
Abstract
Jojoba (Simmondsia chinensis) is cultivated for its seeds, which contain a high-value liquid wax. There is little known regarding irrigation requirements of intensively cultivated jojoba. The project’s objectives were to evaluate the effects of irrigation regime on water status, growth, yield, and water productivity (WP, wax yield per unit of water applied) of intensely cultivated jojoba. An experiment was conducted over six years in a 14-year-old commercial plantation in Israel’s Northern Negev Desert. Treatments included: Control irrigation according to best commercial practice of returning reference evapotranspiration multiplied by a crop coefficient (Kc) of 0.5; Low irrigation providing 75% (Kc = 0.375) of the control; High irrigation providing 125% (Kc = 0.625) of the control; Regulated deficit irrigation (RDI) according to the control treatment, terminated for one to two months, after the wax accumulation stage. Stem water potential (SWP) consistently indicated jojoba water status. Increasing water application from − 25% to the control and further to + 25% led to augmented vegetative growth and number of seeds, but yield was increased at rates lower than the differences in water application by the treatments, such that WP was negatively associated. Effects on yield were limited to higher yielding ‘On’ years. The RDI treatment resulted in yield not different from the control and 15% higher WP, but significantly inhibited seed removal during mechanical harvesting. The results suggest commercial potential for increasing profits through water savings. Further research is needed to determine RDI regimes without detrimental effects on harvesting efficiency.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Jojoba (Simmondsia chinensis) is the sole species of the family Simmondsiaceae, which belongs to the order Caryophyllales. It is native to the arid regions of Sonora, Colorado, and Mojave deserts in the southwest US and northwest Mexico. A perennial, dioecious, evergreen desert shrub, jojoba is cultivated as an oil crop for the liquid wax, commonly called jojoba oil, in its seeds (Benzioni 2006; Sandouqa and Al-Hamamre 2019). Plant size can reach up to 2–3 m in height and slightly less in width. The shrub branching is prolific near its base, resulting in several stems rising from the root crown.
Leaf epidermal cells are thick and covered with hair and a prominent waxy coating, thus reducing water loss from evaporation and making the plant tolerant to dehydration of up to 35% of its leaf water (Benzioni and Dunstone 1986; Khattab et al. 2019). Jojoba uses the C3 pathway of photosynthesis and is apparently water-efficient in comparison to similar plants (Dunstone and Begg, 1979).
The fruit is a capsule, usually bears one seed, and contains mainly undifferentiated cotyledons tissue with little or no endosperm. Fruit ripens during the spring and the summer and is harvested in the autumn. Wax accumulation is rapid during the outset of fruit maturation, rising gradually in a sigmoid pattern. The capsule and the seed mature together until the seed is about 0.7–1.2 g, and the color changes from green to dark brown (Yermanos and Duncan 1976; Perry et al. 2021). Fifty to sixty days after pollination, the capsule reaches its full size, and seed dehydration begins (Benzioni 2006). Eventually, the seed coat dries out, enabling the seeds to fall to the ground with any light shake of the bush, as by animals or wind. In the ‘Hazerim’ cultivar, seed wax content was found to reach its maximum level in mid-June, followed by rapid dehydration of the seed (Perry et al. 2021). This suggests that application of deficient irrigation once dehydration begins would not significantly impact wax accumulation and production over time.
Data obtained in experimental plots near Kibbutz Hazerim in Israel’s northern Negev suggests that early seed development is associated with larger seed size and higher wax content. In early work in Israel (Benzioni et al. 1980) monthly irrigation of individual jojoba plants was found to enhance vegetative growth and induce bud dormancy breakage. Jojoba production exhibits biennial bearing with typical successive low and high yield seasons (Lazare et al. 2021). In high yield seasons, seeds cease growing earlier, resulting in lower wax content than the following year (Perry et al. 2021). A similar trend was observed in Arizona, where seeds that matured over five months were smaller and low in oil compared to seeds that went through a full seven months of maturation (Gentry 1958). Rapid maturation is associated with increasing evaporative demand and earlier seed dehydration, resulting in a reduction of final seed wax content (Benzioni et al. 1980).
Work in Israel’s Negev Desert and specifically in vicinity of Kibbutz Hatzerim/Beer Sheva has been important in the introduction, selection, and intensification of modern agricultural production of jojoba (Benzioni and Dunstone 1986; Benzioni 2006). Two of the most successful and therefore popular and important intensively grown cultivars in Israel today are “Hatzerim” and “Benzioni” (Agarwal et al. 2023). Since the 1960s jojoba cultivation and industry for wax extraction and marketing have been developed in the region (Benzioni 1995, 2006). Work includes investigation of all aspects of cultivation and development of mechanization for pruning and harvesting, leading to a culture with low dependence on labor. Today, about 40% of global jojoba oil production is in Israel, and yields are expected to grow as young plantations, planted between 2010 and 2020, reach maturity and full production. Despite the successful domestication and intensification over the last decades and despite its high profitability, answers to questions regarding preferable irrigation practices under modern intensive cultivation remain open. Current irrigation regimes include use of subsurface drip systems, which allow seed collection from the soil surface, and common irrigation protocols based on return of reference evapotranspiration (ET0) with a crop factor (Kc) of ~ 0.5. This crop factor is loosely based on research in the early 1980’s by Benzioni and Nerd (Benzioni et al. 1980; Benzioni and Nerd 1985), who found that annual water requirements for an adult jojoba plantation then, with smaller plants of different varieties and architecture due to pruning than today, was 500–600 mm. In the years since, the Kc has been adjusted according to grower experience.
Sensor-based irrigation scheduling is becoming popular for many crops, including fruit trees. Water status or stress can be monitored by sensors in the soil or in the plant. Of such sensors, dendrometers, measuring shrinking and swelling of trunks or branches, have been shown to be sensitive to water status and stress and threshold values for indices generated from dendrometers are used commercially to guide irrigation decision-making in tree crops including avocado and olive (Fernández 2017).
The main objective of this study was to evaluate jojoba irrigation requirements by measuring and comparing water potential, growth, and yield under different irrigation regimes. Since irrigation amounts for optimal production have yet to be determined, evaluating the plant response to different irrigation regimes can help optimize field water management. Three hypotheses were specifically tested in the study:
-
1.
Higher irrigation increases yield by promoting flowering and fruit set and delaying seed dehydration, allowing a longer period of maturation and heavier seeds with more wax.
-
2.
Regulated deficit irrigation (RDI) as an approach in water-limited areas would increase water productivity (WP; wax yield as a function of applied irrigation), and that inducing stress in jojoba during fruit dehydration would not significantly damage wax yield over time.
-
3.
