
Abstract
Apex carnivores that rely primarily on predation play a central but complex role within scavenging ecology by potentially suppressing intra-guild competitors, but also facilitating them by providing a reliable supply of carrion. We investigated the competitive relationship between sympatric wolves (Canis lupus) and wolverines (Gulo gulo) in Norway across three seasons. We deployed remote cameras at fresh wolf kills (n = 29) and built Bayesian generalized linear mixed models (GLMMs) to explore the use of fresh wolf-killed prey by sympatric wolves and wolverines. Our results showed that wolves facilitated wolverines by providing scavenging opportunities. Biomass available from wolf kills was influenced by seasonal wolf prey preference and group size. Wolverines visited 100% of wolf kills in fall and winter, whereas only 18% in summer. We found that in winter, wolverines visited wolf kills 3.6 and 6.7 times more often than single wolves and wolf groups revisited their kills, and spent 10 and 25 times as much time at carcasses compared to single wolves and groups of wolves. Thus, wolverines played an important role in the depletion of wolf-killed prey, with potential effects on the scavenging behavior of other guild members. Understanding how globally threatened top predators may function as key species in scavenging processes is important to conservation as this may have community-wide cascading effects and support important ecosystem functions and services.
Significance statement
Large carnivores serve a central role within scavenging ecology through the suppression and facilitation of intraguild competitors. The wolf, as an apex obligate predator, can provide a reliable supply of carrion, that can serve as an important resource to facultative scavengers. However, while facultative behavior helps to mitigate the effects of limited prey for scavengers, it can also increase exposure to competition and intraguild predation. Across three seasons, we explored the use of fresh wolf-killed prey by sympatric wolves and wolverines. Our findings reveal that wolves facilitate wolverines by providing scavenging opportunities, where biomass available from kills is influenced by wolves’ prey preference and group size. Wolverines, like wolves, utilized kills heavily during winter, when increased access to food is important to wolverine reproductive rates. Wolverines exhibited caching behavior, possibly reducing exposure to interspecific competition, while serving an important role in the depletion of carrion biomass.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Apex carnivores that rely primarily on predation are often considered keystone species because they can have a disproportionally large impact on the structure and functioning of ecological systems relative to their abundance (Paine 1969; Mills et al. 1993). This includes their central but complex role in the scavenging community, with their ability to outcompete and kill interspecific competitors, but also to facilitate them by provisioning a reliable source of food (Wilmers and Post 2006; Schmitz et al. 2008; Allen et al. 2015; Prugh and Sivy 2020).
Scavenging is an important process by which organisms consume dead organic material, e.g., carrion. This ecological phenomenon, often overshadowed by more conspicuous trophic interactions, like predation, serves an integral function in ecosystem health by recycling nutrients, preventing disease vectors, and creating food-web linkages (Barton et al. 2013; Beasley et al. 2015; Barton and Bump 2019; Vicente and Vercauteren 2019). Almost all predators are scavengers to some extent, i.e., facultative scavengers, (DeVault et al. 2003; Wilson and Wolkovich 2011), but scavenging differs from predation as it does not involve the act of killing prey before consumption (Schmitz et al. 2008; Selva et al. 2019). Facultative scavenging is advantageous in times or areas of sparse prey availability, but comes with risks associated with intraguild competition and aggression when multiple carnivores utilize the same carcasses (Selva and Fortuna 2007; Ordiz et al. 2020; Prugh and Sivy 2020). This may be especially relevant for carnivores in seasonal environments or with unpredictable food resources, as this may alter the degree of interspecific competition in both time and space (Polis et al. 1989; Palomares and Caro 1999; Linnell and Strand 2000; Gomo et al. 2017).
In boreal ecosystems, the wolf (Canis lupus) and the wolverine (Gulo gulo) make up a species pair consisting of an apex predator and a facultative scavenger. The wolf is a cursorial, group-living carnivore whose primary social unit consists of a territorial, adult breeding pair, forming a pack when accompanied by offspring from the current or previous litters (Mech and Boitani 2003). As an apex obligate predator, the wolf is adapted to predating on ungulate species of considerable size and hazard (Peterson and Ciucci 2003), as well as a variety of other prey, while they can utilize facultative scavenging (Peterson et al. 1984; Selva et al. 2003; Wikenros et al. 2023). In Scandinavia, wolves show a strong prey preference for moose calves throughout the year, with calves’ body size increasing from summer (13 kg), when calves are newly born, to winter when calves weigh around 150 kg (Sand et al. 2008; Zimmermann et al. 2015). As successful predators, wolves may offer beneficial scavenging opportunities (Wilmers et al. 2003a, 2003b; Pereira et al. 2014; Prugh and Sivy 2020). However, as a dominant carnivore, they have been documented to exhibit one of the highest degrees of intraguild predation on sympatric carnivore species representing a significant source of risk to subordinate carnivores (Palomares and Caro 1999).
The wolverine is the world’s largest terrestrial mustelid. The species is territorial and solitary and resides in boreal forests and tundra in the northern hemisphere (Inman et al. 2012; Mattisson et al. 2016). The wolverine is adapted to environments with unpredictable food resources, capable of killing a variety of prey, but also specialized on scavenging by searching vast areas for carcasses (Banci 1994; van Dijk et al. 2008a). Ungulate carrion represents an important food source for wolverines across its range (Magoun 1987; van Dijk et al. 2008b; Dalerum et al. 2009; Mattisson et al. 2016). Wolverines also exhibit caching behavior by removing carrion pieces from carcasses and/or hiding them in different locations, taking advantage of spatial and temporal resource surpluses to secure food predictability (Vander Wall 1990; Inman et al. 2012; van der Veen et al. 2020). Studies on interspecific interactions with other guild members show that wolverines successfully utilize carcasses killed by larger carnivores, including wolves (van Dijk et al. 2008b; Mattisson et al. 2011; Sivy et al. 2017; Klauder et al. 2021; Wallace et al. 2021). However, the competitive relationship between wolves and wolverines is not clear (Prugh and Sivy 2020; Klauder et al. 2021; Wallace et al. 2021).
In this study, we deployed remote cameras at fresh kills from GPS-collared wolves to explore the competitive relationships between wolves and wolverines. To better understand the intraguild interactions between these sympatric large carnivore species, we evaluate to what degree coexistence with wolves provided beneficial scavenging opportunities to wolverines (Wilmers et al. 2003a, 2003b; Pereira et al. 2014; Prugh and Sivy 2020), and whether these scavenging opportunities changed with seasonally varying prey biomass. Furthermore, focusing on winter when wolverines may experience low food availability (Magoun 1985; Persson 2005), we looked for evidence that wolverine utilization of wolf kills may have been hampered by their avoidance of direct interaction with wolves.
