
Abstract
We used video playback of courting male Schizocosa ocreata wolf spiders to examine responses of intended receivers (conspecific females) and eavesdroppers (competitor males, predatory spiders, toads) to manipulations of spider color (natural color, monochromatic gray, monochromatic RBG average) displayed against complex leaf litter backgrounds (color, grayscale). Models of chromatic and achromatic contrast between spider stimuli and backgrounds were used to predict receiver responses. The results support the hypothesis that interactions between spider and background coloration affect detection and recognition, although responses varied with receiver type. Detection responses of intended receivers (female S. ocreata) did not fit predictions of the chromatic contrast model in some cases, but showed a fair fit to the achromatic model. Detection responses of social eavesdroppers (male S. ocreata) fit the chromatic and achromatic contrast models slightly better than did female responses (poor fit and very good fit, respectively). Eavesdropping wolf spider predators (Rabidosa) exhibited detection responses that significantly matched predictions of the chromatic (very good fit) and achromatic (excellent fit) models. Whereas jumping spiders (Phidippus) showed a good fit to the chromatic and achromatic contrast models, toad predators had a good fit only to the chromatic model. Recognition responses revealed a different pattern of fit to the chromatic and achromatic models across receiver types, although Rabidosa again indicated a significant fit to both models. Taken together, the results of this study identify both chromatic and achromatic features of spider appearance as likely explanations for differences in behavioral responses of intended and unintended receivers. This outcome suggests the possibility that both sexual and natural selection likely target different features of male appearance during courtship.
Significance statement
Some of the most striking examples of animal displays involve conspicuous male courtship signals, although such signals often can be exploited by eavesdropping competitors and predators. In this study we manipulated the coloration of courting male Schizocosa ocreata wolf spiders and their leaf-litter backgrounds from pre-existing video footage. We then played back the altered video clips to female conspecifics and eavesdroppers (competing conspecific males and potential predators: non-conspecific wolf spiders, jumping spiders, and toads) and recorded their responses. From these responses we scored both stimulus detection (latency to orient) and recognition (receptivity movements in conspecific females, tapping bouts in conspecific males, and attack in other viewers). Color differences in the courting male stimuli and leaf-litter backgrounds proved to be important for detection and recognition, even for wolf spiders with limited dichromatic vision. Moreover, for jumping spiders and predatory toads with tetrachromatic and trichromatic vision respectively, color of the prey spider stimulus was particularly salient. Overall, our experimental results support the hypothesis that receivers with different visual capacities should vary in their responses to color and contrast of courting male spiders against complex backgrounds.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Animals use a variety of signaling methods and modalities, including chemical, acoustic, visual and seismic (Partan and Marler 2005; Hebets and Papaj 2005; Hebets et al. 2016). These signals are used in many different situations, but the signals used by males to attract females may be the most important for male reproductive success (Choi et al. 2022). Much evidence suggests that male courtship signals are selected to be conspicuous to females against a complex background of environmental noise (Rosenthal 2007; Wiley 2013; Ord et al. 2021), but such signals also may be detected by others (predators, competitors) in the communication network (McGregor 2005; Searcy and Nowicki 2005; Roberts et al. 2007; Cummings et al. 2008; Roberts and Uetz 2008; Marshall et al. 2015; Bernal and Page 2022). Thus, evolution of signaling behavior is likely to be shaped by selection from both intended receivers and eavesdroppers.
In communication networks, unintended receivers may include social eavesdroppers - i.e., conspecifics that recognize and exploit the information content of signals (Peake 2005; Dabelsteen 2005; Searcy and Nowicki 2005), or interceptive eavesdroppers - i.e., predators for which signals reveal cues to the presence of potential prey (Zuk and Kolluru 1998; Haynes and Yeargan 1999; Rosenthal et al. 2001; McGregor 2005; Rosenthal 2007; White et al. 2022). Most research on communication networks has examined eavesdropping in several highly specific contexts (see McGregor 2005 and chapters within), including: bystanders observing male-male contests (Johnstone 2001; Tibbetts et al. 2020), female mate choice copying (Dugatkin 1992; Dugatkin and Godin 1993; Jones and DuVal 2019), and predator detection of male courtship signals (Kotiaho et al. 1998; Koga et al. 2001; Macedonia et al. 2002, 2009; Peake 2005; White et al. 2022). Even in more general communication contexts, intended receivers and different types of eavesdroppers might independently impose selection on different signals, or act together to intensify selection on one signaling trait. Much less is known about the combined influence of targeted receivers, social eavesdroppers and interceptive eavesdroppers, especially on signals in multiple sensory modes (Roberts et al. 2007; Roberts and Uetz 2008; Bernal and Page 2022; Postema et al. 2023).
Studies of prey detection against complex backgrounds have suggested that prey may achieve maximum concealment by using optical patterns that combine background matching and disruptive coloration (Bond and Kamil 2006; Stevens and Cuthill 2006; Stevens et al. 2006; Endler 2006; Stevens et al. 2008; Dimitrova and Merilaita 2009; Stevens and Merilaita 2009a, b; Price et al. 2019; Galloway et al. 2020; Postema et al. 2023). At the same time, aspects of male visual courtship signals that augment female detection and discrimination (movement, contrast, pattern) may also affect detection by predators. A predator’s response might be avoided if signals disrupt the signaler’s visual outline or enhance its crypsis against complex backgrounds (Stevens et al. 2006; Endler 2006; Cummings et al. 2008; Dimitrova and Merilaita 2009; Merilaita et al. 2017; Bernal and Page 2022). Moreover, different aspects of visual perception are related in complex and sometimes opposing ways (see McGregor 2005; Endler 2006; Rosenthal 2007; Davis et al. 2022). For example, a female may be able to detect male signals against a complex background, but whether or not she is able to recognize the signaling species or discriminate mate quality information from those signals will influence her response (receptivity or sexual cannibalism). The female’s response will, in turn, impact male survival, mating success, and ultimately signal evolution (Guilford and Dawkins 1991; Endler et al., 2005; Peters et al. 2009; Wiley 2017; Caves et al. 2024).
Experimentally isolating and manipulating interrelated signal components, as well as features of the physical environment in which they are transmitted, can determine how different aspects of male signals function. Such manipulations have the potential to reveal how the environment has shaped the design of signals as well as their perception – a process known as “sensory drive” (Endler 1992; Cummings and Endler 2018; Fleishman et al. 2022). In our particular case, we wish to know whether selection has maximized signal perception by targeted female receivers, as well as whether it has affected exploitation by social and interceptive eavesdroppers. Rosenthal (2007) has suggested separating chromatic (color) and spatial (achromatic contrast and background complexity) information in testing hypotheses about the role of spatiotemporal aspects of visual signals and backgrounds. Video playback is ideal for such an approach, as it allows independent manipulation of color and background complexity. In addition, moving animal images can be superimposed against experimental backgrounds.
Following the suggestions of Rosenthal (2007), we used digital video playback to examine responses of intended and unintended receivers to video playback of courting male Schizocosa ocreata wolf spiders. We combined altered male morphology (color / pattern) with aspects of environmental backgrounds (chromatic / spatial information) to test how visual signals affect detection and discrimination by: (a) intended receivers (female S. ocreata); (b) social eavesdroppers (male S. ocreata); and (c) interceptive eavesdroppers (predators). We formulated an overarching hypothesis that addresses how visual signal coloration will affect detection and recognition by receivers against a complex visual background. H1: The magnitude of contrast (chromatic, achromatic) between visual signals and complex visual backgrounds differing in color (naturalistic, grayscale) will affect detection (indicated by orientation latency) and recognition (indicated by appropriate behaviors) by receivers. Given the variation in visual systems of the vertebrate and several invertebrate receivers studied here, we created chromatic and achromatic contrast visual models tailored to the specific spectral sensitivities of the receivers we studied (described in detail below in Methods) and used them to generate specific predictions about receiver behaviors.
Background – study species
Studies of the leaf litter-dwelling wolf spider, Schizocosa ocreata (Hentz), have demonstrated that males may benefit by courting females against backgrounds on which they are most conspicuous (Clark et al. 2011; Uetz et al. 2011). Importantly, the coloration of male wolf spiders renders them both conspicuous and cryptic simultaneously (Fig. 1), depending on perception from lateral (female spiders, spider predators, toads) or dorsal (avian predators) view (Clark et al. 2011, 2016; Uetz et al. 2011; Rubi et al. 2019). When viewed from above, the color of the male dorsal median stripe matches dead brown leaf litter and soil, suggesting cryptic or disruptive coloration when viewed by potential predators (Rubi et al. 2019).

Male Schizocosa ocreata (Araneae: Lycosidae). The photo of the spider from this angle shows both the lateral (dark) and dorsal (leaf-colored) coloration. A study by Clark et al. (2011) examines this coloration pattern and its role in detectability in detail
However, when viewed from a lateral perspective male coloration contrasts highly with the leaf litter background, which suggests greater conspicuousness from the perspective of a female conspecific (Clark et al. 2011) as well as (other) potential predators (Clark et al. 2016). This relationship between viewing perspective and detectability poses some interesting questions about environmental constraints on visual signals – particularly the interaction of signals and visual background complexity and how that interaction affects detection and discrimination by different receivers (Roberts and Uetz 2008; Marshall et al. 2015; Clark et al. 2016; Rubi et al. 2019; White et al. 2022; Rowe et al. 2024).
