
Abstract
The string-pulling paradigm is an approach commonly used in the study of animal cognition to investigate problem-solving abilities. This test involves an out-of-reach reward that can only be acquired through pulling a string. Australian magpies (Gymnorhina tibicen tyrannica) can solve cognitive tasks requiring associative and reversal learning, spatial memory, and inhibitory control. Nonetheless, whether magpies can pass a test of means-end understanding—the string-pull test—is unclear. We tested wild magpies on a string-pulling task with five configurations, including a long loose string that required several pulls to obtain food, a short string that only required a single pull to complete, and two short strings—one intact and connected to the food reward; the other broken and unable to yield food when pulled. Of the 11 magpies tested, none solved the long string task at first presentation. Two naïve birds and three, subsequently trained, birds succeeded at the short string. Once proficient at the short string, four of these five birds then solved the long string task; in addition, three learned to choose a functional, intact string over a non-functional, broken string. Overall, these observations provide evidence that Australian magpies have the ability to solve string-pulling tasks but it remains unclear whether they do so by trial-and-error or if they possess means-end understanding.
Significance statement
We found, for the first time, that wild, free-ranging Australian magpies can learn to solve different versions of a string-pulling task; a version with a short string, a long string, and two strings next to each other where one of them was broken and non-functional (broken string task). Some magpies spontaneously solved the task with the short string, where other magpies required training, and magpies only managed to solve the task with the long string after they had succeeded on the short string task. Furthermore, some of the magpies solved the broken string task but required a high number of trials to do so. Overall, our findings show that Australian magpies have the ability to solve string-pulling tasks but it remains unclear whether they do so by trial-and-error or if they understand causal relations between objects.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Causal reasoning is adaptive for animals that live in complex environments where learning about reoccurring sequences of events (physical or social) is important for survival (Emery and Clayton 2004; Huber and Gajdon 2006). Herein, we define causal reasoning as the construction of mental representations of sequences of events, where a particular event brings about a second event via a certain action or process. Such representations lead to the understanding of how the world works and why it works the way it does (Povinelli 2000). On the other hand, an alternative adaptation to causal reasoning is associative learning which links together adjoining events. Associative learning is restricted to effects that immediately precede causes, such that A always leads to B, though associative learning can also occur over longer time intervals with only probabilistic outcomes, but without any understanding of how and why that is (Pearce and Bouton 2001; Gershman 2015). However, in some habitats it may be more adaptive to understand how the world works, for example in rapidly changing urban environments where the habitat might change drastically within the lifetime of an individual. Causal reasoning is therefore adaptive for animals living in complex environments, faced with situations where important events are separated in time and space (Huber and Gajdon 2006). Detecting causality requires that the animal can assign and comprehend cause-and-effect to objects or other animals in the environment and that there are specific means leading to an end goal. Such means-end comprehension requires knowledge of causal relations between objects and may lead to representations of abstract concepts such as connectivity and connectedness between objects, and an understanding that a chain of events or actions results in a specific outcome (Huber and Gajdon 2006; Krasheninnikova 2019). Means-end reasoning and understanding involves finding means for achieving goals and is expressed behaviourally as a deliberate and planned execution of a sequence of actions to attain a desirable outcome. Understanding of such means-end relations can be identified in circumstances where an obstacle preventing an animal from achieving a goal must be removed (Huber and Gajdon 2006; Jacobs and Osvath 2015; Krasheninnikova 2019).
String-pulling is a common experimental paradigm used to evaluate means-end understanding and problem-solving performance of animals in cognition research (Jacobs and Osvath 2015). In this task, a reward (usually food or a food container) is attached to the end of a piece of string that must be pulled to bring the reward within reach. In the case of string-pulling, the aforementioned obstacle that must be removed is the distance to the out-of-reach reward. To have means-end understanding, the animal must recognize the string as a means to obtain the reward, and understand that there is a causal relationship between string and reward (Krasheninnikova 2019). There are many variations on the string-pulling task, for instance, strings can be oriented horizontally or vertically, and with a single string or multiple strings. Multiple strings can be presented in a variety of patterns (patterned-string tasks: e.g., perpendicular, slanted, crossed, contact/no contact, etc.) that tests how animals solve the problem of reaching the reward (Wasserman et al. 2013).
