
Abstract
Rapid climate change occurring in the Arctic may affect the diet of ecologically and culturally important northern fish species. Here, a systematic literature review was completed for eight fish species found across the North American Arctic, with a focus on Inuit Nunangat, to identify major prey items, summarize feeding strategies, and highlight data gaps. Arctic Char (Salvelinus alpinus), Dolly Varden Char (Salvelinus malma), Lake Trout (Salvelinus namaycush), Bull Trout (Salvelinus confluentus), Inconnu (Stenodus leucichthys), Lake Whitefish (Coregonus clupeaformis), Broad Whitefish (Coregonus nasus), and Burbot (Lota lota), were selected as species of interest due to their ecological and cultural importance. The 74 studies reviewed indicate that these species are generalist feeders that demonstrate wide dietary niches, as well as the tendency to avoid agonistic interactions by partitioning resources when they co-occur with an overlapping species. Across coastal, lacustrine, and riverine systems, the most commonly consumed prey items are insects (Diptera spp.), as well as benthic forage fishes such as sculpins (Family: Cottidae). Insects are major prey items in riverine systems, where diets appear to be more generalized, compared to lakes. Anadromous species in coastal waters most commonly feed on various crustaceans and forage fishes. Benthic forage fishes, insects, zooplankton, and mollusks are widely consumed prey items in Arctic lakes. Burbot, Inconnu, and resident Dolly Varden had the most specialized feeding strategies, due in part to their habitat requirements and morphology, while Lake Trout and resident Arctic Char often form multiple ecotypes in lakes, some with different feeding behaviors. Knowledge gaps regarding northern fish trophic ecology are highlighted, and in particular include riverine systems and winter foraging behavior. This review is intended to inform predictions regarding the impacts of climate change on fish tropic ecology and to guide future research.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Climate change is predicted to affect trophic dynamics of fishes and influence geographic range shifts (Brandt et al. 2002; Jackson and Mandrak 2002; Chu et al. 2005), with outcomes anticipated to be particularly acute for the Arctic (Prowse et al. 2006; Hayden et al. 2015; Jansen et al. 2020; IPCC 2022). As temperatures continue to rise, for instance, anadromous life histories may become less common, potentially increasing competition in freshwater (Reist et al. 2006b). The increased productivity of warmer waters may result in longer growing seasons (Reist et al. 2006a) but may also increase metabolism and therefore energy intake, potentially exacerbating current food limitations (McDonald et al. 1996). Environmental change and any resulting shifts in energy intake may also influence fish growth (Huang et al. 2021). Northward range shifts of sub-Arctic species can also impact the diet of Arctic fishes, as has already occurred with a shift to Capelin (Mallotus villosus) and Pacific Sandlance (Ammodytes hexapterus), from Arctic Cod (Boreogadus saida) and crustaceans as prey for Arctic Char (Salvelinus alpinus) in some parts of the species’ range (Falardeau et al. 2017; Yurkowski et al. 2018; Ulrich and Tallman 2021a). Range-expanding species may also benefit native species by providing an alternate source of nutrients in typically oligotrophic Arctic systems, as may occur with the continued expansion of Pacific salmon into the Canadian Arctic (Dunmall et al. 2013). These range-expanding fish may also, however, compete with Arctic fishes for food, as demonstrated with a high level of diet overlap between increasingly common Capelin and endemic Arctic Cod (McNicholl et al. 2016), and there is potential for trophic interactions between Arctic Char and Atlantic Salmon (Salmo salar) in the fresh water (Bilous and Dunmall 2020). The predicted northward expansion of sub-Arctic cyprinids, such as Lake Chub (Couesius plumbeus), may also result in competition with native planktivores, such as ciscoes (Coregonus spp.), and result in a shift in the prey of top predators (Reist et al. 2006a). Changes in species composition due to northward range expansions may be balanced, however, by potential losses of temperature-sensitive northern fishes, with implications cascading through the trophic web (Reist et al. 2006b). The predicted and apparent impacts of climate change on the trophic ecology of Arctic fishes are indeed complex, with reverberations extending across ecosystems (Reist et al. 2006a) and influencing the food security of northern communities (Lynch et al. 2016; Loring et al. 2019).
The feeding ecology of northern fishes is shaped by their environment, making them adapted to harsh Arctic conditions and, at the same time, particularly vulnerable to warming temperatures (Power et al. 2008; Reist et al. 2006a). Many species in Arctic waters display generalist feeding strategies, allowing them to exploit temporal variance in prey availability, as well as resources from multiple habitat types and trophic levels (Laske et al. 2018). This adaptation also minimizes agonistic interactions with other species, which conserves energy (Power et al. 2008). Generalist feeding strategies are often further coupled with migratory and facultative life history strategies, such as seasonal migrations for feeding or the presence of resident and anadromous forms, that allow selection of optimal feeding environments and food resources (Power et al. 2008). As such, most northern aquatic food chains are short, and end with one to three generalist terminal predator species. These predators often partition prey both within (e.g., specialized ecotypes), or between species when they occupy a similar trophic level (Power et al. 2008; Guzzo et al. 2016).
Arctic Char, Dolly Varden Char (Salvelinus malma), Lake Trout (Salvelinus namaycush), Bull Trout (Salvelinus confluentus), Broad Whitefish (Coregonus nasus), Lake Whitefish (Coregonus clupeaformis), Inconnu (Stenodus leucichthys), and Burbot (Lota lota) are ecologically and culturally important fishes in Arctic freshwater or coastal systems. These species are harvested for subsistence among many Indigenous communities and are identified as important species by northern stakeholders throughout the Canadian Arctic (Stewart et al. 2009; McNicholl et al. 2020). In addition to the importance of these species for the food security and nutrition of northern Indigenous communities, fishing is also an important part of Indigenous ways of life, helping to strengthen community networks, facilitates the transmission of knowledge, and preserve cultural traditions (Proverbs et al. 2020; Galappaththi et al. 2022). The western Arctic populations of Bull Trout and Dolly Varden Char (hereafter: “Dolly Varden”) are also categorized as Special Concern species under the Committee on the Status of Endangered Wildlife in Canada (COSEWIC) (COSEWIC 2011, 2013). Additionally, some species, such as Arctic Char, are commercially fished (Galappaththi et al. 2022). Finally, these fishes serve as an important energetic link as a mid-trophic species among coastal ecosystems and as upper trophic predators in freshwater ecosystems.
In order to inform predictions regarding the effects of climate change on culturally and ecologically important fishes in the North American Arctic, a comprehensive overview of available information is required. This was done using a systematic literature review and focusing on eight key Arctic fish species occurring in riverine, lacustrine, and coastal habitats across Inuit Nunangat (traditional Inuit homelands consisting of Nunavut, the Inuvialuit Settlement Region, Nunavik, and Nunatsiavut), while also considering information gathered in the rest of the North American Arctic (the Yukon, Alaska, and the remainder of the Northwest Territories). Our objectives here are to summarize the diet information and feeding strategies of fish species with varying life-history types (resident or anadromous, where applicable) by geographic region, and highlight data gaps related to trophic ecology for these Arctic fishes. This review focuses on the North American Arctic because it was conceptualized around fish species relevant in Inuit Nunangat, and is intended to be most applicable to that region, while also including information on fish species that may become more relevant in Inuit Nunangat as thermal barriers are lowered by climate change. While focused here on the North American Arctic, these results will aid predictions regarding the impacts of climate change on northern fishes more broadly and guide future research that will, together, inform resulting management decisions and improve our understanding regarding the effects of climate change on Arctic aquatic ecosystems.
Methods
A systematic literature review was conducted to gain a thorough appraisal of diet information on eight selected fish species (Arctic Char, Dolly Varden, Lake Trout, Bull Trout, Inconnu, Lake Whitefish, Broad Whitefish, and Burbot) throughout Inuit Nunangat, as well as the rest of the North American Arctic. The geographic scope of this review was restricted to studies conducted in the Yukon, Northwest Territories (NWT), Nunavut, Nunavik, Nunatsiavut, and Alaska, which encompasses the entirety of the selected species’ geographic distributions in the North American Arctic. While the eight species selected were chosen primarily due to their relevance in Inuit Nunangat, the large spatial scale of this study was required in order to compile a sizeable base of information for these species, given that there is limited knowledge of their feeding ecology in northern ecosystems. Studies investigating the diets of both freshwater populations, as well as those that make marine migrations to feed, were included. To maintain consistency with the language used in the majority of studies on fish that make marine feeding migrations, the term “anadromous” was used, rather than more specific terms such as “diadromous” and “amphidromous.”
Web of Science and the Canadian Department of Fisheries and Oceans (DFO) internal catalogue were chosen as the main literature sources to use a publicly available search engine, provide a broader temporal coverage, and to search technical reports that are not published as scientific journal articles. The search string terms (Appendix 1) were the same for both searches in Web of Science and within the DFO catalogue, but the syntax was modified by removing the Web of Science-specific field tags for the DFO catalogue search. The search results were imported into an Excel (2016) spreadsheet in CSV (comma separated value) format and arranged into a database that contained title, authors, journal, abstract, and DOI.
Papers were subjected to an initial screening of titles and abstracts, followed by full-text screening. Initially, only titles and abstracts of the papers were screened according to the exclusion criteria (Appendix 2) to eliminate papers that did not discuss the target species, were not found within the geographic range, and/or did not have diet information obtained through stable isotope, stomach content, and/or fatty acid analyses. Stomach contents were searched for in order to meet the objective of determining specific prey items, while stable isotope and fatty acid analyses were searched for to give insight into trophic levels and broader patterns of resource use. After screening of abstracts and titles, remaining papers were then subjected to full text screening. First, papers were assessed if they contained feeding data, which was done by examining the methods and results sections, followed by a CTRL + F word search for the terms “diet,” “forag*,” and “feed,” in case diet information was available elsewhere in the paper.
Following screening, the included papers were coded according to a list of questions (Appendix 3) into a database that collated the relevant information contained in the papers. The coding questions were developed to record the requisite data for each species of interest, establish regional and temporal trends in feeding, as well as list diet items from stomach contents to the lowest taxonomic level (i.e., order for invertebrates, species for fishes).
The information about diet and feeding strategies (defined for the purposes of this study as the degree of generalism or specialization in the types of prey consumed, as well as factors that affect feeding behavior such as habitat use, daily or seasonal migrations, and interactions with other species) was summarized by species and region. The range of prey items was assessed, which allowed for primary prey items to be identified, as well as variations resulting from habitat type or region. If applicable, interactions with other species were summarized to provide context for the feeding strategies displayed by the focal species, especially in cases where they interacted with one another. Tables 1–10 (available as supplementary files) were constructed to give a more thorough list of recorded prey items, while also highlighting knowledge gaps by showing where diet information has been collected in each species’ range (denoted by a “1”—present or “0”—absent, respectively). Regions outside of a species’ range were denoted with “N/A,” the ranges being verified in Lindsey (1962), Scott and Crossman (1973), Wilson and Hebert (1996), Sawatzky et al. (2007), Stapanian et al. (2010), Mee et al. (2015), and Taylor (2016).
