Fig. 5). Dietary niche overlap was at the high end of the intermediate interval (Oobs = 0.59, Oexp = 0.45, p = 0.25), though the diet composition of the two salmonids was significantly different (PERMANOVA: F1 = 3.81, p = 0.025, stress = 0.04). Both salmonids in this subset consumed a lot of intertidal invertebrates. However, Arctic char ate more pelagic fish, while the fish prey targeted by pink salmon were mostly demersal fish (Fig. 6). Further, the diet composition of the Arctic char in this subset consisted of 32.9% Atlantic prey while pink salmon consumed only Arctic prey (Online Resource 4).

Diet composition of Arctic char (Salvelinus alpinus) and pink salmon (Oncorhynchus gorbuscha) in Kongsfjorden and Krossfjorden, Svalbard during 2017, presented as reconstructed biomass proportions of five different prey classes

Non-metric multidimensional scaling (NMDS) of diet composition based on reconstructed biomass proportions of Arctic char (Salvelinus alpinus) and pink salmon (Oncorhynchus gorbuscha) from Kongsfjorden/Krossfjorden, Svalbard 2017. Large points with black borders represent group centroids (Arctic char/pink salmon) and ellipses show 95% confidence area for mean position of each group. Rhombi with black borders show (prey) species; demersal invertebrates (DI), pelagic invertebrates (PI), intertidal invertebrates (II), demersal fish (DF) and pelagic fish (PF)
Regional dietary comparisons were only possible for Arctic char (n = 272, excluding individuals without identifiable prey in their stomachs), because the sample sizes by location for pink salmon were too low. Pelagic invertebrates dominated the diet of Arctic char in North Spitsbergen (B = 65.4%) and made up almost half of their diet in Outer Isfjorden (B = 49.3%, Fig. 7). Intertidal invertebrates dominated the diet composition in Inner Isfjorden (B = 66.1%) and Wahlenbergfjorden (B = 51.9%), and demersal fish was the dominant prey class in Forlandsundet (B = 64.8%, Fig. 7). The highest proportion of demersal invertebrates was in Wahlenbergfjorden (B = 28.2%) and the highest contribution of pelagic fish was in North Spitsbergen (B = 27.7%, Fig. 7). There was overlap in diet composition between all areas, except North Spitsbergen (Fig. 8), which was significantly different from the other areas (PERMANOVA: F5 = 18.96, p = 0.0001, stress = 0.17). Pairwise comparisons revealed that the diet of Arctic char was significantly different in each area (padj = 0.15). In North Spitsbergen Atlantic prey made up the highest proportion in the diet (B = 46.3%), followed by Outer Isfjorden (B = 29.8%), while Arctic prey species dominated at all other locations (Online Resource 5).

Diet composition of Arctic char (Salvelinus alpinus) from different areas in Svalbard (2015–2018), presented as reconstructed biomass (proportions) of five different prey classes. Isfjorden (In. Is), Outer Isfjorden (Out. Is), Forlandsundet (Forl.), Kongsfjorden/Krossfjorden (K/K), North Spitsbergen (No. Spi.), and Wahlenbergfjorden (Wahl.)

Non-metric multidimensional scaling (NMDS) of diet composition based on reconstructed biomass proportions of Arctic char (Salvelinus alpinus) and pink salmon (Oncorhynchus gorbuscha) from six areas in Svalbard (2015–2018). Large points represent group centroids and ellipses show 95% confidence areas for each area. Rhombi show (prey) species; demersal invertebrates (DI), pelagic invertebrates (PI), intertidal invertebrates (II), demersal fish (DF), and pelagic fish (PF)
Discussion
This study presents novel information about the diet of Arctic char and pink salmon in Svalbard as well as assessing dietary overlap between these two salmonids. It confirmed previous findings that anadromous Arctic char in Svalbard are generalist feeders that exhibit a slight preference for pelagic invertebrates, with amphipods, euphausiids, and small marine fish species being their most important prey (Skogstad and Skogstad 2006; Ebne 2009; Bergane 2018). It also revealed that pink salmon in Svalbard feed on many of the same species as Arctic char, but have a more specialized diet, comprised mainly of intertidal invertebrates, with the amphipods G. setosus and O. litoralis being especially important in terms of biomass. Pink salmon in its native range in the North Pacific, as well as in the Norwegian Sea have a varied diet, consisting of amphipods, euphausiids, small fish, copepods, pteropods, and squid (Heard 1991; Kaeriyama et al. 2004; Diaz Pauli et al. 2023). This species is generally considered to be a dietary generalist (Radchenko et al. 2018). The apparent contrast in feeding mode between North Pacific pink salmon and pink salmon in this study (generalist vs specialist), is likely explained by study “scale”. All pink salmon in this study were caught close to the coast, which might limit the number of prey classes recorded; in the Norwegian Sea, the diet for the whole population was varied, but areas tended to be dominated by one type of prey (Diaz Pauli et al. 2023). In Svalbard, G. setosus is found in inner fjord basins and O. litoralis is almost exclusively associated with soft bottom, tidal flats in front of rivers and glaciers (Węsławski 1994; Węsławski et al. 2000), strongly indicating that pink salmon in Svalbard feed in these areas. This would also explain the absence of prey associated with AW—which are more pelagic—in the pink salmon diet.