String-pulling, a naturally occurring behaviour in some species, may provide a means by which to explore in which environments and sociobiological contexts cognitive abilities such as causal reasoning evolved (Jacobs and Osvath 2015). In the wild, some bird species pull on twigs, caterpillar threads, and seedlings to obtain food items (Altevogt 1954; Thorpe 1963; Dickinson 1969; Bossema 1979; Seibt and Wickler 2006), and crows and ravens (Corvus spp.) have been observed to learn how to tug on fishing lines to obtain fish (Larsson 1958; Boswall 1977). In a laboratory setting, several avian groups such as passerines, parrots, pigeons, and birds-of-prey have succeeded on variations of the string-pulling task, albeit with high between-individual variation in performance (reviewed in Wasserman et al. 2013; Jacobs and Osvath 2015; Wakonig et al. 2021). Some species of passerines, parrots, and pigeons learn to pull strings by trial-and-error (Vince 1958, 1961; Schmidt and Cook 2006; Chaves Molina et al. 2019). In contrast, ravens (Corvus corax) and kea (Nestor notabilis) show rapid problem-solving in that they spontaneously solve string-pulling tasks on the first attempt and seem to understand means-end relationships between string and reward (Heinrich 1995; Heinrich and Bugnyar 2005; Werdenich and Huber 2006; Bastos et al. 2021). An interesting example comes from a study on African grey parrots (Psittacus erithacus), where performance on the task was compromised in individuals that received language training and could verbally request the reward from the researcher instead of pulling the string (Pepperberg 2004). In addition to foraging ecology, the socio-ecology of a species might also be a factor for evolving skills that can prove useful for solving string-pulling problems. At least in parrots, social organization seems to predict string-pulling performance better than other ecological factors, phylogeny, or brain size (Krasheninnikova et al. 2013; Krasheninnikova 2014). Similar cognitive skills used for string pulling might have evolved in different species through convergent or parallel evolution of socioecological variables (Osvath et al. 2014).
In this paper, we address the relevance of the string-pulling test in the Australian magpie (Gymnorhina tibicen; sometimes referred to as Cracticus t.). Australian magpies are passerine birds and members of the Artamidae which also includes butcherbirds, currawongs, and wood swallows (Christidis and Boles 2008; Kearns et al. 2013; Cake et al. 2018; Gill et al. 2023). Magpies sometimes breed cooperatively and live in territorial groups ranging in size from 2 up to 26 individuals (depending on the subspecies), but usually with at least a dominant male and female pair (Kaplan 2019). They are highly territorial and groups cooperate to defend their territory and subordinate magpies within the group are known to participate in alloparental care. Juveniles usually remain within the group’s territory for at least a year (but in some cases longer), and by the next breeding season are forced to leave by the adults (Kaplan 2019). Magpies exhibit complex behaviours such as play in addition to living in social structures, and having a long developmental period before fledging fully (before leaving the territory), all of which are associated with complex cognition in birds (Emery 2006; Kaplan 2019, 2020). In addition, Australian magpies have an impressive vocal repertoire, are capable of vocal learning, and are even able to discriminate different human voices (Brown et al. 1988; Brown and Farabaugh 1991; Kaplan 2005; Dutour et al. 2021). Australian magpies are curious and eagerly participate in experiments presented to them in both wild and laboratory settings; they can succeed on cognitive tasks requiring associative and reversal learning, spatial memory, and inhibitory control, and can learn to pull on sticks to extract food (Mirville et al. 2016; Ashton 2017; Ashton et al. 2018; Blackburn et al. 2022; Connelly et al. 2022; Johnsson et al. 2022a, b). These features make magpies an excellent model species for cognition research. Furthermore, magpies are also highly social birds, which may drive the evolution of more advanced cognition (Humphrey 1976; Dunbar 1998). Some of these social drivers for evolution of complex cognition include the need to maintain and coordinate relationships, keep track of other group members, recognize cooperative partners, and the need to outwit rivals in competitive interactions (Byrne and Whiten 1988; Brosnan et al. 2010; Massen et al. 2014). Accordingly, wild Australian magpies that live in larger groups show increased cognitive performance, which is also associated with increased reproductive success (Ashton et al. 2018). Therefore, based on previous cognitive studies on magpies and because of the social group living in this species, we predict that Australian magpies have evolved the ability to understand causality and exhibit means-end behaviour (Emery and Clayton 2004).
