
Abstract
Numerous biological processes involve proteins capable of transiently assembling into subcellular compartments necessary for cellular functions. One process is the RNA polymerase II transcription cycle which involves initiation, elongation, co-transcriptional modification of nascent RNA, and termination. The essential yeast transcription termination factor Nab3 is required for termination of small non-coding RNAs and accumulates into a compact nuclear granule upon glucose removal. Nab3 nuclear granule accumulation varies in penetrance across yeast strains and a higher Nab3 granule accumulation phenotype is associated with petite strains, suggesting a possible ATP-dependent mechanism for granule disassembly. Here, we demonstrate the uncoupling of mitochondrial oxidative phosphorylation by drug treatment or deletions of nuclear-encoded ATP synthase subunit genes were sufficient to increase Nab3 granule accumulation and led to an inability to proliferate during prolonged glucose deprivation, which requires respiration. Additionally, by enriching for respiration competent cells from a petite-prone strain, we generated a low granule-accumulating strain from a relatively high one, providing another link between respiratory competency and Nab3 granules. Consistent with the resulting idea that ATP is involved in granule accumulation, the addition of extracellular ATP to semi-permeabilized cells was sufficient to reduce Nab3 granule accumulation. Deleting the SKY1 gene, which encodes a kinase that phosphorylates nuclear SR repeat-containing proteins and is involved in efficient stress granule disassembly, also resulted in increased granule accumulation. This observation implicates Sky1 in Nab3 granule biogenesis. Taken together, these findings suggest there is normally an equilibrium between termination factor granule assembly and disassembly mediated by ATP-requiring nuclear machinery.





Similar content being viewed by others


Data availability
All data generated or analyzed during this study are included in this published article [and its supplementary information files].
References
Alberti S, Gladfelter A, Mittag T (2019) Considerations and challenges in studying liquid–liquid phase separation and biomolecular condensates. Cell 176:419–434. https://doi.org/10.1016/j.cell.2018.12.035
Arndt KM, Reines D (2015) Termination of transcription of short noncoding RNAs by RNA polymerase II. Annu Rev Biochem 84:381–404. https://doi.org/10.1146/annurev-biochem-060614-034457
Ashe MP, De Long SK, Sachs AB (2000) Glucose depletion rapidly inhibits translation initiation in yeast. Mol Biol Cell 11:833–848. https://doi.org/10.1091/mbc.11.3.833
Baruffini E, Lodi T, Dallabona C, Puglisi A, Zeviani M, Ferrero I (2006) Genetic and chemical rescue of the Saccharomyces cerevisiae phenotype induced by mitochondrial DNA polymerase mutations associated with progressive external ophthalmoplegia in humans. Hum Mol Genet 15:2846–2855. https://doi.org/10.1093/hmg/ddl219