All of the pink salmon in this study were mature 1-year-olds, with well-developed gonads. It is well known that as pink salmon prepare to spawn, most of their energy is allocated towards gonad maturation and feeding rates decline (Heard 1991), so it was perhaps not surprising that a large proportion of the pink salmon caught in this study had empty stomachs (11 of 28). Both their empty stomachs and their gonad development state suggest that they were preparing to migrate upstream to spawn. This fact might bias their distribution toward the intertidal zone. Prey from this zone dominated the stomach contents (B = 65.1%) of the pink salmon that were still feeding.
Interestingly, T. abyssorum, was rare in the diet of Arctic char in this study and completely absent in the diet of pink salmon, while both species had consumed relatively high proportions of T. libellula. This is despite the high abundance of T. abyssorum in Svalbard fjords, especially in areas with large volumes of AW (Dalpadado et al. 2016; Hop et al. 2019). This suggests that both salmonids have a strong preference for T. libellula over T. abyssorum. This is also known to be the case for polar cod (Boreogadus saida) in Svalbard (Cusa et al. 2019). T. libellula is generally larger than T. abyssorum, which could explain this preference, though the two amphipods have similar energy/lipid content per biomass (Percy and Fife 1981; Auel et al. 2002; Węsławski et al. 2006). It could also suggest that the two salmonids feed in areas with relatively high abundances of T. libellula, e.g., inner fjord basins, as seen in Kongsfjorden (Dalpadado et al. 2016; Hop et al. 2019).
Dietary niche overlap was relatively high between Arctic char and pink salmon (Oobs = 0.59) in Svalbard when the two species co-occurred in the same fjord system, with both species consuming high proportions of intertidal invertebrates (pink salmon: B = 77.5%, Arctic char: B = 34.9%). What separated the diets of the two salmonids was the dominance of pelagic fish in the diet of Arctic char (B = 62.6%). Fish comprised a smaller portion of the pink salmon diet (B = 21.9%) and the fish they did eat were demersal species. This suggests that Arctic char and pink salmon feed in different areas and in different parts of the water column at least some of the time. Pink salmon feed mainly in intertidal areas on bottom-dwelling prey, while Arctic char feed both in pelagic areas further offshore and in the intertidal zone. Pink salmon in the North Pacific are known to compete with other salmonid species via intense consumption of shared resources, mainly zooplankton, during years with high abundances of pink salmon (Shiomoto et al. 1997; Ruggerone and Nielsen 2004). In 2017, there was a record number of pink salmon in rivers in mainland Norway (Sandlund et al. 2019), which corresponds well with the relatively high numbers of pink salmon caught in Svalbard during this year compared to the other years in this study. Interestingly, zooplankton were practically absent in the diet of both Arctic char and pink salmon (< 1% in pink salmon) in Kongsfjorden/Krossfjorden in 2017. The limited sample size, especially for pink salmon, and time scope of this study do not allow any firm conclusions to be drawn but suggest that there is strong potential for competition for resources between Arctic char and pink salmon in Svalbard if pink salmon become more abundant in the archipelago.
Intertidal invertebrates were an important part of the diet for Arctic char in Inner Isfjorden and Wahlenbergfjorden, and to a degree, also in Kongsfjorden/Krossfjorden. These three areas are within fjords, where locally produced water likely makes up a relatively large proportion of the water column, especially deep within the fjords and close to glacier fronts (Bartels et al. 2018; Tverberg et al. 2019; Skogseth et al. 2020). Pelagic invertebrates were more common in the diet of Arctic char sampled in Outer Isfjorden, Forlandsundet and North Spitsbergen, which are less sheltered and more exposed to inflowing water masses which transport zooplankton (Hop et al. 2006; Geoffroy et al. 2017; Skogseth et al. 2020). Outer Isfjorden and North Spitsbergen were also the areas where Atlantic prey contributed most to the Arctic char’s diet. Interestingly, demersal fish were eaten by Arctic char in the areas on the west coast, but not in the north and east, where pelagic fish were more prevalent in the diet compared to demersal fish. Body size might play a role given that the Arctic char from North Spitsbergen were on average larger than from the other areas, making it possible for them to consume pelagic fish, which were larger on average than the demersal fish eaten by either salmonid. However, interannual variation in the diet (prey community) cannot be ruled out, given that Arctic char from North Spitsbergen and Wahlenbergfjorden were only caught in 2017 and 2018, which were the years with the highest relative proportion of pelagic fish in the diet. Likewise, most Arctic char from Inner Isfjorden, where the diet was dominated by intertidal invertebrates, were caught in 2015. There appeared to be a size dependent trend in the diet of Arctic char as previously shown by Ebne (2009) and Bergane (2018). However, due to inconsistencies in the data in terms of year and area where different sized fish were caught—this trend was not investigated further.
Arctic char that were not sexually mature (juveniles) were significantly younger and smaller than adult males and females, but the differences in mean age and length were not great between juveniles of the two sexes (1.3 years and 4.3 cm compared to adult females, and 1.7 years and 6.1 cm compared to adult males). There was considerable variation in size at a given age. Considering that the age at first migration can vary between 4 and > 10 years for Arctic char in Svalbard (Gulseth and Nilssen 2001; Ebne 2009; Johnsen et al. 2021), this overlap was not unexpected. However, the relatively high average age and large size of juveniles in this study indicates that most were not first-time migrators. Mortality of anadromous Arctic char is high while at sea, especially for small individuals (< 25 cm), and only 10–50% of first-time migrators survive to return (Finstad and Heggberget 1993; Gulseth and Nilssen 2000). It is likely that smaller Arctic char are disproportionally eaten by seals and predatory fish. Atlantic cod in mainland Norway are known to eat salmonid smolts (Hvidsten and Møkkelgjerd 1987; Hvidsten and Lund 1988) and Arctic char with bitemarks, that probably come from seals, have been observed in Svalbard (Svenning et al. 2020). With continued increases in the volume and temperature of AW and increasing abundances of predators associated with AW, such as Atlantic cod, haddock, saithe and harbor seals, all of which are increasing in Svalbard fjords (Renaud et al. 2012; Bengtsson et al. 2021), the predatory pressure of Arctic char is also likely to increase. This could have negative effects on the local Arctic char population, especially for small, first-time migrators.