In this study, we tested the ability of means-end understanding in wild Australian magpies by their ability to succeed in several configurations of the string-pulling paradigm. We tested magpies naïve to string-pulling tasks on a single long string, which required multiple pulls on the string to obtain the food reward. If magpies failed, we tested whether they could solve a simpler task that involved a short string that only required a single pull. The reasoning for presenting magpies with a longer string before a short string was to see if they could innovate a solution to a novel and more difficult problem before being presented with an easier problem. Then, after the short string we, again, presented magpies with a long string to investigate whether magpies that were no longer naïve to string-pulling tasks could find success. Finally, we tested whether magpies understood string continuity by presenting them with a task that required them to discriminate between two strings: one being broken and thus non-functional.
Methods
Study species
We studied 11 free-ranging, wild, adult Australian magpies (7 females) of the Victorian subspecies Gymnorhina tibicen tyrannica (also known as Cracticus t. t.; see Cake et al. 2018; Gill et al. 2023; but see also Kearns et al. 2013) at La Trobe University Bundoora campus and Albert Park, Melbourne, Australia, from October 2021 to May 2022. Both study sites were city parklands consisting of large lawns, trees, bodies of water, walkways, roads, buildings, and with anthropogenic disturbances (i.e., people walking, bicycles, street traffic, etc.). The 11 magpies stemmed from 5 separate territories (3 at Albert Park and 2 at La Trobe University Bundoora campus; Table 1). Each territory was approximately 3 – 5 hectares.
Sex and age were identified based on plumage (males have a white plumage on the back and females have a grey back plumage; juveniles have mottled grey plumage), beak coloration (juveniles have darker beak colouration), and behaviour (begging behaviour, parental behaviour, dominance displays) (Kaplan 2019). Seven of the 11 magpies were banded for individual identification; the others were identified based on unique body characteristics and behaviour. Australian magpies are often curious, and in urban habitats they are accustomed to being fed by humans. Therefore, they readily interacted with us and the tasks presented to them. In fact, most of the time they would come towards us as we entered their territories.
String-pulling apparatus
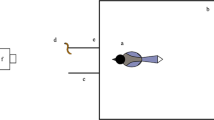
The testing apparatus consisted of a transparent plastic box (30 cm long × 22 cm wide × 8 cm high) attached to a wooden base (33 cm long × 34.5 cm wide × 1.5 cm thick). A white string (3 mm diameter and either long [80 cm] or short [20 cm]) was attached to a transparent plastic dish (7 cm diameter and 3 cm high) containing grated cheese. A rectangular hole (19 cm wide × 3.5 cm high) was cut out of one end of the box which allowed the dish to be placed inside the box (Fig. 1). The birds could see the full string length and food dish through the transparent plastic box.

String-pulling apparatus. (A) Long string configuration used in Experiments 1 and 4, (B) short string configuration used in Experiments 2 and 3, and (C) broken string configuration used in Experiment 5
Training
Australian magpies are ground-foraging birds (Kaplan 2019). Therefore, we presented a horizontal string-pulling apparatus to them on the ground, within each bird’s territory. However, before any of the experiments started, we offered the magpies food on dishes placed on the string-pulling apparatus. This training consisted of two steps. First, we placed the food-filled dish on top of the apparatus. To pass this step, magpies had to eat from the dish three consecutive times. Once completed, we placed the dish containing food half-way into the apparatus, so the magpie could eat from the dish without pulling the string; each magpie had to eat from the dish another three consecutive times. After training, we tested the birds on different string-pulling experiments detailed below (Fig. 2).

Experimental protocol. Flow-chart showing the order of experiments depending on success (green), failure (red), or either (purple)
General experimental procedures and limitations
All birds habituated readily to the experimenters (RDJ and PSV) and would approach to within a meter radius. For training and experiments, we would first enter a magpie territory and search for birds by walking around the area. If we spotted a magpie, we would throw small pieces of cheese to try to attract the bird. If a magpie approached, we would then place the string-pulling apparatus on the ground and then step away approximately 4 m from the apparatus to allow the magpie to interact. All experiments were recorded with a Sony HDR-PJ430VE camera mounted on a tripod. Testing and scoring was done by RDJ and PSV, and RDJ looked through the videos to confirm the scores. It was not possible to record data blind because our study involved focal animals in the field.
Since all magpies were free-ranging wild birds, on some days we could not find them within their territories and on other days they would not opt to interact. We were only able to test them when they willingly interacted with the apparatus. Thus, motivation was not standardized as these were wild birds tested haphazardly over time. Because we tested the magpies within their own territories, we could not isolate the focal individual from other group members during testing, making it likely (or even certain) that group members observed subjects during testing. These limitations were the same for all experiments.