Results
Search string and coding
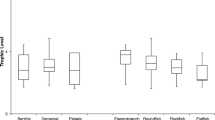
The search string on Web of Science returned 323 papers and the search of the DFO catalogue returned 42, for a total of 365. Of these, 291 were excluded due to not meeting the various inclusion criteria (Appendix 2), leaving 74 inclusions to be coded. Of the included publications, 11 were DFO reports, the rest being academic journal articles, and spanned from 1955 to 2022. Studies were mapped to demonstrate their geographical distribution and the number of studies done per system, with Alaska, Nunavut, and the NWT hosting the majority (Fig. 1). Studies were largely performed in the open-water season from late spring to early fall (Fig. 2). Arctic Char was the most frequently studied species (n = 34), followed by Lake Trout and Dolly Varden (n = 20 and 17, respectively, Fig. 3). Analyses of stomach contents was the most common method used (n = 63), followed by stable isotope analyses (n = 26) and fatty acid analyses (n = 3). The results are summarized by species, and within each species, with the information grouped by life history (i.e., resident or anadromous) if applicable, and throughout the region of known distribution (i.e., Northwest Territories, Yukon, Nunavut, Alaska, Nunavik, and Nunatsiavut) (Online Resource 1, Tables 1–10).

a. Map showing the distribution and frequency of diet studies per-species in Alaska, the Yukon and the mainland NWT. b. Map showing the distribution and frequency of diet studies per-species in Nunavut and Banks Island, NWT. c. Map showing the distribution and frequency of diet studies per-species in Nunavik and Nunatsiavut

Studies performed by season. Some studies included more than one season. Seasons were defined as summer (June–August), Fall (September–October), Winter (November-February), and Spring (March–May)

Number of available diet studies published per species in the North American Arctic. Numbers include anadromous and freshwater resident forms of all applicable species
Arctic Char
Anadromous form
Anadromous Arctic Char feed heavily on marine crustaceans and fishes, as well as mollusks and other invertebrates in some cases (Online Resource 1, Table 1). Anadromous individuals in the NWT feed extensively on forage fishes such as sandlance (Ammodytes spp.) and Capelin (Harwood et al. 2015; Harwood and Babaluk 2014). This is a shift away from diet data collected in the region during the late 1970’s, which saw Arctic Char diets mainly consist of Arctic Cod, mysids, and amphipods (Harwood et al. 2015). It is not known if these differences result from shifts in prey availability, predator preferences, temporal variation, or some combination thereof. Similarly, prior to 2005, most Arctic Char in Nunavut primarily fed on amphipods and mysids, which composed of up to ~ 80% of prey biomass, with other prey, such as Arctic Cod and sculpins (Family: Cottidae) making up the remainder (Ulrich and Tallman 2021a). After 2005, Capelin began to make up 65–75% of Arctic Char diets, while previous prey items were consumed in far lower numbers, or not at all (Yurkowski et al. 2018; Ulrich and Tallman 2021a). A similar increase in Capelin as prey, however, has not occurred across all of Nunavut. In Frobisher Bay for instance, amphipods and mysids made up nearly 95% of prey biomass consumed by Arctic Char as recently as 2009 (Spares et al. 2012) (Online Resource 1, Table 1). Individuals in Tariyunnuaq in the Kitikmeot region of Nunavut also feed on Capelin, as well as Pacific Herring (Clupea pallasii) and zooplankton but the dominant prey species is unknown (Swanson et al. 2011). Anadromous individuals in the northern areas of the archipelago consume mostly crustaceans, but information in these regions is sparse (Peet 1979). Arctic Char are opportunistic feeders in the marine environment, and their diets will likely shift in response to climate-driven changes to the marine Arctic food web. Falardeau et al. (2022) observed changes in the ∂13C and ∂15N signatures of individuals in the Kitikmeot Sea, Nunavut, which potentially reflect climate-driven diet shifts. Explanations for this shift could be a result the expansions in the distribution of boreal prey species, increased pelagic offshore foraging to escape warming nearshore waters, or an increase of pelagic prey availability due to earlier sea ice breakup and melting (Falardeau et al. 2022).
Anadromous Arctic Char in Nunatsiavut, off the north coast of Labrador, are largely piscivorous but also feed on crustaceans, depending on the population, and individual size (Cote et al. 2021). Long-term data collected in this region suggests that fishes such as Capelin and various Sculpins make up ~ 72% of prey biomass for Arctic Char, averaged across years and individual populations (Dempson et al. 2002, 2008). Amphipods, mysids, and other invertebrates are also preyed upon, but only make up the majority of diets in individuals smaller than 300 mm fork length (FL) (Dempson et al. 2002).
Resident form
Resident Arctic Char are primarily lacustrine generalist predators but, unlike their anadromous form, they feed in freshwater exclusively and tend to be smaller and less piscivorous. When another top predator is present (e.g., Lake Trout), resident Arctic Char forego piscivory entirely and prey more heavily on invertebrates (Fraser and Power 1989; Klobucar and Budy 2020). Such invertebrates include insects (especially chironomids and other aquatic emergents), zooplankton, and gastropods (Fraser and Power 1989; Kidd et al. 1998).
Data on resident Arctic Char in the NWT are limited to summer foraging. Individuals are noted to prey on a variety of invertebrates in the Thomsen River on Banks Island, with 65% of prey items being stoneflies, craneflies, or chironomids (Stephenson 2010) (Online Resource 1, Table 2). Overall, there is limited information on the diet of resident Arctic Char in the NWT.
Resident Arctic Char in Nunavut most frequently consume insects, zooplankton, and crustaceans (Gallagher and Dick 2011, 2010; Curtis et al. 1995) (Online Resource 1, Table 3). Larger individuals (> 350 mm FL) feed on small fishes (including cannibalism), except when co-occurring with Lake Trout (Kidd et al. 1998; Young et al. 2021; Vanriel and Johnson 1995). During the winter, feeding becomes less frequent but piscivorous individuals often become more cannibalistic during the winter, with smaller Arctic Char being eaten by up to 33% of large-form individuals, even in cases where they had not been cannibals during the summer (Gallagher and Dick 2010; Young et al. 2021; Young and Tallman 2021). In high-Arctic lakes such as Lake Hazen and Char Lake, which do not feature a forage fish species, cannibalism becomes the only form of piscivory (Hobson and Welch 1995; Guiguer et al. 2002; Gallagher et al. 2009; Sinnatamby et al. 2012).
Resident Arctic Char in Nunavik and Nunatsiavut (particularly Gander Lake) show similar diet proclivities, with larger individuals being more piscivorous, feeding on smaller Arctic Char, Threespine Stickleback (Gasterosteus aculeatus), and Ninespine Stickleback (Pungitus pungitus), among other species (O’Connell and Dempson 2002; Power et al. 2005) (Online Resource 1, Table 2). Smaller individuals feed more on invertebrates, such as chironomids and other aquatic and emergent insects (Online Resource 1, Table 2). Resident Arctic Char in southwest Alaska are more frequently piscivorous, perhaps due to a greater diversity of forage fishes being present in these lower-latitude systems (Woods et al. 2013; Fournier and Schindler 2021). Feeding on salmon eggs and juveniles by Arctic Char occasionally occurs in these areas but to a lesser extent than do Dolly Varden, which may feed extensively on salmon eggs and juveniles found in the streams of this region. Arctic Char tend to occupy lakes and are therefore restricted to preying on Sockeye Salmon (Oncorhynchus nerka) populations which have runs connected to such lakes (Morton 1982; Stewart et al. 2009; Dennert et al. 2016; Fournier and Schindler 2021). Arctic Char in this region have also been recorded to consume small mammals such as shrews (Woods et al. 2013).
At the species level, resident Arctic Char are generalists but in many lakes they diverge into 2–4 distinct morphs or ecotypes which can be segregated on the basis of habitat use and feeding strategies (Reist et al. 1995; Power et al. 2005, 2008). Arctic Char become piscivorous between 200 and 350 mm FL, although this can be affected by the presence of other large piscivores such as Lake Trout which will often exclude even large Arctic Char from piscivory (Fraser and Power 1989; Klobucar and Budy 2020). Outside of this scenario, large Arctic Char can form a piscivorous ecotype, often cannibalizing smaller individuals, especially in high-latitude systems with no other fish species available. Additionally, in some cases a large planktivore ecotype will also arise, as well as small and large benthivorous ecotypes (Power et al. 2008).
Dolly Varden
Anadromous form
Anadromous Dolly Varden feed primarily in coastal areas, where they consume fishes and crustaceans, feeding less frequently as they overwinter in freshwater (Online Resource 1, Table 3) (Stewart et al. 2009). Available information indicates anadromous Dolly Varden are heavily piscivorous at larger sizes (400 mm FL) (Stewart et al. 2009). Smaller individuals will primarily prey upon crustaceans such as mysids and amphipods, as well as chironomids in areas with a strong freshwater influence. In the Beaufort Sea, Dolly Varden often feed on sympagic (i.e., those which spent at least part of their life cycles in, or on, the sea ice) amphipods, and follow sea ice movements to do so, indicating their feeding habitat use is influenced by sea ice position and condition (Gallagher et al. 2021). Large, piscivorous individuals in the Beaufort Sea feed on a variety of fishes such as Fourhorn Sculpin (Myoxocephalus quadricornis), small whitefishes (Coregonus spp.), and others (Stewart et al. 2009) (Online Resource 1, Table 3). Large anadromous Dolly Varden in western Alaska feed primarily on marine fishes, such as Pacific Sandlance and Capelin, while smaller individuals feed on crustaceans, especially amphipods and malacostracans (Narver and Dahlberg 1965; Hart et al. 2015). When returning to freshwater in the fall, anadromous Dolly Varden feed infrequently until the spring migration, although various insects may be consumed during this time (Stewart et al. 2009). An exception occurs in southwestern Alaska, where some anadromous Dolly Varden make shorter coastal migrations and time their return to freshwater in the early fall to coincide with Pacific salmon runs (Sergeant et al. 2014). This allows these individuals to gorge themselves on lipid-rich salmon eggs to sustain themselves during spawning and overwintering (Sergeant et al. 2015). Salmon smolts are occasionally consumed during the winter as well (Hart et al. 2015).