We hypothesized that available carrion biomass and hence the presence of wolves and wolverines at wolf kills would follow the seasonal change in prey size (H1), predicting that co-utilization of wolf kills by scavenging wolves and wolverines would be highest during winter when moose calves are large and lowest during summer (P1). We further hypothesized that the utilization of wolf kills by scavenging wolverines in winter would be negatively influenced by the risk perceived by wolverines at wolf kills (Zimmermann et al. 2015; Tallian et al. 2022) (H2). As most Scandinavian wolf pups dissociate from their natal pack in late winter (i.e., gradually spending less time with their pack members), and subsequently disperse from their natal territory in their first year (Nordli et al. 2023), we expected wolves to appear at their own kills both alone and in groups. We predicted that wolverines would exhibit delayed arrival at carcasses compared to single wolves or wolf groups revisiting the carcass, possibly due to the necessity of allocating time for carcass detection or to minimize chances of direct encounters with wolves (P2). Furthermore, we predicted that wolves, as the larger group-living predator, would visit carcasses more often (P3), spend more time scavenging (P4), and consume more of the remaining biomass (P5) when revisiting a carcass than wolverines. For carnivores utilizing the same food resources, such as when scavenging on carrion, temporally segregating activity patterns can reduce competitive interactions through lowered risk of encounter with a dominant predator (Palomares and Caro 1999; Allen et al. 2015; Karanth et al. 2017). We therefore hypothesized that wolverines in winter would reduce competition by adjusting their diel activity to avoid time periods when wolves are more active (H3). Hence, we predicted high diel activity partitioning between wolves and wolverines at carcasses (P6).
By examining the intraguild interactions between these two sympatric carnivores, we provide new insight into the dynamics of large carnivore guilds and the degree to which suppression or facilitation occurred between wolves and wolverines. Our findings offer important information on how changing carnivore populations may alter these effects and potentially shape ecological communities through coexistence.
Methods
Study area
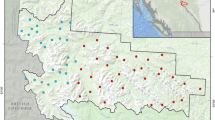
Our study area encompassed a wolf territory of 904 km2 in Innlandet County, southeastern Norway (Fig. 1; 61 °N, 08 °E). We collected data during four study periods in 2014–2015: one summer (June 2015), one fall (August–September 2014), and two late winter periods (March–April 2014 and 2015). The area is one of the few places in Europe with a complete large carnivore guild of four species: brown bear (Ursus arctos), lynx (Lynx lynx), wolf, and wolverine (May et al. 2008; Chapron et al. 2014). Throughout the study periods, the wolf territory was occupied and defended by one adult breeding pair, accompanied by their litter of pups, and one yearling. The total wolf pack size was estimated to be 8 in winter 2013/2014 and 7 in winter 2014/2015 through snow tracking and DNA sampling by the Scandinavian wolf monitoring program (for further detail see: Åkesson et al. 2022; Nordli et al. 2023). Based on wolverine monitoring by identifying active breeding sites (i.e., dens), there were ≥ 3 reproducing female wolverine territories within the wolf territory (Fig. 1) during the study period (Rovbase 2020), as resident wolverines have high territorial fidelity (Aronsson and Persson 2018). The area is characterized by boreal coniferous forest dominated by spruce (Picea abies) and pine (Pinus sylvestris), interspersed with deciduous trees dominated by birch (Betula pubescens, Betula pendula), willow (Salix caprea), aspen (Populous tremula), and rowan (Sorbus aucuparia). Elevation ranges from 215 to 1009 m.a.s.l., with tree line at approximately 800–900 m.a.s.l. The area has a continental inland climate with dry and cold winters, averaging −9.0 °C mid-winter and 15.0 °C mid-summer, and snow covered the ground from mid-November to the end of April (Johnsen et al. 2017). Within the study system, moose were the largest and most abundant ungulate and represented the main prey of wolves throughout the year (Sand et al. 2005, 2008). However, moose also have a strong economic and recreational value and are considered the most important game species in Norway (Storaas et al. 2001). The average moose density was estimated to be 1.2 moose/km2 during winter 2002–2003 (Zimmermann et al. 2007). Other potential prey for wolves were red deer (Cervus elaphus) and roe deer (Capreolus capreolus), both at low densities of 0.01 per km2 (Zimmermann et al. 2007). The human density within the study area is low, with the majority of the area being inhabited with < 1 human per km2 (Wabakken et al. 2001)

The study area defined as 100% minimum convex polygon (MCP) in black outlines the single wolf territory (904 km2), derived from all GPS-positions of the breeding pair, during March 2014 to June 2015. Gray dots show the adult-breeder’s GPS positions throughout the remote camera’s operational times during summer, fall, and the two winter study periods. The locations of fresh ungulate wolf kills (n = 29) monitored with remote cameras are indicated with triangles (summer, n = 11), squares (fall, n = 5), and black circles (winter, n = 13) (Supplementary: S1 Table 1). The bottom left indicates the wolf territory location in Norway on the Scandinavian peninsula
Carcass identification and camera setup
In winter 2013/2014, we captured and equipped five wolves in the pack (the adult breeding pair and three 9-month-old pups) with GPS/GSM collars (Vertex GPS plus, Vectronics Aerospace, Germany) as part of the long-term Scandinavian wolf research project (SKANDULV). All collars were programmed to take six GPS positions per day (UTC+1 00:00, 04:00, 08:00, 12:00, 16:00, 20:00). In order to identify wolf-killed ungulate carcass sites, during the four study periods, wolf GPS/GSM positions were downloaded daily and entered into ArcGIS (ESRI 2017), where each position was buffered with 100 m. Overlapping buffers, i.e., positions within 200 m of each other, were defined as “clusters” (Sand et al. 2005; Zimmermann et al. 2007), while positions > 200 m from the closest neighbor were termed “single positions”. Based on knowledge of wolf movement patterns, time spent at a location and the time of day, we set up a list of prioritized clusters and single positions likely to be wolf-killed ungulate carcasses (Zimmermann et al. 2007). All prioritized clusters were then visited in the field the following day(s), depending on the current location of the wolves, which was verified with a handheld VHF tracker (RX98, Followit, Sweden). In order to minimize disturbance at carcasses, we postponed checks of positions if the most recent wolf position or VHF signal was within 1 km. Postponed sites were checked once wolf movement away from the area was verified via GPS or VHF positions. During the summer period, we also checked single positions in addition to clusters using a dog on a leash to enhance the detection of smaller prey (Sand et al. 2008). The inclusion of single positions was motivated by wolf preference for killing neonate moose calves in this period (Sand et al. 2008), and the extra effort allowed us to identify kill sites with smaller prey remains. We did not visit positions closer than 1 km to the wolf natal den in summer. We estimated the time of death of the ungulate in ArcGIS by identifying the first adult breeder wolf position within 100 m of the carcass. Moreover, we estimated the time when wolves first left the carcass by using a rolling 24-hour (h) time window of adult breeders’ absence within 1 km of the carcass. Wolf carcass occupancy was defined as the time difference between the prey time of death and the time of wolf carcass abandonment.
