
Abstract
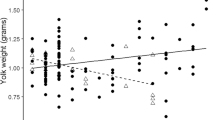
Maternal allocation of androgens to the egg yolk allows mothers to adaptively manipulate offspring phenotype. Increases of egg androgen levels have often been shown to induce sex-specific effects. Some previous studies suggest that females specifically may suffer a fitness reduction after early exposure to high androgen levels. In this study, we explored whether female birds that developed exposed to high yolk androgen concentrations would reduce their maternal investment later in life. We explored two different aspects of female reproduction in spotless starlings (Sturnus unicolor): egg coloration (a post mating sexual signal in this species) and egg size. We manipulated androgen yolk levels in a large sample of clutches of this species. We monitored the reproduction of the females that hatched from these eggs, measuring the size and colour of the eggs they laid throughout their life (from 1 to 7 years of age). We found no overall difference in egg colour in relation to treatment, clutch number or age. However, females exposed to high egg androgen levels showed a steeper decrease in egg size along the laying order than controls. This pattern likely results in a more unbalanced distribution of resources within the clutch, possibly favouring brood size reductions in experimental females. In addition, control and experimental females differed in how egg volume changed with age. These results show that some egg characteristics may be affected by the female’s exposure to yolk androgens during her own embryonic development. Our research calls for further long-term research on the influence of prenatal androgens on the fitness mechanisms regulating reproductive investment and its potential signalling role in a perspective of sexual selection.
Significance statement
In birds, mothers prepare the offspring to the environment they will encounter at hatching by depositing a mixture of compounds into the eggs. Androgens specifically exert a variety of effects during embryo development and chick growth, but some of these may be negative in adult life. In particular, early androgen may be detrimental for females in the long term, decreasing maternal investment and fitness. Using the spotless starling as study species, we tested whether an experimental increase in yolk androgens negatively affects breeding investment, measured by egg colour (a sexual signal) and egg size. We found no effects of the treatment on egg colour. However, females exposed to high androgen levels during their development reduced egg size along the laying order more strongly than controls. This may affect nestling survival, and indicates that prenatal yolk androgens may influence some egg characteristics with known relevance for offspring viability.




Similar content being viewed by others

Data availability
The datasets generated and/or analysed during the current study are available in the Figshare repository, at the permanent link: https://doi.org/10.6084/m9.figshare.13574312.v1.
References
Alonso-Alvarez C, Bertrand S, Faivre B, Chastel O, Sorci G (2007) Testosterone and oxidative stress: the oxidation handicap hypothesis. Proc R Soc B 274:819–825. https://doi.org/10.1098/rspb.2006.3764
Alonso-Alvarez C, Canelo T, Romero-Haro AÁ (2017) The oxidative cost of reproduction: theoretical questions and alternative mechanisms. BioScience 67:258–270. https://doi.org/10.1093/biosci/biw176
Bates D, Mächler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4. J Stat Softw 67:1–48. https://doi.org/10.18637/jss.v067.i01
Calvo JM, Pascual JA, Deceuninck B, Peris SJ (2000) Intraspecific nest parasitism in the spotless starling Sturnus unicolor. Bird Study 47:285–294. https://doi.org/10.1080/00063650009461188
Christians JK (2002) Avian egg size: variation within species and inflexibility within individuals. Biol Rev 77:1–26. https://doi.org/10.1017/S1464793101005784