Resident form
Resident Dolly Varden are primarily insectivorous and almost exclusively inhabit rivers and streams in Alaska and the Canadian Arctic. Chironomids are an important prey item, alongside other emergent insects, making up 75% or more of prey items consumed by resident Dolly Varden in many cases (Milner 1994; Parker and Huryn 2006; Stewart et al. 2009) (Online Resource 1, Table 4). Terrestrial insects and aquatic beetles are also occasionally eaten, with terrestrial insects becoming more prevalent as prey items later in the year (Stewart et al. 2009; Wipfli 1997). Resident Dolly Varden in the Babbage and Firth rivers in the Yukon occasionally eat amphipods and oligochaetes, which is not commonly recorded elsewhere (Stewart et al. 2009). Some large individuals will feed on small fishes, such as Arctic Grayling (Thymallus arcticus), as well as fish eggs, but this is not common compared to feeding on insects (Stewart et al. 2009). An exception to this is in western Alaska (e.g., Unalakleet, Chena, Iliamna, and upper Yukon rivers), where Dolly Varden ranges overlap with spawning Pacific salmon, and eggs and out-migrating juveniles, as well as scavenged flesh and dipteran larvae from dead adults make up the majority of diets while available (Armstrong 1970; Denton et al. 2009, 2010; Jaecks and Quinn 2014; Schoen et al. 2021). Resident individuals in an Alaskan stream feed less during winter, though they still feed on larval dipterans and stoneflies (Huryn and Benstead 2019).
Resident Dolly Varden are riverine insect specialists, with other types of invertebrates and small fishes only consumed incidentally (and only by large individuals in the case of the latter) and there appear to be no instances of resident Dolly Varden shifting their primary prey away from insects (Stewart et al. 2009). This feeding strategy is possibly shaped by morphology, given that resident Dolly Varden have smaller gapes than similarly sized Bull Trout and Arctic Char, which would necessitate feeding on smaller prey, even for larger individuals (Stewart et al. 2007). The feeding habitat use of Dolly Varden while drift foraging can be shaped by dominance hierarchies among individuals (Cullen and Grossman 2019). Larger dominant individuals occupy upstream positions in streams which allows for the first opportunity to consume drifting prey and consequently, higher foraging rates (Cullen and Grossman 2019).
Lake Trout
In Great Bear Lake, NWT, Lake Trout are noted to differentiate among habitat use and diet, forming several distinct morphs or ecotypes (Blackie et al. 2003). These include a more generalist ecotype and more specialized ecotypes that feed on more specific prey or at specific lake areas, although fatty acid analysis indicates that there is significant diet variation within the morphs (Chavarie et al. 2016). The generalist morph feeds on crustaceans as well as Round Whitefish (Prosopium cylindraceum), Slimy Sculpin (Cottus cognatus), adult caddisflies, ants (Formicidae spp.), and small amounts of other insects (Chavarie et al. 2016). The specialized benthic morph feed heavily on ants, as well as bivalves and mysids, but fewer fish compared to other morphs (Chavarie et al. 2016, 2021). The generalist pelagic morph feed on pelagic forage fishes (presumably Coregonus spp.), as well as crustaceans and terrestrial ants (Chavarie et al. 2016). The fourth morph, a pelagic piscivore, feeds on coregonids and small Northern Pike (Esox lucius) but also occasionally switches to Trichoptera spp. and cannibalism of the benthic morph (Chavarie et al. 2016). Each of these morphs feeds on terrestrial insects to some degree, demonstrating that, despite being more specialized ecotypes, they are still opportunistic.
In Great Slave Lake, NWT, Lake Trout exist in three different morphs but their preferred feeding habitat appears to be more related to size rather than morph (Zimmerman et al. 2009). There are shallow and deep-water piscivorous morphs, and a zooplanktivorous morph, but all three tend to forage benthically, and shift to pelagic feeding at approximately 430 mm standard length (Zimmerman et al. 2009). Smaller individuals of piscivorous morphs prey on sculpins during their benthic phase, but switch to coregonids as they grow larger and shift to pelagic feeding (Zimmerman et al. 2009). All morphs in Great Slave Lake also consume terrestrial insects opportunistically (13% occurrence averaged across morphs) (Moshenko and Gilman 1983; Zimmerman et al. 2009). Diets of Lake Trout in Great Slave Lake change throughout the year in accordance to thermal conditions (MacKenzie et al. 2022). Early in the season, foraging for terrestrial insects and littoral fishes in shallow water is more common, but Lake Trout become barricaded from nearshore prey later in the summer when the lake’s upper layer reaches 15 °C (MacKenzie et al. 2022).
In smaller lakes in the NWT, Nunavut, and the Yukon, Lake Trout will feed on zooplankton, insects and fishes such as stickleback, Slimy Sculpin, and others ( Kidd 1996; Kidd et al. 1998; Hulsman et al. 2016) (Online Resource 1, Table 5). The ability of Lake Trout to resource partition is displayed in Alexie Lake, NWT, where Lake Trout, Burbot, and Northern Pike all inhabit similar trophic levels, but feed in different areas of the lake, according to stable isotope data (Guzzo et al. 2016). The diet of Lake Trout in the Thomsen River, NWT appears to be very generalized, likely due to the homogenous nature of a riverine system and contain an array of insects and fishes (Stephenson 2010). On rare occasions, Lake Trout are known to consume small mammals, as lemmings (Lemmini spp.) were found in several Lake Trout stomachs in the Thomsen River (Stephenson 2010).
Nunavut also features rare examples of anadromous Lake Trout populations that migrate to the sound of Tariyunnuaq, Nunavut, for summer feeding (Swanson et al. 2011). During these migrations to coastal areas, Lake Trout feed on Pacific Herring, Capelin, and marine crustaceans (Swanson et al. 2011). Anadromous populations of Lake Trout are present in Husky Lakes, NWT (Kissinger et al. 2016), but there is little detailed information published about their diets.
Lake Trout in Alaska and in Nunavik are also generalist predators. Most Alaskan systems featuring Lake Trout are similar to Toolik Lake, which are relatively shallow and lack pelagic prey (MacDonald and Hershey 1992). This leads larger Lake Trout to feed on benthic fishes and mollusks, while smaller individuals feed on zooplankton closer to shore (McDonald and Hershey 1992; Merrick et al. 1992; McDonald et al. 1996; Keyse et al. 2007). In Nunavik, Quebec, larger individuals (> 600 mm FL) prey heavily on other fishes and occasional crustaceans (branchiopods) (Benoit and Power 1983; Murdoch et al. 2013) (Online Resource 1, Table 5). Mid-size individuals (300–600 mm FL) are generalist foragers (stickleback, small Burbot, and juvenile salmonids) but also feed heavily on insects (craneflies and chironomids; Benoit and Power 1983). The smallest size class of Lake Trout in these Nunavik lakes (< 300 mm FL) feed mostly on dipterans (Benoit and Power 1983).
Bull Trout
The diet and feeding habits of Bull Trout in the Mackenzie River drainage are poorly understood, especially outside of summer. Juvenile Bull Trout feed on a broad array of adult and larval insects (Stewart et al. 2007) (Online Resource 1, Table 6). As Bull Trout grow larger, they become more piscivorous, with the shift in prey occurring when the individual is between 100 and 200 mm FL, and many individuals in lakes become exclusively piscivorous as adults (Stewart et al. 2007). In the NWT and Yukon, Slimy Sculpin, Arctic Grayling, Longnose Sucker (Catostomus catostomus), and juvenile or small Bull Trout are the preferred prey species. If available, large adults will also eat small or young birds, amphibians, snakes, and small mammals such as shrews and mice (Stewart et al. 2007). It is unknown if this species shifts its diet to avoid interspecific competition after it has reached typical piscivory sizes or if it exhibits prey preferences once it reaches such a size (Stewart et al. 2007).
Inconnu
Inconnu are a large, primarily piscivorous freshwater coregonid that are found in both resident and anadromous forms (Tallman and Howland 2017); however, diet data on Inconnu are sparse. Inconnu in both rivers and lakes feed almost entirely on a diverse range of fishes, while invertebrates tend to only be consumed by some juveniles (Fuller 1955; Little et al. 1998; Gallagher and Dick 2015; Stuby et al. 2018). An exception to this has been recorded in Selawik Lake, Alaska, where very large individuals feed on mysids, alongside fishes (Alt 1965). Additionally, Inconnu in the Upper Yukon and Chena rivers feed heavily on juvenile Chinook Salmon (Oncorhychus tshawytcha) when they are present. (Online Resource 1, Table 7). Anadromous individuals in the nearshore Beaufort Sea similarly feed primarily on fish, though some mysids are also consumed (32% by number of prey items consumed) (Lacho 1991).
Inconnu are comparatively specialized to other large predators because of their heavy reliance on piscivory, even in smaller individuals, in their relatively narrow habitat range (shallow, silty areas of lakes; large rivers, and brackish areas) (Fuller 1955; Smith and Sutton 2015). The linkage of individual size to level of piscivory is difficult to establish in Inconnu because small individuals occasionally eat invertebrates, but even smaller individuals have been discovered feeding on fish (Fuller 1955; Alt 1965). Thus, more work is needed to contextualize the prey preferences of Inconnu.
Lake Whitefish
Lake Whitefish is a benthic and occasionally pelagic, mid-trophic level coregonid species, primarily found in lacustrine environments and is a generalist benthic predator of aquatic invertebrates (Jessop et al. 1993). A distinct pelagic zooplanktivore ecotype forms in some instances (Bernatchez et al. 1996). It is limited by its small, subterminal mouth which is adapted for benthivory, though instances of piscivory have been recorded outside the Arctic (Pothoven and Madenjian 2013). The species is primarily freshwater but anadromous populations exist in the Mackenzie River Delta and in Alaska (Sawatzky et al. 2007). Benthic prey such as mollusks, aquatic insects, and crustaceans make up the majority of Lake Whitefish diets (Jessop et al. 1993; Kidd et al. 1996; Little et al. 1998; Mackenzie et al. 2022) In some systems, such as the Slave River, the relative importance of different prey items shifts throughout the year – ostracods make up over half of the diet during the spring, while corixids make up 68% of diets by the fall (Little et al. 1998). Lake Whitefish have a lower salinity tolerance than other coregonids, meaning that anadromous feeding is not especially common, individuals that do migrate to coastal areas to feed remain in nearshore areas (Sawatzky et al. 2007). These anadromous Lake Whitefish in Tuktoyaktuk Harbour and Liverpool Bay, NWT feed primarily on crustaceans and other invertebrates (Lacho 1991; Bond and Erickson 1993) (Online Resource 1, Table 8).
Broad Whitefish
Broad Whitefish are primarily an anadromous coregonid species that are generalist benthic foragers found throughout the coastal Beaufort Sea and Mackenzie River Delta; however, a lacustrine form also exists (Sawatzky et al. 2007). Anadromous Broad Whitefish found in the marine environment near Tuktoyaktuk, Northwest Territories, were found to feed heavily on copepods (98% by number), as well as some nematodes and Diptera larvae (Lacho 1991) (Online Resource 1, Table 9). Anadromous Broad Whitefish in Prudhoe Bay, Alaska feature a more varied diet, also feeding on copepods, but with other crustaceans and chironomids being present as well (Fechhelm et al. 1995). An exception in Broad Whitefish diets occurs in the Arctic Coastal Plain of Alaska, where adult dipterans made up 35% of the diets of several individuals (Laske et al. 2018). In comparison to Least Cisco (Coregonus sardinella) and Dolly Varden, Broad Whitefish occupy a lower trophic level which limits them to feeding on small mollusks and other invertebrates (Kline et al. 1998).