Dendrometer-generated trunk circumference data could provide threshold values to guide irrigation scheduling, and lead to improved production and WP.
Materials and methods
Experimental setup
The experiment was conducted from 2016–2021 in a 14-year-old commercial cultivated jojoba plantation next to Kibbutz Hazerim in the Northern Negev in Israel (31°14’08.2"N 34°43’45.0"E). Average root zone soil texture was loam, consisting of 36% sand, 46% silt, and 18% clay. The soil’s water saturation percentage and the CaCO3 content were 37% and 19%, respectively. The plantation was planted with the cultivar ‘Hazerim’ in straight rows 4.5 m apart with 2 m between plants in a row, producing a density of 1100 plants/ha. Every fifth row had a male plant every five plants as a pollen source, giving a density of 2.5–5% male plants for the entire plantation. Plants were pruned every year, after harvest around November, to enable agricultural machine locomotion and mechanical harvesting, removing the lower part of the plant and its sides. The experiment was conducted in a 0.25 ha portion of the plantation. The field was designed to test different irrigation treatments in a randomized block design, with five plots per block, producing five replicated plots per treatment. Each plot contained three rows, each with nine plants, and only the middle five plants of the middle row were measured.
Each irrigation treatment had its own pump and water meter. Irrigation frequency was once a week on Sundays from November to March and twice a week, on Sundays and Wednesdays during the rest of the year. The irrigation was with freshwater (EC = 0.31 dS/m) via a subsurface drip system, with two laterals buried 40 cm deep in every other row. Drippers were inline “Ram” pressure compensated 1.6 l/h spaced every 0.7 m (Netafim, Hazerim, Israel). Daily ET0 (modified Penman-Monteith, Allen et al., 1998) values were obtained from the Gilat-24 station of the Meteorological Service of Israel’s Ministry of Agriculture and Rural Development (www.meteo.co.il), located 11 km from the experimental field.
Fertilizer was applied continuously and proportionally between March and November via the irrigation system (fertigation), respective to the expected irrigation amount, determined based on the 10-year evaporation daily average from the data from the Israel Meteorological Service. Total seasonal fertilizer applied was equal for all the irrigation treatments. Seasonal fertilizer application was 250 kg/ha N, 100 kg/ha P and 300 kg/ha K. Fe, Mn, Zn, Cu, and Mo were given with the fertilizer as chelates (3%) with a concentration of 0.3, 0.15, 0.07, 0.01, and 0.008 g/L, respectively.
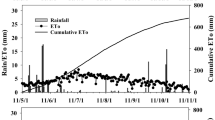
The site is characterized by a semi-arid Mediterranean climate receiving average annual precipitation of about 200 mm. Almost all rainfall falls during a small number of events between November-February. Annual precipitation varied during the experiment with 159, 281, 113, 304, and 97 mm in 2016–2017, 2017–2018, 2018–2019, 2019–2020, and 2020–2021 winter seasons, respectively (Fig. 1). Average daily maximum and minimum temperatures were approximately similar in all years and were ~ 28.1 and ~ 14.3 ℃ and daily reference normal ET ranged from ~ 1.5 mm in the winter to ~ 7.5 mm in the summer (Fig. 1).

Daily reference ET (ET0) and rainfall over the course of the experiment. Data from the Gilat-24 station of the Ministry of Agriculture Meteorological Service (meteo.co.il/)
Irrigation treatments
The control treatment (control) was established according to the common commercial practice of returning the previous week’s mean calculated ET0 with a Kc of 0.5 and a cover factor of 0.85%. Other treatments were as follows: low-irrigation amount was set at 75% of the control treatment using Kc of 0.375; high - irrigation amount was set at 125% of the control treatment using Kc of 0.625; and RDI- irrigation schedule was set according to the control treatment with a cessation of irrigation during the last stage of fruit maturation after wax accumulation ceased. The 2017 and 2018 water deficit periods were from June 18th to September 3rd and from June 27th -August 1st, respectively. After 2019, the periods of irrigation halt were restricted to one month during July. Following resumption of irrigation, an irrigation regime equal to the control (Kc = 0.5) was re-established. Due to the irrigation cessation, the overall fertilization dose of the RDI treatment was divided into fewer fertilization events. Therefore, N amounts in every irrigation event were slightly higher than other treatments to achieve an equivalent annual dose. A fifth treatment, which was largely unsuccessful and therefore its results presented briefly separately from the others, tested an irrigation strategy based on dendrometer data and associated threshold values.
Patterns of accumulated irrigation are shown in Fig. 2 and total annual water applied given in Table 1. Average annual application of all six years of the experiment for the 100% treatment was 664 mm.

Accumulated annual irrigation per treatment. In 2020, seasonal irrigation stopped on 29 November following theft and vandalism of the automation system and irrigation head equipment. In 2021, recording of irrigation ceased with the end of the experiment on the 10th of October
Physiological response
Stem water potential
Stem water potential (SWP) using a Scholander-type pressure chamber-ARIMAD 3000, manufactured by MRC lab, Israel was evaluated midday (11–12 A.M.). Two western branches from each plot were covered with an aluminum bag for at least one hour before they were removed and immediately transferred to the chamber. In 2017–2019 measurements were done on three dates: May, July, and August. In 2020 and 2021, measurement frequency increased to represent the plant’s response more accurately during the growth season. All measurements took place on Tuesdays, two days after the last irrigation (Sunday).
Photosynthesis
Diurnal curves of CO2 assimilation rate were determined on two days, 19 July and 04 August, in 2020. Assimilation rates of sunlit diagnostic (youngest fully mature) leaves from the 75%, 100%, 125%, and RDI treatments in the same block (one with the most distinctive SWP differences) were determined five times between 08:00 and 17:00 on 19 July and six times between 07:00 and 17:00 along the day on 04 August. Measurements were done using a portable photosynthesis system- CIRAS 3 (PP-Systems, U.K) under ambient light, at 400 ppm CO2 and temperature and relative humidity set to ambient.
Vegetative growth
Leaf area index
The leaf area index (LAI) was measured each season using Accupar LP-80, produced by Decagon Devices, following the manufacturer’s instructions. The analyzer directly and automatically calculates the LAI value with an array of 80 PAR (photosynthetically active radiation) sensors installed on an 0.8 m long linear probe that measures how much light is intercepted by the canopy. Five chest height measurements per plot were taken in August 2017–2018 and four in the 2020 and 2021 seasons. In the 2021 season, LAI was measured monthly during vegetative growth from February to August.
Pruned branches
The weight of the pruned material was determined annually (end of November) after commercial, mechanized pruning was done uniformly in all experiment plots in the 2018–2020 seasons. Branches and leaves were collected from the ground and weighed to evaluate and quantify seasonal vegetative growth per plot.