Cameras were placed at a total of 29 wolf-killed ungulates, of those 11 neonate moose calves in summer, 5 moose calves in fall, and 12 moose (10 calves and 2 female yearlings) and 1 adult male red deer in winter (Supplementary: S1 Table 1). At each confirmed wolf-killed ungulate carcass, we set up a Reconyx HC600 (Reconyx, WI, USA) camera, programmed to take three motion-triggered pictures with a minute (min) delay, and a time-lapse picture every fifth minute. All cameras were placed so that the carcass was within the center point of the camera’s detection area at a distance not exceeding 12 m (range = 3–9 m). Camera pictures (n = 377,220) were reviewed using the program Reconyx MapView Professional (Reconyx, WI, USA). For each picture, we manually registered the species and the number of individuals per species. We standardized the camera monitoring to include only the first 21 days after the wolves first left the carcass.
We classified wolves and wolverines into three carnivore categories: single wolf, wolf group (≥ 2 pack members), and wolverine. A visit was defined as a time sequence of presence pictures of the species in question; if more than 30 min elapsed before the next presence picture, we considered this a new visit. We rounded all visits to the closest minute. For visits of less than 30 s, we considered the carnivore present for 1 min. We calculated the duration of a given visit at the carcass by subtracting the time of the start of the visit from the time of the end of the visit. When pictures of only one wolf were interspersed with pictures of a group of wolves during a specific wolf visit, all wolf pictures were classified as belonging to the same wolf group visit. We calculated the time to the first visit at the carcass as the time from when the wolves first left the carcass to the first record of presence. To quantify the visit-specific biomass reduction, we reviewed each 5-min time-lapse picture before and after each visit and visually estimated the reduction of biomass to the closest 5% and attributed that to the specific visit. For each visit, we also reviewed all pictures of wolves and wolverines to determine if any carnivore category showed caching behavior by removing carrion remains from the carcass site. It was not possible to record data blind because our study involved focal animals in the field.
Data analysis
To investigate patterns of foraging behavior at wolf kills according to our predictions, we set up six models. In the first model, we included all seasons: summer, fall, and winter, to estimate the probability of the presence of wolves and wolverines at carcasses across seasons (1) (Table 1). Due to sample size constraints (i.e., few presence pictures of the species in summer and fall), we used only carcasses from the winter season (n = 13) for estimating the time to first revisit/visit (2), number of visits (3), visit duration (4), and visit specific biomass reduction (5) (Table 1). Time to first revisit/visit (2) describes the time until either wolves (single wolf or wolf group) had returned to the carcass they had previously left or the time until the initial visit of a wolverine. Number of visits (3) presents how many times single wolves, wolf groups, or wolverines revisited/visited the carcass and visit duration (4) describes how much time the carnivores spent at the carcass per visit, whereas visit-specific biomass reduction (5) details how much biomass the carnivores ate/removed during each visit (in %), including caching behavior by carnivore category. We performed all data analyses in R, version 4.0.2 (R Development Core Team 2021). For data management and data preparation, we used the R packages “lubridate” (Grolemund and Wickham 2011) and “tidyverse” (Wickham et al. 2019). We used generalized linear mixed models (GLMMs) in a Bayesian framework using Stan from the R package “brms” (Bürkner 2017) and fitted all models with a group-level effect for carcass ID, to account for repeated samples across sites (Table 1). We checked final model diagnostics by visually comparing the fit of the posterior distribution to the observed data (Supplementary: S2 Figs. 1-5), checking for high effective sample sizes (ESS) > 100 and Rhat values not exceeding 1.01 (McElreath 2020; Vehtari et al. 2021). We used the function “add_epred_draws” from the “tidybayes” package to calculate empirical predictive draws from each model (Kay 2020). We expressed all predictions with medians and 95% credible intervals (CI) (McElreath 2020). To assess differences between carnivore categories, we used posterior samples from the estimated marginal means and compared the highest probability density intervals (HPDI) using the package “emmeans” (Lenth and Lenth 2018). We considered significant differences between groups if the 95% HPDI did not include zero (Bürkner 2017). We used default non-informative uniform (flat) priors for all models, except for model 1 to estimate the probability of presence among seasons for which a weakly informative prior was used (Table 1).
For the last model, to analyze the diel activity patterns of carcass use and to determine if single wolves, groups of wolves, and wolverines showed temporal partitioning of carcass use (6), we used the R package “overlap” (Ridout and Linkie 2009; Meredith and Ridout 2021). We explored the average daily activity pattern of each species from the standardized sample of time-lapse pictures pre-programmed to take pictures every fifth minute. We transformed the predictor variable time of day to a circular variable and fitted kernel density curves of activity to determine the coefficient of overlap (∆). We estimated the mean coefficient of overlap using Dhat1 (∆1) as suggested for small sample sizes and bootstrapped this to 10,000 replications from the R package “boot” (Canty 2002) for each pairwise combination of carnivore categories to extract 95% confidence intervals (Ridout and Linkie 2009). We considered overlap estimates of Δ1 ≤ 0.50 to be a low overlap value (i.e., high temporal partitioning), Δ1 < 0.70 to be moderate, and Δ1 ≥ 0.70 to be high (i.e., low temporal partitioning) (Monterroso et al. 2014). For all plot visualizations, we used the R package “ggplot2” (Wickham 2016).
Results
Carnivore presence across three seasons
We identified 2865 pictures containing either a single wolf, a group of wolves, or a wolverine. Wolves occupied a fresh kill for an average of 4.4 h (range = 0–20.0 h) in summer, 10.0 h (range = 0–40 h) in fall, and 23.5 h (range = 0–100 h) in winter before leaving the carcass for the first time. Cameras were set up, on average, 33.6 h (range = 8–100 h) after the time of death of the ungulate and 19.2 h (range = 4–92 h) after wolves had first left the carcass. Carcass revisits by single wolves and visits by wolverines had a predicted probability of 100% (CI = 98–100 and 99–100, respectively) in winter, whereas the predicted probability that wolf groups revisited the carcasses in winter was 56% (CI = 15–92) (Fig. 2A, Supplementary: S3Table 1). Carcass revisits by single wolves and wolf groups were less likely in fall with a predicted probability of 92% (CI = 38–100%) for single wolves and 0% (CI = 0–12) for wolf groups, whereas carcass visits by wolverines were equally likely in fall compared to winter, with a predicted probability of 100% (CI = 87–100) (Fig. 1A, Supplementary: S3 Table 1, S4 Fig. 1). Carcass (re)visits by all three carnivore categories were less likely in summer compared to winter with a predicted probability for wolverines of 18% (CI = 1–61), single wolves 17% (CI = 1–59), and 0% (CI = 0–2) for wolf groups (Fig. 2A, Supplementary: S3 Table 1, S4 Fig. 1A-C).