Stomach content analysis has inherent biases that are important to address when studying diet (Trites and Spitz 2018; Amundsen and Sánchez‐Hernández 2019). Differences in degradation and evacuation rates due to fat content, particle size and digestibility of different prey items can lead to an overrepresentation of large prey with easily identifiable hard parts, and an underrepresentation of small and/or soft bodied prey (Elliott 1972; Hyslop 1980; Legler et al. 2010; Baker et al. 2014). The diet of both Arctic char and pink salmon in this study consisted of relatively large crustaceans and fish, while smaller crustaceans (e.g., copepods and decapod larvae) and gelatinous organisms (e.g., pteropods) were absent or only made up a tiny proportion of the diet despite being abundant in Svalbard waters (Weydmann et al. 2014; Gluchowska et al. 2016). Neither copepods nor pteropods have been reported to be particularly important as food for Arctic char in Svalbard, but copepods are consumed by Arctic char in mainland Norway (Rikardsen et al. 2000), and both copepods and pteropods are important prey for pink salmon in the North Pacific (Heard 1991; Kaeriyama et al. 2004). Further, methods for quantitative estimation of diet, e.g., through reconstruction of biomass contribution of prey items introduces potential bias by making assumptions of prey condition and length to body mass relationships (Baker et al. 2014; Amundsen and Sánchez‐Hernández 2019). Due to the state of digestion of prey in the stomachs, this method was chosen over the more robust gravimetric method (Amundsen and Sánchez‐Hernández 2019) when analyzing differences in diet composition. It is important to note that all fish in this study were caught relatively close to shore, which might discriminate against any potential individuals feeding further out in the fjords. There might also have been an underrepresentation of small individuals, which would be especially relevant for (juvenile) Arctic char, due to the mixed mesh size of the gillnets (smallest mesh size 35 mm).
Further studies should focus on a limited number of areas sampled during the same time intervals and include methods that can determine diet over longer periods, e.g., stable isotope and fatty acid analyses to estimate the diet during periods when Arctic char and high numbers of pink salmon coexist in the marine environment compared to periods without high numbers of pink salmon. Parallel sampling of the local potential prey community should also be considered to investigate possible selectivity by the salmonids.
Conclusions
Novel findings of the pink salmon’s diet in Svalbard revealed that they share important prey species with the native Arctic char population. Differences in the proportions of different prey classes indicated that dietary overlap was moderately high, but also that the two salmonid species likely use different areas and different parts of the water column for at least some of their foraging. High proportions of empty stomachs in the pink salmon that were mature indicate that they were preparing to migrate to freshwater to spawn. Although the Arctic char in this study showed a preference for the Arctic pelagic amphipod T. libellula over its Atlantic counterpart T. abyssorum, the Arctic char did consume other species associated with AW, e.g., T. inermis, capelin, and Atlantic herring. With continued Atlantification of the marine environment in Svalbard, it is likely that the diet of the native Arctic char population will resemble that of the population in mainland Norway, which consists of herring, euphausiids, copepods, gadids, and other Atlantic fish species (Rikardsen et al. 2000, 2007; Rikardsen and Amundsen 2005). Continued warming of the Arctic is also likely to increase the level of competition between the salmonid species, to the detriment of the Arctic char. If pink salmon start to spawn in the Svalbard Archipelago it is likely that they will have negative impacts on the anadromous char populations from lakes that have access to the sea.
Data availability
The data are available via contact with the corresponding author.