Using video playbacks, Uetz et al. (2011) found that courtship displays and leg decorations of male S. ocreata are highly conspicuous against leaf-litter backgrounds, and that both complexity and light level of the display background affect the detection of male visual signals. Results also showed that the presence of male leg tufts increases chances of detection and receptivity by conspecific females. Previous research by Roberts et al. (2008) demonstrated that males with ornaments (leg tufts) are at greater risk of detection by predatory toads than are males without ornaments, but little is known about how a predator might cue into the courtship behavior itself. More recent video playback studies with American Toads (Clark et al. 2016) and Blue Jays (Rubi et al. 2019) also have shown that spider coloration (i.e., cryptic vs. conspicuous) and courtship behavior movements affect predator detection. Notably, most of the earlier studies were focused on male ornamentation (leg tufts) as a signal and were limited to a single type of receiver.
In this study we examine the differential detectability and discriminability of male courtship behavior that arises from visual contrast with complex environmental backgrounds. To address our questions, we tested viewers with different spectral sensitivities, which include intended receivers (conspecific females), social eavesdroppers (conspecific male competitors), and interceptive eavesdroppers (predators).
Methods
Animal subjects, care
Wolf spiders were collected from the Cincinnati Nature Center Rowe Woods facility near Milford, Clermont County, OH (39˚ 7’31.15” N; 84˚ 15’4.29” W), in May and June of 2013–2015. Female S. ocreata (N = 69) were collected as juveniles and raised to maturity in the lab to ensure lack of exposure to males and prevent bias from experience (Stoffer and Uetz 2015). Male S. ocreata (N = 118) were collected as adults so that they were previously exposed to male courtship during breeding season, when eavesdropping behavior is learned (Clark et al. 2015).
Eavesdropping predators are common and sympatric with S. ocreata in deciduous forests of eastern North America. Our test species (other than S. ocreata) were chosen because they hunt using visual cues (Beck and Ewert 1979; Aho et al. 1993), readily accept Schizocosa wolf spiders as prey (Uetz, pers. obs.), and react to videos of wolf spider courtship with predatory behavior (Roster et al. 1995; Roberts et al. 2007; Roberts and Uetz 2008).
We studied two spider species that are known predators of S. ocreata and which have highly acute vision: Phidippus clarus jumping spiders (Family Salticidae: Zurek et al. 2015); and Rabidosa rabida wolf spiders (Family Lycosidae: DeVoe et al. 1969). Specimens of R. rabida wolf spiders (N = 42) were field-collected from New Richmond, OH (39° 0’56” N, − 84°5’82” W), and specimens of P. clarus jumping spiders (N = 34) were field-collected from Pymatuning, PA (41° 38’ 33.72” N, -80° 25’ 39.72” W) during July 2014. All spiders were housed in opaque, plastic deli dish containers (10 cm diameter) and maintained at room temperature (22–25˚C) with stable humidity and a 13:11 h light/dark cycle to approximate late spring/early summer lighting conditions. Spiders were supplied water ad libitum (although predatory Rabidosa and Phidippus were starved for three to four days before being used in a trial).
The vertebrate predator we chose for this study was the American toad (Anaxyrus americanus). Toads are well known to feed on a variety of arthropods including spiders (Clarke 1974), and field observations (G. Uetz) have shown that S. ocreata is a common prey item for toads. Previous studies have demonstrated that A. americanus responds to digital sequences of spider behavior with typical toad predatory behavior (Roster et al. 1995; Roberts and Uetz 2008; Clark et al. 2016).
Juvenile and adult toads either were collected locally in Central Michigan (43o 330 46.43′′ N; 84o 370 15.43′′ W; n = 100 total collected) during spring and summer months of 2013–2016 or were purchased from a commercial dealer (Charles D. Sullivan Company, Nashville, TN, USA; n = 150 total purchased). All toads were housed at Alma College, Alma Michigan and maintained according to a protocol approved by the Alma College Animal Care Committee (Protocol #A4162-01). Each toad was kept individually in a 35 cm (length) x 18 cm (width) x 12 cm (height) plastic container layered with peat and sphagnum moss bedding. Ventilation was provided by drilling small holes into each end of the container. Toads were misted daily and fed crickets (Acheta domesticus) twice weekly.
Experimental methods – video Playback
Following the suggestions in Rosenthal (2007), we digitally-manipulated pre-existing courting spider and background videos used in many previous studies (see Uetz et al. 2015). We compared female S. ocreata and eavesdropper responses to video playback of a courting male in the following treatments: (1) an unmanipulated digital color video of a courting male spider (i.e., natural unmanipulated spider = control), (2) the same video as in “1” with chromatic information “removed” (i.e., a grayscale monochromatic spider = gray mono), and (3) the same video as in “1” with surface detail information removed (i.e., a standardized average RGB mono chromatic value spider = RGB mono). These courting male video images were displayed against two background color treatments: (1) natural complex color leaf litter (= color litter); and (2) grayscale complex leaf litter (= grayscale litter). LCD screen intensity was adjusted to match closely that of the natural environment from measurements collected under field conditions using an Ocean Optics USB2000 spectroradiometer. Video playback trials were conducted using digital video images, as all receivers, i.e., female and male wolf spiders (S. ocreata), predatory wolf spiders (Rabidosa rabida), jumping spiders (Phidippus clarus) and American toads (Anaxyrus americanus) perceive moving video stimuli and interpret them as real (Clark and Uetz 1990; Roster et al. 1995; McClintock and Uetz 1996; Roberts et al. 2007; Roberts and Uetz 2008).
Spider and toad subjects were presented with video playback of the three male spider color treatments (control, gray mono and RGB mono) against a background of either natural color litter or grayscale litter. For each presentation, subjects were chosen randomly and used only once so that all trials were independent. The manipulated video stimuli of male S. ocreata courtship and background color treatments created a 3 × 2 factorial design, presenting subjects with three different levels of chromatic contrast against two backgrounds (Fig. 2). For all spider trials, we presented test subjects with a lateral view perspective of male performing courtship behavior, as that is how they typically would be perceived.

Experimental design, with still images from each video playback treatment of spider color x complex litter background. Video playback exemplars were manipulated to provide varying degrees of chromatic and achromatic contrast with backgrounds
Experimental methods – measuring light Spectra and calculating Visual contrast
General procedure - spectral data - Our methods for gathering spectral data from digital stimuli presented on an Apple iPod® Touch (5th Generation, Apple Inc.) are like those used to collect spectral data from live spiders and natural backgrounds (see Clark et al. 2011). In the present study we acquired emission spectra from the LCD light-emitting iPod® screen and modified our measurement methodology to calculate reflectance of digital leaf litter backgrounds and spider stimuli.
Adapting background, Leaf litter, and spider image measurements
To estimate how an animal perceives a visual scene, several types of spectral data must be obtained and converted to units appropriate for vision (Endler 1990). For our study these data include irradiance spectra of the visual adapting background (i.e., the entire background scene), reflectance samples of leaf litter, and reflectance measurements of spider stimuli. All spectral data were recorded using an Ocean Optics USB2000 spectroradiometer and an Apple® laptop computer running the OceanView (v2.0.12) software program (Ocean Optics, Inc.). We sampled the irradiance spectrum of the iPod® screen (N = 3 each for our color and grayscale backgrounds) using an Ocean Optics CC3-UV cosine-corrected irradiance probe. The three samples for each background type then were averaged. Next, we obtained reflectance measurements of leaf litter in the visual background (N = 17 each for the color and grayscale backgrounds) using an Ocean Optics 400 nm fiber optic probe. The adapting background (= screen irradiance excluding the spider image) then was converted to radiance by multiplying the averaged irradiance spectrum of the background by the reflectance samples of leaf litter. We likewise collected reflectance spectra of the digital spider images (N = 5 for each spider stimulus), which were converted to radiance spectra by multiplying the averaged irradiance of a specific background (color or grayscale) by the reflectance samples of each spider stimulus (i.e., control, gray mono and RGB mono).
To calibrate the reflectance program prior to sampling, we used a digital pure white image set to RGB values of 255 (R), 255 (G), 255 (B), which equals pure white (to human vision) on a digital screen. We set the white standard curve in “scope mode” to a maximum count of approximately 3,500 units and removed the electric dark current (electrical noise in the absence of light) by covering the probe and taking a dark sample. Finally, we set spectrum averaging to five samples and boxcar width to three. To maintain a constant distance between the end of the probe and the iPod® screen the probe was clamped 1.25 cm above the screen, resulting in a sample spot size of approximately 1.0 mm diameter. All measurements were performed in a darkened room.
Visual modeling of Chromatic and Achromatic contrast
Each class of photoreceptors exhibits a visual pigment that absorbs light over a specific range of wavelengths. Whereas the summed response from one or more photoreceptor classes accounts for luminance (achromatic brightness) perception, color perception requires overlap in the absorption spectra of at least two different photoreceptor classes (Osorio and Vorobyev 2005). Across animal taxa, larger numbers of photoreceptor classes provide greater color discrimination ability than fewer classes (Osorio and Vorobyev, 2008).