Predicted posterior distributions with median (black dot) and associated 95% credible intervals (CI, black bars) of metrics of carcass use by wolverines, single wolves, and group wolves at wolf kills (n = 29 for all seasons, and n = 13 for winter) 21 days from initial wolf abandonment. A Probability of presence of each carnivore category in each season, B time until first visit/revisit in winter (days), C number of (re)visits in winter, and D duration per (re)visit (minutes) in winter
Carcass utilization in winter
Visits
The predicted time until the first visit of wolverines at a carcass was 7 days after wolf abandonment (CI = 4.2–12.9). After the wolves had first left the carcass, revisits by single wolves and wolf groups were predicted at the carcass after 9.1 days (CI = 5–17.4) and 8.9 days (CI = 4.5–21.4), respectively. However, the arrival times of wolverines and revisiting wolves did not differ (Fig. 2B, Supplementary: S3 Table 2, S4 Fig. 2A).
Number and duration of visits
We recorded a total of 91 visits by wolverines and 39 revisits by wolves in winter (14 and 25 for wolf groups and single wolves, respectively). The predicted number of (re)visits at wolf-killed carcasses was on average 5.4 (CI = 3.2–8.8) for wolverines, 1.5 (CI = 0.81–2.7) for single wolves, and 0.8 (CI = 0.4–1.6) for wolves in groups (Fig. 2C, Supplementary: S3 Table 3). Wolverine visits at carcasses were significantly more frequent than single wolf and wolf group revisits, but no significant difference in the number of revisits was detected among single or group of wolves (Supplementary: S4 Fig. 2B). We only observed single wolverines at carcasses, while wolves revisited carcasses in groups of up to 4 individuals (Supplementary: S4 Fig. 3), i.e., the total pack was never recorded during revisit at a kill site. The longest (re)visit duration was predicted for wolf groups, lasting on average 20 min (CI = 9.3–45.9) followed by but not significantly different from wolverines with 15.3 min (CI = 7.9–26.2) (Fig. 2D, Supplementary: S3 Table 4). The shortest predicted (re)visit duration, and significantly different to wolverine and wolf groups, was found among single wolves with 3.3 min (CI = 1.7–6.4, Fig. 2D) (Supplementary: S3 Table 4, S4 Fig. 2C). Wolverines spent a cumulative total of 42.6 h scavenging on wolf kills, compared to single wolves of 1.7 h and group foraging wolves of 4.2 h.
Biomass reduction and caching behavior
In 72 visits by wolverines and 27 revisits by wolves where we could determine visit-specific biomass reduction and caching behavior (n = 11 carcasses), caching incidents were exclusively recorded among wolverines (n = 20, 27.8% of the wolverine visits). In visits where wolverines exhibited caching behavior, the predicted biomass reduction was 5.6% (CI = 4.0–7.0, Fig. 3) per visit, which was significantly more than when wolverines that did not cache (Fig. 4) (reduced biomass 1.8%, CI = 1.0–2.0, Fig. 3, Supplementary: S3 Table 5, S4 Fig. 4). Biomass reduction per revisit was predicted to be 2.4% (CI = 2.0–3.0) for single wolves and 10.5% (CI = 8.0–14) for wolf groups (Fig. 3, Supplementary: S5, Fig. 1). Multiplying the predicted visit-specific biomass reduction per carnivore category by accounting for the proportion of caching behaviour in wolverines with the average predicted number of (re)visits per carcass in each category yielded a total average biomass reduction per carcass of 12% for wolves (3.6% for single wolves and 8.4% for wolf groups) and 15.4% for wolverines.

Predicted posterior distributions with median (black dot) and associated 95% credible intervals (CI, black bars) of visit-specific biomass reduction (%) at wolf-killed carcasses in winter, 21 days from initial wolf abandonment

Examples of wolverines displaying caching behavior by carrying away food from moose carcasses. The pictures are from cameras mounted at kill sites of GPS-collared wolves in southeast Norway. For more examples on carcass utliization by wolves and wolverines see Supplementary: S5 Fig. 1
Temporal activity pattern and overlap
We never observed wolves and wolverines at carcasses simultaneously, nor within the same unique visit (Supplementary: S5 Fig. 1). The minimum observed time from a wolf revisit until a wolverine visit was 0.8 h (48 min). The activity patterns of wolves and wolverines showed moderate to low temporal partitioning, where single wolves and groups of wolves overlapped with wolverines by ∆1 = 0.74, and ∆1 = 0.71, respectively (Fig. 5A, B, Table 2). Wolverines showed the most nocturnal activity pattern at carcasses, with their activity peaking in the darkest hours of the day (Fig. 5A, B). Single wolf activity peaked during dusk, and single wolves had generally higher activity during daylight compared to wolverines and wolves in groups (Fig. 5A–C). Wolf groups showed a bimodal crepuscular activity pattern that peaked at both dusk and dawn, and they never appeared during midday (Fig. 5B, C). The lowest coefficient of overlap was estimated between single wolves and wolf groups at ∆1 = 0.59, indicating a moderate to high temporal partitioning among wolves (Table 2).

Diel activity patterns centered on midnight, and estimated overlap of paired combinations of wolverine (solid line), single wolf (dashed line), and wolf group (dotted line) at wolf-killed carcasses in winter 21 days after initial wolf abandonment (A-C). The rug beneath each plot represents presence observations of the carnivore category from 5-min time-lapse camera trap pictures. The gray-shaded area represents the estimated activity overlap ∆1 on the circular transformed variable time of day
Discussion
Our study shows that wolves facilitated wolverines through carcass provisioning and that wolverines alongside wolves may alter carrion persistence from wolf kills, influencing availability for other scavengers. In this low large carnivore and high prey density system (Bischof et al. 2020; Wikenros et al. 2020), where human-caused mortality is the primary driver of moose population dynamics (Solberg et al. 2000; Sand et al. 2012; Ausilio et al. 2022), we suggest that seasonal variability in available biomass at wolf kills can be an important predictor influencing wolverine scavenging behavior where these species coexist. Wolves provided scavenging opportunities by increasing carrion availability, but the amount of biomass that could be obtained from wolf kills was subject to major seasonal fluctuations, ultimately driven by wolves selecting moose calves as their main prey year-round and changes in wolf group size.
Consistent with our first hypothesis (H1), the presence of both wolverines and wolves at wolf kills was highest during winter (P1), when more biomass was remaining on kills. By generating resource surpluses, territorial wolves may buffer the unpredictability of winter carrion available to wolverines in a changing climate (Wilmers et al. 2003b; Wilmers and Getz 2005; Wikenros et al. 2013). This may be particularly important for wolverine demographics because increased access to food during this time of year has shown a positive effect on wolverine reproductive rates (Persson 2005; Rauset et al. 2015). Nevertheless, it is important to recognize that our study spanned only a limited number of monitored seasons and one wolf territory.