References
Amundsen PA, Gabler HM, Staldvik F (1996) A new approach to graphical analysis of feeding strategy from stomach contents data—modification of the Costello (1990) method. J Fish Biol 48:607–614. https://doi.org/10.1111/j.1095-8649.1996.tb01455.x
Amundsen PA, Sánchez-Hernández J (2019) Feeding studies take guts–critical review and recommendations of methods for stomach contents analysis in fish. J Fish Biol 95:1364–1373. https://doi.org/10.1111/jfb.14151
Anderson MJ (2001) A new method for non-parametric multivariate analysis of variance. Austral Ecol 26:32–46. https://doi.org/10.1111/j.1442-9993.2001.01070.pp.x
Anderson MJ (2006) Distance-based tests for homogeneity of multivariate dispersions. Biometrics 62:245–253. https://doi.org/10.1111/j.1541-0420.2005.00440.x
Auel H, Harjes M, Da Rocha R, Stübing D, Hagen W (2002) Lipid biomarkers indicate different ecological niches and trophic relationships of the Arctic hyperiid amphipods Themisto abyssorum and T. libellula. Polar Biol 25:374–383. https://doi.org/10.1007/s00300-001-0354-7
Baker R, Buckland A, Sheaves M (2014) Fish gut content analysis: robust measures of diet composition. Fish Fish 15:170–177. https://doi.org/10.1111/faf.12026
Barber WE, McFarlane GA (1987) Evaluation of three techniques to age Arctic char from Alaskan and Canadian waters. Trans Am Fish Soc 116:874–881. https://doi.org/10.1577/1548-8659(1987)116%3c874:EOTTTA%3e2.0.CO;2
Bartels M, Titschack J, Fahl K, Stein R, Hebbeln D (2018) Wahlenbergfjord, eastern Svalbard: a glacier-surrounded fjord reflecting regional hydrographic variability during the Holocene? Boreas 47:1003–1021. https://doi.org/10.1111/bor.12325
Bengtsson O, Hamilton CD, Lydersen C, Andersen M, Kovacs KM (2021) Distribution and habitat characteristics of pinnipeds and polar bears in the Svalbard Archipelago, 2005–2018. Polar Res 40:5326. https://doi.org/10.33265/polar.v40.5326
Bergane VÅ (2018) Sjørøya (Salvelinus alpinus) i Linnévassdraget, Svalbard: livshistorietrekk og populasjonsstruktur ti år etter innføring av kvotebasert fangst. MSc thesis, Norwegian University of Life Sciences, Ås
Beszczynska-Möller A, Fahrbach E, Schauer U, Hansen E (2012) Variability in Atlantic water temperature and transport at the entrance to the Arctic Ocean, 1997–2010. ICES J Mar Sci 69:852–863. https://doi.org/10.1093/icesjms/fss056
Chernova N, Spiridonov V, Syomin V, Gavrilo M (2021) Notes on the fishes of the Severnaya Zemlya archipelago and the spawning area of polar cod Boreogadus saida (Gadidae). Proc Zool Inst RAS 325:248–268
Chipps SR, Garvey JE (2007) Assessment of food habits and feeding patterns. In: Guy C, Brown ML (eds) Analysis and interpretation of freshwater fisheries data. American Fisheries Society, Bethesda, pp 473–514
Cokelet ED, Tervalon N, Bellingham JG (2008) Hydrography of the West spitsbergen current, svalbard branch: autumn 2001. J Geophys Res Oceans 113:C01006. https://doi.org/10.1029/2007JC004150
Costello M (1990) Predator feeding strategy and prey importance: a new graphical analysis. J Fish Biol 36:261–263. https://doi.org/10.1111/j.1095-8649.1990.tb05601.x
Cusa M, Berge J, Varpe Ø (2019) Seasonal shifts in feeding patterns: individual and population realized specialization in a high Arctic fish. Ecol Evol 9:11112–11121. https://doi.org/10.1002/ece3.5615
Dahl K (1917) Studier og forsøk over ørret og ørretvand. Centraltrykkeriet, Kristiania (Oslo)
Dahl K (1926) Contributions to the biology of the Spitsbergen char. Skrifter Om Svalbard Og Ishavet 7:1–12
Dalpadado P, Hop H, Rønning J, Pavlov V et al (2016) Distribution and abundance of euphausiids and pelagic amphipods in Kongsfjorden, Isfjorden and Rijpfjorden (Svalbard) and changes in their relative importance as key prey in a warming marine ecosystem. Polar Biol 39:1765–1784. https://doi.org/10.1007/s00300-015-1874-x
Diaz Pauli B, Berntsen HH, Thorstad EB, Lusseau SM, Wennevik V, Utne KR (2023) Geographic distribution, abundance, diet, and body size of invasive pink salmon (Oncorhynchus gorbuscha) in the Norwegian and Barents Seas, and in Norwegian rivers. ICES J Mar Sci 80:76–90. https://doi.org/10.1093/icesjms/fsac224
Dunn OJ (1964) Multiple comparisons using rank sums. Technometrics 6:241–252
Ebne I (2009) Anadrom røye (Salvelinus alpinus) i Linnévassdraget, Svalbard; Diett, oppvandring, bestandssammensetning og fangbarhet. MSc thesis, Norwegian University of Life Sciences, Ås
Elliott J (1972) Rates of gastric evacuation in brown trout, Salmo trutta L. Freshw Biol 2:1–18. https://doi.org/10.1111/j.1365-2427.1972.tb01575.x
Ferguson A, Adams CE, Jóhannsson M, Kelly F, King RA, Maitland P, Winfield I (2019) Trout and char of the North Atlantic Isles. In: Kershner JL, Williams JE, Gresswell RE, Lobón-Cerviá J (eds) Trout and char of the world. American Fisheries Society, Bethesda