Reproductive development
Fruit set
To determine the fruit set rate, branches from each plot were marked before flowering (December-January), and the number of flower buds in each branch was recorded. In May, after pollination and fruit set, branches were further surveyed, and the number of fruit per branch was counted. Fruit set (%) was calculated as number of fruit per number of flowers on each branch. Nine, four, and ten branches per plot were marked for seasons 2017–2018, 2019, and 2020–2021, respectively.
Yield
Harvesting was done after the seeds fully matured, and the plants were shaken by commercial mechanical means to release attached fruit to the ground. In 2017 and 2019, a second harvest was done following observation of many fruit that remained on the plants after the first harvest. Fruit were collected in boxes, then separated from leaves and litter using a conveyor belt and a blower. The yield was calculated per plant by dividing the yield per plot by the number of measured plants in each.
A random sample of 100 fruit was taken from each plot. Seeds were counted and weighed for average seed mass.
Wax content
A random 1 kg sample of seeds was taken from each plot and was dried in an oven at 70℃ for three days. The dried sample was ground, and a subsample weighed and transferred to a paper tube capped with cotton wool to determine the wax content using a Soxhlet extractor apparatus heated to 70℃ and filled with n-hexane for 6 h. The tube was oven-dried and weighed again with the weight difference after the wax extraction represented the wax content.
Wax yield per hectare was calculated by first calculating wax yield per plant (seed mass X average wax percent) and then multiplying by number of plants per hectare (1100).
Trunk/branch size fluctuations
An additional set of five replicate plots was set aside for monitoring by dendrometers with the goal of developing thresholds for irrigation scheduling (sensor-based irrigation, SBI). Dendrometers (Phytech, Israel) were installed on main branches/trunks and monitored in 2017 and 2018 to develop a plant-based irrigation regime, which was applied in 2019. The plants were irrigated according to the control treatment in 2017 and 2018 and their average maximum daily shrinkage (MDS) was used to detect irregular values compared to desirable values, determined in the preceding years. Irrigation in 2019 was halted in March with the intention of re-applying irrigation once plants reached this threshold (MDS = 200 μm). After nearly two months, when the threshold was still not reached and following measurement of very low SWP values, the decision to stop the SBI treatment was made, leading to a single year of early season drought stress.
Statistical analysis
Average and standard deviation calculations were performed with JMP statistical software (JMP pro-15.0.0). Treatments were initially compared by repeated measures MANOVA and evaluated using the Pillai’s Trace test. When interactions were found between time (year) and treatments, treatments in each year were evaluated by ANOVA and comparisons between groups were done using the Tukey-HSD test. Data presented as percentages (fruit set and wax content) underwent arcsin root transformation prior to analysis. All tests had p-value for significance set to 0.05.
Results
Physiological measurements
Stem water potential
Repeated measures ANOVA showed significant interaction between time and irrigation treatment as well as significant influence of both treatment and time. Stem water potential measured on three dates in each of the years 2017–2019, showed consistent significant differences between the low (Kc 0.375) and high (Kc 0.625) irrigation treatments (Fig. 3). Differences between treatments reflected the amount of water applied in each treatment, and seasonal changes reflected the increased evaporative demand in the summer months. Each year, the values measured in summer averaged − 4.9, − 4.7, and − 4.4 MPa for the low, control, and high treatments, respectively. Irrigation cessation in the RDI treatment lowered the SWP, with a mean of − 5.9, − 5.3, and − 5.7 MPa in the latter measurements in 2017, 2018, and 2019, respectively. Those values were significantly lower compared to those of the other irrigation treatments in all years.
In the 2020 and 2021 seasons, SWP measurements were taken more frequently and reflected the dynamics of the treatment application (Fig. 4). All treatments decreased to a similar level in both seasons. In 2020, treatments low, control, and high had maximum values of − 2.4, − 2.3, and − 2.2 MPa (low and high were significantly different), respectively, in late March. Treatment-related differences developed over time, dropping to a minimum of − 4.7, − 4.4, and − 4.1 MPa in mid-August for the low, control, and high treatments, respectively, where the high irrigation level was significantly different from the others. No significant difference was visible between the low and control treatments in all measurements, while a significant difference between the high and control treatments started to develop in mid-July and continued throughout the season. A steep drop in SWP value of 0.29 MPa/week occurred once irrigation stopped in July in the RDI treatment, reaching a minimum of − 5.1 MPa on 28 July. Resumed irrigation resulted in a recovery of the plants’ water status to a level similar to the control on 16 August. A month later, SWP rose to − 4.2 MPa, higher than the control treatment (− 4.4 MPa), but not significantly different.

The effect of irrigation treatments on stem water potential (SWP) on three dates during seasons 2017–2019. Values are means of five plots per treatment. RDI is regulated deficit irrigation. Different letters represent a significant difference (p < 0.05, Tukey -HSD) between treatments for each date separately
On approximately the same date as in 2020, the first measurement in 2021 was 0.79 MPa, lower in average for all treatments (Fig. 4b). A significant difference developed between the high irrigation treatment with − 3.3 MPa and the control with − 3.8 MPa in late May, and unlike 2020, between the low treatment (− 4.3 MPa) and the control (− 4.6 MPa) in mid-July. Both differences lasted until the last measurement in late August. RDI (irrigation halt in July) resembled the control and low treatments, until the effect of the stoppage of irrigation caused a significant difference compared to other treatments. RDI treatment SWP values dropped to a minimum of − 5.4 MPa in late July. Ten days after irrigation resumed, the RDI treatment reached a statistically similar SWP to the low irrigation level (− 4.4 MPa).

The effect of irrigation treatments on stem water potential (SWP) in 2020 (a), and 2021 (b). Values are means of five plots per treatment. Apart from 16 March 2021, significant differences (p < 0.05, Tukey-HSD) between treatments were observed on all other dates
CO2 assimilation
The assimilation rate was related to the irrigation treatments (Fig. 5). In Fig. 5 diurnal results for two days in 2020, the 19th of July and the 04th of August are presented. The meteorological conditions on the two days were similar. On the 19th of July, minimum and maximum temperatures were 19.4 and 35.3 °C, minimum relative humidity was 28.3%, maximum daily radiation was 962 W/m2 and reference evapotranspiration was 6.79 mm. On the 20th of August, minimum and maximum temperatures were 21.9 and 34.9 °C, minimum relative humidity was 37%, maximum daily radiation was 995 W/m2 and reference evapotranspiration was 6.59 mm. Assuming that fruit maturation was complete by mid-July and therefore carbon demand was equal between the days, the explanation for differences between the days is that 19 July (Fig. 5a) was a single day after an irrigation event while 04 August (Fig. 5b) was two days after irrigation. On the first day (Fig. 5a), the RDI treatment showed a statistically significant difference compared to other irrigation regimes in the morning and again in the afternoon. By 14:45 there were no longer significant differences. While the control treatment had its highest assimilation rate at 12:30, the photosynthesis rates of the low and high irrigation treatments peaked earlier, at 10:15. The plants’ assimilation rates decreased in all treatments on the second measurement day (Fig. 5b). On August 4 2020, at 11:00, only the high treatment had significantly greater CO2 assimilation compared to the RDI treatment. At 15:00, after a steady decline in the low irrigation treatment, three separated significance groups were visible (RDI, low and control, and high).