Boehning M, Dugast-Darzacq C, Rankovic M, Hansen AS, Yu T, Marie-Nelly H, McSwiggen DT, Kokic G, Dailey GM, Cramer P, Darzacq X, Zweckstetter M (2018) RNA polymerase II clustering through carboxy-terminal domain phase separation. Nat Struct Mol Biol 25:833–840. https://doi.org/10.1038/s41594-018-0112-y
Buchan JR, Kolaitis RM, Taylor JP, Parker R (2013) Eukaryotic stress granules are cleared by autophagy and Cdc48/VCP function. Cell 153:1461–1474. https://doi.org/10.1016/j.cell.2013.05.037
Buchan JR, Yoon JH, Parker R (2011) Stress-specific composition, assembly and kinetics of stress granules in Saccharomyces cerevisiae. J Cell Sci 124:228–239. https://doi.org/10.1242/jcs.078444
Campos-Melo D, Hawley ZCE, Droppelmann CA, Strong MJ (2021) The integral role of RNA in stress granule formation and function. Front Cell Dev Biol 9:621779. https://doi.org/10.3389/fcell.2021.621779
Carroll KL, Ghirlando R, Ames JM, Corden JL (2007) Interaction of yeast RNA-binding proteins Nrd1 and Nab3 with RNA polymerase II terminator elements. RNA 13:361–373. https://doi.org/10.1261/rna.338407
Cho WK, Spille JH, Hecht M, Lee C, Li C, Grube V, Cisse II (2018) Mediator and RNA polymerase II clusters associate in transcription-dependent condensates. Science 361:412–415. https://doi.org/10.1126/science.aar4199
Chong S, Dugast-Darzacq C, Liu Z, Dong P, Dailey GM, Cattoglio C, Heckert A, Banala S, Lavis L, Darzacq X, Tjian R (2018) Imaging dynamic and selective low-complexity domain interactions that control gene transcription. Science. https://doi.org/10.1126/science.aar2555
Corden JL (2008) Yeast Pol II start-site selection: the long and the short of it. EMBO Rep 9:1084–1086. https://doi.org/10.1038/embor.2008.192
Costanzo M, VanderSluis B, Koch EN, Baryshnikova A, Pons C, Tan G, Wang W, Usaj M, Hanchard J, Lee SD, Pelechano V, Styles EB, Billmann M, van Leeuwen J, van Dyk N, Lin ZY, Kuzmin E, Nelson J, Piotrowski JS, Srikumar T, Bahr S, Chen Y, Deshpande R, Kurat CF, Li SC, Li Z, Usaj MM, Okada H, Pascoe N, San Luis BJ, Sharifpoor S, Shuteriqi E, Simpkins SW, Snider J, Suresh HG, Tan Y, Zhu H, Malod-Dognin N, Janjic V, Przulj N, Troyanskaya OG, Stagljar I, Xia T, Ohya Y, Gingras AC, Raught B, Boutros M, Steinmetz LM, Moore CL, Rosebrock AP, Caudy AA, Myers CL, Andrews B, Boone C (2016) A global genetic interaction network maps a wiring diagram of cellular function. Science. https://doi.org/10.1126/science.aaf1420
Darby MM, Serebreni L, Pan X, Boeke JD, Corden JL (2012) The Saccharomyces cerevisiae Nrd1-Nab3 transcription termination pathway acts in opposition to ras signaling and mediates response to nutrient depletion. Mol Cell Biol 32:1762–1775. https://doi.org/10.1128/MCB.00050-12
Dichtl B (2008) Transcriptional shortcuts. Mol Cell 31:617–618. https://doi.org/10.1016/j.molcel.2008.08.014
Dimitrov LN, Brem RB, Kruglyak L, Gottschling DE (2009) Polymorphisms in multiple genes contribute to the spontaneous mitochondrial genome instability of Saccharomyces cerevisiae S288C strains. Genetics 183:365–383. https://doi.org/10.1534/genetics.109.104497
Gietz RD, Schiestl RH, Willems AR, Woods RA (1995) Studies on the transformation of intact yeast cells by the LiAc/SS-DNA/PEG procedure. Yeast 11:355–360. https://doi.org/10.1002/yea.320110408