We generated photoreceptor absorbance curves for our study species using peak absorptance (λmax) values (per Govardovskii et al. 2000) provided in previously published research of our study taxa (see below). Although the light emittance spectrum of the iPod® is constrained to the human-visible range of 400–700 nm, we included wavelengths at and above 400 nm for the UV photoreceptor in our visual models of the jumping spider and wolf spiders. We included this range because the visible-wavelength tail of the UV absorptance curve extends to approximately 450 nm in jumping spiders and 475 nm in wolf spiders. Importantly, the UV photoreceptors of these spiders overlap the sensitivities of one or more other photoreceptors, and thus likely aid in chromatic discrimination of images displayed on the iPod® screen.
In modeling animal color perception, researchers often use either receptor noise (Vorobyev and Osorio, 1998) or Euclidean distance (Endler and Mielke 2005). Fleishman et al. (2016a, b) have shown that although the two approaches produce similar results, the receptor noise method requires information that frequently is unknown (e.g., ratios of different photoreceptor classes) and a surrogate species or simply a guess is substituted. As we lacked data on several receptor noise parameters for some of our study species, we chose the simpler Euclidean distance approach. For details of the procedure and equations used to generate our visual models for our study species, see Appendix I.
Wolf Spider Visual Model
Wolf spiders of the family Lycosidae have well developed visual systems and are known to be active diurnal and nocturnal hunters that rely on eyesight (Rovner 1993, 1996). While few studies have been conducted on the spectral sensitives of wolf spiders, DeVoe et al. (1969) recorded ERGs from the eyes of three different species (Lycosa baltimoriana, L. miami and L. carolinensis). These authors reported that the principal eyes of their wolf spider species exhibited spectral sensitivity peaks at 380 nm (UV) and from 505 to 510 nm. Based on the findings of DeVoe et al. (1969), we used peak spectral sensitives of 380 and 507.5 nm in our visual models for Schizocosa ocreata and Rabidosa wolf spiders.
Jumping spider visual model
Jumping spiders are known for their excellent vision used in hunting and in viewing their colorful displays (Land and Fernald 1992; Land and Nilsson 2002; Lim and Li 2004). Several studies conducted on jumping spider spectral sensitivity have revealed highly variable visual capacities, ranging from tetrachromats with four classes of photoreceptors to dichromats with two photoreceptor classes (Lim and Li 2006; Glenszczyk et al. 2022). DeVoe (1975) reported the spectral sensitivity of the jumping spider Phidippus regius as being dichromatic, with a UV-sensitive (UVS) photoreceptor that peaks at 370 nm and a green-sensitive photoreceptor (MWS) that peaks at 532 nm. However, Yamashita and Tateda (1976) as well as Lim and Li (2006) argued that DeVoe (1975) likely was incorrect about Phidippus possessing only two photoreceptor classes, given that Land’s (1969) findings suggested the presence of tetrachromacy in Phidippus and Metaphidippus, ranging from ultraviolet-sensitivity (UVS) to red-sensitivity (LWS). Thus, in our visual model for the jumping spider Phidippus clarus, we used the four photoreceptor classes reported by Yamashita and Tateda (1976) for Menemerus confusus of 360 nm (UVS), 490 nm (SWS), 530 nm (MWS), and 580 nm (LWS).
Toad Visual Model
The visual systems of frogs and toads have been well studied and it is known that these amphibians use vision primarily for hunting and prey capture (Ingle, 1968). More recent studies (Yovanovich et al. 2017) have demonstrated that anurans exhibit color vision in dimly lit environments due to the presence of multiple classes of rods. Yovanovich et al. (2017) also reported the properties of photoreceptors from several different species of frogs and toads, including three species in the genus Bufo. As our study species, Anaxyrus americanus (formerly Bufo americanus), is closely related to the Bufo species reported by Yovanovich et al. (2017), we used spectral sensitivity peaks at 432 nm (SWS), 502 nm (MWS), and 562 nm (LWS) for our toad visual model.
Experimental methods - spiders
The trials involving spiders (female and male S. ocreata, predatory Rabidosa and Phidippus) were conducted in standard video playback arenas identical to other studies (Fig. 3): circular clear plastic arenas (15 cm diameter, 6.5 cm high) used in previous studies (McClintock and Uetz 1996; Uetz et al. 2011) with images presented on an iPod Touch® screen.

Diagram of clear plastic arena (15 cm diameter, 6.5 cm high) with iPod Touch® as video playback source
Focal subjects were gently released from a vial placed below the arena and introduced to the video playback arenas through a small hole in the empty arena floor (to mimic their natural movements within the leaf litter of the forest floor) and presented immediately with one of the randomly assigned videos. Trials were recorded from above at a 45° angle using a Sony Handycam HDR-XR260. However, because video screens were visible (i.e., when filmed from a 45° angle), it was impossible not to see which spider and background stimulus was being used and so trials could not be scored blind. We therefore conducted inter-observer analyses and determined a lack of observer bias (p > 0.05 for all variables measured) before further data analysis.
Experimental methods - vertebrate predators (toads)
The American Toad (Anaxyrus americanus) hunts visually and responds to prey items meeting a minimum size threshold (approx. 2.5 mm), but are sensitive to small differences in orientation and size in any prey items large enough to stimulate a response (Beck and Ewert 1979). Toads readily stalk and consume S. ocreata, in leaf litter mesocosms, and constitute a significant source of predation in the field (Clarke 1974). Results of previous studies show that toads recognize video spiders as prey and respond to differences in tuft size (Roster et al. 1995; Roberts and Uetz 2008).
Toads were tested in a similar manner to spiders. We presented digital stimuli to toads in a plastic chamber that measured 28 cm (width) x 40 cm (length) x 22 cm (height), where they were released from a small tube positioned perpendicular to an iPod® screen. Toads were presented with a lateral perspective of a courting male wolf spider against leaf litter backgrounds as in the other experiments. Using this method, the latency to first detection, i.e., or the optomotor response of the toad, could easily be observed and scored by the turning of the head toward the stimulus. Within the chamber, the iPod® was positioned at approximately toad eye level 30 cm from the release tube.
Experimental methods - description of behaviors and Scoring
Video playback trials with S. ocreata were conducted within the first week after field-collected males were captured, or one week after lab-reared females reached sexual maturity. For spider test subjects, video files were scored following trials using an event recorder (Spectator Go!® Biobserve.com on an iPad®). Female responses to video stimuli and receptivity displays were scored as in previous studies (Uetz and Norton 2007). The following female response variables were recorded for each trial: (1) detection - latency to orient to the male; (2) recognition/receptivity - number of individual receptivity displays (slow pivot, settle, tandem leg extend) summed as a composite receptivity score. When orientation or approach was not seen, trial length (300 s) values were substituted.
For eavesdropping competitor males, investment in courtship is indicated by the frequency and duration of leg tapping and other behaviors. As in previous studies (Clark et al. 2012, 2015), we scored the number of courtship tapping bouts of test subject in response to the different video exemplars. Each male was tested only once to control for experience effects with the video apparatus.
For eavesdropping invertebrate predators, the following response variables were recorded for each trial of wolf spiders (Rabidosa) and jumping spiders (Phidippus): (1) detection - latency to orient to the male; (2) recognition - latency to approach and attack the male on the screen. When orientation or attack were not seen, trial length (300 s) values were substituted.
Individual toads from the laboratory population were exposed to each experimental video treatment for a single exemplar (N = 15 toads for each of 3 exemplars). Toads were tested once / day and fed after trials to control for hunger effects. We recorded toad behavior and scored latency (secs) of the optomotor response as a proxy for attack. The results of previous studies indicated that when toads oriented and approached in a stalking pose, they were highly likely (> 85%) to lunge (attack) at the stimulus (Roster et al. 1995; Roberts and Uetz 2008; Clark et al. 2016). Because toads habituate quickly following unsuccessful attacks on video prey and are unlikely to provide realistic measures of response when used repeatedly (Aho et al. 1993), the latency of optomotor response (orientation/initiation of attack) was recorded, and the trial was terminated before any capture attempt before any capture attempt. Each experimental trial lasted 5 min or when orientation occurred, whichever came first. This rule minimized habituation and allowed toads to be used more than once. We used Scribe 4.2 (Duke and Stammen 2011) as a computer event recording program.
Statistical analyses
Data from the behavioral studies cited above were analyzed using JMP 16.0© 2021 (SAS Institute, JMP Statistical Discovery LLC). Data on behaviors were first tested for fit to a normal distribution. After determination of goodness of fit (or lack thereof), behavioral data were subjected to a Generalized Linear Model (GLM) with either a Poisson distribution (for continuous data, e.g., counts and rates of behaviors) or a binomial distribution (for categorical data, e.g., mate/not). The latency of receiver responses was subjected to a parametric survival analysis via GLM. We used a full factorial design (spider color, background type and color x background interaction term) for most analyses.
Results
Chromatic and achromatic contrast
Predictions of the chromatic contrast model (Fig. 4a) suggest that against the color background, contrast values are similar for different receiver types (jumping spiders, wolf spiders, toads), but show considerable variation based on spider exemplar types (control, gray mono and RGB mono). Against a color background, control spider exemplars show higher contrast values and would be very dark, while gray mono and RGB mono spider exemplars have lower contrast values and are similar to each other. Consequently, we predict that orientation latency for all receiver types should be shorter for control spiders than other spider exemplars.
Against a grayscale litter background, predicted contrast values are lower overall for control and gray mono spider exemplars than they are against the color background (Fig. 4a). However, control spider and RGB mono spider exemplars have greater contrast than gray mono spiders (which show very little contrast against the grayscale background). Orientation latency therefore should be shorter for control and RGB mono spiders against grayscale litter, and behaviors indicating recognition should be greater.