Costantini D, Goutte A, Barbraud C, Faivre B, Sorci G, Weimerskirch H, Delord K, Chastel O (2015) Demographic responses to oxidative stress and inflammation in the wandering albatross (Diomedea exulans). PLoS One 10:e0133967. https://doi.org/10.1371/journal.pone.0133967
Cunningham EJA, Russell AF (2000) Egg investment is influenced by male attractiveness in the mallard. Nature 404:74–77. https://doi.org/10.1038/35003565
Dewailly D, Robin G, Peigne M, Decanter C, Pigny P, Catteau-Jonard S (2016) Interactions between androgens, FSH, anti-Müllerian hormone and estradiol during folliculogenesis in the human normal and polycystic ovary. Hum Reprod Update 22:709–724. https://doi.org/10.1093/humupd/dmw027
Eising CM, Groothuis TGG (2003) Yolk androgens and begging behaviour in black-headed gull chicks: an experimental field study. Anim Behav 66:1027–1034. https://doi.org/10.1006/anbe.2003.2287
Eising CM, Eikenaar C, Schwabl H, Groothuis TGG (2001) Maternal androgens in black-headed gull (Larus ridibundus) eggs: consequences for chick development. Proc R Soc B 268:839–846. https://doi.org/10.1098/rspb.2001.1594
von Engelhardt N, Carere C, Dijkstra C, Groothuis TGG (2006) Sex-specific effects of yolk testosterone on survival, begging and growth of zebra finches. Proc R Soc B 273:65–70. https://doi.org/10.1098/rspb.2005.3274
Falchuk KH, Contin JM, Dziedzic TS, Feng Z, French TC, Heffron GJ, Montorzi M (2002) A role for biliverdin IX in dorsal axis development of Xenopus laevis embryos. P Natl Acad Sci U S A 99:251–256. https://doi.org/10.1073/pnas.012616099
Folstad I, Karter AJ (1992) Parasites, bright males, and the immunocompetence handicap. Am Nat 139:603–622. https://doi.org/10.1086/285346
Galván I, Alonso-Alvarez C (2010) Yolk testosterone shapes the expression of a melanin-based signal in great tits: an antioxidant-mediated mechanism? J Exp Biol 213:3127–3130. https://doi.org/10.1242/jeb.045096
Gibson KF, Williams TD (2017) Intraclutch egg size variation is independent of ecological context among years in the European starling Sturnus vulgaris. J Ornithol 158:1099–1110. https://doi.org/10.1007/s10336-017-1473-4
Gil D (2008) Hormones in avian eggs: physiology, ecology and behavior. Adv Study Behav 38:337–398. https://doi.org/10.1016/S0065-3454(08)00007-7
Gil D, Marzal A, de Lope F, Puerta M, Møller AP (2006) Female house martins (Delichon urbica) reduce egg androgen deposition in response to a challenge of their immune system. Behav Ecol Sociobiol 60:96–100. https://doi.org/10.1007/s00265-005-0145-1
Gil D, Alfonso-Iñiguez S, Pérez-Rodríguez L, Muriel J, Monclús R (2019) Harsh conditions during early development influence telomere length in an altricial passerine: links with oxidative stress and corticosteroids. J Evol Biol 32:111–125. https://doi.org/10.1111/jeb.13396
Giraudeau M, Ziegler A-K, Pick JL, Ducatez S, Canale CI, Tschirren B (2017) Interactive effects of yolk testosterone and carotenoid on prenatal growth and offspring physiology in a precocial bird. Behav Ecol 28:31–38. https://doi.org/10.1093/beheco/arw127
Gronstol G, Blomqvist D, Wagner RH (2006) The importance of genetic evidence for identifying intra-specific brood parasitism. J Avian Biol 37:197–199. https://doi.org/10.1111/j.2006.0908-8857.03574.x
Groothuis TGG, Schwabl H (2002) Determinants of within- and among-clutch variation in levels of maternal hormones in black-headed gull eggs. Funct Ecol 16:281–289. https://doi.org/10.1046/j.1365-2435.2002.00623.x
Groothuis TGG, Eising CM, Dijkstra C, Müller W (2005) Balancing between costs and benefits of maternal hormone deposition in avian eggs. Biol Lett 1:78–81. https://doi.org/10.1098/rsbl.2004.0233
Groothuis TGG, Hsu BY, Kumar N, Tschirren B (2019) Revisiting mechanisms and functions of prenatal hormone-mediated maternal effects using avian species as a model. Philos Trans R Soc B 374:20180115. https://doi.org/10.1098/rstb.2018.0115