Experiment 1: Long string
In Experiment 1, we presented naïve magpies with a long string (80 cm) coiled inside the apparatus (Fig. 3A). Because the string was long, the magpies had to pull it several times in order to obtain the dish containing the food reward, and thus successfully make one extraction. Five successful extractions served as our criterion for success; maximum time-on-task per extraction before defaulting was 10 min. The timer and apparatus were reset after a successful extraction, however, if the time was reached but the extraction was unsuccessful the bird failed the experiment. Within the time limit, the bird was allowed to try and fail to extract food by pulling the string. If a bird attempted to pull the string, without obtaining food, and then walked away from the apparatus, then the string was quickly reset to its original configuration (< 1 min). The timer for time-on-task commenced when the bird was within a ca. 1 m radius of the apparatus, and the timer was paused when the bird went outside the radius. If a bird stopped interacting and flew away, the timer was stopped, the apparatus was removed, and we presented the test again on a following day. On some occasions we had to motivate the magpies to participate by placing small pieces of cheese near (but not touching) the string, or on, or in front of, the apparatus. The definition for time-on-task and method of motivation was the same for all experiments.

Top view diagram of the five experiments. (A) Experiment 1, (B) Experiment 2, (C) Experiment 3, (D) Experiment 4, and (E) Experiment 5
Experiments 2 & 3: Short string and training
If magpies failed Experiment 1, they moved on to the simpler Experiment 2 where we presented magpies with a short string (20 cm) that only required a single pull to succeed (Fig. 3B). If magpies failed Experiment 2, we trained them to complete the short string task as an Experiment 3, although, it is important to note that after having received training, the magpies where again presented with an identical string configuration as in Experiment 2, the only difference being that the magpies had now been trained to complete the task (Fig. 3C). Here, we trained magpies in a stepwise manner by first placing the food-containing dish outside the apparatus (as in the training phase), and then for each time they fed from the dish, we placed the dish deeper into the apparatus until it reached the same position as in Experiment 2. To encourage interaction with the string, we also put cheese in contact with the string, so the string would move when the magpies pecked the cheese. Once the dish had reached the same position as in Experiment 2, provided the magpie successfully extracted the food five times in total, they advanced to the next experiment.
Experiment 4: Long string after short string
If magpies failed on Experiment 1 and succeeded on Experiments 2 or 3, they moved on to Experiment 4. Here, we presented magpies with exactly the same long string test as in Experiment 1, the only difference being that now the magpies were no longer naïve given their experience on successfully pulling a short string (Fig. 3D). The success criterion was five successful extractions in total.
Experiment 5: Broken string
Lastly, if magpies succeeded on the short string (Experiments 2 or 3), they were tested on Experiment 5. Here we presented the magpies with two parallel short strings: one string was intact and connected to the food dish; the other was broken in the middle and thus non-functional (Fig. 3E). Whether the broken string was on the left or right was pseudo-randomized; the same string could only be presented on the same side at most twice in a row. We used 10 out of 12 (83%) correct choices as our criterion for successfully completing the task. Each magpie was tested until reaching 10/12 correct choices and was given a maximum of 100 choices to reach criterion after which the task was recorded as a failure. As per all previous experiments, maximum time-on-task was 10 min with the timer reset whenever the magpie pulled a string. A binomial test was done in IBM SPSS Statistics 28.0.
Results
None of the 11 Australian magpies were able to spontaneously solve the long string task within the given timeframe (Experiment 1) (Table 1). Some of the magpies occasionally pulled the string, but not enough to extract the food dish from the apparatus. Two naïve birds (Experiment 2) and three trained birds (Experiment 3), succeeded at the short string. Once proficient at the short string, four of these five birds could then solve the long string (Experiment 4). Furthermore, of these five birds, three magpies learned to discriminate between an intact string and a broken string within 100 trials (Experiment 5) (Table 2; Supplementary Table 1). The magpies required a high number (54, 71, and 77, respectively) of trials to solve the task, with a close-to-chance performance in the first twelve trials compared to the above-chance levels in the last twelve trials (10/12 successful trials represents a significant deviation from random binomial probability; binomial test: p = 0.039).