Predictions based on achromatic (brightness) contrast (Fig. 4b) suggest that, like chromatic contrast, achromatic contrast between spiders and backgrounds will vary more with spider exemplar color than with background or receiver type. Achromatic contrast values for the natural dark coloration of the control spider exemplar far exceed the other two spider exemplars (gray mono, RGB mono), regardless of background. Thus, we predict that orientation latency for all receiver types should be briefer for control spiders and much longer for other spider exemplars. Likewise, we predict that recognition responses should follow this same pattern.

Mean (± S.E.) contrasts of the spider stimuli (control, gray mono, RGB mono) against a leaf litter (color, grayscale) background. a Chromatic contrast, b Achromatic contrast. Contrasts are Euclidean distances between the spider stimuli and backgrounds (see text for details of calculations)
Behavioral responses of receivers
Given the predictions of the chromatic and achromatic contrast models, we examined the behavioral responses of each receiver type here, then summarized the fit of behavioral responses to model predictions in Table I at the end of the Results section.
Intended receivers (female S. ocreata)
For female S. ocreata detection (orientation latency) responses, results of a Generalized Linear Model (GLM) with a Poisson distribution showed significance overall: Χ25 = 1235.430; P < 0.0001, with significance for spider exemplar color (Χ22 = 167.842; P < 0.0001) but not background (Χ21 = 1.060; P = 0.321). In addition, the interaction of spider color x background was significant (Χ22 = 1184.551; P < 0.0001), reflecting differences in patterns of female responses (Fig. 5). Responses of female S. ocreata to playback of male courtship (Fig. 5a) indicated that, against a color litter background, the gray mono spider took longer to detect, whereas the control and RGB mono spider were seen more quickly. By comparison, against a grayscale litter background the RGB mono spider elicited longer detection latencies than the control and RGB mono stimuli.

Responses of female S. ocreata to video playback of courting males against complex color and grayscale litter backgrounds. a Detection, measured as orientation latency in seconds; b Recognition, measured as receptivity score (sum of displays). Letters represent results of post-hoc Tukey tests; different letters represent significant differences of P < 0.05
For female S. ocreata receptivity responses (sum of displays), results of GLM showed significance overall (Χ25 = 53.732; P < 0.0001), with significance for spider exemplar color (Χ22 = 25.183; P < 0.0001) but not background (Χ21 = 1.333; P = 0. 248). The interaction of spider color x background again was significant (Χ22 = 71.061; P < 0.0001; Fig. 5b). Against the color litter background, the composite receptivity score (sum of displays) for the gray mono spider was very small, while responses to the control and RGB spider exemplars were equivalently large. Against the grayscale litter background, the RGB mono spider evoked very little receptivity, whereas the responses to the control and gray mono stimuli were similarly large.
Social eavesdroppers (male S. ocreata)
Eavesdropping male S. ocreata (Fig. 6) show a somewhat different response pattern than females. GLM analysis of detection latency (Fig. 6a) showed significance overall (Whole model: Χ25 = 2294.346; P < 0.0001), as well as for spider color (Χ25 = 1686.665; P < 0.0001), background (Χ21 = 7.625; P = 0.0058) and the interaction of spider color x background (Χ22 = 514.957; P < 0.0001).

Responses of eavesdropping male S. ocreata (social eavesdroppers) to video playback of courting males against complex color and grayscale litter backgrounds. a Detection, measured as orientation latency; b Recognition, measured as mean number of tapping bouts. Letters represent results of post-hoc Tukey tests; different letters represent significant differences of P < 0.05
Results of a GLM analysis of eavesdropping male tapping bouts (Fig. 6b) were similar and showed significance overall (Χ25 = 290.572; P < 0.0001) as well as spider color (Χ22 = 39.113; P < 0.0001), but not video background (Χ22 = 1.892; P = 0.169). However, the interaction of spider color x background was significant (Χ25 = 247.514; P < 0.0001), revealing the differences in patterns of eavesdropping male responses. Whereas the control spider stimulus elicited short orientation times when viewed both against the color and grayscale backgrounds (Fig. 6a), the RGB mono and gray mono exemplars appeared to be more difficult to detect against both backgrounds. Recognition, as evidenced by courtship behavior (tapping), showed dissimilar patterns to detection (Fig. 6b; see below).
Comparison of female and male S. ocreata responses
Stimulus Detection. In females, the control and RGB mono stimuli elicited significantly shorter orientation latencies than did the gray mono stimulus (Fig. 5a). In eavesdropping males the control stimulus evoked significantly shorter orientation latency than did the gray mono stimulus, while the RGB mono was not significantly different from either the control or gray stimuli (Fig. 6a). Against a grayscale litter background, orientation latencies to the three exemplar types did not differ significantly for females (Fig. 5a), whereas latencies were significantly shorter in response to the control stimulus than to the RGB stimulus for males (Fig. 6a). Stimulus Recognition. Females showed strong, equivalent recognition (receptivity) to all stimuli except the gray mono exemplar against the color litter background and the RGB mono exemplar against the grayscale background (Fig. 5b). By contrast, eavesdropping males exhibited recognition responses (tapping bouts) that are most interpretable in terms of stimulus conspicuousness (or lack thereof): few tapping bouts were elicited by the RGB mono exemplar against the color litter background and by the gray mono exemplar against the gray litter background; other exemplars evoked intermediate numbers of tapping bouts (Fig. 6b).
Interceptive eavesdroppers (predators)
Jumping spiders and wolf spider predators showed significant differences in the frequency of orientation (χ2 = 7.041; P = 0.008) and approach/attack (χ2 = 7.656; P = 0.0057) behaviors, so we analyzed each species separately. Orientation latency (secs) varied significantly for each predator species depending on color of the spider stimulus and the background against which it was displayed.
Wolf spiders (Rabidosa)
For Rabidosa wolf spiders, orientation latency (secs) varied significantly with spider color and background (Fig. 6a). Results of a GLM analysis showed significance overall (Whole model: Χ25 = 629.899; P < 0.0001), as well as for spider color (Χ22 = 555.780; P < 0.0001), background (Χ21 = 46.822; P < 0.0001) and the interaction of spider color x background (Χ22 = 37.444; P < 0.0001).
For Rabidosa approach/attack latency (Fig. 7b), GLM results were significant overall (Whole model: Χ25 = 852.853; P < 0.0001), as well as for spider color (Χ22 = 771.139; P < 0.0001), background (Χ21 = 60.755; P < 0.0001) and the interaction of spider color x background (Χ22 = 61.912; P < 0.0001).
Jumping spiders (Phidippus)
For Phidippus jumping spiders, a GLM analysis of latency to orient (Fig. 8a) showed significance overall (Whole model: Χ25 = 425.408; P < 0.0001), as well as for spider color (Χ22 = 297.722; P < 0.0001), background (Χ21 = 98.698; P < 0.0001), and for the interaction of spider color x background (Χ22 = 52.229; P < 0.0001). For Phidippus approach/attack latency (Fig. 8b), GLM results were similar to the detection results (Whole model: Χ25 = 253.559; P < 0.0001), as well as for spider color (Χ22 = 83.176; P < 0.0001), background (Χ21 = 112.454; P < 0.0001) and the interaction of spider color x background (Χ22 = 72.824; P < 0.0001).

Responses of eavesdropping wolf spider predators (Rabidosa rabida) to video playback of courting males against complex color and grayscale litter backgrounds. a Detection, measured as orient latency in seconds; a Recognition, measured as latency to attack in seconds. Letters represent results of post-hoc Tukey tests; different letters represent significant differences of P < 0.05

Responses of eavesdropping jumping spider predators (Phidippus clarus) to video playback of courting males against complex color and grayscale litter backgrounds. a Detection, measured as orient latency in seconds; b Recognition, measured as latency to attack in seconds. Letters represent results of post-hoc Tukey tests; different letters represent significant differences of P < 0.05
Toads (A. americanus)
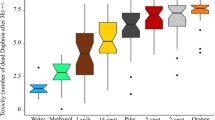
A GLM analysis showed that latency of visual detection (optomotor response) by toads of a courting male S. ocreata video (Fig. 9), was significant overall (Whole model: Χ25 = 17.817; P < 0.0001). Individual treatment effects were not significant (spider color: Χ22 = 1.090; P = 0.579; background: Χ21 = 2.000; P < 0.157), although the interaction term was significant (spider color x background: Χ22 = 10.479; P < 0.0053).

Visual detection of a courting male stimulus by toads, as indicated by a head tilt / orient response. Letters represent results of post-hoc Tukey tests; different letters represent significant differences of P < 0.05
Against the color litter background, the gray mono spider elicited the shortest orientation latency from toad subjects, whereas detection time for the control and RGB mono spider exemplars did not differ significantly. Against the grayscale litter background, orientation latency was much longer to the gray mono stimulus than to the control and RGB mono stimulus, which were not significantly different.
Discussion
Results of this study support our general hypothesis that visual contrast between a spider and its background will affect receiver detection and recognition and will be strongly influenced by viewer spectral sensitivity. Every statistical test of receiver response included a significant interaction term between spider coloration and background coloration – an outcome underscoring the importance of visual contrast in stimulus detection and recognition. As a consequence, selection by different types of receivers on male coloration and courtship behavior may act together or in opposition on animal color and motion signals (Earley and Dugatkin 2005; Thery and Casas 2009).