Hargitai R, Herényi M, Torok J (2008) Eggshell coloration in relation to male ornamentation, female condition and egg quality in the collared flycatcher Ficedula albicollis. J Avian Biol 39:413–422. https://doi.org/10.1111/j.0908-8857.2008.04337.x
Honza M, Požgayová M, Procházka P, Cherry MI (2011) Blue-green eggshell coloration is not a sexually selected signal of female quality in an open-nesting polygynous passerine. Naturwissenschaften 98:493–499. https://doi.org/10.1007/s00114-011-0790-3
Hoyt DF (1979) Practical methods of estimating volume and fresh weight of bird eggs. Auk 96:73–77
Krist M (2011) Egg size and offspring quality: a meta-analysis in birds. Biol Rev 86:692–716. https://doi.org/10.1111/j.1469-185X.2010.00166.x
Krist M, Grim T (2007) Are blue eggs a sexually selected signal of female collared flycatchers? A cross-fostering experiment. Behav Ecol Sociobiol 61:863–876. https://doi.org/10.1007/s00265-006-0315-9
Kumar N, Lohrentz A, Gahr M, Groothuis TGG (2019) Steroid receptors and their regulation in avian extraembryonic membranes provide a novel substrate for hormone mediated maternal effects. Sci Rep 9:11501. https://doi.org/10.1038/s41598-019-48001-x
Kuznetsova A, Brockhoff PB, Christensen RHB (2017) lmerTest package: tests in linear mixed effects models. J Stat Softw 2:1–26. https://doi.org/10.18637/jss.v082.i13
López-Rull I, Celis P, Gil D (2007) Egg colour covaries with female expression of a male ornament in the spotless starling (Sturnus unicolor). Ethology 113:926–933. https://doi.org/10.1111/j.1439-0310.2007.01400.x
López-Rull I, Mikšík I, Gil D (2008) Egg pigmentation reflects female and egg quality in the spotless starling Sturnus unicolor. Behav Ecol Sociobiol 62:1877–1884. https://doi.org/10.1007/s00265-008-0617-1
López-Rull I, Salaberria C, Gil D (2010) Seasonal decline in egg size and yolk androgen concentration in a double brooded passerine. Ardeola 57:321–332
McDonagh AF (2001) Turning green to gold. Nat Struct Mol Biol 8:198–200. https://doi.org/10.1038/84915
Monclús R, Muriel J, Pérez-Rodríguez L, Møller AP, Gil D (2017) The role of the mating system and intraspecific brood parasitism in the costs of reproduction in a passerine bird. Oecologia 185:629–639. https://doi.org/10.1007/s00442-017-3977-2
Morales J (2020) Eggshell biliverdin as an antioxidant maternal effect. BioEssays 42:2000010. https://doi.org/10.1002/bies.202000010
Morales J, Velando A, Moreno J (2008) Pigment allocation to eggs decreases plasma antioxidants in a songbird. Behav Ecol Sociobiol 63:227–233. https://doi.org/10.1007/s00265-008-0653-x
Morales J, Velando A, Torres R (2011) Biliverdin-based egg coloration is enhanced by carotenoid supplementation. Behav Ecol Sociobiol 65:197–203. https://doi.org/10.1007/s00265-010-1025-x
Moreno J, Osorno JL (2003) Avian egg colour and sexual selection: does eggshell pigmentation reflect female condition and genetic quality? Ecol Lett 6:803–806. https://doi.org/10.1046/j.1461-0248.2003.00505.x
Moreno J, Veiga JP, Cordero PJ, Mínguez E (1999) Effects of paternal care on reproductive success in the polygynous spotless starling Sturnus unicolor. Behav Ecol Sociobiol 47:47–53. https://doi.org/10.1007/s002650050648
Moreno J, Osorno JL, Morales J, Merino S, Tomás G (2004) Egg colouration and male parental effort in the pied flycatcher Ficedula hypoleuca. J Avian Biol 35:300–304. https://doi.org/10.1111/j.0908-8857.2004.03407.x
Moreno J, Morales J, Lobato E, Merino S, Tomás G, Martínez-de la Puente J (2005) Evidence for the signaling function of egg color in the pied flycatcher Ficedula hypoleuca. Behav Ecol 16:931–937. https://doi.org/10.1093/beheco/ari072
Moreno J, Lobato E, Morales J, Merino S, Tomás G, Martínez-de la Puente J, Sanz JJ, Mateo R, Soler JJ (2006) Experimental evidence that egg color indicates female condition at laying in a songbird. Behav Ecol 17:651–655. https://doi.org/10.1093/beheco/ark014