Burbot
Burbot are specialized piscivores that primarily inhabit benthic areas, often emerging at night to feed in littoral zones. In the NWT, adult Burbot in the Mackenzie Delta are exclusively piscivorous during the winter, feeding largely on juvenile Burbot and Northern Pike, as well as Ninespine Stickleback, among others (Gallagher and Dick 2015) (Online Resource 1, Table 10). Juveniles are also piscivorous, being highly cannibalistic, and eating young Northern Pike and Ninespine Stickleback (Gallagher and Dick 2015). To a lesser extent, juveniles also consume invertebrates (Online Resource 1, Table 10). This combination of invertebrates and small fishes has also been recorded in juveniles in the Slave River system (Little et al. 1998). Burbot piscivory increases with size, between 100 and 460 mm FL, reinforced by the ∂15N differences between adults and juveniles (Gallagher and Dick 2015). Young Burbot that prey on fish appear to grow faster, which may explain the prevalence of cannibalism, as rapid growth would enable juveniles to grow to where they are at less risk of predation (Gallagher and Dick 2015). Winter cannibalism may also be favored by sexually mature individuals, as the energy gained would be beneficial for spawning (Gallagher and Dick 2015). Cannibalism among Burbot appears to be more prevalent in the Mackenzie River Delta than throughout most of its North American range (Gallagher and Dick 2015). In Great Slave Lake, Burbot feed on juvenile coregonids, as well as forage fishes such as Ninespine Stickleback, Trout-Perch, and various minnows (MacKenzie et al. 2022). In the Yukon, Burbot are similarly piscivorous, with individuals in Fox and Laberge lakes consuming Slimy Sculpin and various coregonids (Kidd 1996). In Alaska, Burbot in the Chena and Upper Yukon rivers feed on coregonids as well as juvenile Chinook Salmon when available, while in the Kuskokwim River, individuals consume various forage fishes (Rausch and Adams 2000; Schoen et al. 2021) (Online Resource 1, Table 10). Burbot in Nunavik consume littoral forage fishes, presumably during nightly feeding migrations (Power et al. 2009).
Burbot are the most specialized among the species reviewed. These fish are almost solely piscivorous as adults, typically feeding on littoral and some benthic fishes after reaching 460 mm total length at the latest (Power et al. 2009; Gallagher and Dick 2015; Guzzo et al. 2016). The feeding strategies of Burbot are further specialized by its photophobic and crepuscular nature, they remain at the benthos in cooler waters and protected from sunlight during the days of the open water season until nightfall, when they migrate to littoral zones to forage (Cott et al. 2015; Guzzo et al. 2016). Burbot also prefer low-complexity and soft substrates at the bottom of these lake slopes that it can burrow in during the day, which further shapes its hunting behavior (Cott et al. 2015).
Knowledge gaps
While consolidating available information, this review also highlighted a number of knowledge gaps relating to the diets and feeding strategies of Arctic fishes. For instance, basic year-round diet information is still needed for many Arctic species across all life-stages, especially given than current information is primarily focused on the open-water season (Fig. 2). This is particularly apparent for Inconnu, Broad Whitefish, Lake Whitefish, and Bull Trout, which lack even basic diet data for much of their Arctic ranges (Online Resource 1, Tables 1–10). Assessing ontogenetic diet shifts is important across species as several fishes (e.g., Arctic Char, Bull Trout, Burbot) feature significant diet shifts after reaching a certain size, but it is unknown if such shifts occur in Inconnu, for example. As well, the available information focuses on lacustrine and coastal ecosystems, highlighting the need for an expansion of information about the diets of fishes in riverine habitats in the Arctic.
The limited number of fish diet studies in the Arctic leads to several geographic knowledge gaps. For instance, studies in the NWT tend to be focused on the northern Great Lakes and the Mackenzie Delta, while the majority of smaller lakes and the rest of the Mackenzie River upstream of the delta remain unstudied (Fig. 1a). The Yukon features a severe lack of diet studies, with key species such as Burbot, Inconnu, Lake Trout, Lake Whitefish, and Bull Trout having virtually no available diet information in the territory (Fig. 1a). Additionally, there were no studies found in this review for Nunavut that consider species other than Arctic Char and Lake Trout (Fig. 1b). The logistics of performing diet studies in remote northern areas are a limiting factor, which highlights the need to focus research efforts on constructing a baseline for each species across seasons and habitat types, to reduce redundant efforts. When comparing the diet studies reviewed here, it is apparent that variation in overall diets and feeding strategies within and among regions is low, with the biggest factor in differences between regions being latitude and its limitation on prey diversity. Therefore, studies conducted in one region are likely to be applicable to fish diets in other regions, provided the type of environment is broadly similar (i.e., data collected on a lacustrine population is unlikely to be relevant to riverine individuals of the same species). Research efforts should therefore be focused on assessing fish diets and feeding strategies on species and environments where information is sparse, extremely dated, or subject to rapid current changes, given the more extensive body of literature on the most well-studied species (i.e., lacustrine Arctic Char; Lake Trout in the northern Great Lakes and smaller Arctic lakes; resident Dolly Varden).
Discussion
This review gives a thorough appraisal of the diet and feeding behaviors of eight ecologically and culturally important mid- to upper-trophic fish species throughout the Canadian Arctic and Alaska. The species included in this review largely display generalist feeding strategies, though within-species specialization did occur in several instances (e.g., Power et al. 2005; Zimmerman et al. 2009). Diet generalization is regarded as an adaptation common in northern fishes as it both reduces costly agonistic interaction with other species and allows for rapid exploitation of varied prey (Power et al. 2008; Ulrich and Tallman 2021a). The majority of fish dietary research in the region is focused on lake-resident or anadromous species during the summer. Therefore, several data gaps were identified, the most prominent being the need for information about winter feeding across species and ecosystems, as well as a lack of information available for riverine species. While some species such as Lake Trout and Arctic Char are well-studied, available information for other species is either dated (e.g., Inconnu) or limited (e.g., Lake Whitefish and Bull Trout). Diet studies on Broad Whitefish are also few in number, although two studies were recently published (Leppi et al. 2022; Stanek et al. 2022). Brewster et al. (2016) also contains information about diets and trophic niches in Dolly Varden, Broad Whitefish, Lake Whitefish, Burbot, and Inconnu, although the information was not included here as the study was not detected during keyword searches in Web of Science. There is also very little diet information available in the Yukon for the species considered, aside from Dolly Varden. Assessing the diets of fishes that are predicted to expand into the Arctic with continued warming is also important to better predict the resulting interactions with native fishes in Arctic habitats (e.g., McNicholl et al. 2016; Harris et al. 2022). Documentation of Indigenous knowledge of fish diets, as well as parameters related to diet (e.g., flesh color and condition), is needed and would expand upon and provide additional context to current understandings (e.g., Zerehi 2016; Cott et al. 2018).
In anadromous species, crustaceans (especially copepods, amphipods, and mysids) are the most widely consumed type of prey during marine feeding migrations. This is likely due to their presence in high density patches in marine environments and ideal for northern anadromous fishes looking to maximize food intake during their short feeding periods (Ulrich and Tallman 2021a). Crustaceans make up a portion, and in many cases the majority, of diets for Arctic Char, Broad Whitefish, Lake Whitefish, Dolly Varden, and Inconnu during their marine feeding migrations. Larger individuals of these species also eat an array of fishes, most of which are benthic-dwelling coastal species such as sandlances, blennies, pricklebacks, and sculpins. The recent increase in Capelin in the diet of some Arctic Char populations appears to be an exception to this tendency (Dempson et al. 2008; Ulrich and Tallman 2021a). This is possibly a result of Capelin aggregating in large spawning shoals when Arctic Char begin their feeding migrations (Ulrich and Tallman 2021a).
Changes in sea ice breakup timing, movements, and melt onset may have impacts on the diets and feeding strategies of Arctic Char and Dolly Varden (Harris et al. 2022). In the Beaufort Sea, predation of crustaceans by these species is connected to sea ice conditions, because both species prey on sympagic crustacean species (Gallagher et al. 2021; Falardeau et al. 2022). In the Kitikmeot Sea, changes in ∂13C values in Arctic Char reflect a shift toward a higher level of pelagic prey being consumed, compared to sympagic or benthic prey sources. A potential mechanism for this shift is earlier sea ice breakups and melt onset, which results in a phytoplankton bloom and in turn, a greater abundance of pelagic crustaceans present when Arctic Char arrive at sea to feed (Falardeau et al. 2022). This pattern has been observed in other Arctic predators, such as Belugas (Delphinapterus leucas) (Brown et al. 2017). Additionally, the shift toward more ∂13C-poor prey could be explained by the warming of nearshore coastal waters, which necessitates individuals moving to cooler, offshore areas, where pelagic prey lower in carbon enrichment (Falardeau et al. 2022). Conversely, ∂15N signatures have increased over time, indicating that Arctic Char occupy higher trophic levels in the Kitikmeot Sea than in the past. Falardeau et al. (2022) suggest this may be due to the increased distribution of boreal prey species (i.e., Capelin and Pacific Sandlance) into Arctic waters. Boreal species have higher tropic levels and pelagic ∂13C signatures, which may explain the isotopic shift seen in Arctic Char, although further research is needed to fully understand this pattern (Falardeau et al. 2022). While Arctic Char is an adaptable feeder that can react to shifts in the food web, the species has a narrow thermal range, and the physiological consequences of warming waters will likely restrict the species’ adaptability (Gilbert and Tierney 2018).
In riverine environments, diet overlap seems to be more extensive and resource partitioning is less common, possibly due to the more confined nature of such systems, or because resources may be partitioned by territorial behavior over prime feeding areas rather than species feeding on different prey items (Hearn 1987; Arostegui and Quinn 2018). In the Thomsen River, NWT, Lake Trout and resident Arctic Char share a large number of invertebrate prey sources, with chironomids and arachnids being the only ones exclusive to Arctic Char (Stephenson 2010). In Iliamna Lake, Alaska and its surrounding steams, Arctic Char exclude Rainbow Trout (Oncorhynchus mykiss) from piscivory and from feeding on benthic invertebrates alongside forage fishes; the Rainbow Trout are limited to insects (Arostegui and Quinn 2018). Conversely, in the tributaries of the same system, stream-resident Dolly Varden have significant diet overlap with stream-resident Rainbow Trout, indicating that these environments are less conducive to resource partitioning (Arostegui and Quinn 2018). Insects appear to be the most important group of prey for most fish in riverine systems, especially chironomids, trichopterans, and plecopterans, as well as other emergent and aquatic insects. Even high-level predators, such as Lake Trout and Inconnu, appear to feed upon insects more commonly when in rivers (Alt 1965; Stephenson 2010).