Generalist predators will switch prey as a functional response to changing prey availability (Holling 1959). Correspondingly, we would expect facultative scavengers to transition between scavenging and predation depending on the changing availability of prey or carrion (Pereira et al. 2014; Mattisson et al. 2016). Despite high reported levels of carcass use by wolves (Klauder et al. 2021), we did not find support for the hypothesis (H2) that the wolverine utilization of wolf kills in winter would be negatively influenced by the risk perceived by wolverines at wolf kills. Contrary to our expectations, wolverines were the most present at wolf kills followed by single wolves scavenging at carcasses, while group foraging wolves revisited only to a limited degree and only in winter. In winter, we found no significant difference between time until the first revisit/visit for either wolves or wolverines (P2). In other words, the wolverines’ initial use of the carcasses occurred around the same time as the wolves’ revisit of their kills, indicating the potential for concurrent utilization. However, wolves, both single and in groups, had significantly fewer revisits at carcasses than wolverines (P3) suggesting that the risk of encountering wolves at carcasses may be low. Our prediction for visit duration at carcasses (P4) was not supported, as wolverines spent as much time at the carcass as wolf groups during visits and significantly more than single wolves. Wolverines visited carcasses on average 3.6 times more often than single wolves, and 6.7 times more often than wolves in a group. Single wolves and groups of wolves did not differ in the number of visits but showed differing durations of visits. We found that wolverines cumulatively spent more than 10 times the amount of time at carcasses in winter compared to groups of wolves, and 25 times more than single wolves.
Our prediction (P5) that wolves would have a larger impact on biomass reduction than wolverines was supported when assessing biomass consumption per visit in winter. Indeed, on the visit-specific level, wolves in groups exhibited the highest consumptive rates, followed by single wolves and wolverines, owing to their extended visit durations, numerical advantage from social foraging, and body size (Kane et al. 2017; Gutiérrez-Cánovas et al. 2020). However, due to the lower number of wolf visits compared to wolverines, the total impact on carcass biomass by wolverines was like that of wolves combined. Beyond direct consumption at carcasses, caching was observed in about 28% of the wolverine visits. Typically, food caching is a behavioral strategy to buffer unpredictability in food availability by storing away food for later use (Vander Wall 1990), though it may also serve as a spatiotemporal strategy to reduce exposure to interspecific competition (van der Veen et al. 2020). Our results show that both wolves and wolverines may reduce carcass biomass after wolf abandonment in winter, but that wolverines may impact biomass amount disproportionally relative to their size or numbers. Interestingly, if carcass persistence is substantially altered, this may have community-wide cascading effects (Cunningham et al. 2018), potentially changing species composition and trophic interactions.
Scavengers can impact wolf kill rates by interference or exploitative competition (Tallian et al. 2017) or shape group foraging behaviors by offsetting costs of intraspecific competition in food sharing among pack members versus food loss to interspecific competitors (Vucetich et al. 2004). However, wolves consumed most biomass from their kills before they first left the carcass (Supplementary: S1 Table 1). As the primary predator, wolves may monopolize carcass resources before leaving unconsumed carcass remains in search of the next hunting opportunity (Wikenros et al. 2013; Ordiz et al. 2020). The kill rate by adult wolves on moose in Scandinavia during winter has been estimated to 4.6 days between consecutive kills, our study area included (Sand et al. 2008; Zimmermann et al. 2015). Presumably, unless the next hunt is unsuccessful (Wikenros et al. 2009), there may be reduced energetic incentive to return to a previous carcass, at least for the adult breeding pair, explaining the low visit rate of wolf groups. While the adult breeding pair remains cohesive, young wolves usually dissociate from their pack in later winter, while remaining in the natal territory (Nordli et al. 2023), without killing ungulate prey (Zimmermann et al. 2015). Consequently, the young wolves may be considered more scavengers than obligate predators for as long as they are provisioned by their parents. The predominance of single wolves at carcasses and the limited degree to which adult breeding wolves (i.e., ≥ 2 wolves) visited older kills in winter can support the idea of carcass partitioning, where the wolves may be occupied handling a fresher kill by the time a wolverine has had time to locate a carcass. As a result, the staggered carcass use may indirectly reduce the chances of direct interactions and promote coexistence.
Similar to sympatric wolves and brown bears feeding at ungulate kills (Ordiz et al. 2020), we did not observe wolves and wolverines at carcasses simultaneously, indicating that direct interactions are rare (van Dijk et al. 2008a). However, wolverines have been observed at carcasses simultaneously with wolves and lynx (López-Bao et al. 2016; Klauder et al. 2021; Jung et al. 2023). A strategy of species to avoid interference competition at a common resource is to shift their diel activity patterns to avoid the activity periods of competitors (H3) (Zalewska et al. 2021). Wolverines were the most nocturnal carcass visitors in winter, with the highest activity during the darkest hours, corresponding to previous studies (Mattisson et al. 2010; Thiel et al. 2019). Single wolves were most active at carcasses during dusk with relatively higher activity in daytime compared to wolves in groups and wolverines. Wolf group revisits to carcasses were never recorded during daytime but followed a crepuscular activity pattern with peaks at both dawn and dusk, as previously reported by Theuerkauf (2009). Contrary to our expectation (P6), we found only moderate to low temporal partitioning between wolverine and wolf activity patterns at carcasses in winter, indicating limited temporal niche separation. But we found moderate to high temporal partitioning between single and groups of wolves, indicating differences in activity patterns among single and wolves in groups at carcasses. In general, our findings show that both species were most active at similar times of the day, suggesting that differing diel activity patterns are not the primary cause limiting their direct interactions. Interestingly, Klauder et al. (2021) found that wolverines were the least vigilant species scavenging at wolf kills and wolverines can escape direct interactions and intraguild predation from wolves by climbing trees in forested landscapes (Grinnell 1926) such as our study area. However, following their historical persecution and recent recolonization (Chapron et al. 2014), large carnivores have evolved avoidance towards human activity (Ordiz et al. 2021); thus the nocturnality exhibited by both species may rather reflect temporal partitioning with their common diurnal human predator (Gaynor et al. 2018).
In conclusion, our findings indicate that the wolf, as an apex predator, can provide a reliable supply of carrion that can serve as an important resource to facultative scavengers such as the wolverine, especially in winter when wolves’ preferred prey are large and wolverine access to other food sources may be low. We also found a potential for, but no observation of, concurrent carcass utilization by wolverines and re-visiting wolves. Hence, wolverines seem to avoid direct interactions with wolves at carcasses without any apparent impact on their overall carcass utilization pattern. In general, large carnivores serve a central role within scavenging ecology because they can suppress and facilitate interspecific competitors. A reduction of large predators and dominant scavengers is expected to substantially alter the ecosystem functions and services they provide (Estes et al. 2011; Mateo-Tomás et al. 2017). Thus, a better understanding of how globally threatened and rare top predators may function as key species in scavenging processes is therefore imperative to future conservation (Ripple et al. 2014; Mateo-Tomás et al. 2017).