Moreno J, Lobato E, Morales J (2011) Eggshell blue-green colouration fades immediately after oviposition: a cautionary note about measuring natural egg colours. Ornis Fenn 88:51–56
Moreno J, Morales J, Martínez J (2013) HSP70 level in blood is associated with eggshell blue-green colouration in the pied flycatcher. Avian Biol Res 6:297–301. https://doi.org/10.3184/175815513X13805559244972
Mousseau TA, Fox CW (1998a) Maternal effects as adaptation. Oxford University Press, Oxford
Mousseau TA, Fox CW (1998b) The adaptive significance of maternal effects. Trends Ecol Evol 13:403–407. https://doi.org/10.1016/S0169-5347(98)01472-4
Müller W, Eising CM, Dijkstra C, Groothuis TGG (2002) Sex differences in yolk hormones depend on maternal social status in leghorn chickens (Gallus gallus domesticus). Proc R Soc Lond B 269:2249–2255. https://doi.org/10.1098/rspb.2002.2159
Müller W, Deptuch K, López-Rull I, Gil D (2007) Elevated yolk androgen levels benefit offspring development in a between-clutch context. Behav Ecol 18:929–936. https://doi.org/10.1093/beheco/arm060
Müller W, Vergauwen J, Eens M (2009) Long-lasting consequences of elevated yolk testosterone levels on female reproduction. Behav Ecol Sociobiol 63:809–816. https://doi.org/10.1007/s00265-009-0714-9
Muriel J, Pérez-Rodríguez L, Puerta M, Gil D (2015a) Diverse dose-response effects of yolk androgens on embryo development and nestling growth in a wild passerine. J Exp Biol 218:2241–2249. https://doi.org/10.1242/jeb.118257
Muriel J, Salmón P, Nunez-Buiza A, de Salas F, Pérez-Rodríguez L, Puerta M, Gil D (2015b) Context-dependent effects of yolk androgens on nestling growth and immune function in a multibrooded passerine. J Evol Biol 28:1476–1488. https://doi.org/10.1111/jeb.12668
Muriel J, Pérez-Rodríguez L, Ortiz-Santaliestra ME, Puerta M, Gil D (2017) Sex-specific effects of high yolk androgen levels on constitutive and cell-mediated immune responses in nestlings of an altricial passerine. Physiol Biochem Zool 90:106–117. https://doi.org/10.1086/688445
Muriel J, Pérez-Rodríguez L, Gil D (2019) Age-related patterns of yolk androgen deposition are consistent with adaptive brood reduction in spotless starlings. Behav Ecol Sociobiol 73:160. https://doi.org/10.1007/s00265-019-2770-0
Navara KJ, Hill GE, Mendonça MT (2006) Yolk testosterone stimulates growth and immunity in house finch chicks. Physiol Biochem Zool 79:550–555. https://doi.org/10.1086/501054
Navarro C, Pérez-Contreras T, Avilés JM, McGraw KJ, Soler JJ (2011) Blue-green eggshell coloration reflects yolk antioxidant content in spotless starlings Sturnus unicolor. J Avian Biol 42:538–543. https://doi.org/10.1111/j.1600-048X.2011.05293.x
Nilsson JÅ, Svensson E (1993) Causes and consequences of egg mass variation between and within blue tit clutches. J Zool 230:469–481. https://doi.org/10.1111/j.1469-7998.1993.tb02699.x
Nilsson JF, Tobler M, Nilsson JÅ, Sandell MI (2011) Long-Lasting consequences of elevated yolk testosterone for metabolism in the zebra finch. Physiol Biochem Zool 84:287–291. https://doi.org/10.1086/659006
Noguera JC, Alonso-Alvarez C, Kim S-Y, Morales J, Velando A (2011) Yolk testosterone reduces oxidative damages during postnatal development. Biol Lett 7:93–95. https://doi.org/10.1098/rsbl.2010.0421
Parolini M, Possenti CD, Romano A, Caprioli M, Rubolini D, Saino N (2018) Physiological increase of yolk testosterone level does not affect oxidative status and telomere length in gull hatchlings. PLoS One 13:e0206503. https://doi.org/10.1371/journal.pone.0206503
Pilz KM, Smith HG, Sandell MI, Schwabl H (2003) Interfemale variation in egg yolk androgen allocation in the European starling: do high-quality females invest more? Anim Behav 65:841–850. https://doi.org/10.1006/anbe.2003.2094