Discussion
Some wild Australian magpies can spontaneously solve, or be trained to solve, a string-pulling task that requires the bird to pull a horizontally-oriented, single rewarded short string where only one pull is needed to succeed. Magpies can also be trained to solve a long string task that requires several pulls to reel-in the reward, and thus does not rely on immediate feedback. However, magpies were only able to solve this long string task after they had had experience on pulling a short string. Taken together, these tasks that involve a single string provide evidence that Australian magpies possess the necessary sensorimotor skills required to obtain the reward. Because only those individuals that had experience pulling a short string were able to solve the long string task, Australian magpies could, in principle, rely on a perceptual-motor feedback loop when pulling strings. Such a feedback loop does not rely on any mental modelling of the effects of an animal’s actions; instead, it may involve only associative learning or operant conditioning (Taylor et al. 2010). However, at least to our knowledge Australian magpies do not exhibit behaviour reminiscent of continuous pulling in the wild, and it is unlikely that magpies naturally pull on an object to get to another object. Therefore, it is also possible that once a magpie realises that pulling a string produces a food reward, then they are primed to learn more complex pulling tasks.
Importantly, Australian magpies can, likely facilitated through trial-and-error, learn to discriminate between broken and intact strings. The broken string test (also called “contact/no contact” e.g., Jacobs and Osvath 2015) necessitates that the animal has the capacity to visually determine whether objects are physically connected. This task was solved by three of five Australian magpies. The magpies probably solved this task by trial-and-error learning as indicated by the high number of trials needed to succeed on the task, which is also shown by the close-to-chance performances in the first twelve trials compared to the above-chance levels in the last twelve trials. Nevertheless, this result suggests that wild Australian magpies might be able to learn the concept of connectedness between two objects (Piaget 1954), which in turn is evidence consistent with the idea that they have or can acquire means-end understanding (as per Jacobs and Osvath 2015), though further study is needed.
An alternative, and perhaps more parsimonious, explanation is that Australian magpies learnt to associate the perceptual features of the gap in the broken string with the reward rather than obtaining any understanding of means-end relations (Povinelli 2000). Indeed, this explanation would be similar to results from pigeons and African grey parrots that can learn to succeed on a means-end task, but do so via focus on visual differences between the strings instead of functionally understanding the means-end problem (Schmidt and Cook 2006; Chaves Molina et al. 2019). In order to test whether magpies rely on perceptual cues or have means-end understanding, one could test them on a task where they need to pull a broken string to receive the reward, which could be done using thin, visually undetectable fishing line (or some other string) to connect the broken string. If magpies learn to solve this problem that violates causal understanding of connectedness between objects, that will provide evidence for that they do not have means-end understanding, but rely on perceptual features.
Notwithstanding, the result that some magpies could succeed on the broken string task is comparable to that of corvids and parrots, such as hooded crows (Corvus cornix; n = 6 successful birds of 8 studied; Bagotskaya et al. 2012), azure-winged magpies (Cyanopica cyanus; n = 3 of 6; Wang et al. 2019), Lear’s macaw (Anodorhynchus leari; n = 1 of 4) and Hyacinth macaws (Anodorhynchus hyacinthinus; n = 4 of 4; Schuck-Paim et al. 2009), galahs (Eolophus roseicapilla; n = 3 of 6) and a cockatiel (Nymphicus hollandicus; n = 1 of 10; Krasheninnikova 2013), spectacled parrotlets (Forpus conspicillatus; n = 7 of 8; Krasheninnikova et al. 2013), and peach-fronted conures (Eupsittula aurea; n = 3 of 4; Torres Ortiz et al. 2019). One can also see the great between-individual variation in success among the species that solve the broken string task; variability that is reflected in our magpies (n = 3 of 5).
In contrast, some birds that failed on the broken string task were: New Caledonian crows (Corvus moneduloides; n = 11; Taylor et al. 2012), great-tailed grackles (Quiscalus mexicanus; n = 8; Logan 2016), blue-fronted amazons (Amazona aestiva; n = 2; Schuck-Paim et al. 2009), rainbow lorikeets (Trichoglossus haematodus; n = 10), green-winged macaws (Ara chloroptera; n = 4) and sulphur-crested cockatoos (Cacatua galerita triton; n = 3; Krasheninnikova et al. 2013), African grey parrots (Psittacus erithacus; n = 3; Chaves Molina et al. 2019), and kea (Nestor notabilis; n = 7; Bastos et al. 2021). It is curious that New Caledonian crows and kea which are known for their complex cognition do not solve the broken string task. Although it should be noted that the New Caledonian crows and kea were presented with a more difficult task that involved a coiled long string (as in our Experiment 1) that was broken rather than a short string as in our study (Taylor et al. 2012; Bastos et al. 2021; but see Auersperg et al. 2009 where kea solve a similar means-end task). Certainly, it would be interesting to test our Australian magpies that solved the broken string task on the same task using a coiled long string.