Fit of behavioral results to model predictions
Results of our behavioral analyses showed that the fit of receiver responses to predictions of the visual contrast models varied dramatically, both by type of receiver and by stimulus-background contrast (Table 1). Although behavioral responses of some receivers matched the predictions of the contrast models better than did others, we note that our Spearman rank correlation index summarizes that fit into a single value. Thus, unanticipated levels of response to one or two of the six experimental spider exemplar/background combinations can have a large impact on the behavior-contrast correlations (see below).
For instance, certain unexpected behavioral responses in our experiments may be explained by recent studies demonstrating that the magnitude of chromatic and achromatic contrast between a signal and its background has direct effects on color signal detectability and discriminability (Davis et al. 2022; Caves et al. 2024). Interestingly, although increasing contrast with the background makes a color patch (i.e., spider exemplars in our study) more detectable, greater stimulus-background contrast makes discriminating color variation more difficult.
Among the spider exemplars, the naturally-colored control spider was one of the most easily detected stimuli against each background. For example, leg tufts and dark colors of male S. ocreata increase both chromatic and achromatic contrast against all backgrounds, as evidenced here and in previous studies (Clark et al. 2011). Even with the limited color vision of lycosids (DeVoe 1972), contrast may be important for recognition, and control spiders exhibited maximum chromatic contrast values against both experimental backgrounds.
Against a color background, the gray mono and RGB mono spider exemplars are predicted to be more difficult to detect (i.e., lower chromatic contrast) than the control spider (Fig. 4a). Against a grayscale background, however, the control and RGB mono spider exemplars should be more readily detectable than the gray mono spider, which exhibits substantially lower chromatic contrast (Fig. 4a). For achromatic contrast, only the control spider stimulus is predicted to be easily detectable against both color and grayscale litter backgrounds (Fig. 4b).
Intuitively, the gray mono spider exemplar should be very difficult to detect against a grayscale background – a supposition that is verified in the chromatic and achromatic contrast models.
Intended receivers (female S. ocreata)
Although detection responses of intended receivers (female S. ocreata) showed no fit to the predictions of the chromatic model (Table 1a), the absence of fit arose from two seemingly anomalous responses: a brief orientation latency to the RGB mono stimulus against the color litter background and a similarly short orientation latency to the gray mono stimulus against the grayscale litter background (Figs. 4a and 5a). Detection responses exhibited a fair match to the achromatic contrast model, with the Spearman Rho value approaching the “good fit” range (Table 1a). Recognition responses also exhibited a fair fit to the chromatic model, in which receptivity to the gray mono stimulus against the color litter background were weaker than predicted, and receptivity to the gray mono stimulus against the grayscale litter background was stronger than predicted (Table 1b; Figs. 4a and 5b). By comparison, female S. ocreata receptivity scores showed an excellent (and statistically significant fit) to the achromatic model, despite relatively strong responses to the RGB mono stimulus against the color litter background and gray mono stimulus against the grayscale litter background (Table 1b; Figs. 4b and 5b). We can offer a partial explanation here by speculating that movement of the RGB mono exemplar between the different hues of the color leaf litter background might be more detectable and recognizable than we predicted. Likewise, the higher receptivity response to the gray mono spider against a grayscale background, which seems counter-intuitive, is also hard to explain. Interestingly, motion has been shown to readily reveal the presence of animal color patterns that are cryptic against their visual backgrounds (Ioannou and Krause 2009; Hall et al. 2013). In our experiment, a monochromatic gray stimulus moving against a grayscale (i.e., not monochromatic) complex visual background may be more visible than we had anticipated. However, were this explanation generally applicable to our results, we would anticipate having observed brief detection times for the gray stimulus against the grayscale background in receivers other than female S. ocreata (which was not the case).
Social eavesdroppers (male S. ocreata)
Detection (orientation latency) showed a poor fit to predictions of the chromatic contrast model for male S. ocreata (Table 1a). This low level of fit appears to stem from male responses to the gray mono stimulus against the color litter background, as well as to the gray mono and RGB mono stimuli against the grayscale background (Figs. 4a and 6a). By comparison, male detection had a very good (and statistically significant) fit to predictions of the achromatic contrast visual model (Table 1a). Regarding stimulus recognition (tapping bouts), male responses showed a good fit to the chromatic model (Fig. 6a) but poor fit to the achromatic model (Table 1b), in which responses to a number of stimulus/background combinations ran counter to prediction (Fig. 6b).
Eavesdropping predators
For eavesdropping spider predators – wolf spiders (Rabidosa) and jumping spiders (Phidippus) – fit to the chromatic and achromatic contrast models in detection was better overall than for S. ocreata: whereas Phidippus showed a good fit to predictions from both contrast models, Rabidosa had a very good (and significant) fit to the chromatic model, and an excellent (and significant) fit to the achromatic model (Table 1a; Figs. 4, 7a and 8a and a). Note that despite the better overall fit of these predators’ detection responses to the visual models of contrast (as compared to S. ocreata), actual orientation latencies were much longer than in S. ocreata, especially for Phidippus (Figs. 7a and 8a and a). The fit of recognition responses for Rabidosa was identical to their detection responses: a very good (and significant) fit to the chromatic model, and an excellent (and significant) fit to the achromatic model (Table 1b). Recognition responses in Phidippus fit predictions of the contrast models less well than did their detection responses (i.e., poor for the chromatic model and fair for the achromatic model: Table 1b). However, the color of the prey spider stimulus may have mattered more for Phidippus than for our wolf spider receivers. Jumping spiders have well developed color vision (Zureck et al. 2015), and P. clarus responded more quickly to control and RGB mono spiders color than to gray mono exemplars in all stimulus/background combinations except for recognition (attack latency) against the grayscale litter background.
The vertebrate eavesdropper - the predatory toad, Anaxyrus, showed a different overall pattern of responses to our stimuli. Given that the optomotor responses (head tilt) we measured indicate both detection and recognition, we considered only orientation latency (Table 1a). Like Phidippus, Anaxyrus exhibited a good fit to the chromatic contrast model (Table 1a): relatively brief responses latencies to stimuli with high and moderate contrast, and a longer response latency to the stimulus with very low contrast (gray stimulus against the grayscale background; Figs. 4a and 9). Interestingly, most orientation latency values were much shorter than those of the invertebrate predators. Unlike Phidippus, however, orientation latencies for Anaxyrus fell into the “no fit” category for the achromatic contrast model (Table 1a). Importantly, despite a lack of overall fit, not all responses of our toad subjects ran counter to predictions of the achromatic contrast model. Specifically, Anaxyrus exhibited (1) brief orientation latencies to the control stimulus, which has strong (negative) achromatic contrast against the color and grayscale backgrounds, and (2) exhibited comparatively long orientation latencies to the low achromatic contrast of the gray stimulus against the grayscale background (Figs. 4b and 9).
It is not surprising that our representative vertebrate predator responded differently from our arthropod viewers. For predatory toads, previous studies have shown that latency in visual detection of a courting male stimulus varied significantly with the background against which they were viewed (Roberts and Uetz 2008; Clark et al. 2016). These results suggest both spider color and background complexity affect detection, and that courting male wolf spiders may face higher predation risk from visually acute toads.
Together, results of our experiments suggest that both chromatic and achromatic contrast are likely explanations for differences in the behavioral responses of intended and unintended receivers. Consequently, males must balance fitness gains from conspicuous courtship behaviors and visual background contrast with losses from detection by both social eavesdroppers (conspecific competitors) and interceptive eavesdroppers (predators). To the human eye, visibility of the RGB and gray mono spiders against the color and grayscale backgrounds (respectively) differs from what behavioral results for non-human receivers in our experimental treatments might suggest, challenging some of our initial assumptions (Rosenthal 2007). For example, against the color litter background, the visual contrast of RGB mono and gray mono spiders appears roughly equivalent to us. Likewise, the appearance of the RGB mono spider against the grayscale litter background seems much more obvious to human vision than would be expected based on the responses of non-human receivers. Even in light of the fact that spectral sensitivities of spider photoreceptors in our visual models differ from those of normal trichromatic human vision, these perceptual differences are intriguing.
One issue that we have not considered is detection of visual courtship displays against a moving background, as has been examined in studies of signal detection in other organisms (Fleishman 1986; Peters et al. 2008; Ioannou and Krause 2009; Bian et al. 2019; Tan and Elgar 2021). For example, the movement of leaves and branches in wind creates a background against which motion signals of lizards are difficult to detect (Peters and Evans 2003; Peters 2008), resulting in directional selection on signal morphology (Fleishman et al. 1986) or real-time compensation mechanisms (Ord et al. 2007). To be fair, there are issues of scale, and background motion of leaves on the forest floor is limited except in the winter, when winds are greatest (Boerner and Kooser 1989). However, even small amounts of wind movement within leaf litter, along with changing light and shadows might be important to tiny animals like spiders (Théry 2001; Matchette et al. 2019; Aguilar-Arguello et al. 2021). In addition, for wolf spiders in the genus Schizocosa, where species use vibratory and visual communication, the complexity of the leaf litter environment has an influence on efficacy of multimodal signals (Elias et al. 2010; Uetz et al. 2013).