Gilks N, Kedersha N, Ayodele M, Shen L, Stoecklin G, Dember LM, Anderson P (2004) Stress granule assembly is mediated by prion-like aggregation of TIA-1. Mol Biol Cell 15:5383–5398. https://doi.org/10.1091/mbc.e04-08-0715
Hunn JC, Hutchinson KM, Kelley JB, Reines D (2022) Variable penetrance of Nab3 granule accumulation quantified by a new tool for high-throughput single-cell granule analysis. Curr Genet. https://doi.org/10.1007/s00294-022-01234-2
Jain S, Wheeler JR, Walters RW, Agrawal A, Barsic A, Parker R (2016) ATPase-modulated stress granules contain a diverse proteome and substructure. Cell 164:487–498. https://doi.org/10.1016/j.cell.2015.12.038
Jenks MH, O’Rourke TW, Reines D (2008) Properties of an intergenic terminator and start site switch that regulate IMD2 transcription in yeast. Mol Cell Biol 28:3883–3893. https://doi.org/10.1128/MCB.00380-08
Kaliszewska M, Kruszewski J, Kierdaszuk B, Kostera-Pruszczyk A, Nojszewska M, Lusakowska A, Vizueta J, Sabat D, Lutyk D, Lower M, Piekutowska-Abramczuk D, Kaniak-Golik A, Pronicka E, Kaminska A, Bartnik E, Golik P, Tonska K (2015) Yeast model analysis of novel polymerase gamma variants found in patients with autosomal recessive mitochondrial disease. Hum Genet 134:951–966. https://doi.org/10.1007/s00439-015-1578-x
Kopcewicz KA, O’Rourke TW, Reines D (2007) Metabolic regulation of IMD2 transcription and an unusual DNA element that generates short transcripts. Mol Cell Biol 27:2821–2829. https://doi.org/10.1128/MCB.02159-06
Kuehner JN, Brow DA (2008) Regulation of a eukaryotic gene by GTP-dependent start site selection and transcription attenuation. Mol Cell 31:201–211. https://doi.org/10.1016/j.molcel.2008.05.018
Li H (2018) Minimap2: pairwise alignment for nucleotide sequences. Bioinformatics 34:3094–3100. https://doi.org/10.1093/bioinformatics/bty191
Longo VD, Liou LL, Valentine JS, Gralla EB (1999) Mitochondrial superoxide decreases yeast survival in stationary phase. Arch Biochem Biophys 365:131–142. https://doi.org/10.1006/abbi.1999.1158
Loya TJ, O’Rourke TW, Degtyareva N, Reines D (2013) A network of interdependent molecular interactions describes a higher order Nrd1-Nab3 complex involved in yeast transcription termination. J Biol Chem 288:34158–34167. https://doi.org/10.1074/jbc.M113.516765
Loya TJ, O’Rourke TW, Simke WC, Kelley JB, Reines D (2018) Nab3’s localization to a nuclear granule in response to nutrient deprivation is determined by its essential prion-like domain. PLoS ONE 13:e0209195. https://doi.org/10.1371/journal.pone.0209195
Lu H, Yu D, Hansen AS, Ganguly S, Liu R, Heckert A, Darzacq X, Zhou Q (2018) Phase-separation mechanism for C-terminal hyperphosphorylation of RNA polymerase II. Nature 558:318–323. https://doi.org/10.1038/s41586-018-0174-3
Matheson K, Parsons L, Gammie A (2017) Whole-genome sequence and variant analysis of W303, a widely-used strain of Saccharomyces cerevisiae. G3 (bethesda) 7:2219–2226. https://doi.org/10.1534/g3.117.040022
MATLAB, 2020. (R2021a), Natick, Massachusetts: The MathWorks, Inc.