To conclude that an animal has means-end understanding, performance has to be investigated using multiple strings (patterned-string problems). Four criteria have been proposed that establishes means-end understanding: (1) goal-directedness, (2) no proximity errors, (3) flexible solutions, and (4) no dependence on perceptual feedback (Jacobs and Osvath 2015). Here, we argue that Australian magpies may fulfill criterion 2 in our broken string task, and criterion 4 in our long string task. If the magpies had relied solely on proximity to the reward as a rule for which string to pull and paid no attention to the connection between string and reward, they would have performed at chance level because both the intact and broken strings had the same distance from the end of the string to the reward. However, with our results we are not able to fully rule out proximity errors and future studies should test magpies on patterned-string problems where only one string is baited and the strings are presented perpendicular or slanted to each other (see Jacobs and Osvath 2015 for details). In the long string task, magpies had to pull the string multiple times before the reward moved. Therefore, the magpies could not rely on immediate visual feedback to solve the problem (i.e., the food did not move closer on the initial pull). That said, it remains possible that magpies relied on operant conditioning (of pulling a string) given they solved the long string task only after they had succeeded on the short string task. Consequently, our study would have benefitted from testing magpies on different patterned-string tasks that further test the four criteria for means-end understanding (Jacobs and Osvath 2015). For example, future studies should present magpies with two strings next to one another with only one string baited to provide evidence for goal-directedness, and slanted or crossed strings to strengthen our results on the absence of proximity errors condition. To test flexible solutions, a string with a weight too heavy to pull can be presented to see whether magpies inhibit their pulling response, and to more rigorously test for dependence on perceptual feedback, a task that restricts visual feedback to the reward can be used.
Conclusion
Some Australian magpies can solve string-pulling tasks that require them to pull single short and long strings to receive a reward. They were also able to learn a patterned-string task that require them to discriminate between a broken and an intact string. Overall, our results indicate that Australian magpies can learn to solve a means-end task, and may have the capacity to understand contact between objects. However, after the magpies had learnt to discriminate broken strings, whether the magpies relied on an association or means-end reasoning to continue to solve the problem remains to be tested.
Data availability
For raw data see Supplementary material.
References
Altevogt R (1954) Über das “Schöpfen” einiger Vogelarten. Behaviour 6:147–152
Ashton BJ, Ridley AR, Edwards EK, Thornton A (2018) Cognitive performance is linked to group size and affects fitness in Australian magpies. Nature 554:364–367
Ashton BJ (2017) The causes and consequences of individual variation in cognitive ability in the cooperatively breeding Australian magpie (Cracticus tibicen australis). Dissertation, The University of Western Australia
Auersperg AMI, Gajdon GK, Huber L (2009) Kea (Nestor notabilis) consider spatial relationships between objects in the support problem. Biol Lett 5:455–458
Bagotskaya MS, Smirnova AA, Zorina ZA (2012) Corvidae can understand logical structure in baited string-pulling tasks. Neurosci Behav Physiol 42:36–42
Bastos APM, Wood PM, Taylor AH (2021) Kea (Nestor notabilis) fail a loose-string connectivity task. Sci Rep 11:15492
Blackburn G, Broom E, Ashton BJ, Thornton A, Ridley AR (2022) Heat stress inhibits cognitive performance in wild Western Australian magpies, Cracticus tibicen dorsalis. Anim Behav 188:1–11
Bossema I (1979) Jays and oaks: an eco-ethological study of a symbiosis. Behaviour 70:1–116
Boswall J (1977) Tool-using by birds and related behaviour. Avicult Mag 83:88–97
Brosnan SF, Salwiczek L, Bshary R (2010) The interplay of cognition and cooperation. Phil Trans R Soc B 365:2699–2710