Understanding animal communication and its evolution requires consideration of inter-relationships among senders, signals, and receivers, as well as the social and physical environments in which communication occurs (Endler 1992, 1993; Searcy and Nowicki 2005; Hebets et al. 2016; Rosenthal et al. 2018). In the present work we have examined the effects of viewer spectral sensitivity and visual contrast on the perception of Schizocosa wolf spiders as potential prey or mates, yet much remains to be explored in studies of animal signal detection and recognition. Additional insights might be gained from studies of receiver responses to chromatic and achromatic contrast in other species that exhibit striking coloration and movement in courtship (Rosenthal 2007; Stevens and Merilaita 2009b; Maiditsch and Ladich 2022; White et al. 2022). Another possibility is to investigate how recently developed approaches to animal communication can address these questions (Rosenthal 2007; Hebets et al. 2016; Hoke et al. 2021; Uetz et al. 2022; Caves et al. 2024). Ultimately, further exploration is required to determine how sexual and natural selection might target different features of male appearance during courtship signaling.
Data availability
The datasets generated and/or analyzed during the current study are available from the authors upon request and are archived in Mendeley Data: https://data.mendeley.com/datasets/yhs8yv3wj7/1.
References
Aguilar-Arguello S, Taylor AH, Nelson XJ (2021) Jumping spiders attend to information from multiple modalities when preparing to jump. Anim Behav 171:99–109. https://www.sciencedirect.com/science/article/abs/pii/S0003347220303353
Aho AC, Donner K, Helenius S, Olesen-Larsen L, Reuter T (1993) Visual performance of the toad (Bufo bufo) at low light levels: retinal ganglion cell responses and prey-catching accuracy. J Comp Physiol A 172:671–682. https://doi.org/10.1007/BF00195393
Beck A, Ewert JP (1979) Prey selection by toads (Bufo bufo L.) in response to configurational stimuli moved in the visual field z,y-coordinates. J Comp Physiol A 129:207–209. https://link.springer.com/article/10.1007/BF00657655
Bernal XE, Page RA (2022) Tactics of evasion: strategies used by signallers to deter eavesdropping enemies from exploiting communication systems. Biol Rev 98:222–242. https://doi.org/10.1111/brv.12904
Bian X, Chandler T, Pinilla A, Peters RA (2019) Now you see me, now you don’t: environmental conditions, signaler behavior, and receiver response thresholds interact to determine the efficacy of a movement-based animal signal. Front Ecol Evol 7:130. https://doi.org/10.3389/fevo.2019.00130/full
Boerner RE, Kooser JG (1989) Leaf litter redistribution among forest patches within an Allegheny Plateau watershed. Landsc Ecol 2:81–92. https://link.springer.com/article/10.1007/BF00137152
Bond AB, Kamil AC (2006) Spatial heterogeneity, predator cognition, and the evolution of color polymorphism in virtual prey. Proc Natl Acad Sci USA 103:3214–3219. https://doi.org/10.1073/pnas.0509963103
Caves EM, Davis AL, Nowicki S, Johnsen S (2024) Backgrounds and the evolution of visual signals. Trends Ecol Evol. 39:188–198. https://doi.org/10.1016/j.tree.2023.09.006
Choi N, Adams M, Fowler-Finn K, Knowlton E, Rosenthal M, Rundus A, Santer RD, Wilgers D, Hebets EA (2022) Increased signal complexity is associated with increased mating success. Biol Lett 18:20220052. https://doi.org/10.1098/rsbl.2022.0052
Clark DL, Uetz GW (1990) Video image recognition by the jumping spider Maevia inclemens (Araneae: Salticidae). Anim Behav 40:884–890. https://www.sciencedirect.com/science/article/abs/pii/S000334720580990X
Clark DL, Roberts JA, Rector M, Uetz GW (2011) Spectral reflectance properties and visual communication in the wolf spider, Schizocosa ocreata (Hentz): simultaneous crypsis and background contrast in visual signals. Behav Ecol Sociobiol 65:1237–1247. https://link.springer.com/article/10.1007/s00265-011-1137-y
Clark DL, Roberts JA, Uetz GW (2012) Eavesdropping and signal matching in visual courtship displays of spiders. Biol Lett 8:375–378. https://doi.org/10.1098/rsbl.2011.1096
Clark DL, Kizer C, Sabovodny G, Hollenberg A, Roberts JA, Uetz GW (2015) The role of social experience in eavesdropping by male wolf spiders (Lycosidae). Anim Behav 106: 89–97. https://www.sciencedirect.com/science/article/pii/S0003347215001803?casa_token=zMIMdrL7HwkAAAAA:gxCyRSHbofPUz0Qin03fOLHHtae3r8MEeLa3Db-hIo8JMKg-U8jwmHusnNTg9DUXcgS4xWw1ai8g
Clark DL, Kizer C, Karson A, Roberts JA, Uetz GW (2016) Risky courtship: background contrast, ornamentation, and display behavior of wolf spiders affect visual detection by vertebrate predators. Ethol 122:1–12. https://onlinelibrary.wiley.com/doi/abs/
Clarke RD (1974) Food habits of toads, genus Bufo (Amphibia: Bufonidae). Amer Midl Natur 91:140–147. https://www.jstor.org/stable/2424517
Cummings ME, Endler JA (2018) 25 years of sensory drive: the evidence and its watery bias. Curr Zool 64:471–484. https://academic.oup.com/cz/article/64/4/471/5034003
Cummings ME, Jordão JM, Cronin TW, Oliveira RF (2008) Visual ecology of the fiddler crab, Uca tangeri: effects of sex, viewer and background on conspicuousness. Anim Behav 75: 175–188. https://www.sciencedirect.com/science/article/pii/S0003347207003569?casa_token=8fQzu-jSwMEAAAAA:MIM02G-W2S8Y55TkvSVNEmJV-Sj90KdqqJM6mHY72fD4xAnp4d8zcZ7Hg3HktzF0uiB5Ab4K36WO
Dabelsteen T (2005) Public, private, or anonymous? Facilitating and countering eavesdropping. Pp. 38–62. In McGregor P K Animal Communication Networks. Cambridge Univ. Press, Cambridge, USA. https://www.cambridge.org/core/books/abs/animal-communication-networks/public-private-or-anonymous-facilitating-and-countering-eavesdropping/E715EED98235040060868ACA868C8857
Davis A, Zipple MN, Diaz D, Peters S, Nowicki S, Johnsen S (2022) Influence of visual background on discrimination of signal-relevant colours in zebra finches (Taeniopygia guttata). Proc R Soc Lond B 1976:1–8. https://doi.org/10.1098/rspb.2022.0756
DeVoe RD (1972) Dual sensitivities of cells in wolf spider eyes at ultraviolet and visible wavelengths of light. J Gen Physiol 59:247–269. https://rupress.org/jgp/article-pdf/59/3/247/1809493/247.pdf
DeVoe RD (1975) Ultraviolet and green receptors in principle eyes of jumping spiders. J Gen Physiol 66:193–207. https://rupress.org/jgp/article-pdf/66/2/193/1810525/193.pdf
DeVoe RD, Small RJ, Zvargulis JE (1969) Spectral sensitivities of wolf spider eyes. J Gen Physiol 54:1–32. https://rupress.org/jgp/article-pdf/54/1/1/1808679/1.pdf
Dimitrova M, Merilaita S (2009) Prey concealment: visual background complexity and prey contrast distribution. Behav Ecol 21:176–181. https://academic.oup.com/beheco/article/21/1/176/181694
Dugatkin LA (1992) Sexual selection and imitation: females copy the mate choice of others. Am Nat 139:1384–1389. https://www.journals.uchicago.edu/doi/abs/10.1086/285392
Dugatkin LA, Godin JGJ (1993) Female mate copying in the guppy (Poecilia reticulata): age-dependent effects. Behav Ecol 4:289–292. https://academic.oup.com/beheco/article-abstract/4/4/289/206631
Elias DO, Mason AC, Hebets EA (2010) A signal-substrate match in the substrate-borne component of a multimodal courtship display. Curr Zool 56:370–378. https://academic.oup.com/cz/article/56/3/370/5559827
Endler JA (1990) On the measurement and classification of colour in studies of animal colour patterns. Biol J Linn Soc 41:315–352. https://academic.oup.com/biolinnean/article-abstract/41/4/315/2654288