Data availability
The dataset analyzed during the current study is available in the supplementary information and on https://doi.org/10.18710/PGZHZJ
References
Åkesson M, Svensson L, Flagstad Ø, Wabakken P, Frank J (2022) Wolf monitoring in Scandinavia: evaluating counts of packs and reproduction events. J Wildlife Manage 86:e22206
Allen ML, Elbroch LM, Wilmers CC, Wittmer HU (2015) The comparative effects of large carnivores on the acquisition of carrion by scavengers. Am Nat 185:822–833
Aronsson M, Persson J (2018) Female breeding dispersal in wolverines, a solitary carnivore with high territorial fidelity. Eur J Wildlife Res 64:1–10
Ausilio G, Wikenros C, Sand H, Wabakken P, Eriksen A, Zimmermann B (2022) Environmental and anthropogenic features mediate risk from human hunters and wolves for moose. Ecosphere 13:e4323
Banci VA (1994) Wolverine. In: Ruggiero LF, Aubrey KB, Buskirk SW, Lyon J, Zielinski WJ (eds) The scientific basis for conserving forest carnivores: American marten, fisher, lynx and wolverine in the western United States. U.S. Department of Agriculture, Forest Service, Fort Collins, CO, USA, pp 99–127
Barton PS, Bump JK (2019) Carrion decomposition. In: Olea P, Mateo-Tomás P, Sánchez-Zapata J (eds) Carrion ecology and management. Springer, Cham, pp 101–124
Barton PS, Cunningham SA, Lindenmayer DB, Manning AD (2013) The role of carrion in maintaining biodiversity and ecological processes in terrestrial ecosystems. Oecologia 171:761–772
Beasley JC, Olson ZH, DeVault TL (2015) Ecological role of vertebrate scavengers. In: Benbow ME, Tomberlin JK, Tarone AM (eds) Carrion ecology, evolution, and their applications. CRC Press, Boca Raton, pp 107–127
Bischof R, Milleret C, Dupont P et al (2020) Estimating and forecasting spatial population dynamics of apex predators using transnational genetic monitoring. P Natl Acad Sci USA 117:30531–30538
Bürkner P-C (2017) brms: An R package for Bayesian multilevel models using Stan. J Stat Softw 80:1–28
Canty AJ (2002) Resampling methods in R: the boot package. R News 2:2–7
Chapron G, Kaczensky P, Linnell JDC et al (2014) Recovery of large carnivores in Europe's modern human-dominated landscapes. Science 346:1517–1519
Cunningham CX, Johnson CN, Barmuta LA, Hollings T, Woehler EJ, Jones ME (2018) Top carnivore decline has cascading effects on scavengers and carrion persistence. Proc R Soc B 285:20181582
Dalerum F, Kunkel K, Angerbjörn A, Shults BS (2009) Diet of wolverines (Gulo gulo) in the western Brooks Range, Alaska. Polar Res 28:246–253
DeVault TL, Rhodes J, Olin E, Shivik JA (2003) Scavenging by vertebrates: behavioral, ecological, and evolutionary perspectives on an important energy transfer pathway in terrestrial ecosystems. Oikos 102:225–234
ESRI (2017) ArcGIS Desktop Release 10.5.1. Environmental Systems Research Institute, Redlands, CA
Estes JA, Terborgh J, Brashares JS et al (2011) Trophic downgrading of planet Earth. Science 333:301–306
Gaynor KM, Hojnowski CE, Carter NH, Brashares JS (2018) The influence of human disturbance on wildlife nocturnality. Science 360:1232–1235
Gomo G, Mattisson J, Hagen BR, Moa PF, Willebrand T (2017) Scavenging on a pulsed resource: quality matters for corvids but density for mammals. BMC Ecol 17:22
Grinnell GB (1926) Some habits of the wolverine. J Mammal 7:30–34
Grolemund G, Wickham H (2011) Dates and times made easy with lubridate. J Stat Softw 40:1–25
Gutiérrez-Cánovas C, Moleón M, Mateo-Tomás P, Olea PP, Sebastián-González E, Sánchez-Zapata JA (2020) Large home range scavengers support higher rates of carcass removal. Funct Ecol 34:1921–1932
Holling CS (1959) The components of predation as revealed by a study of small-mammal predation of the European Pine Sawfly. Can Entomol 91:293–320
Inman RM, Magoun AJ, Persson J, Mattisson J (2012) The wolverine's niche: linking reproductive chronology, caching, competition, and climate. J Mammal 93:634–644
Johnsen K, Boonstra R, Boutin S, Devineau O, Krebs CJ, Andreassen HP (2017) Surviving winter: Food, but not habitat structure, prevents crashes in cyclic vole populations. Ecol Evol 7:115–124
Jung TS, Peers MJ, Drummond R, Taylor SD (2023) Dining with a glutton: an intraguild interaction between scavenging wolverine (Gulo gulo) and lynx (Lynx canadensis). Ecosphere 14:e4491
Kane A, Healy K, Guillerme T, Ruxton GD, Jackson AL (2017) A recipe for scavenging in vertebrates–the natural history of a behaviour. Ecography 40:324–334
Karanth KU, Srivathsa A, Vasudev D, Puri M, Parameshwaran R, Kumar NS (2017) Spatio-temporal interactions facilitate large carnivore sympatry across a resource gradient. Proc R Soc B 284:20161860
Kay M (2020) tidybayes: Tidy data and geoms for Bayesian models. R package version 3.0.5. http://mjskay.github.io/tidybayes/. Accessed 01.12.2023
Klauder KJ, Borg BL, Sivy KJ, Prugh LR (2021) Gifts of an enemy: scavenging dynamics in the presence of wolves (Canis lupus). J Mammal 102:558–573
Lenth R, Lenth MR (2018) Package ‘lsmeans’. Am Stat 34:216–221
Linnell JD, Strand O (2000) Interference interactions, co-existence and conservation of mammalian carnivores. Divers Distrib 6:169–176
López-Bao JV, Mattisson J, Persson J, Aronsson M, Andrén H (2016) Tracking neighbours promotes the coexistence of large carnivores. Sci Rep 6:23198
Magoun AJ (1985) Population characteristics, ecology, and management of wolverines in northwestern Alaska. PhD thesis,. University of Alaska, Fairbanks
Magoun AJ (1987) Summer and winter diets of wolverines, Gulo gulo, in arctic Alaska. Can Field-Nat 101:392–397
Mateo-Tomás P, Olea PP, Moleón M, Selva N, Sánchez-Zapata JA (2017) Both rare and common species support ecosystem services in scavenger communities. Global Ecol Biogeogr 26:1459–1470
Mattisson J, Andrén H, Persson J, Segerström P (2010) Effects of species behavior on global positioning system collar fix rates. J Wildlife Manage 74:557–563
Mattisson J, Andrén H, Persson J, Segerström P (2011) Influence of intraguild interactions on resource use by wolverines and Eurasian lynx. J Mammal 92:1321–1330