Brown ED, Farabaugh SM (1991) Song sharing in a group-living songbird, the Australian magpie, Gymnorhina tibicen. Part III. Sex specificity and individual specificity of vocal parts in communal chorus and duet songs. Behaviour 118:244–274
Brown ED, Farabaugh SM, Veltman CJ (1988) Song sharing in a group-living songbird, the Australian magpie, Gymnorhina tibicen. Part I. Vocal sharing within and among social groups. Behaviour 104:1–28
Byrne RW, Whiten A (1988) Machiavellian intelligence: social expertise and the evolution of intellect in monkeys, apes, and humans. Clarendon Press, Oxford, UK
Cake M, Black A, Joseph L (2018) The generic taxonomy of the Australian magpie and Australo-Papuan butcherbirds is not all black-and-white. Bull Brit Ornithol Club 138:346–359
Chaves Molina AB, Cullell TM, Mimó MC (2019) String-pulling in African grey parrots (Psittacus erithacus): performance in discrimination tasks. Behaviour 156:847–857
Christidis L, Boles WE (2008) Systematics and Taxonomy of Australian Birds. CSIRO Publishing, Collingwood, Australia
Connelly F, Hall ML, Johnsson RD, Elliot-Kerr S, Dow BR, Lesku JA, Mulder RA (2022) Urban noise does not affect cognitive performance in wild Australian magpies. Anim Behav 188:35–44
Dickinson JC Jr (1969) A string-pulling tufted titmouse. Auk 86:559
Dunbar RIM (1998) The social brain hypothesis. Evol Anthropol 6:178–190
Dutour M, Walsh SL, Speechley EM, Ridley AR (2021) Female Western Australian magpies discriminate between familiar and unfamiliar human voices. Ethology 127:979–985
Emery NJ (2006) Cognitive ornithology: the evolution of avian intelligence. Phil Trans R Soc B 361:23–43
Emery NJ, Clayton NS (2004) The mentality of crows: convergent evolution of intelligence in corvids and apes. Science 306:1903–1907
Gershman SJ (2015) A unifying probabilistic view of associative learning. PLoS Comput Biol 11:e1004567
Gill F, Donsker D, Rasmussen P (2023) IOC World Bird List (v13.1), https://doi.org/10.14344/IOC.ML.13.1
Heinrich B (1995) An experimental investigation of insight in common ravens (Corvus corax). Auk 112:994–1003
Heinrich B, Bugnyar T (2005) Testing problem solving in ravens: string-pulling to reach food. Ethology 111:962–976
Huber L, Gajdon GK (2006) Technical intelligence in animals: the kea model. Anim Cogn 9:295–305
Humphrey N (1976) The social function of intellect. In: Bateson PPG, Hinde RA (eds) Growing points in ethology. Cambridge University Press, Cambridge, UK, pp 303–317
Jacobs IF, Osvath M (2015) The string-pulling paradigm in comparative psychology. J Comp Psychol 129:89–120
Johnsson RD, Connelly F, Gaviraghi Mussoi J, Vyssotski AL, Cain KE, Roth TC II, Lesku JA (2022a) Sleep loss impairs cognitive performance and alters song output in Australian magpies. Sci Rep 12:6645
Johnsson RD, Connelly F, Lesku JA (2022b) Preliminary evidence of tool use in an Australian magpie? Behaviour 159:1483–1497
Kaplan G (2019) Australian magpie: biology and behaviour of an unusual songbird, 2nd edn. CSIRO Publishing, Australia
Kaplan G (2020) Play behaviour, not tool using, relates to brain mass in a sample of birds. Sci Rep 10:20437
Kaplan G (2005) Vocal behaviour of Australian magpies (Gymnorhina tibicen): a study of vocal development, song learning, communication and mimicry in the Australian magpie. Dissertation, The University of Queensland
Kearns AM, Joseph L, Cook LG (2013) A multilocus coalescent analysis of the speciational history of the Australo-Papuan butcherbirds and their allies. Mol Phylogenet Evol 66:941–952
Krasheninnikova A (2013) Patterned-string tasks: relation between fine motor skills and visual-spatial abilities in parrots. PLoS ONE 8:e85499
Krasheninnikova A (2019) Means-end reasoning. In: Vonk J, Shackelford T (eds) Encyclopedia of animal cognition and behavior. Springer, Cham, pp 1–6
Krasheninnikova A, Bräger S, Wanker R (2013) Means-end comprehension in four parrot species: explained by social complexity. Anim Cogn 16:755–764
Krasheninnikova A (2014) Physical cognition in parrots: a comparative approach. Dissertation, University of Hamburg, Hamburg, Germany
Larsson E (1958) Fiskande kråkor och korpar [Fishing crows and ravens]. Fauna Och Flora 53:92–94