Patel A, Malinovska L, Saha S, Wang J, Alberti S, Krishnan Y, Hyman AA (2017) ATP as a biological hydrotrope. Science 356:753–756. https://doi.org/10.1126/science.aaf6846
Rasmussen AK, Chatterjee A, Rasmussen LJ, Singh KK (2003) Mitochondria-mediated nuclear mutator phenotype in Saccharomyces cerevisiae. Nucleic Acids Res 31:3909–3917. https://doi.org/10.1093/nar/gkg446
Sabari BR, Dall’Agnese A, Boija A, Klein IA, Coffey EL, Shrinivas K, Abraham BJ, Hannett NM, Zamudio AV, Manteiga JC, Li CH, Guo YE, Day DS, Schuijers J, Vasile E, Malik S, Hnisz D, Lee TI, Cisse II, Roeder RG, Sharp PA, Chakraborty AK, Young RA (2018) Coactivator condensation at super-enhancers links phase separation and gene control. Science. https://doi.org/10.1126/science.aar3958
Sathyanarayanan U, Musa M, Bou Dib P, Raimundo N, Milosevic I, Krisko A (2020) ATP hydrolysis by yeast Hsp104 determines protein aggregate dissolution and size in vivo. Nat Commun 11:5226. https://doi.org/10.1038/s41467-020-19104-1
Schindelin J, Arganda-Carreras I, Frise E, Kaynig V, Longair M, Pietzsch T, Preibisch S, Rueden C, Saalfeld S, Schmid B, Tinevez JY, White DJ, Hartenstein V, Eliceiri K, Tomancak P, Cardona A (2012) Fiji: an open-source platform for biological-image analysis. Nat Methods 9:676–682. https://doi.org/10.1038/nmeth.2019
Shattuck JE, Cascarina SM, Paul KR, Ross ED (2020) Sky1: at the intersection of prion-like proteins and stress granule regulation. Curr Genet 66:463–468. https://doi.org/10.1007/s00294-019-01044-z
Shattuck JE, Paul KR, Cascarina SM, Ross ED (2019) The prion-like protein kinase Sky1 is required for efficient stress granule disassembly. Nat Commun 10:3614. https://doi.org/10.1038/s41467-019-11550-w
Siebel CW, Feng L, Guthrie C, Fu XD (1999) Conservation in budding yeast of a kinase specific for SR splicing factors. Proc Natl Acad Sci U S A 96:5440–5445. https://doi.org/10.1073/pnas.96.10.5440
Sridharan S, Kurzawa N, Werner T, Gunthner I, Helm D, Huber W, Bantscheff M, Savitski MM (2019) Proteome-wide solubility and thermal stability profiling reveals distinct regulatory roles for ATP. Nat Commun 10:1155. https://doi.org/10.1038/s41467-019-09107-y
Takaine M, Imamura H, Yoshida S (2022) High and stable ATP levels prevent aberrant intracellular protein aggregation in yeast. Elife. https://doi.org/10.7554/eLife.67659
Tauber D, Tauber G, Khong A, Van Treeck B, Pelletier J, Parker R (2020) Modulation of RNA condensation by the DEAD-Box protein eIF4A. Cell 180(411–426):e416. https://doi.org/10.1016/j.cell.2019.12.031
Tigano M, Ruotolo R, Dallabona C, Fontanesi F, Barrientos A, Donnini C, Ottonello S (2015) Elongator-dependent modification of cytoplasmic tRNALysUUU is required for mitochondrial function under stress conditions. Nucleic Acids Res 43:8368–8380. https://doi.org/10.1093/nar/gkv765
Wach A, Brachat A, Pohlmann R, Philippsen P (1994) New heterologous modules for classical or PCR-based gene disruptions in Saccharomyces cerevisiae. Yeast 10:1793–1808
Webb S, Hector RD, Kudla G, Granneman S (2014) PAR-CLIP data indicate that Nrd1-Nab3-dependent transcription termination regulates expression of hundreds of protein coding genes in yeast. Genome Biol 15:R8. https://doi.org/10.1186/gb-2014-15-1-r8
Weis K, Hondele M (2022) The role of DEAD-box ATPases in gene expression and the regulation of RNA-protein condensates. Annu Rev Biochem. https://doi.org/10.1146/annurev-biochem-032620-105429
Wilson WA, Hawley SA, Hardie DG (1996) Glucose repression/derepression in budding yeast: SNF1 protein kinase is activated by phosphorylation under derepressing conditions, and this correlates with a high AMP:ATP ratio. Curr Biol 6:1426–1434. https://doi.org/10.1016/s0960-9822(96)00747-6
Young MJ, Court DA (2008) Effects of the S288c genetic background and common auxotrophic markers on mitochondrial DNA function in Saccharomyces cerevisiae. Yeast 25:903–912. https://doi.org/10.1002/yea.1644
Acknowledgements
The authors acknowledge Drs. Jennifer Kwong, Homa Galei, Gerald Shadel, Joanna Goldberg and Judy Fridovich-Keil for helpful discussions, plate reader usage, or a critical reading of the manuscript. The technical expertise of Laura Fox-Goharioon is also appreciated. Additionally, the authors acknowledge Emory University’s Integrated Cellular Imaging Core (ICI) and Integrated Genomics Core.