The effect of irrigation treatments on assimilation rate. Measurements were taken under ambient light on 19 July 2020 (a) and on 04 Aug 2020 (b). RDI plots were not irrigated during July. Values are means of five and seven measurements per treatment for a and b, respectively. Different letters represent a significant difference (p < 0.05, Tukey-HSD) between treatments for each hour separately
Vegetative growth
LAI
The LAI measured each year in August was correlated to the amount of water applied in the irrigation treatments (Fig. 6), with greater amount of applied irrigation water leading to higher LAI values. There was no significant interaction between time (year) and treatment regarding LAI while both year and treatment influenced overall LAI significantly. Generally, the treatments were found to be divided as High (A), Control (AB), Low (C) and RDI (BC).

The effect of irrigation treatments on leaf area index (LAI) measured in August every year. Measurement was not taken in 2019. Values are means of five plots per treatment with each plot value represent an average of five readings for 2017 and 2018, and an average of four readings for 2020 and 2021. Repeated measures ANOVA resulted in division between High (A), Control (AB), Low (C), and RDI (BC) with different upper case letters signifying significant differences (p < 0.05). Different letters in the figure represent significant differences (p < 0.05, Tukey-HSD) between treatments for each year separately
Pruned branches
The mass of the pruned branches measured annually starting in 2018 (Fig. 7) showed a consistent increase every year except 2021 in all treatments. Repeated measures ANOVA indicated significant influence of year but not treatment and no interaction between treatment and time. Despite a significant difference between the low and high treatments in 2018 and 2019, branches mass in 2020 was almost identical in all treatments. Results for the RDI treatment resembled the control treatment. The total four-year average for the low, control and high treatments were generally related to their irrigation application amounts, average pruned branches was 6.7 kg/plant for the low treatment, 8.9 kg/plant for the control, and 10.0 kg/plant for the high irrigation treatment.

The effect of irrigation treatments on pruned branches weight. Weighing took place after pruning in November. Values are means of five plots per treatment. Different letters represent a significant difference (p < 0.05, Tukey-HSD) between treatments within each year
Reproductive development
Fruit set, seed weight and wax content
Fruit set was significantly influenced by year but not by irrigation treatment and there was no interaction found between time and treatment. Apart from 2017, when fruit set level of the low irrigation treatment was lower (67.1%) compared to the control (80.6%) and high (81.8%) treatments, no significant differences were found between treatments (Table 2). The total average percent fruit set for all treatments in 2017–2021 was 80.2%.
Seed mass (Table 2) was significantly affected by season, but not by treatment and time and treatment showed positive interaction. The only significant effect of treatments on seed mass was for the RDI treatment in 2018 with average mass per seed of 0.99 g compared to 1.1 g for all other treatments. Seed mass was inversley related to number of fruit per plant. In lower yielding years 2016, 2018, 2020 average seed weight was 1.10, 1.09, and 1.25 g, respectively while in the higher yielding years 2017, 2019 and 2021, corresponding seed weight was 0.88, 0.93 and 1.00 g.
Irrigation treatment was additionally found to be an insignificant factor influencing wax content in the dry seeds (Table 2). Time (year) affected wax content while treatment did not, with no interaction found between time and treatment. The average dry seed wax content for all treatments from 2016 to 2020 was 50.5%, with variations between years that were not related to bearing level.
Yield
Seed was harvested in the high yielding seasons 2017 and 2019 in two stages because many remained on the plants after the first commercial harvest. The second harvest of the RDI treatment represented a significantly greater portion of the total yield in both 2017 and 2019 compared to the other treatments (Fig. 8). Biennial bearing behavior was evident with relatively low yields in 2016, 2018 and 2020, each followed by relatively high yields in 2017, 2019 and 2021. Yield was significantly influenced by both year and irrigation treatment with significant interaction between them. In the ‘On ' years, plants produced ~ 2.9 times more fruit on average for all treatments, compared to the ‘Off’ years. Yield in the high treatment was significantly greater in 2017 and 2021 compared to the low and RDI treatments. In 2017 the control treatment was also had significantly higher yield than the low and RDI treatments. In 2021 the three water amount treatments were significantly different from one another with the low treatment having 19% less and the high treatment 14% more yield than the control treatment. Total six year accumulated seed and wax yields were related to water application, the low irrigation treatment reached 26.9 kg seed/plant (15.0 ton wax/ha), the control treatment 27.8 kg seed/plant (15.5 ton wax/ha), and the high treatment 30.63 kg seed/plant (17.1 ton wax/ha). The accumulated yield for the RDI treatment was 26.7 kg seed/plant (14.5 ton wax/ha).
Water productivity
Water productivity (WP) was calculated as kg jojoba wax per m3 of water irrigated (Fig. 9). Like yield, overall WP was significantly influenced by both irrigation treatment and time, with significant interaction found between them. Greater WP was consistently observed in the low irrigation treatment compared to the control and high treatments, where the decrease in yield was lower than the decrease in irrigation. Treatment-related differences in productivity were observed in high yield years (2017, 2019, and 2021) and significantly increased in all treatments compared to low yield years. One exception was 2018, when the RDI treatment showed significantly higher WP compared to the high treatment. Irrigation treatments in 2017 resulted in four significantly different groups, with RDI showing the highest WP values (0.74 kg wax/m3 irrigation water), while in 2019, only the low treatment was significantly different (0.72 kg wax/m3 water). Total six-year accumulated WP ranged from 0.33 for the high irrigation treatment and 0.38 for the control treatment to a significantly higher 0.47 kg wax/m3 irrigation water in the low irrigation treatment.