Finstad B, Heggberget T (1993) Migration, growth and survival of wild and hatchery-reared anadromous Arctic charr (Salvelinus alpinus) in Finnmark, northern Norway. J Fish Biol 43:303–312. https://doi.org/10.1111/j.1095-8649.1993.tb00430.x
Fossheim M, Primicerio R, Johannesen E, Ingvaldsen RB, Aschan MM, Dolgov AV (2015) Recent warming leads to a rapid borealization of fish communities in the Arctic. Nat Clim Change 5:673–677. https://doi.org/10.1038/nclimate2647
Geoffroy M, Cottier FR, Berge J, Inall ME (2017) AUV-based acoustic observations of the distribution and patchiness of pelagic scattering layers during midnight sun. ICES J Mar Sci 74:2342–2353. https://doi.org/10.1093/icesjms/fsw158
Gjelland KØ, Sandlund OT (2012) Pukkellaks (Oncorynchus gorbuscha) Artsdatabanken Faktaark 283, http://www2.artsdatabanken.no/faktaark/Faktaark283.pdf. Accessed 3 Nov 2022
Gluchowska M, Kwasniewski S, Prominska A, Olszewska A et al (2016) Zooplankton in Svalbard fjords on the Atlantic-Arctic boundary. Polar Biol 39:1785–1802. https://doi.org/10.1007/s00300-016-1991-1
Gordeeva N, Salmenkova E, Prusov S (2015) Variability of biological and population genetic indices in pink salmon, Oncorhynchus gorbuscha transplanted into the White Sea basin. J Ichthyol 55:69–76. https://doi.org/10.1134/S0032945215010051
Gotelli N, Hart E, Ellison A, Hart ME (2015) Package ‘EcoSimR’. https://github.com/GotelliLab/EcoSimR. Accessed 3 Nov 2022
Grossman GD (1986) Food resource partitioning in a rocky intertidal fish assemblage. J Zool 1:317–355. https://doi.org/10.1111/j.1096-3642.1986.tb00642.x
Gullestad N (1968) Observasjoner av pukkellaks (Oncorhynchus gorbuscha) på Svalbard i tiden 1960–1965. In: Gjelsvik T, Hagevold P, Hisdal V, Holmsen E, Norderhaug M, Winsnes TS (eds) Norsk Polarinstitutt årbok 1968. The Norwegian Polar Institute, Oslo, pp 131–134
Gullestad N (1975) On the biology of char (Salmo alpinus L.) in Svalbard. I. Migratory and non-migratory char in Revvatnet, Spitsbergen. In: Gjelsvik T, Hagevold P, Hisdal V, Larsen T (eds) Norsk Polarinstitutt årbok 1973. The Norwegian Polar Institute, Oslo, pp 125–140
Gulseth OA, Nilssen KJ (2000) The brief period of spring migration, short marine residence, and high return rate of a northern Svalbard population of Arctic char. Trans Am Fish Soc 129:782–796. https://doi.org/10.1577/1548-8659(2000)129%3c0782:TBPOSM%3e2.3.CO;2
Gulseth OA, Nilssen KJ (2001) Life–history traits of charr, Salvelinus alpinus, from a high arctic watercourse on Svalbard. Arctic 54:1–11
Hard JJ, Kope RG, Grant WS, Waknitz FW, Parker LT, Waples RS (1996) Status review of pink salmon from Washington, Oregon, and California. U.S Department of Commerce, NOAA Tech. Memo. NMFS-AFSC-25, Seattle, WA
Heard WR (1991) Life history of pink salmon (Oncorhynchus gorbuscha). In: Groot C, Margolis L (eds) Pacific salmon life histories. UBC Press, Vancouver, pp 121–230
Hesthagen T, Sandlund O (2015) Spredning av ferskvannsfisk i Norge, En fylkesvis oversikt og nye registreringer i 2015. Report No. 1205, Norwegian Institute for Nature Research, Trondheim
Hindar K, Ryman N, Ståhl G (1986) Genetic differentiation among local populations and morphotypes of Arctic charr, Salvelinus alpinus. Biol J Linn Soc 27:269–285. https://doi.org/10.1111/j.1095-8312.1986.tb01737.x
Hindar K, Jonsson B (1993) Ecological polymorphism in Arctic charr. Biol J Linn Soc 48:63–74. https://doi.org/10.1111/j.1095-8312.1993.tb00877.x
Hollander M, Wolfe DA, Chicken E (2013) Nonparametric statistical methods, 3rd edn. Wiley, Hoboken, NJ
Holm S (1979) A simple sequentially rejective multiple test procedure. Scand J Stat 6:65–70
Hop H, Falk-Petersen S, Svendsen H, Kwasniewski S, Pavlov V, Pavlova O, Søreide JE (2006) Physical and biological characteristics of the pelagic system across Fram Strait to Kongsfjorden. Prog Oceanogr 71:182–231. https://doi.org/10.1016/j.pocean.2006.09.007
Hop H, Wold A, Vihtakari M, Daase M et al (2019) Zooplankton in Kongsfjorden (1996–2016) in relation to climate change. In: Hop H, Wiencke C (eds) The ecosystem of Kongsfjorden. Springer, Svalbard, pp 229–300
Hvidsten N, Møkkelgjerd P (1987) Predation on salmon smolts, Salmo salar L., in the estuary of the River Surna. Norway J Fish Biol 30:273–280. https://doi.org/10.1111/j.1095-8649.1987.tb05752.x
Hvidsten N, Lund R (1988) Predation on hatchery-reared and wild smolts of Atlantic salmon, Salmo salar L., in the estuary of River Orkla. Norway J Fish Biol 33:121–126. https://doi.org/10.1111/j.1095-8649.1988.tb05453.x
Hyslop E (1980) Stomach contents analysis—a review of methods and their application. J Fish Biol 17:411–429. https://doi.org/10.1111/j.1095-8649.1980.tb02775.x
Jensen J, Rikardsen A (2008) Do northern riverine anadromous Arctic charr Salvelinus alpinus and sea trout Salmo trutta overwinter in estuarine and marine waters? J Fish Biol 73:1810–1818. https://doi.org/10.1111/j.1095-8649.2008.02042.x