Lakes demonstrate clearer patterns of diet resource use and partitioning. Benthic fishes such as sculpins and sticklebacks are important prey for most piscivores, as many higher latitude Arctic lakes lack pelagic forage fishes (McDonald and Hershey 1992). Littoral foraging on prey fishes is relatively uncommon among the species considered here, limited mostly to Burbot in accordance to their nightly migrations to such areas to feed (Power et al. 2009; Guzzo et al. 2016). Inconnu also appear to specialize in piscivory in shallower waters in Great Slave Lake, although it is not known if they are specifically targeting littoral species (Fuller 1955). The importance of benthic feeding in many of these lakes is reinforced by the prevalence of mollusks (gastropods and bivalves) in the diets of Lake Whitefish, Arctic Char, and Lake Trout, with the latter two species forming benthivore-specialist ecotypes in some cases (Power et al. 2008; Chavarie et al. 2016). Zooplankton is also an important food source, especially for Arctic Char and small Lake Trout, the former of whom form a zooplanktivorous ecotype to avoid competition with piscivorous conspecifics or Lake Trout, while the latter are sometimes restricted to nearshore planktivory by larger individuals (Merrick et al. 1992; Kidd et al. 1998; Power et al. 2008).
Differences in prey items across the ranges in each species appears to be governed mostly by latitude, as food chains in the higher Arctic are simpler and less diverse (Power et al. 2008). Beyond the differences in prey diversity inherent to latitude, overall diet preferences and feeding strategies do not vary markedly within, and across, regions. Piscivorous resident Arctic Char feed on a more diverse array of fishes in western Alaska, Nunatsiavut, and Nunavik than in the High Arctic, due to the larger variety of different forage fishes at lower latitudes (O’Connell and Dempson 2002; Power et al. 2009; Dennert et al. 2016). There is a greater array of invertebrates eaten by Arctic Char in these regions as well, such as leeches, hemipterans, Odonata spp., Ephemoptera spp., and various Gastropods (O’Connell and Dempson 2002; Power et al. 2009; Woods et al. 2013; Dennert et al. 2016). Arctic Char in places such as northern Nunavut have simpler diets, relying heavily on emergent insects and bivalves, with piscivory restricted only to the large piscivore ecotype which preys on smaller Arctic Char and the few available forage fishes such as sticklebacks (Guiger et al. 2002; Gallagher and Dick 2011; Ulrich and Tallman 2021b). Lake Trout also show some differences in their diets across their range due to changes in environment. The Lake Trout of large, deep lakes, such as Great Slave Lake and Great Bear Lake, feature a greater emphasis for some individuals on pelagic feeding upon various fishes such as ciscoes and whitefishes, as well as various crustaceans (Zimmerman et al. 2009; Chavarie et al. 2016), but at higher latitudes in smaller and shallower lakes, Lake Trout instead feed very generally on mollusks, oligochaetes, and benthic fishes (McDonald and Hershey 1992; Keyse et al. 2007; Stephenson 2010).
Continued understanding of trophic ecology and interactions of Arctic fishes for food resources will assist predictions of how native fish populations may respond to a rapidly changing Arctic (Niemi et al. 2019), or potential outcomes resulting from climate-driven distributional shifts to the north (Dunmall et al. 2013; Bilous and Dunmall 2020). Future research that addresses these gaps will contribute toward a robust baseline of diet information for northern fishes and improve understanding regarding Arctic fishes and the potential effects of climate change on Arctic ecosystems.
References
Alt KT (1965) Food habits of Inconnu in Alaska. Trans Am Fish Soc 94:272–274. https://doi.org/10.1577/1548-8659(1965)94[272:FHOIIA]2.0.CO;2
Armstrong RH (1970) Age, food, and migration of Dolly Varden smolts in Southeastern Alaska. J Fish Board Can 27:991–1004. https://doi.org/10.1139/f70-116
Arostegui MC, Quinn T (2018) Trophic ecology of nonanadromous rainbow trout in a post-glacial lake system: partial convergence of adfluvial and fluvial forms. Can J Zool 96:818–827. https://doi.org/10.1139/cjz-2017-0201
Benoit J, Power G (1983) Biology of two arctic populations of lake trout Salvelinus namaycush (Walbaum) of the Lake Minto region of new Quebec. Can Transl Fish Aquat Sci 4920:ii + 16 p
Bernatchez L, Vuorinen JA, Bodaly RA, Dodson JJ (1996) Genetic evidence for reproductive isolation and multiple origins of sympatric trophic ecotypes of whitefish (Coregonus). Evolution 50:624–635. https://doi.org/10.1111/j.1558-5646.1996.tb03873.x
Bilous M, Dunmall KM (2020) Atlantic salmon in the Canadian Arctic: potential dispersal, establishment, and interaction with Arctic char. Rev Fish Biol Fish 30:463–483. https://doi.org/10.1007/s11160-020-09610-2
Blackie CT, Weese DJ, Noakes DLG (2003) Evidence for resource polymorphism in the lake charr (Salvelinus namaycush) population of Great Bear Lake, Northwest Territories, Canada. Ecoscience 10:509–514. https://doi.org/10.1080/11956860.2003.11682799
Bond WA, Erickson RN (1993) Fisheries investigations in the coastal waters of Liverpool Bay, Northwest Territories. Can Tech Rep Fish Aquat Sci 2204:vi + 51 p
Brandt SB, Mason DM, McCormick MJ, Lofgren B, Hunter TS (2002) Climate change: Implications for fish growth performance in the Great Lakes. Am Fish Soc Symp 32:61–76
Brewster JD, Giraldo C, Swanson HK, Walkusz W, Loewen TN, Reist JD, Stern GA, Loseto LL (2016) Ecological niche of coastal Beaufort Sea fishes defined by stable isotopes and fatty acids. Mar Ecol Prog Ser 559:159–173. https://doi.org/10.3354/meps11887
Brown TA, Chrytsal E, Ferguson SH, Yurkowski DJ, Watt C, Hussey NE, Kelley TC, Belt ST (2017) Coupled changes between the H-Print biomarker and δ15N indicates a variable sea ice carbon contribution to the diet of Cumberland Sound beluga whales. Limnol Oceanogr 62:1606–1619. https://doi.org/10.1002/lno.10520
Chavarie L, Howland KL, Gallagher CP, Tonn W (2016) Fatty acid signatures and stomach contents of four sympatric Lake Trout: assessment of trophic patterns among morphotypes in Great Bear Lake. Ecol Freshw Fish 25:109–124. https://doi.org/10.1111/eff.12195
Chavarie L, Howland KL, Harris LN, Gallagher CP, Hansen MJ, Tonn WM, Muir AM, Krueger CC (2021) Among-individual diet variation within a lake trout ecotype: lack of stability of niche use. Ecol Evol 11:1457–1475. https://doi.org/10.1002/ece3.7158
Chu C, Mandrak NE, Minns CK (2005) Potential impacts of climate change on the distributions of several common and rare freshwater fishes in Canada. Divers Distrib 11:299–310. https://doi.org/10.1111/j.1366-9516.2005.00153.x
COSEWIC (2011) COSEWIC Response Statement – Dolly Varden (Western Arctic populations) in Canada. Committee on the Status of Endangered Wildlife in Canada, Ottawa. https://registrelep-sararegistry.gc.ca/virtual_sara/files/statements/rs_1122_429_2011-9_e.pdf. Accessed 20 May 2022
COSEWIC (2013) COSEWIC Response Statement - Bull Trout (Western Arctic populations) in Canada. Committee on the Status of Endangered Wildlife in Canada, Ottawa. https://www.sararegistry.gc.ca/virtual_sara/files/statements/rs_1202_620_2013-9_e.pdf. Accessed 20 May 2022
Cote D, Dempson JB, Piersiak M, Layton K, Roul S, Laing R, Angnatok J, Bradbury I (2021) Using movement, diet, and genetic analyses to understand Arctic charr responses to ecosystem change. Mar Ecol Prog Ser 673:135–149. https://doi.org/10.3354/meps13775
Cott PA, Guzzo MM, Chapelsky AJ, Milne SW, Blanchfield PJ (2015) Diel bank migration of Burbot (Lota lota). Hydrobiologia 757:3–20. https://doi.org/10.1007/s10750-015-2257-6
Cott PA, Amos AL, Guzzo MM, Chavrie L, Goater CP, Muir DCG, Evans MS (2018) Can traditional methods of selecting food accurately assess fish health? Arct Sci 4:205–222. https://doi.org/10.1139/as-2017-0052
Cullen JL, Grossman GD (2019) Aggressive interactions affect foraging and use of space in a drift foraging salmonid, Salvelinus malma (Salmoniformes: Salmonidae). Zool J Linn Soc 187:774–781. https://doi.org/10.1093/zoolinnean/zlz050
Curtis MA, Berube M, Stenzel A (1995) Parasitological evidence for specialized foraging behavior in lake-resident arctic char (Salvelinus alpinus). Can J Fish Aquat Sci 52:186–194. https://doi.org/10.1139/f95-526
Dempson JB, Shears M, Bloom M (2002) Spatial and temporal variability in the diet of anadromous Arctic charr, Salvelinus alpinus, in northern Labrador. In: Magnan P, Audet C, Glemet H, Legault M, Rodriguez MA, Taylor EB (eds) Ecology, behaviour and conservation of the charrs, genus Salvelinus. Dev Env Biol Fishes, vol 22, Springer, Dordrecht, pp 49–62. https://doi.org/10.1007/978-94-017-1352-8_4