Logan CJ (2016) How far will a behaviourally flexible invasive bird go to innovate? R Soc Open Sci 3:160247
Massen JJM, Szipl G, Spreafico M, Bugnyar T (2014) Ravens intervene in others’ bonding attempts. Curr Biol 24:2733–2736
Mirville MO, Kelley JL, Ridley AR (2016) Group size and associative learning in the Australian magpie (Cracticus tibicen dorsalis). Behav Ecol Sociobiol 70:417–427
Osvath M, Kabadayi C, Jacobs I (2014) Independent evolution of similar complex cognitive skills: the importance of embodied degrees of freedom. Anim Behav Cogn 1:249–264
Pearce JM, Bouton ME (2001) Theories of associative learning in animals. Annu Rev Psychol 52:111–139
Pepperberg IM (2004) “Insightful” string-pulling in grey parrots (Psittacus erithacus) is affected by vocal competence. Anim Cogn 7:263–266
Piaget J (1954) The construction of reality in the child. Basic Books, New York, USA
Povinelli DJ (2000) Folk physics for apes. Oxford University Press, Oxford, UK
Schmidt GF, Cook RG (2006) Mind the gap: means-end discrimination by pigeons. Anim Behav 71:599–608
Schuck-Paim C, Borsari A, Ottoni EB (2009) Means to an end: neotropical parrots manage to pull strings to meet their goals. Anim Cogn 12:287–301
Seibt U, Wickler W (2006) Individuality in problem solving: string pulling in two Carduelis species (Aves: Passeriformes). Ethology 112:493–502
Taylor AH, Medina FS, Holzhaider JC, Hearne LJ, Hunt GR, Gray RD (2010) An investigation into the cognition behind spontaneous string pulling in New Caledonian crows. PLoS ONE 5:e9345
Taylor AH, Knaebe B, Gray RD (2012) An end to insight? New Caledonian crows can spontaneously solve problems without planning their actions. Proc R Soc Lond B 279:4977–4981
Thorpe WH (1963) Learning and instinct in animals. Methuen, London
Torres Ortiz S, Maxwell A, Krasheninnikova A, Wahlberg M, Larsen ON (2019) Problem solving capabilities of peach-fronted conures (Eupsittula aurea) studied with the string-pulling test. Behaviour 156:815–846
Vince MA (1958) String-pulling in birds. II. Differences related to age in greenfinches, chaffinches and canaries. Anim Behav 6:53–59
Vince MA (1961) ‘String-pulling’ in birds. III. The successful response in greenfinches and canaries. Behaviour 17:103–129
Wakonig B, Auersperg AMI, O’Hara M (2021) String-pulling in the Goffin’s cockatoo (Cacatua goffiniana). Learn Behav 49:124–136
Wang L, Luo Y, Wang X, Maierdiyali A, Chang H, Li Z (2019) Azure-winged magpies solve string-pulling tasks by partial understanding of the physical cognition. Curr Zool 65:385–392
Wasserman EA, Nagasaka Y, Castro L, Brzykcy SJ (2013) Pigeons learn virtual patterned-string problems in a computerized touch screen environment. Anim Cogn 16:737–753
Werdenich D, Huber L (2006) A case of quick problem solving in birds: string pulling in keas, Nestor notabilis. Anim Behav 71:855–863
Acknowledgements
We would like to thank José Henrique Provazi das Neves for helping to video record magpies. We also thank the two anonymous reviewers for their comments and suggestions on the manuscript.
Funding
Open Access funding enabled and organized by CAUL and its Member Institutions
Author information
Authors and Affiliations
Contributions
Conceived and designed experiment: RDJ, FC, JAE, TCR and JAL. Investigation: RDJ and PSV. Analysis: RDJ. Writing- Original draft: RDJ. Writing- Review & Editing: RDJ, PSV, FC, JAE, TCR and JAL.
Corresponding author
Ethics declarations
Ethical approval
All applicable international, national, and/or institutional guidelines for the use of animals were followed. All procedures were carried out with permission from the Department of Environment, Land, Water and Planning (permit number: 10010344), the Australian Bird and Bat Banding Scheme (#1405), and the La Trobe University Animal Ethics Committee (AEC22002).
Competing interests
The authors declare no competing interests.
Additional information
Communicated by N. Clayton.
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Johnsson, R.D., Veillet, P.S., Connelly, F. et al. Wild Australian magpies learn to pull intact, not broken, strings to obtain food. Behav Ecol Sociobiol 77, 49 (2023). https://doi.org/10.1007/s00265-023-03326-6
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00265-023-03326-6