Pilz KM, Smith HG, Andersson M (2005) Brood parasitic European starlings do not lay high-quality eggs. Behav Ecol 16:507–513. https://doi.org/10.1093/beheco/ari017
R Core Team (2020) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. https://www.R-project.org/
van de Pol M, Verhulst S (2006) Age-dependent traits: a new statistical model to separate within- and between-individual effects. Am Nat 167:766–773. https://doi.org/10.1086/503331
van de Pol M, Wright J (2009) A simple method for distinguishing within- versus between-subject effects using mixed models. Anim Behav 77:753–758. https://doi.org/10.1016/j.anbehav.2008.11.006
Rubolini D, Martinelli R, Von Engelhardt N, Romano M, Groothuis TGG, Fasola M, Saino N (2007) Consequences of prenatal androgen exposure for the reproductive performance of female pheasants (Phasianus colchicus). Proc R Soc B 274:137–142. https://doi.org/10.1098/rspb.2006.3696
Rutkowska J, Cichoń M, Puerta M, Gil D (2005) Negative effects of elevated testosterone on female fecundity in zebra finches. Horm Behav 47:585–591. https://doi.org/10.1016/j.yhbeh.2004.12.006
Ruuskanen S, Laaksonen T (2010) Yolk hormones have sex-specific long-term effects on behavior in the pied flycatcher (Ficedula hypoleuca). Horm Behav 57:119–127. https://doi.org/10.1016/j.yhbeh.2009.09.017
Ruuskanen S, Doligez B, Pitala N, Gustafsson L, Laaksonen T (2012) Long-term fitness consequences of high yolk androgen levels: sons pay the costs. Funct Ecol 26:884–894. https://doi.org/10.1111/j.1365-2435.2012.01994.x
Ruuskanen S, Lehikoinen E, Nikinmaa M, Siitari H, Waser W, Laaksonen T (2013) Long-lasting effects of yolk androgens on phenotype in the pied flycatcher (Ficedula hypoleuca). Behav Ecol Sociobiol 67:361–372. https://doi.org/10.1007/s00265-012-1456-7
Saino N, Ferrari RP, Romano M, Martinelli R, Lacroix A, Gil D, Møller AP (2006) Maternal allocation of androgens and antagonistic effects of yolk androgens on sons and daughters. Behav Ecol 17:172–181. https://doi.org/10.1093/beheco/arj023
Schwabl H (1993) Yolk is a source of maternal testosterone for developing birds. P Natl Acad Sci U S A 90:11446–11450. https://doi.org/10.1073/pnas.90.24.11446
Schwabl H (1996) Maternal testosterone in the avian egg enhances postnatal growth. Comp Biochem Physiol A 114:271–276. https://doi.org/10.1016/0300-9629(96)00009-6
Siefferman L, Navara KJ, Hill GE (2006) Egg coloration is correlated with female condition in eastern bluebirds (Sialia sialis). Behav Ecol Sociobiol 59:651–656. https://doi.org/10.1007/s00265-005-0092-x
Smith HG, Ottosson U, Ohlsson T (1993) Interclutch variation in egg mass among starlings Sturnus vulgaris reflects female condition. Ornis Scand 24:311–316. https://doi.org/10.2307/3676793
Sockman KW, Schwabl H (2000) Yolk androgens reduce offspring survival. Proc R Soc B 267:1451–1456. https://doi.org/10.1098/rspb.2000.1163
Soler JJ, Navarro C, Contreras TP, Avilés JM, Cuervo JJ (2008) Sexually selected egg coloration in spotless starlings. Am Nat 171:183–194. https://doi.org/10.1086/524958
Soler JJ, Ruiz-Castellano C, Figuerola J, Martínez-de la Puente J, Ruiz-Rodríguez M, Tomás G (2018) Egg colouration predicts brood size, telomere length and body condition of spotless starling fledglings. J Avian Biol 49:jav-012512. https://doi.org/10.1111/jav.01686
Stoffel MA, Nakagawa S, Schielzeth H (2017) rptR: repeatability estimation and variance decomposition by generalized linear mixed-effects models. Methods Ecol Evol 8:1639–1644. https://doi.org/10.1111/2041-210X.12797
Tobler M, Sandell MI, Chiriac S, Hasselquist D (2013) Effects of prenatal testosterone exposure on antioxidant status and bill color in adult zebra finches. Physiol Biochem Zool 86:333–345. https://doi.org/10.1086/670194
Treidel LA, Whitley BN, Benowitz-Fredericks ZM, Haussmann MF (2013) Prenatal exposure to testosterone impairs oxidative damage repair efficiency in the domestic chicken (Gallus gallus). Biol Lett 9:20130684. https://doi.org/10.1098/rsbl.2013.0684