Seed yield per plant for each treatment in each year. Columns are averages of five replicate plots. Black horizontal lines in 2017 and 2019 differentiate between seeds harvested with commercial equipment (below) and seeds remaining on the plants and harvested afterwards manually (above). Different letters represent a significant difference (p < 0.05, Tukey-HSD) between treatments for each year separately

Water productivity (WP) per year and accumulate total in the irrigation experiment for all treatments. Calculated as kg wax per cubic meter irrigation water. Values are means of five plots per treatment. Different letters represent significant differences (p < 0.05, Tukey-HSD) between treatments within each year
Sensor-based irrigation
Maximum daily trunk shrinkage values determined for each treatment based on dendrometers fluctuated in response to the biweekly irrigation events (Fig. 10). In general, the low irrigation treatment showed greater amplitude than the high treatment. The oscillation trend disappeared in the RDI plots once irrigation ceased and returned when irrigation resumed.
The attempt to irrigate according to MDS threshold using the dendrometers (SBI treatment) led to low application amounts of irrigation early in the season, from April until mid-May, of 2019. On 12 May 2019 the SBI treatment had SWP of − 4.6 MPa (Table 3), significantly lower than that of the control treatment (− 4.0 MPa) or the low treatment (− 4.1). In light of this, the treatment was stopped and irrigation returned to irrigation as the control. On 02 July the SWP of the SBI treatment was still lower than the other treatments, − 5.0 MPa compared to − 4.4 MPa (control) and − 4.6 (low). Compared to the control treatment, the SBI irrigation regime in 2019 had no significant effect on growth but did influence two yield components. The SBI plants witnessing the early season stress produced lighter seeds and accumulated less wax (Table 3).

Maximum daily shrinkage (MDS) from June to August in 2019. Values are mean of two or three sensors per treatment. Average (min – max) standard deviations for the entire period shown were 6.2 (0–20), 22.0 (3–50), 7.2 (0–19), and 27.5 (0–56) µm for the Low, Control, High, and RDI treatments, respectively
Discussion
Physiological measurements
Stem water potential and assimilation rate reflected the given irrigation amounts, and both showed the same trend of lower values with less irrigation and in response to increasing evaporative demand (Figs. 3, 4 and 5). The difference in SWP between the three levels of irrigation (low, control, and high) remained roughly constant, while seasonal SWP followed a pattern of high (less negative) values in spring decreasing during fruit maturation toward the end of the summer. A similar seasonal trend of decreasing SWP was reported in olives (Ben-Gal et al. 2011). Values of SWP fluctuated between years, however, they did not follow the alternate bearing cycles and were probably responding to meteorological conditions in each year. Compared to other crops, jojoba plants reach very low SWP values, drawing moisture from very dry soils. This is probably made possible by high resistance to embolism (Jones and Sutherland 1991). In addition, jojoba’s leaf arrangement with perpendicular orientation decreases radiation interception at noon when ET is high and allows maximum light penetration when ET is low and water potential is high (Wardlaw et al. 1983).
As expected, the RDI treatment had a strong effect on SWP in all years. The prolonged period of irrigation cessation in 2017 resulted in the lowest value measured of − 5.9 MPa at the end of August. In 2019, the same effect was observed, with the RDI treatment having its lowest values at the end of July. Midday SWP for detecting water stress in jojoba was suggested previously by Benzioni and Nerd (1985) to be between − 4.0 to − 4.2 MPa on 12 year old “Schneider” variety plants. This reported range may be considered skeptically, since, in our experiment, SWP was often lower, even in well-watered plants. Minimum threshold SWP values for irrigation should address the timing of the measurements in regards to seasonal dynamics since the maximum difference in values along the year reached 2.1 and 1.2 MPa in the control treatment in 2020, and 2021, respectively. In 2020 and 2021, plants under RDI regimes showed a fast recovery once regular irrigation resumed, even to a level higher than the control in 2020 (Fig. 4a), which may suggest a positive effect of the RDI treatment on plant water status in the long term. Similarly, moderate deficit irrigation during pit hardening in olives has been reported to improve water savings (Fernández et al., 2013). In 2021, the recovery of SWP following the RDI period could also be attributed to environmental factors affecting the ET demand as the SWP in all treatments rose in the measurement after irrigation resumed. According to the results in the current experiment, local thresholds of SWP for guiding irrigation would range from − 3 MPa in April and May to − 4 MPa in June and August-September and reach as low as − 4.5 in July (Figs, 3, 4).
Irrigation increased the photosynthesis rate and extended the time of maximum CO2 assimilation, as significant differences between the low and control treatments occurred only at 15:00 (Fig. 5b). The ability of jojoba plants to photosynthesize at very low levels of leaf water potential compared to other mesophytic plants has been reported in other studies (Benzioni and Dunstone 1986; Adams et al. 1978; Glat et al. 1983). Our results demonstrate that in jojoba, like other plants, water availability affects stomatal closure, which intensifies under water stress, decreasing the CO2 assimilation rate (Downton et al. 1988; Cochard et al. 2002). The assimilation rate was strongly affected by, in addition to environmental conditions, the immediate water availability. This is evident from comparison of diurnal measurement from two days, the first, a single day following an irrigation event, and the second, two days after irrigation (Fig. 5). The additional day resulted in approximately half of the previously measured assimilation rate.
Vegetative growth
The LAI and pruning material results confirm the positive effect of irrigation on vegetative growth (Figs. 6 and 7), although reductions in LAI could be due to enhanced rate or extent of leaf abscission . These findings are supported by previous studies of enhanced growth in well-irrigated jojoba plants (Benzioni and Nerd 1985; Nelson et al. 1993). Bearing levels and patterns likely influence vegetative growth due to restrictive and local effect of fruit development on vegetative growth because of competition for resources between neighboring branches, as reported previously in jojoba (Perry et al. 2021) and olives (Dag et al. 2010). While there is little data concerning jojoba’s biomass accumulation in commercial plantations reported in the literature, the sensitivity of cell elongation (growth) to water stress is well known in other crops (Nonami 1998). Like us, Ehrler et al. (1978) reported a significant increase in canopy volume in jojoba with higher water availability using water catchments after four years of treatments. The evaluated growth parameters were not affected by the RDI treatments, except the LAI results in August 2017. This confirms that halting irrigation for two months was too long and restricted growth. Nevertheless, and according to the SWP increase once irrigation resumed (Fig. 4), water-stressed plants seem to recover well, and the negative effect on growth was not present in the following years when irrigation discontinuation was reduced to one month. Fast recovery of physiological processes following water stress was reported previously for olives (Ben-Gal et al. 2010), a crop sharing jojoba’s evolutionary adaptation to harsh dry climate with prolonged drought conditions and intermittent or rare precipitation.
Reproductive growth
We hypothesized that higher irrigation amounts would delay seed dehydration, thus allowing a longer period of maturation, which is associated with heavier seeds. Our results did not support this, as seed mass was not affected by increasing irrigation. Higher irrigation levels were reported to produce heavier seeds in a previous study (Ayerza 1993). In addition to being maybe being cultivar specific, a possible explanation is the difference between the evaluated irrigation treatments in Ayerza (1993), which were significantly higher (0, 300, 600, and 900 mm ha− 1 year− 1) compared to ours.