Endler JA (1992) Signals, signal conditions, and the direction of evolution. Am Nat 139:S125–S153. https://www.journals.uchicago.edu/doi/abs/10.1086/285308
Endler JA (1993) The color of light in forests and its implications. Ecol Monogr 63:1–27. https://esajournals.onlinelibrary.wiley.com/doi/abs/10.2307/2937121
Endler JA (2006) Disruptive and cryptic coloration. Proc R Soc Lond B 273:2425–2426. https://doi.org/10.1098/rspb.2006.3650
Endler JA, Mielke JR (2005) Comparing entire colour patterns as birds see them. Biol J Linn Soc 86:405–431. https://academic.oup.com/biolinnean/article/86/4/405/2706048
Fleishman LJ (1986) Motion detection in the presence and absence of background motion in an Anolis lizard. J Comp Physiol A 159:711–720. https://link.springer.com/article/10.1007/BF00612043
Fleishman LJ, Persons M (2001) The influence of stimulus and background colour on signal visibility in the lizard Anolis cristatellus. J Exp Biol 204:1559–1575. https://journals.biologists.com/jeb/article/204/9/1559/8818/The-Influence-of-Stimulus-and-Background-Colour-on
Fleishman LJ, Perez CW, Yeo AI, Cummings KJ, Dick S, Almonte E (2016a) Perceptual distance between colored stimuli in the lizard Anolis sagrei: comparing visual system models to empirical results. Behav Ecol Sociobiol 70:541–555. https://doi.org/10.1007/s00265-016-2072-8
Fleishman L, Ogas B, Steinberg D, Leal M (2016b) Why do Anolis dewlaps glow? An analysis of a translucent visual signal. Funct Ecol 30:345–355. https://besjournals.onlinelibraryhttps://doi.org/10.1111/1365-2435.12502
Fleishman LJ, Perez-Martinez CA, Leal M (2022) Can sensory drive explain the evolution of visual signal diversity in terrestrial species? A test with Anolis lizards. Am Nat 200:236–249. https://www.journals.uchicago.edu/doi/full/10.1086/720267
Galloway JA, Green SD, Stevens M, Kelley LA (2020) Finding a signal hidden among noise: how can predators overcome camouflage strategies? Phil Trans R Soc B 375:20190478. https://doi.org/10.1098/rstb.2019.0478
Glenszczyk M, Outomuro D, Gregorič M, Kralj-Fišer S, Schneider J, Nilsson DE, Morehouse NI, Tedore C (2022) The jumping spider Saitis barbipes lacks a red photoreceptor to see its own sexually dimorphic red coloration. Sci Nat 109:6. https://doi.org/10.1007/s00114-021-01774-6
Govardovskii VI, Fyhrquist N, Reuter T, Kuzmin DG, Donner K (2000) In search of the visual pigment template. Vis Neurosci 17:509–528. https://www.cambridge.org/core/journals/visual-neuroscience/article/abs/in-search-of-the-visual-pigment-template/A4738E821720092B7F5A233C4AB4962B
Hall JR, Cuthill IC, Baddeley R, Shohet AJ, Scott-Samuel NE (2013) Camouflage, detection and identification of moving targets. Proc R Soc Lond B 280:1–7. https://doi.org/10.1098/rspb.2013.0064
Haynes KF, Yeargan KV (1999) Exploitation of intraspecific communication systems: illicit signalers and receivers. Ann Entomol Soc Am 92:960–970. https://academic.oup.com/aesa/article-abstract/92/6/960/93493
Hebets EA, Papaj DR (2005) Complex signal function: developing a framework of testable hypotheses. Behav Ecol Sociobiol 57:197–214. https://link.springer.com/article/10.1007/s00265-004-0865-7
Hebets EA, Barron AB, Balakrishnan CN, Hauber ME, Mason PH, Hoke KL (2016) A systems approach to animal communication. Proc R Soc Lond B 283:20152889. https://doi.org/10.1098/rspb.2015.2889
Hoke KL, Hensley N, Kanwal JK, Wasserman S, Morehouse NI (2021) Spatio-temporal dynamics in animal communication: A special issue arising from a unique workshop-symposium model. Integr Comp Biol 61:783–786. https://academic.oup.com/icb/article/61/3/783/6313282
Ioannou CC, Krause J (2009) Interactions between background matching and motion during visual detection can explain why cryptic animals keep still. Biol Lett 5:191–193. https://doi.org/10.1098/rsbl.2008.0758
Johnstone RA (2001) Eavesdropping and animal conflict. Proc Natl Acad Sci USA 98:9177–9180. https://www.pnas.org/doi/abs/10.1073/pnas.161058798
Jones BC, DuVal EH (2019) Mechanisms of social influence: a meta-analysis of the effects of social information on female mate choice decisions. Front Ecol Evol 7:390. https://doi.org/10.3389/fevo.2019.00390
Koga T, Backwell PRY, Murai M, Kasuya E (2001) Male-biased predation of a fiddler crab. Anim Behav 62:201–207. https://www.sciencedirect.com/science/article/abs/pii/S0003347201917403
Kotiaho J, Alatalo RV, Mappes J, Parri S, Rivero A (1998) Male mating success and risk of predation in a wolf spider: a balance between sexual and natural selection? J Anim Ecol 67:287–291. https://besjournals.onlinelibrary.wiley.com/doi/abs/https://doi.org/10.1046/j.1365-2656.1998.00192.x
Land MF, Fernald RD (1992) The evolution of eyes. Ann Rev Neurosci 15:1–29. https://www.annualreviews.org/doi/abs/10.1146/annurev.ne.15.030192.000245?journalCode=neuro
Land MF, Nilsson DE (2002) Animal eyes. Oxford University Press, Oxford. https://books.google.com/books?hl=enamplr=&id=uXSK6hDKFC0C&oi=fnd&pg=PP1&dq=Land+Nilsson+2002+Animal+eyes&ots=dMEr4zjxHM&sig=-9fyGMhKUVvV6Yer8L7CiTwYIZU#v=onepage&q=LandNilsson2002Animaleyes&f=false
Lim MLM, Li D (2004) Courtship and male–male agonistic behaviour of Cosmophasis umbratica Simon, an ornate jumping spider (Araneae: Salticidae) from Singapore. RaZes B Zool 52:435–448. https://www.researchgate.net/profile/Daiqin-Li/publication/253321808_Courtship_and_male-male_agonistic_behaviour_of_Cosmophasis_umbratica_Simon_an_ornate_jumping_spider_Araneae_Salticidae_from_Singapore/links/0046352312cad56cbe000000/Courtship-and-male-male-agonistic-behaviour-of-Cosmophasis-umbratica-Simon-an-ornate-jumping-spider-Araneae-Salticidae-from-Singapore.pdf
Lim LM, Li D (2006) Extreme ultraviolet sexual dimorphism in jumping spiders (Araneae: Salticidae). Soc 89:397–406. https://academic.oup.com/biolinnean/article/89/3/397/2691690
Macedonia JM, Brandt JM, Clark DL (2002) Sexual dichromatism and differential conspicuousness in two populations of the common collard lizard (Crotaphytus collaris) from Utah and New Mexico, USA. Biol J Linn Soc 77:67–85. https://academic.oup.com/biolinnean/article/77/1/67/2639806
Macedonia JM, Lappin AK, Loew ER, McGuire JA, Hamilton PS, Plasman M, Brandt Y, Lemos-Espinal JA, Kemp DJ (2009) Conspicuousness of Dickerson’s collared lizard (Crotaphytus dickersonae) through the eyes of conspecifics and predators. Biol J Linn Soc 97:749–765. https://academic.oup.com/biolinnean/article/97/4/749/2447941
Maiditsch IP, Ladich F (2022) Acoustic and visual adaptations to predation risk: a predator affects communication in vocal female fish. Curr Zool. 2022 68:149–157. https://doi.org/10.1093/cz/zoab049
Marshall KL, Philpot KE, Stevens M (2015) Conspicuous male coloration impairs survival against avian predators in Aegean wall lizards, Podarcis Erhardii. Ecol Evol 5:4115–4131. https://onlinelibrary.wiley.com/doi/abs/https://doi.org/10.1002/ece3.1650
Matchette SR, Cuthill IC, Scott-Samuel NE (2019) Dappled light disrupts prey detection by masking movement. Anim Behav 155:89–95. https://www.sciencedirect.com/science/article/pii/S0003347219302143
McClintock WJ, Uetz GW (1996) Female choice and pre-existing bias: visual cues during courtship in two Schizocosa wolf spiders (Araneae: Lycosidae). Anim Behav 52:167–181. https://www.sciencedirect.com/science/article/abs/pii/S0003347296901621
McGregor PK (2005) Animal Communication Networks. Cambridge Univ. Press, Cambridge, p 657. https://books.google.com/books?hl=en&lr=&id=B58DSFCaQpwC&oi=fnd&pg=PR8&dq=McGregor+PK+(2005)+Animal+Communication+Networks&ots=vRMi1pCis0&sig=CSV5ESA2xKHOcnn8f_AV7ylU690#v=onepage&q=McGregor%20PK%20(2005)%20Animal%20Communication%20Networks&f=false.