Dempson JB, Shears M, Furey G, Bloom M (2008) Resilience and stability of north Labrador Arctic charr, Salvelinus alpinus, subject to exploitation and environmental variability. Env Biol Fish 83:57–67. https://doi.org/10.1007/s10641-007-9261-2
Dennert AM, May-McNally SL, Bond MH, Quinn TP, Taylor EB (2016) Trophic biology and migratory patterns of sympatric Dolly Varden (Salvelinus malma) and Arctic char (Salvelinus alpinus). Can J Zool 94:529–539. https://doi.org/10.1139/cjz-2016-0004
Denton KP, Rich HB, Quinn TP (2009) Diet, movement, and growth of Dolly Varden in response to sockeye salmon subsidies. Trans Am Fish Soc 138:1207–1219. https://doi.org/10.1577/T09-006.1
Denton KP, Rich HB, Moore JW, Quinn TP (2010) The utilization of a Pacific salmon Oncorhynchus nerka subsidy by three populations of charr Salvelinus spp. J Fish Biol 77:1006–1023. https://doi.org/10.1111/j.1095-8649.2010.02746.x
Dunmall KM, Reist JD, Carmack EC, Babaluk JA, Heide-Jørgensen MP, Docker MF (2013) Pacific salmon in the Arctic: Harbingers of change. In: Mueter FJ, Dickson DMS, Huntington HP, Irvine JR, Logerwell EA, MacLean SA, Quakenbush LT, Rosa C (eds) Responses of Arctic marine ecosystems to climate change. University of Alaska Fairbanks, Alaska Sea Grant, pp 141–163
Falardeau M, Bouchard C, Robert D, Fortier L (2017) First records of Pacific sand lance (Ammodytes hexapterus) in the Canadian Arctic Archipelago. Polar Biol 40:2291–2296. https://doi.org/10.1007/s00300-017-2141-0
Falardeau M, Bennett EM, Else B, Fisk A, Mundy CJ, Choy ES, Ahmed MM, Harris LN, Moore J (2022) Biophysical indicators and Indigenous and Local Knowledge reveal climatic and ecological shifts with implications for Arctic Char fisheries. Glob Env Chang 74:102469. https://doi.org/10.1016/j.gloenvcha.2022.102469
Fechhelm RG, Griffiths WB, Bryan JD, Gallaway BJ, Wilson WJ (1995) Application of an in-situ growth-model: Inferred instance of interspecific trophic competition between anadromous fishes of Prudhoe Bay, Alaska. Trans Am Fish Soc 124:55–69. https://doi.org/10.1577/1548-8659(1995)124%3C0055:AOAISG%3E2.3.CO;2
Fournier EB, Schindler DE (2021) Depth-specific benthic specialization of Arctic char in an oligotrophic subarctic lake. Aquat Sci 83:1–11. https://doi.org/10.1007/s00027-021-00827-2
Fraser NC, Power G (1989) Influences of lake trout on lake-resident arctic char in northern Quebec, Canada. Trans Am Fish Soc 118:36–45. https://doi.org/10.1577/1548-8659(1989)118%3C0036:IOLTOL%3E2.3.CO;2
Fuller WA (1955) The Inconnu (Stenodus-leucichthys mackenziei) in Great Slave Lake and adjoining waters. J Fish Res Board Can 12:768–780. https://doi.org/10.1139/f55-042
Galappaththi EK, Falardeau M, Harris LN, Rocha JC, Moore JS, Berkes F (2022) Resilience-based steps for adaptive co-management of Arctic small-scale fisheries. Env Res Lett 17:083004. https://doi.org/10.1088/1748-9326/ac7b37
Gallagher CP, Dick TA (2010) Trophic structure of a landlocked Arctic char Salvelinus alpinus population from southern Baffin Island, Canada. Ecol Freshw Fish 19:39–50. https://doi.org/10.1111/j.1600-0633.2009.00387.x
Gallagher CP, Dick TA (2011) Ecological characteristics of ninespine stickleback Pungitius pungitius from southern Baffin Island, Canada. Ecol Freshw Fish 20:646–655. https://doi.org/10.1111/j.1600-0633.2011.00516.x
Gallagher CP, Dick TA (2015) Winter feeding ecology and the importance of cannibalism in juvenile and adult burbot (Lota lota) from the Mackenzie Delta, Canada. Hydrobiologia 757:73–88. https://doi.org/10.1007/s10750-015-2227-z
Gallagher CP, Dick TA, Babaluk JA, Reist JD (2009) Parasite community of Arctic Char, Salvelinus alpinus, from Lake Hazen and Craig Lake, Quttinirpaaq National Park, Nunavut. Can Tech Rep Fish Aquat Sci 2854:vi + 24 p
Gallagher CP, Courtney MB, Seitz AC, Lea EV, Howland KL (2021) Ocean-entry timing and marine habitat-use of Canadian Dolly Varden: Dispersal among conservation, hydrocarbon exploration, and shipping areas in the Beaufort Sea. Estuar Coast Shelf Sci 262:107609. https://doi.org/10.1016/j.gloenvcha.2022.102469
Gilbert MJH, Tierney KB, Ferry L (2018) Warm northern river temperatures increase post-exercise fatigue in an Arctic migratory salmonid but not in a temperate relative. Funct Ecol 32:687–700. https://doi.org/10.1111/1365-2435.13006
Guiguer KRRA, Reist JD, Power M, Babaluk JA (2002) Using stable isotopes to confirm the trophic ecology of Arctic charr morphotypes from Lake Hazen, Nunavut, Canada. J Fish Biol 60:348–362. https://doi.org/10.1111/j.1095-8649.2002.tb00285.x
Guzzo MM, Blanchfield PJ, Chapelsky AJ, Cott PA (2016) Resource partitioning among top-level piscivores in a sub-Arctic lake during thermal stratification. J Gt Lakes Res 42:276–285. https://doi.org/10.1016/j.jglr.2015.05.014
Harris L, Moore JS, Dunmall KM, Evans MS, Falardeau M, Gallagher CP, Gilbert MJH, Kenny T, McNicholl DG, Norman M, Lyall G, Kringayark L (2022) Arctic char in a rapidly changing North. Polar Knowledge: Aqhaliat Report 4:34–57. https://doi.org/10.35298/pkc.2021.02.eng
Hart LM, Bond MH, May-McNally SL, Miller JA, Quinn TP (2015) Use of otolith microchemistry and stable isotopes to investigate the ecology and anadromous migrations of Northern Dolly Varden from the Egegik River, Bristol Bay, Alaska. Environ Biol Fish 98:1633–1643. https://doi.org/10.1007/s10641-015-0389-1
Harwood LA, Smith TG, George JC, Sandstrom SJ, Walkusz W, Divoky GJ (2015) Change in the Beaufort Sea ecosystem: Diverging trends in body condition and/or production in five marine vertebrate species. Prog Oceanogr 136:263–273. https://doi.org/10.1016/j.pocean.2015.05.003
Harwood LA, Babaluk JA (2014) Spawning, overwintering and summer feeding habitats used by anadromous Arctic Char (Salvelinus alpinus) of the Hornaday River, Northwest Territories, Canada. Arctic 67:449–461. https://doi.org/10.14430/arctic4422
Hayden B, Harrod C, Sonninen E, Kahilainen KK (2015) Seasonal depletion of resources intensifies trophic interactions in subarctic freshwater fish communities. Freshw Biol 60:1000–1015. https://doi.org/10.1111/fwb.12564
Hearn WE (1987) Interspecific competition and habitat segregation among stream-dwelling trout and salmon: a review. Fisheries 12:24–31. https://doi.org/10.1577/1548-8446(1987)012%3C0024:ICAHSA%3E2.0.CO;2
Hobson KA, Welch HE (1995) Cannibalism and trophic structure in a high Arctic lake: insights from stable-isotope analysis. Can J Fish Aquat Sci 52:1195–1201. https://doi.org/10.1139/f95-116
Huang M, Ding L, Wang J, Ding C, Tao J (2021) The impacts of climate change on fish growth: a summary of conducted studies and current knowledge. Ecol Indic 121:106976. https://doi.org/10.1016/j.ecolind.2020.106976
Hulsman MF, Cahill CL, Erwin AC, Lunn BD, Tonn WM, Howland KL, Bakix ABM (2016) Influence of potential fish competitors on Lake Trout trophic ecology in small lakes of the Barrenlands, NWT, Canada. J Gt Lakes Res 42:290–298. https://doi.org/10.1016/j.jglr.2015.07.014
Huryn AD, Benstead JP (2019) Seasonal changes in light availability modify the temperature dependence of secondary production in an Arctic stream. Ecology 100:1–15. https://doi.org/10.1002/ecy.2690
IPCC (2022) Climate change 2022: Impacts, adaptation and vulnerability. Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. Pörtner HO, Roberts DC, Tignor M, Poloczanska ES, Mintenbeck K, Alegría A, Craig M, Langsdorf S, Löschke S, Möller V, Okem A, Rama B (eds) Cambridge University Press, Cambridge, UK and New York, NY, USA. https://doi.org/10.1017/9781009325844.
Jackson DA, Mandrak NE (2002) Changing fish biodiversity: Predicting the loss of cyprinid biodiversity due to global climate change. Am Fish Soc Symp 32:89–98
Jaecks T, Quinn TP (2014) Ontogenetic shift to dependence on salmon-derived nutrients in Dolly Varden char from the Iliamna River, Alaska. Env Biol Fish 97:1323–1333. https://doi.org/10.1007/s10641-014-0221-3
Jansen E, Christensen JH, Dokken T, Nisancioglu KH, Vinther BM, Capron E, Guo C, Jensen MF, Langen PL, Pedersen RA, Yang S, Bentsen M, Kjær HA, Sadatzki H, Sessford E, Stendel S (2020) Perspectives on the present era of abrupt Arctic climate change. Nat Clim Chang 10:714–721. https://doi.org/10.1038/s41558-020-0860-7
Jessop EF, Chang-Kue KTJ, MacDonald G (1993) Fish resource data from Indin Lake, Northwest Territories. Can Data Rep Fish Aquat Sci 907:v + 52 p
Keyse MD, Fortino K, Hershey AE, O’Brien WJ, Lienesch PW, Luecke C, McDonald ME (2007) Effects of large lake trout (Salvelinus namaycush) on the dietary habits of small lake trout: a comparison of stable isotopes (delta N-15 and delta C-13) and stomach content analyses. Hydrobiologia 579:175–185. https://doi.org/10.1007/s10750-006-0399-2
Kidd KA (1996) Use of stable nitrogen isotope ratios to characterize food web structure and organochlorine accumulation in subarctic lakes in Yukon Territory. PhD Thesis, University of Alberta.