Mattisson J, Rauset GR, Odden J, Andrén H, Linnell JD, Persson J (2016) Predation or scavenging? Prey body condition influences decision-making in a facultative predator, the wolverine. Ecosphere 7:e01407
May R, van Dijk J, Wabakken P et al (2008) Habitat differentiation within the large-carnivore community of Norway's multiple-use landscapes. J Appl Ecol 45:1382–1391
McElreath R (2020) Statistical rethinking: A Bayesian course with examples in R and Stan. Chapman and Hall/CRC, Boca Raton
Mech LD, Boitani L (2003) Wolf social ecology. In: Mech LD, Boitani L (eds) Wolves: Behavior, Ecology, and Conservation. University Chicago Press, Chicago, pp 1–35
Meredith M, Ridout M (2021) Package ‘overlap’. In: Estimates of coefficient of overlapping for animal activity patterns. https://github.com/mikemeredith/overlap. Accessed 15 Dec 2023
Mills LS, Soulé ME, Doak DF (1993) The keystone-species concept in ecology and conservation. Bioscience 43:219–224
Monterroso P, Alves PC, Ferreras P (2014) Plasticity in circadian activity patterns of mesocarnivores in Southwestern Europe: implications for species coexistence. Behav Ecol Sociobiol 68:1403–1417
Nordli K, Wabakken P, Eriksen A, Sand H, Wikenros C, Maartmann E, Zimmermann B (2023) Spatial and temporal cohesion of parents and offspring in a social large carnivore. Anim Behav 197:155–167
Ordiz A, Aronsson M, Persson J, Støen O-G, Swenson JE, Kindberg J (2021) Effects of human disturbance on terrestrial apex predators. Diversity 13:68
Ordiz A, Milleret C, Uzal A, Zimmermann B, Wabakken P, Wikenros C, Sand H, Swenson JE, Kindberg J (2020) Individual variation in predatory behavior, scavenging and seasonal prey availability as potential drivers of coexistence between wolves and bears. Diversity 12:356
Paine RT (1969) A note on trophic complexity and community stability. Am Nat 103:91–93
Palomares F, Caro TM (1999) Interspecific killing among mammalian carnivores. Am Nat 153:492–508
Pereira LM, Owen-Smith N, Moleón M (2014) Facultative predation and scavenging by mammalian carnivores: Seasonal, regional and intra-guild comparisons. Mamm Rev 44:44–55
Persson J (2005) Female wolverine (Gulo gulo) reproduction: reproductive costs and winter food availability. Can J Zool 83:1453–1459
Peterson RO, Ciucci P (2003) The wolf as carnivore. In: Mech LD, Boitani L (eds) Wolves: Behavior, Ecology, and Conservation. University Chicago Press, Chicago, pp 104–130
Peterson RO, Woolington JD, Bailey TN (1984) Wolves of the Kenai peninsula, Alaska. Wildlife Monogr 88:1–52
Polis GA, Myers CA, Holt RD (1989) The ecology and evolution of intraguild predation: potential competitors that eat each other. Annu Rev Ecol Syst 20:297–330
Prugh LR, Sivy KJ (2020) Enemies with benefits: integrating positive and negative interactions among terrestrial carnivores. Ecol Lett 23:902–918
R Development Core Team (2021) R: A language and environment for statistical computing R Foundation for Statistical Computing, Vienna, Austria http://www.R-project.org
Rauset GR, Low M, Persson J (2015) Reproductive patterns result from age-related sensitivity to resources and reproductive costs in a mammalian carnivore. Ecology 96:3153–3164
Ridout MS, Linkie M (2009) Estimating overlap of daily activity patterns from camera trap data. J Agr Biol Envir St 14:322–337
Ripple WJ, Estes JA, Beschta RL et al (2014) Status and ecological effects of the world’s largest carnivores. Science 343:1241484
Rovbase (2020) Wolverine monitoring hedmark county, reproducing females and densities. In: Rovbase 3.0. Norwegian Environmental Agency, Miljodirektoratet. https://rovbase30.miljodirektoratet.no. Accessed 10.05.2023
Sand H, Vucetich JA, Zimmermann B, Wabakken P, Wikenros C, Pedersen HC, Peterson RO, Liberg O (2012) Assessing the influence of prey–predator ratio, prey age structure and packs size on wolf kill rates. Oikos 121:1454–1463
Sand H, Wabakken P, Zimmermann B, Johansson Ö, Pedersen H, Liberg O (2008) Summer kill rates and predation pattern in a wolf-moose system: can we rely on winter estimates? Oecologia 156:53–64
Sand H, Zimmermann B, Wabakken P, Andren H, Pedersen HC (2005) Using GPS technology and GIS cluster analyses to estimate kill rates in wolf-ungulate ecosystems. Wildlife Soc B 33:914–925
Schmitz OJ, Jones HP, Barton BT (2008) Scavengers. In: Joergensen SE, Fath BD (eds) Encyclopedia of Ecology. Academic Press, London, pp 3160–3164
Selva N, Fortuna MA (2007) The nested structure of a scavenger community. Proc R Soc Lond B 274:1101–1108
Selva N, Jedrzejewska B, Jedrzejewski W, Wajrak A (2003) Scavenging on European bison carcasses in Bialowieza primeval forest (eastern Poland). Ecoscience 10:303–311
Selva N, Moleón M, Sebastián-González E, DeVault TL, Quaggiotto MM, Bailey DM, Lambertucci SA, Margalida A (2019) Vertebrate scavenging communities. In: Olea PP, Mateo-Tomás P, Sánchez-Zapata JA (eds) Carrion ecology and management. Springer, Cham, pp 71–99
Sivy KJ, Pozzanghera CB, Grace JB, Prugh LR (2017) Fatal attraction? Intraguild facilitation and suppression among predators. Am Nat 190:663–679
Solberg EJ, Loison A, Sæther BE, Strand O (2000) Age-specific harvest mortality in a Norwegian moose Alces alces population. Wildlife Biol 6:41–52
Storaas T, Gundersen H, Henriksen H, Andreassen HP (2001) The economic value of moose in Norway-a review. Alces 37:97–107
Tallian A, Ordiz A, Metz MC et al (2017) Competition between apex predators? Brown bears decrease wolf kill rate on two continents. Proc R Soc B 284:20162368
Tallian A, Ordiz A, Metz MC et al (2022) Of wolves and bears: Seasonal drivers of interference and exploitation competition between apex predators. Ecol Monogr 92:e1498
Theuerkauf J (2009) What drives wolves: fear or hunger? Humans, diet, climate and wolf activity patterns. Ethology 115:649–657
Thiel A, Evans AL, Fuchs B, Arnemo JM, Aronsson M, Persson J (2019) Effects of reproduction and environmental factors on body temperature and activity patterns of wolverines. Front Zool 16:21
van der Veen B, Mattisson J, Zimmermann B, Odden J, Persson J (2020) Refrigeration or anti-theft? Food-caching behavior of wolverines (Gulo gulo) in Scandinavia. Behav Ecol Sociobiol 74:52