Merilaita S, Scott-Samuel NE, Cuthill CE (2017) How camouflage works. Phil Trans R Soc B 372:20160341. https://doi.org/10.1098/rstb.2016.0341
Ord TJ, Peters RA, Clucas B, Stamps JA (2007) Lizards speed up visual displays in noisy motion habitats. Proc R Soc Lond B 274:1057–1062. https://doi.org/10.1098/rspb.2006.0263
Osorio D, Vorobyev M (2005) Photoreceptor spectral sensitivities in terrestrial animals: adaptations for luminance and colour vision. Proc R Soc Lond B 272:1745–1752. https://doi.org/10.1098/rspb.2005.3156
Partan SR, Marler P (2005) Issues in the classification of multimodal communication signals. Am Nat 166:231–245. https://www.journals.uchicago.edu/doi/abs/10.1086/431246
Peake TM (2005) Eavesdropping in communication networks. pp.13–37. In: McGregor PK (ed) Animal Communication Networks. Cambridge Univ, Cambridge USA
Peters RA (2008) Environmental motion delays the detection of movement-based signals. Biol Lett 4:2–5. https://doi.org/10.1098/rsbl.2007.0422
Peters RA, Evans CS (2003) Design of the Jacky dragon visual display: signal and noise characteristics in a complex moving environment. J Comp Physiol A 189:447–459. https://link.springer.com/article/10.1007/s00359-003-0423-1
Peters RA, Hemmi J, Zeil J (2008) Image motion environments: background noise for movement-based animal signals. J Comp Physiol A 194:441–456. https://link.springer.com/article/10.1007/s00359-008-0317-3
Postema EG, Lippey MK, Armstrong-Ingram T (2023) Color under pressure: how multiple factors shape defensive coloration. Behav Ecol 34:1–13. https://doi.org/10.1093/beheco/arac056
Price N, Green S, Troscianko J, Tregenza T, Stevens M (2019) Background matching and disruptive coloration as habitat-specific strategies for camouflage. Nature: Sci Rep 9:7840. https://www.nature.com/articles/s41598-019-44349-2
Roberts JA, Uetz GW (2008) Discrimination of variation in a male signaling trait affects optomotor response in visual predators. Ethol 114: 557–563. https://onlinelibrary.wiley.com/doi/abs/10.1111/j.1439-0310.2008.01491.x
Roberts JA, Taylor PW, Uetz GW (2007) Consequences of complex courtship display: Predator detection of multimodal signaling. Behav Ecol 18:236–240. https://academic.oup.com/beheco/article/18/1/236/210734
Rosenthal GG (2007) Spatiotemporal dimensions of visual signals in animal communication. Ann Rev Ecol Evol Syst 38:155–178. https://doi.org/10.1146/annurev.ecolsys.38.091206.095745
Rosenthal GG, Flores-Martinez TY, Garcia de Leon F, Ryan M (2001) Shared preferences by predators and females for male ornamental traits in swordtails. Am Nat 158:146–154. https://www.journals.uchicago.edu/doi/abs/10.1086/321309
Rosenthal MF, Wilkins MR, Shizuka D, Hebets EA (2018) Dynamic changes in display architecture and function across environments revealed by a systems approach to animal communication. Evolution 72:1134–1145. https://academic.oup.com/evolut/article/72/5/1134/6726688
Roster NO, Clark DL, Gillingham JC (1995) Prey catching behavior in frogs and toads using video-simulated prey. Copeia 1995:496–498. https://www.jstor.org/stable/1446921
Rovner JS (1993) Visually mediated responses in the lycosid spider Rabidosa rabida: the roles of different pairs of eyes. Mem Qld Mus 33:635–638
Rovner JS (1996) Conspecific interactions in the lycosid spider Rabidosa rabida: the roles of different senses. J Arachnol 24:16–23. https://www.jstor.org/stable/3705862
Rowe ZW, Scott-Samuel NE, Cuthill IC (2024) How background complexity impairs target detection. Anim Behav 210:99–111.
Rubi TL, Clark DL, Keller JS, Uetz GW (2019) Courtship behavior and coloration influence conspicuousness of wolf spiders (Schizocosa ocreata (Hentz)) to avian predators. Behav Proc 162:215–220. https://doi.org/10.1016/j.beproc.2018.12.023
Searcy WA, Nowicki S (2005) The evolution of animal communication: reliability and deception in signaling systems. Princeton Univ, Princeton, NJ, p 270. https://doi.org/10.1515/9781400835720
Stevens M, Cuthill IC (2006) Disruptive coloration, crypsis and edge detection in early visual processing. Proc R Soc Lond B 273:2141–2147. https://doi.org/10.1098/rspb.2006.3556
Stevens M, Merilaita S (2009a) Animal camouflage: current issues and new perspectives. Phil Trans R Soc B 364:423–427. https://doi.org/10.1098/rstb.2008.0217
Stevens M, Merilaita S (2009b) Defining disruptive coloration and distinguishing its functions. Phil Trans R Soc B 364:481–488. https://doi.org/10.1098/rstb.2008.0216
Stevens M, Cuthill IC, Windsor AMM, Walker HJ (2006) Disruptive contrast in animal camouflage. Proc R Soc Lond B 273:2433–2438. https://doi.org/10.1098/rspb.2006.3614
Stevens M, Yule DH, Ruxton GD (2008) Dazzle coloration and prey movement. Proc R Soc Lond B 275:2639–2643. https://doi.org/10.1098/rspb.2008.0877
Stoffer B, Uetz GW (2015) The effects of social experience with varying male availability on female preference in a wolf spider. Behav Ecol Sociobiol 69:927–937. https://link.springer.com/article/10.1007/s00265-015-1904-2
Tan EJ, Elgar MA (2021) Motion: enhancing signals and concealing cues. Biol Open 10:058762. https://journals.biologists.com/bio/article/10/8/bio058762/271863/Motion-enhancing-signals-and-concealing-cues
Théry M (2001) Forest light and its influence on habitat selection. In Tropical Forest Canopies: Ecology and Management: Proceedings of ESF Conference, Oxford University: 251–261. Springer, Netherlands. https://link.springer.com/chapter/10.1007/978-94-017-3606-0_20
Tibbetts EA, Wong E, Bonello S (2020) Wasps use social eavesdropping to learn about individual rivals. Curr Biol 30:3007–3010. https://www.sciencedirect.com/science/article/pii/S0960982220307405
Uetz GW, Clark DL, Roberts JA, Rector M (2011) Effect of visual background complexity and light level on detection of visual signals of male Schizocosa ocreata wolf spiders by female wolf spider receivers. Behav Ecol Sociobiol 65:753–761. https://doi.org/10.1007/s00265-010-1079-9
Uetz GW, Clark DL, Stoffer B (2022) On strengths and limitations of field, semi-natural captive, and laboratory study settings. In The Routledge International Handbook of Comparative Psychology. 34–47 Riley AR, d’Ettorre P, Freeberg T. https://doi.org/10.4324/9781003091868
Vorobyev M, Osorio D (1998) Receptor noise as a determinant of colour thresholds. Proc R Soc Lond B: Biol Sci 265:351–358. https://doi.org/10.1098/rspb.1998.0302
White TE, Latty T, Umbers KD (2022) The exploitation of sexual signals by predators: a meta-analysis. Proc R Soc Lond B 289:20220444. https://doi.org/10.1098/rspb.2022.0444
Wiley RH (2013) Signal detection, noise, and the evolution of communication. In Brumm H (ed) Animal communication and noise: 7–30. https://link.springer.com/chapter/10.1007/978-3-642-41494-7_2
Wiley RH (2017) How noise determines the evolution of communication. Anim Behav 124:307–313. https://www.sciencedirect.com/science/article/pii/S0003347216301403?casa_token=4bv9KIkWanAAAAAA:tOsg3LldRkwLzCf46PZE58rcWLLq5Z-Pcm_H7xhZ10_Mg9Spuc_EHf0JsMSnWdkgWeY0CRzjjPLR
Yamashita S, Tateda H (1976) Spectral sensitivities of jumping spider eyes. J Comp Physiol 105:29–41. https://link.springer.com/article/10.1007/BF01380051
Yovanovich CAM, Koskela SM, Nevala N, Kondrashev SL, Kelber A, Donner K (2017) The dual rod system of amphibians supports colour discrimination at the absolute visual threshold. Phil Trans R Soc B 372:20160066. https://doi.org/10.1098/rstb.2016.0066
Zuk M, Kolluru GR (1998) Exploitation of sexual signals by predators and parasitoids. Q Rev Biol 73:415–438. https://www.journals.uchicago.edu/doi/abs/10.1086/420412
Zurek DB, Cronin TW, Taylor LA, Byrne K, Sullivan ML, Morehouse NI (2015) Spectral filtering enables trichromatic vision in colorful jumping spiders. Curr Biol 25:R403–R404. https://www.sciencedirect.com/science/article/pii/S0960982215003486
Acknowledgements
This research was supported by National Science Foundation grants IOS-1026995 and DBI-1262863 (to G.W.U) and IOS1026817 (to D.L.C.), the UC Department of Biological Sciences and Alma College. We also thank the members of the Uetz and Clark labs for helping collect and care for the spiders. We are grateful to two anonymous reviewers whose comments have greatly improved this manuscript. Finally, thank you to the Cincinnati Nature Center for allowing the collection of spiders from their Rowe Woods property. We acknowledge with respect that our study areas are on forest land originally occupied by the Myaamia (Miami) and Shawandasse (Shawnee) nations prior to European invasion.
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception and design to varying degrees. Material preparation, data collection and analysis were performed by George W. Uetz, David L. Clark, Joseph Macedonia, Rachel Gilbert, Rebecca Wilson, and Jacqueline Mann. The first draft of the manuscript was written by George W. Uetz and David L. Clark and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare they have no financial or non-financial conflicts of interest that are directly or indirectly related to the work submitted for publication. We adhered to the ASAB/ABS Guidelines for the Use of Animals in Research (https://doi.org/10.1016/j.anbehav.2019.11.002). All applicable international, national, and/or institutional guidelines for the care and use of animals were followed. There are no institutional, local, state or federal government rules regarding animal care for invertebrates (spiders). Vertebrates (toads) were housed at Alma College, Alma Michigan and maintained according to a protocol approved by the Alma College Animal Care Committee (Protocol #A4162-01) (see text for details).
Additional information
Communicated by D. Kemp.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Uetz, G.W., Gilbert, R., Wilson, R. et al. Background contrast affects detection and recognition of courting wolf spiders by intended and unintended receivers. Behav Ecol Sociobiol 78, 43 (2024). https://doi.org/10.1007/s00265-024-03453-8
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00265-024-03453-8