Johnsen H, Iversen HR, Sigurdsen T (2021) Svalbardrøye (Salvelinus alpinus): sammenstilling av forvaltningsrelevant kunnskap. Report No. Short report 57, The Norwegian Polar Institute, https://brage.npolar.no/npolar-xmlui/handle/11250/2826637. Accessed 3 Nov 2022
Johnson L (1980) The arctic charr, Salvelinus alpinus. In: Balon EK (ed) Charrs, Salmonid fishes of the genus Salvelinus. Springer, The Hague, pp 15–98
Kaeriyama M, Nakamura M, Yamaguchi M, Ueda H et al (2000) Feeding ecology of sockeye and pink salmon in the Gulf of Alaska. N Pac Anadr Fish Comm Bull 2:55–63
Kaeriyama M, Nakamura M, Edpalina R, Bower J, Yamaguchi H, Walker R, Myers KW (2004) Change in feeding ecology and trophic dynamics of Pacific salmon (Oncorhynchus spp.) in the central Gulf of Alaska in relation to climate events. Fish Oceanogr 13:197–207. https://doi.org/10.1111/j.1365-2419.2004.00286.x
Klekowski RZ, Węsławski JM (1992) Atlas of the marine fauna of Southern Spitsbergen: invertebrates, vol 2. Ossolineum, Gdansk
Klemetsen A, Amundsen PA, Dempson J, Jonsson B, Jonsson N, O’connell M, Mortensen E (2003) Atlantic salmon Salmo salar L., brown trout Salmo trutta L. and Arctic charr Salvelinus alpinus (L.): a review of aspects of their life histories. Ecol Freshwat Fish 12:1–59. https://doi.org/10.1034/j.1600-0633.2003.00010.x
Legler ND, Johnson TB, Heath DD, Ludsin SA (2010) Water temperature and prey size effects on the rate of digestion of larval and early juvenile fish. Trans Am Fish Soc 139:868–875. https://doi.org/10.1577/T09-212.1
Levene H (1960) Robust tests for equality of variances. In: Olkin I (ed) Contributions to probability and statistics essays in honor of Harold Hotelling. Stanford University Press, Stanford, CA, pp 279–292
Lind S, Ingvaldsen RB, Furevik T (2018) Arctic warming hotspot in the northern Barents Sea linked to declining sea-ice import. Nat Clim Change 8:634–639. https://doi.org/10.1038/s41558-018-0205-y
Menze S, Ingvaldsen RB, Nikolopoulos A, Hattermann T, Albretsen J, Gjøsæter H (2020) Productive detours–Atlantic water inflow and acoustic backscatter in the major troughs along the Svalbard shelf. Prog Oceanogr 188:102447–102451. https://doi.org/10.1016/j.pocean.2020.102447
Mo TA, Thorstad EB, Sandlund OT, Berntsen HH, Fiske P, Uglem I (2018) The pink salmon invasion: a Norwegian perspective. J Fish Biol 93:5–7. https://doi.org/10.1111/jfb.13682
Niemelä E, Johansen N, Zubchenko AV, Dempson JB, Veselov A, Ieshko EP, Barskaya Y, Novokhatskaya OV, Shulman BS, Länsman M, Hassinen E, Kuusela J, Haantie J, Kylmäaho M, Kivilahti E, Arvola K-M, Kalske TH (2016) Pink salmon in the Barents region: with special attention to the status in the transboundary rivers Tana and Neiden, rivers in North West Russia and in East Canada. Report no 3-2016, Fylkesmannen i Finnmark. https://www.statsforvalteren.no/siteassets/utgatt/fm-finnmark/dokument-fmfi/miljovern/rapportserie/2016_3-pink-salmon-in-the-barents-region.pdf. Accessed 1 March 2023
Nilsen F, Skogseth R, Vaardal-Lunde J, Inall M (2016) A simple shelf circulation model: intrusion of Atlantic Water on the West Spitsbergen Shelf. J Phys Oceanogr 46:1209–1230. https://doi.org/10.1175/JPO-D-15-0058.1
Nordeng H (1983) Solution to the” char problem” based on Arctic char (Salvelinus alpinus) in Norway. Can J Fish Aquat Sci 40:1372–1387. https://doi.org/10.1139/f83-159
Oksanen J, Blanchet FG, Kindt R, Legendre P and others (2013) Package ‘vegan’: community ecology package, version 2.6-2. https://www.veganr-forger-projectorg/. Accessed 1 March 2023
Percy J, Fife F (1981) The biochemical composition and energy content of Arctic marine macrozooplankton. Arctic 34:307–313
Pianka ER (1974) Niche overlap and diffuse competition. Proc Natl Acad Sci USA 71:2141–2145. https://doi.org/10.1073/pnas.71.5.2141
Pörtner H-O, Roberts DC, Masson-Delmotte V, Zhai P, Tignor M, Poloczanska E, Weyer N (2019) IPCC special report on the ocean and cryosphere in a changing climate, Intergovernmental Panel on Climate Change. https://www.ipcc.ch/site/assets/uploads/sites/3/2019/12/SROCC_FullReport_FINAL.pdf. Accessed 3 Nov 2022
Quinn TP (2018) The behavior and ecology of Pacific salmon and trout, 2nd edn. University of Washington Press, Seattle, WA
R Core Team (2021) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Radchenko V, Beamish R, Heard W, Temnykh O (2018) Ocean ecology of pink salmon. In: Beamish R (ed) The ocean ecology of Pacific salmon and trout. American Fisheries Society, Bethesda, pp 15–160
Renaud PE, Berge J, Varpe Ø, Lønne OJ, Nahrgang J, Ottesen C, Hallanger I (2012) Is the poleward expansion by Atlantic cod and haddock threatening native polar cod, Boreogadus saida? Polar Biol 35:401–412. https://doi.org/10.1007/s00300-011-1085-z
Rikardsen A, Amundsen PA, Bjørn P, Johansen M (2000) Comparison of growth, diet and food consumption of sea-run and lake-dwelling Arctic charr. J Fish Biol 57:1172–1188. https://doi.org/10.1111/j.1095-8649.2000.tb00479.x