Seed mass changed according to the alternate bearing, with lighter seeds in high-yield years (Table 2). A significantly lower mass was recorded in 2018 in the RDI treatment, suggesting that the long period of fruit maturation reported by Perry et al. (2021) in our experimental plots may influence the proper timing of deficit irrigation.
Fruit set was expected to benefit from higher water availability. This hypothesis was based on the findings of Benzioni and Nerd (1985), who found that irrigation promoted flowering and fruit set. In our experiment, no effect on fruit set (Table 2) or flower production (data not shown) was observed, similar to findings in a study conducted in Saudi Arabia on 5 year old jojoba plants (cv Schneider) exhibiting drought conditions instigated by irrigation intervals ranging between 1 and 36 weeks (Osman and AboHassan 2002).
In a study on two olive cultivars, oil content tended to increase due to decreased irrigation (Ben-Gal et al. 2021). In contrast, wax content as a percentage of seed dry weight in jojoba seems to be independent of irrigation regime (Table 2). Other findings suggest this trait is mainly genetic (Purcell et al. 2000; Benzioni and Vaknin 2002). In the present study, wax content was apparently linked to environmental conditions, which varied between years.
The RDI irrigation cut-off date was planned according to the onset of the dehydration process, indicating maximum wax accumulation in the seeds, which we assumed to occur in July. In 2017 and 2019 (‘On’ years), this led to inhibition of seed drop and the need for a second harvest, which was significant in the RDI treatment with 32%, and 22% of total yield, respectively (Fig. 8). This phenomenon has substantial economic implications due to the high cost of each jojoba harvest. Other crops including oranges have been reported to have an opposite response, with fruit detachment force increased as a function of water stress (Morgan et al. 2014).
A maximum annual yield of 5.6 ton seeds/ha (2.85 ton wax/ha) for a six-year average was found for the high irrigation treatment. This with biennial bearing ranging from ~ 1 to 4.9 ton/ha wax. A significant advantage on yield from higher water availability was observed only in ‘On’ years (Fig. 8). This is likely due to increased vegetative growth in “Off” years providing opportunity for more flowers and seeds per plant. Treatments did not significantly affect the 2016–2021 total accumulated yields, although mean values corresponded to the water amounts applied. This suggests that adequate irrigation is more crucial under high fruit load and that the low irrigation regime in jojoba may be implementable over the long-term without significant profit loss. This is also true for the RDI treatment, albeit with the caveat of the issue of a large percentage of seeds failing to drop. Similar results where irrigation treatments affected yield in “On” and not “Off” years were found for olives (Ben-Gal et al.,2011).
Yield is a function of several components. Higher water application showed an advantage in growth, which affected the number of reproductive buds, but no significant effects on seed size, fruit set, or wax concentration were visible, resulting in only mild differences in accumulated yields between the treatments. The expected increase in yield due to increase of number of fruit on plants with higher vegetative growth rates is confined in modern intensive jojoba cultivation due to pruning protocols that remove much of the vegetation each season (Lazare et al. 2021). Hence, depending on local economics, the water saved by reducing the common commercial irrigation practice by 25% may be more valuable to growers than the associated damage to crop yield. Due to the risk of fruit remaining on the plants, we conclude that our approach to the RDI regime in jojoba was not optimal or practical. Yield load affects the duration of wax accumulation and treatment response; thus, we suggest that a more successful RDI regime would consider the wax levels each year in the timing of the water stress period. Potentially, to optimize yield, WP, and profits, jojoba could be irrigated with a Kc of 0.375 (like the low treatment) and reduced in a moderate RDI after wax accumulation and before seed fall to 0.25. But this remains to be tested. Another suggestion is to further investigate the relations between seasonal SWP and fruit load in order to establish an RDI regime based on actual plant water status.
The influence of alternate bearing on yield was evident (Fig. 8). RDI and the low treatment resulted in the highest restraining effect on the yield difference between two consecutive years (Table 2; Fig. 8), but as mentioned, this did not increase yields. Therefore, reduced severity of the alternate bearing phenomena could possibly be related to lower relative yields and not to improved balance between vegetative and reproductive growth.
Water productivity
Regulated deficit irrigation, with lower than optimal amounts of water provided at physiological plant phenological stages found insensitive to water stress, commonly increases WP in fruit tree crops (Kumar et al. 2023). In the current study, WP (Fig. 9) showed a trend of higher values as a function of reduced irrigation, but despite RDI applied at the fruit dehydration stage of jojoba being proven not to reduce yields significantly, its WP was not the highest. The highest WP was obtained in the low irrigation treatment, with a total average for all years of 0.47 kg wax/m3 of irrigated water. The fact that the current experiment utilized good quality water for irrigation is important to the findings regarding WP. In Israel, irrigation of jojoba with water containing high concentrations of salts is common (Benzioni et al. 1996). In such cases, irrigation regimes must include additional water for salt leaching (Ben-Gal et al. 2017).
Sensor based irrigation regime in 2019
The immediate effect of irrigation on plant water status was reflected in the MDS values obtained from the dendrometers (Fig. 10). Treatment-related differences were not apparent, although a general pattern of smaller MDS fluctuations with higher amounts of irrigation were identified. Irrigation cessation in the RDI treatment was well recorded by the sensors, causing a moderate decrease with time in the MDS values. This proves the potential of the daily shrinkage and swelling of the trunk to represent the jojoba plant water status, as was reported in other crops (Fernández and Cuevas 2010). Nonetheless, dendrometer-driven data should be treated carefully as an indicator for water stress in jojoba, as was reported previously in olives (Moriana et al. 2012b). The SBI regime applied in 2019 resulted in prolonged and low SWP, first detected in mid-May. Until late March, the irrigation rate in the control and SBI plots was almost similar. Irrigation ceased in the SBI plots on 31 May 2019 according to threshold MDS values determined based on previous years and other orchards, and resumed to be as the control only in mid-May, once poor water status was detected by the relatively low SWP values that were measured. Because of this, the SBI and the RDI treatments in 2019 were comparable, and any difference in plant performance could be linked to the time in which the irrigation halt period was initiated. Seeds under SBI were smaller and with less wax compared to the control (Table 2). This implies that more than actual values, the time in which low SWP occurred is crucial to jojoba response to water deficit and that wax accumulation processes occurring between March-May are sensitive to water stress.