Tschirren B, Richner H, Schwabl H (2004) Ectoparasite-modulated deposition of maternal androgens in great tit eggs. Proc R Soc B 271:1371–1375. https://doi.org/10.1098/rspb.2004.2730
Veiga JP, Polo V (2008) Fitness consequences of increased testosterone levels in female spotless starlings. Am Nat 172:42–53. https://doi.org/10.1086/587850
Viñuela J (1997) Adaptation vs. constraint: intraclutch egg-mass variation in birds. J Anim Ecol 66:781–792. https://doi.org/10.2307/5995
Wang XT, Deng XM, Zhao CJ, Li JY, Xu GY, Lian LS, Wu CX (2007) Study of the deposition process of eggshell pigments using an improved dissolution method. Poult Sci 86:2236–2238. https://doi.org/10.1093/ps/86.10.2236
Williams TD (1994) Intraspecific variation in egg size and egg composition in birds: effects on offspring fitness. Biol Rev 69:35–59. https://doi.org/10.1111/j.1469-185X.1994.tb01485.x
Williams TD (2012) The hormonal and physiological control of egg production. In: Physiological adaptations for breeding in birds. Princeton University Press, Princeton, pp 8–51
Acknowledgments
We would like to thank two anonymous reviewers for their valuable comments that have substantially improved the manuscript. We are very grateful to all the people that have contributed to collating the data presented in this study, including field technicians: Oscar Magaña, Hector Bintanel and Lara Moreno; MSc students: Blanca Jimeno, Louise Froud, Elodie Faux, Irene García Celada, Thomas Crouchet and Pablo Salmón; and volunteers: Lucia Manzanares, Robyn Womack, Cristiano Gala, Evan Burnett, Hannah van Noordt, Benoit Vallas, Joao Heriberto de Oliveira, Angelica Camacho, Jorge Garcia Campa, Lara de la Cita, Aida Rodrigues and Simon Degrise. This study is a contribution to the research developed at “El Ventorrillo” field station.
Funding
This paper is the result of a long-term study financed by a series of research grants by the Spanish Ministerio de Ciencia, Innovación y Universidades to DG (refs.: CGL2008-03501/BOS, CGL2011-26318 and CGL2017-83843-C2-1-P) and LP-R (ref.: PGC2018-099596-B-I00, co-financed by the European Regional Development Fund -ERDF-). From 2009 to 2013, JM was supported by a FPI grant (BES-2009-021383) from the ICINN and currently by a postdoctoral grant from the Juan De La Cierva Subprogram (FJCI-2017-34109), with the financial sponsorship of the MICINN. RM was funded by a Marie Skłodowska Curie Action within the Horizon 2020 Initiative from the European Commission.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Ethical approval
All applicable international, national and/or institutional guidelines for the care and use of animals were followed (RD 53/2013). Capture and manipulation of birds were authorised by the Consejería de Medio Ambiente (Comunidad de Madrid, Spain) under licence from the Spanish institutional authorities (Consejería de Medio Ambiente and Centro de Migración de Aves de SEO/BirdLife). The androgen manipulation experiment complies with the animal laws in Spain in 2011 (Real Decreto 1201/2005, 10-10-2005). Permission to work in the area was granted by both the Parque Regional de la Cuenca Alta del Manzanares (Consejería de Medio Ambiente de la Comunidad Autónoma de Madrid: Ref. 10/129049.9/11) and the Ayuntamiento de Soto del Real, Spain.
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Communicated by M. Leonard
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
ESM 1
(DOCX 27.6 kb)
Rights and permissions
About this article

Cite this article
D’Arpa, S.R., Muriel, J., Monclús, R. et al. Prenatal manipulation of yolk androgen levels affects egg size but not egg colour in a songbird. Behav Ecol Sociobiol 75, 52 (2021). https://doi.org/10.1007/s00265-021-02991-9
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00265-021-02991-9