Rikardsen A, Elliott J (2000) Variations in juvenile growth, energy allocation and life-history strategies of two populations of Arctic charr in North Norway. J Fish Biol 56:328–346. https://doi.org/10.1111/j.1095-8649.2000.tb02110.x
Rikardsen A, Amundsen PA (2005) Pelagic marine feeding of Arctic charr and sea trout. J Fish Biol 66:1163–1166. https://doi.org/10.1111/j.0022-1112.2005.00655.x
Rikardsen A, Dempson J, Amundsen PA, Bjørn P, Finstad B, Jensen A (2007) Temporal variability in marine feeding of sympatric Arctic charr and sea trout. J Fish Biol 70:837–852. https://doi.org/10.1111/j.1095-8649.2007.01345.x
Ruggerone GT, Nielsen JL (2004) Evidence for competitive dominance of pink salmon (Oncorhynchus gorbuscha) over other salmonids in the North Pacific Ocean. Rev Fish Biol Fish 14:371–390
Ruggerone GT, Connors BM (2015) Productivity and life history of sockeye salmon in relation to competition with pink and sockeye salmon in the North Pacific Ocean. Can J Fish Aquat Sci 72:818–833. https://doi.org/10.1139/cjfas-2014-0134
Sandlund OT, Berntsen HH, Fiske P, Kuusela J et al (2019) Pink salmon in Norway: the reluctant invader. Biol Invasions 21:1033–1054. https://doi.org/10.1007/s10530-018-1904-z
Shapiro SS, Wilk MB (1965) An analysis of variance test for normality (complete samples). Biometrika 52:591–611. https://doi.org/10.2307/2333709
Shiomoto A, Tadokoro K, Nagasawa K, Ishida Y (1997) Trophic relations in the subarctic North Pacific ecosystem: possible feeding effect from pink salmon. Mar Ecol Prog Ser 150:75–85. https://doi.org/10.3354/meps150075
Sisk-Hackworth L, Kelley ST (2020) An application of compositional data analysis to multiomic time-series data. NAR Genom Bioinform 2:lqaa079. https://doi.org/10.1093/nargab/lqaa079
Skogseth R, Olivier LL, Nilsen F, Falck E et al (2020) Variability and decadal trends in the Isfjorden (Svalbard) ocean climate and circulation—an indicator for climate change in the European Arctic. Prog Oceanogr 187:102394. https://doi.org/10.1016/j.pocean.2020.102394
Skogstad OC, Skogstad Ø (2006) Dynamikk og ressursbruk hos anadrom og resident røye (Salvelinus alpinus) i Vårfluesjøen på Svalbard etter flere års fredning av bestanden. MSc thesis, Norwegian University of Life Sciences, Ås
Smedsrud LH, Esau I, Ingvaldsen RB, Eldevik T et al (2013) The role of the Barents Sea in the Arctic climate system. Rev Geophys 51:415–449. https://doi.org/10.1002/rog.20017
Svenning M-A, Borgstrøm R (1995) Population structure in landlocked Spitsbergen Arctic charr. sustained by cannibalism? Nord J Freshw Res 71:4224–4431
Svenning M-A, Gullestad N (2002) Adaptations to stochastic environmental variations: the effects of seasonal temperatures on the migratory window of Svalbard Arctic charr. In: Magnan P, Audet C, Glémet H, Legault M, Rodríguez MA, Taylor EB (eds) Ecology, behaviour and conservation of the charrs, genus Salvelinus. Springer, Dordrecht, pp 165–174
Svenning M-A, Klemetsen A, Olsen T (2007) Habitat and food choice of Arctic charr in Linnévatn on Spitsbergen, Svalbard: the first year-round investigation in a High Arctic lake. Ecol Freshwat Fish 16:70–77. https://doi.org/10.1111/j.1600-0633.2006.00183.x
Svenning M-A (2010) Metodikk for prøvefiske etter røye på Svalbard. Report No. 645, Norwegian Institute for Nature Research. https://brage.nina.no/nina-xmlui/handle/11250/2642595. Accessed 3 Nov 2022
Svenning M-A, Bergane V, Borgstrøm R (2020) Sjørøya i Linnévassdraget. Report No. 1825, Norwegian Institute for Nature Research. https://brage.nina.no/nina-xmlui/bitstream/handle/11250/2653904/1825.pdf?sequence=5&isAllowed=y. Accessed 3 Nov 2022
Tallman RF, Saurette F, Thera T (1996) Migration and life history variation in Arctic charr, Salvelinus alpinus. Ecoscience 3:33–41. https://doi.org/10.1080/11956860.1996.11682312
Trites AW, Spitz J (2018) Diet. In: Würsig B, Thewissen JGM, Kovacs KM (eds) Encyclopedia of marine mammals. Elsevier, Amsterdam, pp 255–259
Tverberg V, Nøst OA, Lydersen C, Kovacs KM (2014) Winter sea ice melting in the Atlantic Water subduction area, Svalbard Norway. J Geophys Res Oceans 119:5945–5967. https://doi.org/10.1002/2014JC010013
Tverberg V, Skogseth R, Cottier F, Sundfjord A et al (2019) The Kongsfjorden transect: seasonal and inter-annual variability in hydrography. In: Hop H, Wiencke C (eds) The ecosystem of Kongsfjorden. Springer, Svalbard, pp 49–104
Vassilenko SV, Petryashov VV (2009) Illustrated keys to free-living invertebrates of Eurasian Arctic seas and adjacent deep waters, vol 1. University of Alaska Fairbanks, Fairbanks, AK
Von Quillfeldt CH, Øseth E (2016) Klimaendringer på Svalbard: effekter på naturmangfold og konsekvenser for den fremtidige naturforvaltningen. Report No. Short report 42, The Norwegian Polar Institute. https://brage.npolar.no/npolar-xmlui/handle/11250/2431605. Accessed 3 Nov 2022