The chosen MDS values from dendrometer data for scheduling irrigation apparently represented severe water stress in jojoba plants in the year they were tested and were obviously not appropriate. Further studies on jojoba’s trunk diameter variations in response to irrigation are needed for correct calibration of the dendrometers data to actual water status. An additional limitation to the method lies in the large variability found between replicate sensors on the same treatment. Apart from natural variations arising from field experiments, the non-symmetrical growth and multi-trunked structure of jojoba add challenges for correct sensor placement and data interpretation.
Characterizing jojoba water demand
Jojoba is a well-adapted plant for surviving in water-limited areas. Desert-adapted plants have a limited assimilation rate and reduced transpiration (Dunstone and Begg, 1979), restricting possible benefits from higher water availability. Genotypes adapted to unfavorable conditions may have superior performance in restrictive environments but be unable to take advantage of favorable conditions (Lortie and Aarssen 1996). Nevertheless, jojoba plants showed improved vegetative growth in response to higher water availability, which may not be reflected in yield because of the annual pruning. SWP was highly sensitive to the irrigation rate, suggesting that although wild stands have been reported to live in areas with very little rainfall (Gentry 1958), plants can uptake, utilize, and benefit from higher amounts of water. Nevertheless, despite an increase in the canopy density (Fig. 6), and vegetative growth (Fig. 7), no significant advantages in any of the yield components were observed with a higher application of water. Therefore, we conclude that, under the conditions of the current experiment, reducing the common irrigation amounts by 25%, and therefore basing irrigation on Kc of 0.375, is possible without significant yield loss. This would further allow benefit from the relatively high WP found in the low irrigation treatment (Fig. 9). The period of fruit maturation is crucial for maintaining proper water status. Inflicting water stress in the onset and during fruit maturation was found to have negative implications on productivity, as opposed to our RDI strategy of drought stress during July, which showed no significant yield reduction. More research is required to determine RDI timing and rates that do not reduce rates of seed drop from the plants.
Conclusions
The three driving hypotheses of this study were each found incorrect. First, higher rates of irrigation, while found to improve plant water status and increase vegetative growth, did not contribute to increased wax production via promotion of flowering, fruit set or delayed seed hydration. Second, regulated deficit irrigation with drought stress applied by stopping irrigation during the period following seed maturation and wax accumulation before harvest, did increase WP as hypothesized but, due to it causing reduced efficiency of mechanical harvesting and fruit that remained on the plants, the approach was not optimal or practical. Third, dendrometer-generated trunk size data was not reliable for generating threshold values to guide irrigation scheduling.
Midday stem water potential was a sensitive indicator of water status for jojoba plants under intensive commercial cultivation. Lower SWP as a function of reduced irrigation supply led to less photosynthesis and carbon assimilation. This in turn, reduced vegetative growth in “Off” years and potential yield in subsequent “On” years. Irrigation level did not affect yield parameters beyond number of fruit per plant and, due to pruning protocols, had only a small effect on commercial yield, limited to high bearing level years. Therefore, WP increased as irrigation application rate decreased. Stress to jojoba caused by severe drought was found to be detrimental to production, whether occurring early in the season and leading to reduced seed size and less wax content, or occurring mid-summer following wax accumulation and leading to inhibition of seed drop and inefficient harvesting.
References
Adams JA, Bingham FT, Kaufmann MR, Hoffman GJ, Yermanos DM (1978) Responses of stomata and water, osmotic, and turgor potentials of jojoba to water and salt stress. Agron J 70:381–387. https://doi.org/10.2134/agronj1978.00021962007000030005x
Agarwal S, Kumari S, Khan S (2023) Jojoba (Simmondsia chinesis): an oil producing cash crop. Neglected and underutilized crops. Academic, pp 757–779
Ayerza R (1993) Effect of irrigation on jojoba production under arid chaco conditions: II-Seed yields and wax quality. J Am Oil Chem Soc 70:1225–1228. https://doi.org/10.1007/BF02564230
Ben-Gal A, Kool D, Agam N, van Halsema GE, Yermiyahu U, Yafe A, Presnov E, Erel R, Majdop A, Zipori I, Segal E (2010) Whole-tree water balance and indicators for short-term drought stress in non-bearing ‘Barnea’ olives. Agr Water Manage 98:124–133. https://doi.org/10.1016/j.agwat.2010.08.008
Ben-Gal A, Yermiyahu U, Zipori I, Presnov E, Hanoch E, Dag A (2011) The influence of bearing cycles on olive oil production response to irrigation. Irrig Sci 29:253–263. https://doi.org/10.1007/s00271-010-0237-1
Ben-Gal A, Beiersdorf I, Yermiyahu U, Soda N, Presnov E, Zipori I, Ramirez Crisostomo R, Dag A (2017) Response of young bearing olive trees to irrigation-induced salinity. Irrig Sci 35:99–109. https://doi.org/10.1007/s00271-016-0525-5
Ben-Gal A, Ron Y, Yermiyahu U, Zipori I, Naoum S, Dag A (2021) Evaluation of regulated deficit irrigation strategies for oil olives: a case study for two modern Israeli cultivars. Agr Water Manage 245:106577. https://doi.org/10.1016/j.agwat.2020.106577
Benzioni A (1995) Jojoba Domestication and Commercialization in Israel. Ch7 In Hortic Rev, J. Janick (Ed.). 17, 233–266. https://doi.org/10.1002/9780470650585
Benzioni A (2006) Jojoba research as basis for domestication of jojoba in Israel. Isr J Plant Sci 54:157–167. https://doi.org/10.1560/IJPS_54_3_157
Benzioni A, Dunstone RL (1986) Jojoba: adaptation to environmental stress and the implications for domestication. Q Rev Biol 61:177–199. https://www.journals.uchicago.edu/doi/abs/10.1086/414898
Benzioni A, Nerd A (1985) Effect of irrigation and fertilization on vegetative growth and yield of jojoba in relation to water status of the plants. Jojoba: Proceedings of the Sixth International Conference on Jojoba and Its Uses, October 21–26, 1984, Beer-Sheva, Israel/Jaime Wisniak and Jacob Zabicky, Editors