Funding
This work was funded by National Institutes of Health (R01 GM120271 to D.R.), the Emory University School of Medicine, the Emory University Research Committee, and the Emory Integrated Cellular Imaging Core. The content is solely the responsibility of the authors and does not necessarily reflect the official views of the National Institute of Health.
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception and design. Material preparation, data collection and analysis were performed by KH, JH, and DR. The first draft of the manuscript was written by all authors and previous versions of the manuscript were commented upon by all authors. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no relevant financial or non-financial interests to disclose.
Ethical approval
No human subjects or vertebrate animals were used in this work.
Consent to participate
None.
Consent to publish
None.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
294_2022_1248_MOESM1_ESM.tiff
Supplementary file1 Fig. S1 A high granule accumulating yeast strain is ρ-. Whole genome sequencing analysis reveals that the high granule accumulating yeast strain DY4746 (parent to DY4756) is ρ- as it possesses only a small portion of its mitochondrial genome compared to its parental strain (DY4736). Sequencing reads in the lower two panels of the NCBI viewer are piled in horizontal lines with gray representing identities to the reference sequence and mismatches shown in red. All but ~15 kbp of mitochondrial DNA are lost from this petite strain (TIFF 6155 KB)
294_2022_1248_MOESM2_ESM.tiff
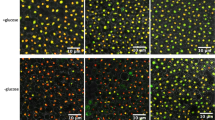
Supplementary file2 Fig. S2 Confocal microscopy of a glucose starved low granule accumulating strain treated with and without the electron transport uncoupler, FCCP. Yeast strain DY4772, which is low Nab3 granule accumulating, was grown to mid-logarithmic phase, washed into starvation media, starved for 2 hrs, treated with mock or 20µM FCCP for 30 min, imaged, and analyzed. DY4772 contains GFP-NAB3 (green) and HTB2-mCherry (red) markers (TIFF 6155 KB)
294_2022_1248_MOESM3_ESM.tiff
Supplementary file3 Fig. S3 Deletions of nuclear encoded genes result in a mitochondrial defect. A Yeast strains DY4772 (control), DY4851 (Δatp1) and DY4856 (Δatp3) were grown to saturation and diluted with sterile water to a concentration of 107 cells/mL. Cells were serially diluted 10-fold and 10µl of each dilution were spotted onto SC-glucose or SC-glycerol/ethanol as indicated and incubated at 30°C. B Yeast strains DY4772 (control) and DY4863 (low ∆sky1) were grown to saturation and diluted with sterile water to a concentration of 107 cells/mL. Cells were serially diluted 10-fold and spotted on SC-glucose or SC-glycerol/ethanol as indicated and incubated at 30°C (TIFF 6155 KB)
294_2022_1248_MOESM4_ESM.tiff
Supplementary file4 Fig. S4 Deletion of the HSP104, disaggregase does not affect Nab3 granule accumulation. Yeast strains DY4772 (control) and DY4862 (low ∆hsp104) were grown to mid-logarithmic phase, washed into starvation media, starved for 2 hours, imaged, and analyzed. Averages and standard deviations are plotted, and p values are presented. [n values for the bar graph (left to right) were, 11 and 3 respectively.] (TIFF 6155 KB)
Rights and permissions
Springer Nature or its licensor holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Hutchinson, K.M., Hunn, J.C. & Reines, D. Nab3 nuclear granule accumulation is driven by respiratory capacity. Curr Genet 68, 581–591 (2022). https://doi.org/10.1007/s00294-022-01248-w
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00294-022-01248-w