Węsławski JM (1994) Gammarus (Crustacea, Amphipoda) from Svalbard and Franz Josef Land. Distribution and density. Sarsia 79:145–150. https://doi.org/10.1080/00364827.1994.10413553
Węsławski JM, Opalinski K, Legezynska J (2000) Life cycle and production of Onisimus litoralis [Crustacea, Amphipoda]: the key species in the Arctic soft sediment littoral. Pol Arch Hydrobiol 47:585–596
Węsławski JM, Kwaśniewski S, Stempniewicz L, Błachowiak-Samołyk K (2006) Biodiversity and energy transfer to top trophic levels in two contrasting Arctic fjords. Pol Polar Res 27:259–278
Weydmann-Zwolicka A, Prątnicka P, Łącka M, Majaneva S, Cottier F, Berge J (2021) Zooplankton and sediment fluxes in two contrasting fjords reveal Atlantification of the Arctic. Sci Total Environ 773:145599. https://doi.org/10.1016/j.scitotenv.2021.145599
Weydmann A, Carstensen J, Goszczko I, Dmoch K, Olszewska A, Kwasniewski S (2014) Shift towards the dominance of boreal species in the Arctic: inter-annual and spatial zooplankton variability in the West Spitsbergen Current. Mar Ecol Prog Ser 501:41–52. https://doi.org/10.3354/meps10694
Witkowski A, Głowacki P (2010) A record of pink salmon, Oncorhynchus Gorbuscha (Actinoptergii, Salmoniformes, Salmonidae), in the Revelva River, Hornsund area (SW Spitsbergen). Acta Ichthyol Piscat 40:87–89
Acknowledgements
We thank Marie-Anne Blanchet and Ulf Lindstrøm for their advice regarding data exploration. Bernt Bye created the map of the study region.
Funding
Open access funding provided by Norwegian Polar Institute. The study was funded by the Norwegian Research Council project ARK (project no. 313678), the Svalbard Environmental Protection Fund (RIS ID: 11416, RIS ID: 10847), the Norwegian Polar Institute, Akvaplan-niva and the Institute of Oceanography Polish Academy of Sciences, Sopot.
Author information
Authors and Affiliations
Contributions
All authors have agreed to be listed and have approved the submitted version of the manuscript. The following authors contributed to study design: GC, OB, KMK and CL; fieldwork: GC, CL, KMK and OB; labwork: JMW and GC; statistical analyses: OB; manuscript preparation: all authors.
Corresponding author
Ethics declarations
Competing interests
The authors have no competing interests to declare.
Ethical approval
No ethics approval was needed for this study, but all national and institutional guidelines and regulations were fulfilled during capture and handling of the animals used in the study – most importantly, the Ethical Guidelines for the Use of Animals in Research issued by the (Norwegian) National Committee for Research Ethics in Science and Technology (NENT), 2018.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Online Resource 1
Length of Arctic char (Salvelinus alpinus) from different areas in Svalbard (2015–2018). Boxes contain 25th, 50th (median line) and 75th percentiles, and whiskers stretch between the minimum and the maximum values (excluding outliers). Outliers (> 1.5 × higher or lower than upper or lower quartile) are represented by dots. Isfjorden (In. Is), Outer Isfjorden (Out. Is), Forlandsundet (Forl.), Kongsfjorden/Krossfjorden (K/K), North Spitsbergen (No. Spi.), and Wahlenbergfjorden (Wahl.) (TIF 345 KB)
Online Resource 2
Diet composition of different length classes of Arctic char (Salvelinus alpinus) from different areas in Svalbard (2015–2018), presented as reconstructed biomass (proportions) of five different prey classes (TIF 1119 KB)
Online Resource 3
Diet composition of different sex/maturity classes of Arctic char (Salvelinus alpinus) from different areas in Svalbard (2015–2018), presented as reconstructed biomass (proportions) of five different prey classes. There were no significant differences between the classes (TIF 844 KB)
Online Resource 4
Diet composition of Arctic char (Salvelinus alpinus) and pink salmon (Oncorhynchus gorbuscha) in Kongsfjorden and Krossfjorden, Svalbard during 2017, presented as reconstructed biomass (proportions) of Arctic and Atlantic prey types (TIF 314 KB)
Online Resource 5
Diet composition of Arctic char (Salvelinus alpinus) from different areas in Svalbard (2015–2018), presented as reconstructed biomass (proportions) of Arctic and Atlantic prey types. Isfjorden (In. Is), Outer Isfjorden (Out. Is), Forlandsundet (Forl.), Kongsfjorden/Krossfjorden (K/K), North Spitsbergen (No. Spi.), and Wahlenbergfjorden (Wahl.) (TIF 931 KB)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Bengtsson, O., Lydersen, C., Christensen, G. et al. Marine diets of anadromous Arctic char (Salvelinus alpinus) and pink salmon (Oncorhynchus gorbuscha) in Svalbard, Norway. Polar Biol 46, 1219–1234 (2023). https://doi.org/10.1007/s00300-023-03196-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-023-03196-8