Benzioni A, Vaknin Y (2002) Effect of female and male genotypes and environment on wax composition in jojoba. JAOCS. J Am Oil Chem Soc 79:297–302. https://doi.org/10.1007/s11746-002-0477-7
Benzioni A, Mizrahi Y, Nerd A (1980) Effects of water and fertilization regimes on floral bud dormancy, fruit set and vegetative growth of jojoba plants. Proc 4th Inter. Conf. on Jojoba and Its Uses, (IM Puebla (Ed.), Hermosillo, Mexico, 162–166
Benzioni A, Ventura M, De Malach Y (1996) Long-term effect of irrigation with saline water on the development and productivity of jojoba clones. J Hortic Sci 71:835–846. https://doi.org/10.1080/14620316.1996.11515466
Cochard H, Coll L, Le Roux X, Améglio T (2002) Unraveling the effects of plant hydraulics on stomatal closure during water stress in walnut. Plant Physiol 128:282–290. https://doi.org/10.1104/pp.010400
Dag A, Bustan A, Avni A, Tzipori I, Lavee S, Riov J (2010) Timing of fruit removal affects concurrent vegetative growth and subsequent return bloom and yield in olive (Olea europaea L.). Scientia Horticulturae 123:469–472. https://doi.org/10.1016/j.scienta.2009.11.014
Downton WJS, Loveys BR, Grant WJR (1988) Non-uniform stomatal closure induced by water stress causes putative non‐stomatal inhibition of photosynthesis. New Phytol 110:503–509. https://doi.org/10.1111/j.1469-8137.1988.tb00289.x
Dunstone RL, Begg JE Biology of jojoba. Proceedings 1st Australian Jojoba Conference, Bathurst NSW (1979) pp. 1–18. http://hdl.handle.net/102.100.100/298134?index=1
Ehrler WL, Fink DH, Mitchell ST (1978) Growth and yield of jojoba plants in native stands using runoff-collecting microcatchments. Agron J 70:1005–1009. https://doi.org/10.2134/agronj1978.00021962007000060028x
Fernández JE (2017) Plant-based methods for irrigation scheduling of woody crops. Horticulturae 3:35
Fernández JE, Cuevas MV (2010) Irrigation scheduling from stem diameter variations: a review. Agr for Meteorol 150:135–151. https://doi.org/10.1016/j.agrformet.2009.11.006
Gentry HS (1958) The natural history of Jojoba (Simmondsia chinensis) and its cultural aspects. Econ Bot 12:261–295. https://doi.org/10.1007/BF02859772
Glat D, Dobrenz A, Palzkill D (1983) Carbon dioxide exchange processes in jojoba. CALS Publications Archive. The University of Arizona
Jones HG, Sutherland RA (1991) Stomatal control of xylem embolism. Plant Cell Environ 14:607–612. https://doi.org/10.1111/j.1365-3040.1991.tb01532.x
Khattab EA, Afifi MH, Amin GA (2019) Significance of nitrogen, phosphorus, and boron foliar spray on jojoba plants. B Natl Res Cen 43:1–8. https://doi.org/10.1186/s42269-019-0109-7
Kumar A, Sharma N, Wani TF, Sharma R (2023) Water Productivity of Temperate fruits in Climate Change scenario. Advances in Water Management under Climate Change. CRC, pp 210–235
Lazare S, Zipori I, Cohen Y, Haberman A, Goldshtein E, Ron Y, Rotschild R, Dag A (2021) Jojoba pruning: new practices to rejuvenate the plant, improve yield and reduce alternate bearing. Sci Hort 277:109793. https://doi.org/10.1016/j.scienta.2020.109793
Lortie CJ, Aarssen LW (1996) The specialization hypothesis for phenotypic plasticity in plants. Int J Plant Sci 157:484–487
Morgan KT, Barkataky S, Kadyampakeni D, Ebel R, Roka F (2014) Effects of short-term drought stress and mechanical harvesting on sweet orange tree health, water uptake, and yield. HortScience 49:835–842. https://doi.org/10.21273/HORTSCI.49.6.835
Moriana A, Pérez-López D, Prieto MH, Ramírez-Santa-Pau M, Pérez-Rodriguez JM (2012) Midday stem water potential as a useful tool for estimating irrigation requirements in olive trees. Agr Water Manage 112:43–54. https://doi.org/10.1016/j.agwat.2012.06.003
Nelson JM, Palzkill DA, Bartels PG (1993) Irrigation cut-off date affects growth, frost damage, and yield of jojoba. J Am Soc Hort Sci 118:731–756. https://doi.org/10.21273/JASHS.118.6.731
Nonami H (1998) Plant water relations and control of cell elongation at low water potentials. J Plant Res 111:373–382. https://doi.org/10.1007/BF02507801
Osman HE, AboHassan A (2002) Effect of drought stress on flowering and on seed yield and its components in jojoba (Simmondsia chinesis) established in Western Saudi Arabia. Meteorol, Environ Arid Land Agr Sci, 13, 121–131
Perry A, Tel-Zur N, Dag A (2021) Vegetative and reproductive response to fruit load in two Jojoba (Simmondsia chinensis) cultivars. Agron 11:889. https://doi.org/10.3390/agronomy11050889
Purcell HC, Abbott TP, Holser RA, Phillips BS (2000) Simmondsin and wax ester levels in 100 high-yielding jojoba clones. Ind Crop Prod 12:151–157. https://doi.org/10.1016/S0926-6690(00)00066-2
Sandouqa A, Al-Hamamre Z (2019) Energy analysis of biodiesel production from jojoba seed oil. Renew Energ 130(2019):831–842. https://doi.org/10.1016/j.renene.2018.07.015
Wardlaw IF, Begg JE, Bagnall D, Dunstone RL (1983) Jojoba: temperature adaptation as expressed in growth and leaf function. Funct Plant Biol 10:299–312. https://doi.org/10.1071/PP9830299
Yermanos DM, Duncan CC (1976) Quantitative and qualitative characteristics of jojoba seed. J Am Oil Chem Soc 53:80–82. https://doi.org/10.1007/BF02637398
Acknowledgements
The research was funded by the Chief Scientist of the Israel Ministry of Agriculture and Rural Development, grants number 20-13-005 and 20-01-0174. We thank Yulia Sobbotin and Talal Hawashla for their technical assistance and Ronen Rotschild and Yizhar Eilat, (Jojoba Israel, Kibbutz Hatzerim, Israel) for their advice and for allowing us to conduct the experiment in their field. We thank ICL and Deshen Gat for providing us with the fertilizers for the experiment.
Funding
Open access funding provided by The Agricultural Research Organization of Israel.
Author information
Authors and Affiliations
Contributions
Study conception and design by A.B-G, A.D. and U.Y. Procurring of funding by A.D. and A.B-G. Material preparation, data collection and analysis were performed by S.B. Y.R. A.P. with guidance from A.B-G. U.Y. and and A.D. The first draft of the manuscript was written by A.B-G. based on a thesis written by S.B. All authors commented on previous versions of the manuscript and read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ben-Gal, A., Badichi, S., Ron, Y. et al. Effects of irrigation amounts and a deficit irrigation strategy on water status and yields of intensively cultivated jojoba (Simmondsia chinensis). Irrig Sci (2024). https://doi.org/10.1007/s00271-024-00937-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00271-024-00937-3