Kidd KA, Hesslein RH, Ross BJ, Koczanski K, Stephens GR, Muir DCG (1998) Bioaccumulation of organochlorines through a remote freshwater food web in the Canadian Arctic. Env Pollut 102:91–103. https://doi.org/10.1016/S0269-7491(98)00068-2
Kissinger BC, Gantner N, Anderson WG, Gillis DM, Halden NM, Harwood LA, Reist JD (2016) Brackish-water residency and semi-anadromy in Arctic lake trout (Salvelinus namaycush) inferred from otolith microchemistry. J Gt Lakes Res 42:267–275. https://doi.org/10.1016/j.jglr.2015.05.016
Kline TC, Wilson WJ, Goering JJ (1998) Natural isotope indicators of fish migration at Prudhoe Bay, Alaska. Can J Fish Aquat Sci 55:1494–1502. https://doi.org/10.1139/f98-012
Klobucar SL, Budy P (2020) Trophic structure of apex fish communities in closed versus leaky lakes of arctic Alaska. Oecologia 194:491–504. https://doi.org/10.1007/s00442-020-04776-9
Lacho G (1991) Stomach Content Analyses of Fishes from Tuktoyaktuk Harbour, N.W.T, 1981. Can Data Rep Fish Aquat Sci 853:iv + 10 p
Laske SM, Rosenberger AE, Wipfli MS, Zimmerman CE (2018) Generalist feeding strategies in Arctic freshwater fish: a mechanism for dealing with extreme environments. Ecol Freshw Fish 27:767–784. https://doi.org/10.1111/eff.12391
Leppi JC, Rinella DJ, Wipfli MS, Whitman MS (2022) Broad whitefish (Coregonus nasus) isotopic niches: Stable isotopes reveal diverse foraging strategies and habitat use in Arctic Alaska. PLoS ONE. https://doi.org/10.1371/journal.pone.0270474
Lindsey CC (1962) Distinctions between the Broad Whitefish, Coregonus nasus, and other North American whitefishes. J Fish Board Can 19:687–714. https://doi.org/10.1139/f62-046
Little AS, Tonn WM, Tallman RF, Reist JD (1998) Seasonal variation in diet and trophic relationships within the fish communities of the lower Slave River, Northwest Territories, Canada. Environ Biol Fish 53:429–445. https://doi.org/10.1023/A:1007433400648
Loring PA, Fazzino DV, Agapito M, Chuenpagdee R, Gannin G, Isaacs M (2019) Fish and food security in small-scale fisheries. In: Chuenpagdee R, Jentoft S (eds) Transdisciplinarity for small-scale fisheries governance: Analysis and Practice. MARE Publ Ser 21, Springer, Cham. https://doi.org/10.1007/978-3-319-94938-3_4
Lynch AJ, Cooke SJ, Deines AM, Bower SD, Bunnell DB, Cowx IG, Nguyen VM, Nohner J, Phouthavong K, Riley B, Rogers MW, Taylor WW, Woelmer W, Youn S, Douglas BT (2016) The social, economic and environmental importance of inland fish and fisheries. Environ Rev 24:115–121. https://doi.org/10.1139/er-2015-0064
MacKenzie CJA, Fortin BL, Stevens CE (2022) Summary of Ecological Information Relevant to Great Slave Lake Fisheries. Can Manuscr Rep Fish Aquat Sci 3214:vii + 63 p
McDonald ME, Hershey AE (1992) Shifts in abundance and growth of slimy sculpin in response to changes in the predator population in an arctic Alaskan lake. Hydrobiologia 240:219–223. https://doi.org/10.1007/978-94-011-2720-2_17
McDonald ME, Hershey AE, Miller MC (1996) Global warming impacts on lake trout in arctic lakes. Limnol Oceanogr 41:1102–1108. https://doi.org/10.4319/lo.1996.41.5.1102
McNicholl DG, Walkusz W, Davoren GK, Majewski JD (2016) Dietary charictaristics of co-occurring polar cod (Boreogadus saida) and capelin (Mallotus villosus) in the Canadian Arctic, Darnley Bay. Polar Biol 39:1099–1108. https://doi.org/10.1007/s00300-015-1834-5
McNicholl DG, Dunmall KM, Majewski AR, Niemi A, Gallagher CP, Sawatzky CD, Reist JD (2020) Distribution of marine and anadromous fishes of Darnley Bay and the Anguniaqvia Niqiqiyuam Protected Area, NT. Can Tech Rep Fish Aquat Sci 3394:x + 90 p.
Mee JA, Bernatchez L, Reist JD, Rogers SM, Taylor EB (2015) Identifying designatable units for intraspecific conservation prioritization: a hierarchical approach applied to the lake whitefish species complex (Coregonus spp.). Evol Appl 8:423–441. https://doi.org/10.1111/eva.12247
Merrick GW, Hershey AE, Mcdonald ME (1992) Salmonid diet and the size, distribution, and density of benthic invertebrates in an arctic lake. Hydrobiologia 240:225–233. https://doi.org/10.1007/BF00013464
Milner AM (1994) Colonization and succession of invertebrate communities in a new stream in Glacier Bay National Park, Alaska. Freshw Biol 32:387–400. https://doi.org/10.1111/j.1365-2427.1994.tb01134.x
Morton WM (1982) Comparative catches and food-habits of Dolly Varden and Arctic charrs, Salvelinus malma and Salvelinus alpinus, at Karluk, Alaska, in 1939–1941. Environ Biol Fish 7:7–28. https://doi.org/10.1007/BF00011819
Moshenko RW, Gillman DV (1983) Creel Census and Biological Data from the Lake Trout Sport Fishery on Great Bear and Great Slave Lakes, Northwest Territories, 1977–78. Can Data Rep Fish Aquat Sci 389:vi + 73 p
Murdoch A, Klein G, Doidge DW, Power M (2013) Assessing the food web impacts of an anadromous Arctic charr introduction to a sub-Arctic watershed using stable isotopes. Fish Manag Ecol 20:302–314. https://doi.org/10.1111/fme.12012
Narver DW, Dahlberg ML (1965) Estuarine food of Dolly Varden at Chignik Alaska. Trans Am Fish Soc 94:405–408. https://doi.org/10.1577/15488659(1965)94[405:EFODVA]2.0.CO;2
Niemi A, Ferguson S, Hedges K, Melling H, Michel C, Ayles B, Azetsu-Scott K, Coupel P, Deslauriers D, Devred E, Doniol-Valcroze T, Dunmall KM, Eerts J, Galbraith P, Geoffery M, Gilchrist G, Hennin H, Howland KL, Kendall M, Kohlbach D, Lea E, Loseto L, Majewski A, Marcoux M, Matthews C, McNicholl DG, Mosnier A, Mundy C, Ogloff W, Perrie W, Richards C, Richardson E, Reist JD, Roy V, Sawatzky CD, Scharffenberg K, Tallman RF, Tremblay J-E, Tufts T, Watt C, Williams B, Worden E, Yurkowski DJ, Zimmerman, S (2019) State of Canada’s Arctic seas. Can Tech Rep Fish Aquat Sci 3344:xv + 189 p
O’Connell MF, Dempson JB (2002) The biology of Arctic charr, Salvelinus alpinus, of Gander Lake, a large, deep, oligotrophic lake in Newfoundland, Canada. Environ Biol Fish 64:115–126. https://doi.org/10.1023/A:1016001423937
Parker SM, Huryn AD (2006) Food web structure and function in two arctic streams with contrasting disturbance regimes. Freshw Biol 51:1249–1263. https://doi.org/10.1111/j.1365-2427.2006.01567.x
Peet RF (1979) Data on the biology of Arctic char from the eastern Arctic, Northwest Territories. Fish Mar Serv Manuscr Rep 1456:vi + 43 p
Pothoven SA, Madenjian CP (2013) Increased piscivory by Lake Whitefish in Lake Huron. N Am J Fish Manag 33:1194–1202. https://doi.org/10.1080/02755947.2013.839973
Power M, O’Connell MF, Dempson JB (2005) Ecological segregation within and among Arctic char morphotypes in Gander Lake, Newfoundland. Env Biol of Fishes 73:263–274. https://doi.org/10.1007/s10641-005-2137-4
Power M, Reist JD, Dempson JB (2008) Fish in high-latitude Arctic lakes. In: Vincent WF, Laybourn-Parry J (eds) Polar lakes and rivers. Oxford University Press, Oxford, pp 249–267
Power M, Power G, Reist JD, Bajno R (2009) Ecological and genetic differentiation among the Arctic charr of Lake Aigneau, Northern Quebec. Ecol Freshw Fish 18:445–460. https://doi.org/10.1111/j.1600-0633.2009.00362.x
Proverbs TA, Lantz TC, Lord SI, Amos A, Gwich’in Tribal Council Department of Cultural Heritage, Ban NC (2020) Social-ecological determinants of access to fish and well-being in four Gwich’in communities in Canada’s Northwest Territories. Hum Ecol 48:155–171. https://doi.org/10.1007/s10745-020-00131-x
Prowse TD, Wrona FJ, Reist JD, Gibson JJ, Hobbie JE, Levesque MJL, Vincent WE (2006) Climate change effects on hydroecology of Arctic freshwater ecosystems. Ambio 35:347–358. https://doi.org/10.1579/0044-7447(2006)35[347:CCEOHO]2.0.CO;2
Rausch RL, Adams AM (2000) Natural transfer of helminths of marine origin to freshwater fishes, with observations on the development of Diphyllobothrium alascense. J Parasitol 86:319–327. https://doi.org/10.1645/0022-3395(2000)086[0319:NTOHOM]2.0.CO;2
Reist JD, Gyselman E, Babaluk JA, Johnson JD, Wissink R (1995) Evidence for two morphotypes of arctic char (Salvelinus alpinus (L.)) from Lake Hazen, Ellesmere Island, Northwest Territories. Can Nord J Freshw Res 71:396–410
Reist JD, Wrona FJ, Prowse TD, Power M, Dempson JB, Beamish RJ, King JR, Carmichael TJ, Sawatzky CD (2006a) General effects of climate change on Arctic fishes and fish populations. Ambio 35:370–380. https://doi.org/10.1579/0044-7447(2006)35[370:GEOCCO]2.0.CO;2
Reist JD, Wrona FJ, Prowse TD, Power M, Dempson JB, King JR, Beamish RJ (2006b) An overview of effects of climate change on selected Arctic freshwater and anadromous fishes. Ambio 35:381–387. https://doi.org/10.1579/0044-7447(2006)35[381:AOOEOC]2.0.CO;2
Sawatzky CD, Michalak D, Reist JD, Carmichael TJ, Mandrak NE, Heuring LG (2007) Distributions of freshwater and anadromous fishes from the mainland Northwest Territories, Canada. Can Manuscr Rep Fish Aquat Sci 2793:xiv + 239 p
Schoen ER, Sellmer KW, Wipfli MS, Lopez JA, Ivanoff R, Meyer BE (2021) Piscine predation on juvenile salmon in sub-arctic Alaskan rivers: associations with season, habitat, predator size and streamflow. Ecol Freshw Fish. https://doi.org/10.1111/eff.12626
Scott WB, Crossman EJ (1973) Freshwater fishes of Canada. Fisheries Research Board of Canada, Ottawa
Sergeant CJ, Armstrong JB, Ward EJ (2014) Predator-prey migration phenologies remain synchronized in a warming catchment. Freshw Biol 60:724–732. https://doi.org/10.1111/fwb.12524
Sinnatamby RN, Babaluk JA, Power G, Reist JD, Power M (2012) Summer habitat use and feeding of juvenile Arctic charr, Salvelinus alpinus, in the Canadian High Arctic. Ecol Freshw Fish 21:309–322. https://doi.org/10.1111/j.1600-0633.2012.00552.x
Smith NJ, Sutton TM (2015) Mixed-stock Inconnu Stenodus leucichthys habitat use in an Arctic estuarine delta complex. Environ Biol Fish 98:857–869. https://doi.org/10.1007/s10641-014-0321-0