van Dijk J, Andersen T, May R, Andersen R, Andersen R, Landa A (2008a) Foraging strategies of wolverines within a predator guild. Can J Zool 86:966–975
van Dijk J, Gustavsen L, Mysterud A et al (2008b) Diet shift of a facultative scavenger, the wolverine, following recolonization of wolves. J Anim Ecol 77:1183–1190
Vander Wall SB (1990) Food hoarding in animals. University of Chicago Press, Chicago
Vehtari A, Gelman A, Simpson D, Carpenter B, Bürkner P-C (2021) Rank-normalization, folding, and localization: An improved R ̂ for assessing convergence of MCMC (with discussion). Bayesian Anal 16:667–718
Vicente J, Vercauteren K (2019) The role of scavenging in disease dynamics. In: Olea PP, Mateo-Tomás P, Sánchez-Zapata JA (eds) Carrion ecology and management. Springer, Cham, pp 161–182
Vucetich JA, Peterson RO, Waite TA (2004) Raven scavenging favours group foraging in wolves. Anim Behav 67:1117–1126
Wabakken P, Sand H, Liberg O, Bjarvall A (2001) The recovery, distribution, and population dynamics of wolves on the Scandinavian peninsula, 1978-1998. Can J Zool 79:710–725
Wallace CF, Golla JM, Allen ML (2021) Direct observations of a wolverine scavenging at an active gray wolf kill site. Northwest Nat 102:157–160
Wickham H (2016) ggplot2: elegant graphics for data analysis. Springer, Cham
Wickham H, Averick M, Bryan J et al (2019) Welcome to the Tidyverse. J Open Source Softw 4:1686
Wikenros C, Di Bernardi C, Zimmermann B et al (2023) Scavenging patterns of an inbred wolf population in a landscape with a pulse of human-provided carrion. Ecol Evol 13:e10236
Wikenros C, Sand H, Ahlqvist P, Liberg O (2013) Biomass flow and scavengers use of carcasses after re-colonization of an apex predator. PLoS ONE 8:e77373
Wikenros C, Sand H, Månsson J, Maartmann E, Eriksen A, Wabakken P, Zimmermann B (2020) Impact of a recolonizing, cross-border carnivore population on ungulate harvest in Scandinavia. Sci Rep 10:21670
Wikenros C, Sand H, Wabakken P, Liberg O, Pedersen HC (2009) Wolf predation on moose and roe deer: chase distances and outcome of encounters. Acta Theriol 54:207–218
Wilmers CC, Crabtree RL, Smith DW, Murphy KM, Getz WM (2003a) Trophic facilitation by introduced top predators: grey wolf subsidies to scavengers in Yellowstone National Park. J Anim Ecol 72:909–916
Wilmers CC, Getz WM (2005) Gray wolves as climate change buffers in Yellowstone. PLoS Biol 3:e92
Wilmers CC, Post E (2006) Predicting the influence of wolf-provided carrion on scavenger community dynamics under climate change scenarios. Global Change Biol 12:403–409
Wilmers CC, Stahler DR, Crabtree RL, Smith DW, Getz WM (2003b) Resource dispersion and consumer dominance: scavenging at wolf-and hunter-killed carcasses in Greater Yellowstone, USA. Ecol Lett 6:996–1003
Wilson EE, Wolkovich EM (2011) Scavenging: how carnivores and carrion structure communities. Trends Ecol Evol 26:129–135
Zalewska K, Waggershauser CN, Kortland K, Lambin X (2021) The best defence is not being there: avoidance of larger carnivores is not driven by risk intensity. J Zool 315:110–122
Zimmermann B, Sand H, Wabakken P, Liberg O, Andreassen HP (2015) Predator-dependent functional response in wolves: from food limitation to surplus killing. J Anim Ecol 84:102–112
Zimmermann B, Wabakken P, Sand H, Pedersen HC, Liberg O (2007) Wolf movement patterns: a key to estimation of kill rate? J Wildlife Manage 71:1177–1182
Acknowledgements
We are indebted to the capture team, J.M. Arnemo, A.L. Evans, T.H. Strømseth, and B. Fuchs, who darted and handled the wolves; to pilot U. Grinde for safe flying of the helicopter; to the Norwegian Elkhound Enya, E. Maartmann, F. Holen, E.R. Dahl, and J. Meiholt who assisted in fieldwork, along with numerous students at Campus Evenstad who participated in the wolf tracking; and to O. Devineau for advice on data management. We also thank A. Thiel, the editor, and two anonymous reviewers for valuable comments on the manuscript.
Funding
Open access funding provided by Inland Norway University Of Applied Sciences This study was supported by the Scandinavian Wolf Research Project (SKANDULV), alongside the Norwegian Boreal Forest Wolverine Project (SKOGSJERV). Financially, the projects have been funded by the Norwegian Research Council, the Norwegian Environment Agency, Inland Norway University of Applied Sciences, County Governor of Innlandet (Norway), Swedish Environmental Protection Agency, Swedish University of Agricultural Sciences, and Swedish Association for Hunting and Wildlife Management.
Author information
Authors and Affiliations
Contributions
KN: field work, conceptualization, methodology, data analyses, writing—original draft, writing—review and editing. ZW: field work, conceptualization, methodology, writing—review and editing. AE: conceptualization, writing—review and editing. MR: field work, conceptualization, methodology, writing—review and editing. BZ: supervision, conceptualization, methodology, writing—review and editing. CW: writing—review and editing. MA: methodology, writing—review and editing. PW: supervision, conceptualization, methodology, writing—review and editing, funding acquisition.
Corresponding author
Ethics declarations
Ethical approval
Captures and handling of wolves were performed by trained and certified personnel with several veterinarians at the site, following standardized biomedical protocols. Captures and capture-related procedures, e.g., biomedical sampling and/or GPS-collaring of wolves, were approved by both the Norwegian Environmental Agency and the ethical committee (FOTS:7224) at the Norwegian Food Safety Authority.
Competing interests
The authors declare no competing interests.
Additional information
Communicated by S. Ciuti
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
ESM 1
(DOCX 2987 kb)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Nordli, K., Walton, Z., Eriksen, A. et al. Carcass provisioning and intra-guild risk avoidance between two sympatric large carnivores. Behav Ecol Sociobiol 78, 17 (2024). https://doi.org/10.1007/s00265-023-03423-6
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00265-023-03423-6