Spares AD, Stokesbury MJW, O’Dor RK, Dick TA (2012) Temperature, salinity and prey availability shape the marine migration of Arctic char, Salvelinus alpinus, in a macrotidal estuary. Mar Biol 159:1633–1646. https://doi.org/10.1007/s00227-012-1949-y
Stanek AE, von Biela VR, Laske SM, Taylor RL, Dunton KH (2022) Barrier islands influence the assimilation f terrestrial energy in nearshore fishes. Estuar Coast Shelf Sci 278:108094. https://doi.org/10.1016/j.ecss.2022.108094
Stapanian MA, Vl P, Madenjian CP, Jackson JR, Lappalainen J, Evenson MJ, Neufeld MD (2010) Worldwide status of burbot and conservation measures. Fish Fish 11:34–56. https://doi.org/10.1111/j.1467-2979.2009.00340.x
Stephenson SA (2010) Fishes of the Thomsen River, Banks Island, Northwest Territories. Can Manuscr Rep Fish Aquat Sci 2944:vi + 44 p
Stewart DB, Mochnacz NJ, Sawatzky CD, Carmichael, Reist JD (2007) Fish diets and food webs in the Northwest Territories: bull trout (Salvelinus confluentus). Can Manuscr Rep Fish Aquat Sci 2800:vi + 18 p
Stewart DB, Mochnacz NJ, Carmichael TJ, Sawatzky CD, Reist JD (2009) Fish diets and food webs in the Northwest Territories: Dolly Varden (Salvelinus malma). Can Manuscr Rep Fish Aquat Sci 2912:vi + 33 p
Stuby L (2018) Contributions to the Life History of Kuskokwim River Inconnu. Trans Am Fish Soc 147:879–890. https://doi.org/10.1002/tafs.10069
Swanson HK, Kidd KA, Reist JD (2011) Quantifying importance of marine prey in the diets of two partially anadromous fishes. Can J Fish Aquat Sci 68:2020–2028. https://doi.org/10.1139/f2011-111
Tallman RF, Howland KL (2017) Factors that influence productivity and vulnerability of Inconnu, Stenodus leucichthys nelma, populations in Canada. Fundam Appl Limnol 189:235–247. https://doi.org/10.1127/fal/2016/0815
Taylor EB (2016) The Arctic Char (Salvelinus alpinus) “complex” in North America revisited. Hydrobiologia 783:283–293. https://doi.org/10.1007/s10750-015-2613-6
Ulrich KL, Tallman RF (2021a) The Capelin invasion: evidence for a trophic shift in Arctic Char populations from the Cumberland Sound region, Nunavut, Canada. Arct Sci 7:413–435. https://doi.org/10.1139/as-2020-0001
Ulrich KL, Tallman RF (2021b) Multi-indicator evidence for habitat use and trophic strategy segregation of two sympatric forms of Arctic char from the Cumberland Sound region of Nunavut, Canada. Arct Sci 7:512–544. https://doi.org/10.1139/as-2019-0039
Vanriel P, Johnson L (1995) Action principles as determinants of ecosystem structure—the autonomous lake as a reference system. Ecol 76:1741–1757. https://doi.org/10.2307/1940707
Wilson CC, Herbert PDN (1996) Phylogeographic origins of Lake Trout (Salvelinus namaycush) in eastern North America. Can J Fish Aquat Sci 53:2764–2775. https://doi.org/10.1139/f96-223
Wipfli MS (1997) Terrestrial invertebrates as salmonid prey and nitrogen sources in streams: contrasting old-growth and young-growth riparian forests in southeastern Alaska, USA. Can J Fish Aquat Sci 54:1259–1269. https://doi.org/10.1139/f97-034
Woods PJ, Young D, Skulason S, Snorrason SS, Quinn TP (2013) Resource polymorphism and diversity of Arctic charr Salvelinus alpinus in a series of isolated lakes. J Fish Biol 82:569–587. https://doi.org/10.1111/jfb.12011
Young AL, Tallman RF (2021) The comparative lake ecology of two allopatric Arctic Charr, Salvelinus alpinus, populations with differing life histories in Cumberland Sound, Nunavut. Arct Sci 7:471–493. https://doi.org/10.1139/as-2019-0037
Young AL, Tallman RF, Ogle DH (2021) Life history variation in Arctic charr (Salvelinus alpinus) and the effects of diet and migration on the growth, condition, and body morphology of two Arctic charr populations in Cumberland Sound, Nunavut, Canada. Arct Sci 7:436–453. https://doi.org/10.1139/as-2019-0036
Yurkowski DJ, Hussey NE, Ferguson SH, Fisk AT (2018) A temporal shift in trophic diversity among a predator assemblage in a warming Arctic. R Soc Open Sci 5:1–12. https://doi.org/10.1098/rsos.180259
Zerehi SS (2016) Odd colouring in Nunavut's Arctic char leaves fishermen stumped. CBCNews. https://www.cbc.ca/news/canada/north/arctic-char-changes-1.3725161. Accessed 26 July 2022
Zimmerman MS, Schmidt SN, Krueger CC, Vander Zanden MJ, Eshenroder RL (2009) Ontogenetic niche shifts and resource partitioning of lake trout morphotypes. Can J Fish Aquat Sci 66:1007–1018. https://doi.org/10.1139/F09-060
Acknowledgements
Funding to support this review was provided by Fisheries and Oceans Canada and the Government of Northwest Territories Cumulative Impact Monitoring Program (CIMP 221). We wish to thank Colin Gallagher as well as two anonymous reviewers for their insightful comments and suggestions to improve this manuscript.
Funding
Open access funding provided by Fisheries & Oceans Canada.
Author information
Authors and Affiliations
Contributions
Conceptualization and design: KD. Study screening: KW. Data extraction: KW. Data analysis: KW. Writing—original draft: KW. Writing. Review and editing: KW, DM, KD.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflicts of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Appendices
Appendices
Appendix 1: Search strings
The following search strings were used to conduct the literature searches on Web of Science and the DFO catalogue. The search terms are identical and modified only to reflect differences in the syntax requirements of each search engine.
Web of Science search string:
(TS=(canad* OR alask* OR newfoundlan* OR quebec OR labrador* OR yukon OR northwest OR nunavut)) AND (TS=("arctic char*" OR "dolly varden" OR “inconnu” OR "sheefish" OR “burbot” OR "lake trout" OR “lake char*” OR "lake whitefish" OR "broad whitefish" OR "bull trout" OR "bull char*")) AND (TS=(diet* OR feed* OR “stable isotop*” OR “fatty acid” OR stomach OR prey OR food OR web OR trophic* OR niche)) NOT TS=(mercury OR contamin* OR pollution)
DFO collection search string:
(canad* OR alask*OR newfoundlan* OR quebec OR labrador* OR yukon OR northwest OR nunavut)) AND ("lake trout" OR “lake char*” OR "arctic char*" OR "inconnu" OR “sheefish” OR "dolly varden" OR "lake whitefish" OR "broad whitefish" OR "bull trout" OR “bull char*” OR "burbot") AND (diet OR feed* OR stomach OR "stable isotop*" OR "fatty acid*" OR food OR web OR trophic* OR niche) NOT (pollution OR contamin* OR mercury)
Appendix 2: Exclusion criteria
Bases on which papers were included or excluded in the sample
Dimension | Inclusion | Exclusion |
|---|---|---|
Geographic scope | Inuit Nunangat and the rest of Arctic Canada and Alaska | BC, Alberta, Saskatchewan, Manitoba, Ontario, Quebec outside of Nunavik, lower 49 US states, the rest of the world |
Language | English and French | Non-English and non-French |
Focus | Diet/Trophic ecology | Anything else |
Publication type | Case studies, research articles, technical reports (DFO), manuscript reports (DFO), and research documents (DFO) | Models, predictions |
Temporal scope | Current and past | Future projections |
Species | Dolly Varden, Arctic Char, inconnu, broad whitefish, lake whitefish, bull trout, lake trout, burbot | Any other fish species, unless the focal species was a prey species for one of the 8 studied species, and included relevant feeding data in the paper |
Appendix 3: Coding questions
Questions designed for extracting diet information from each paper in a standardized manner and how that information was coded into the database
# | Column item | Description | Coding |
|---|---|---|---|
A | Included/excluded | Is it included or excluded from analysis? | 1 = Included; 0 = Excluded |
0.1 | Document Type | What kind of document is it? | J = Journal Article D = DFO report |
0.2 | Geographic focus | What state/province/territory does the study take place in? | Name(s) |
0.3 | Study area(s) | Name the bodies of water under study | Name(s) |
0.4 | Timeframe | When was the data collected? | Most specific dates available |
0.5 | Season(s) | What season(s) did the stufy occur in (Summer: Jun-Aug, Fall: Sept-Oct, Winter: Nov-Feb, Spring: Mar-May) | Sp = Spring, S = Summer, F = Fall, W = Winter |
1.1 | Species studied | Common names of species studies, alphabetically | Name(s) |
1.1.2 | Habitat type | Were specimens collected in still, coastal or flowing water systems? | F = Fluvial, L = Lacustrine,, A = Anadromous |
1.2 | Diet sampling method | What diet sampling technique(s) are used? | Mark “X” if applicable |
1.2.1 | Stomach contents | Were stomach contents analyzed? | “X” |
1.2.2 | Stable isotopes | Were stable isotopes analyzed? | “X” |
1.2.2.1 | Flesh type | Where was the flesh for SI analysis taken from? | List location(s) |
1.2.3 | Fatty acids | Were fatty acids analyzed? | “X” |
1.3 | General diet characteristics | What broad groups of prey were consumed? | Mark “X” if applicable |
1.3.1 | Fish | Were fish consumed? | “X” |
1.3.2 | Insects | Were insects or insect larva consumed? | “X” |
1.3.3 | Worms | Annelids, platyhelminthes, nematodes etc | “X” |
1.3.4 | Crustaceans | Amphipods, copepods etc | “X” |
1.3.5 | Molluscs | Gastropods, bivalves etc | “X” |
1.3.6 | Fish eggs | Eggs of other fish | “X” |
1.3.7 | Plankton | “X” | |
1.3.8 | Other | Other prey items: mammals, lizards, amphibians etc | “X” |
1.3.9 | Please specify | List the additional prey items | List |
1.4 | Specific diet components | List the specific prey items to order for invertebrates, species for fishes | List |
1.5 | Competition/overlap with other species | If the paper describes how species react to competitors, describe here | Describe |
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Wight, K.J., McNicholl, D.G. & Dunmall, K.M. A systematic review of the trophic ecology of eight ecologically and culturally important fish species in the North American Arctic. Polar Biol 46, 409–425 (2023). https://doi.org/10.1007/s00300-023-03133-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-023-03133-9