
Abstract
The pace-of-life syndrome (POLS) hypothesis predicts that behavior and physiology covary with life history. Evidence for such covariation is contradictory, possibly because systematic sources of variation (e.g. sex) have been neglected. Sexes often experience different selection pressures leading to sex-specific allocation between reproduction and self-maintenance, facilitating divergence in life-history. Sex-specific differences in means and possibly variances may therefore play a key role in the POLS framework. We investigate whether sexes differ in means and variances along the fast-slow pace-of-life continuum for life history and physiological and behavioral traits. In addition, we test whether social and environmental characteristics such as breeding strategy, mating system, and study environment explain heterogeneity between the sexes. Using meta-analytic methods, we found that populations with a polygynous mating system or for studies conducted on wild populations, males had a faster pace-of-life for developmental life-history traits (e.g., growth rate), behavior, and physiology. In contrast, adult life-history traits (e.g., lifespan) were shifted towards faster pace-of-life in females, deviating from the other trait categories. Phenotypic variances were similar between the sexes across trait categories and were not affected by mating system or study environment. Breeding strategy did not influence sex differences in variances or means. We discuss our results in the light of sex-specific selection that might drive sex-specific differences in pace-of-life and ultimately POLS.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Selection is assumed to favor the integration of life-history and physiology and behavioral traits, mediating the trade-off between current and future reproduction. This so called pace-of-life syndrome (POLS) hypothesis has been proposed as a unifying framework integrating life-history theory with physiology and behavior (Ricklefs and Wikelski 2002; Réale et al. 2010). POLS can be studied at multiple hierarchical levels of biological variation including among-species (Promislow and Harvey 1990), among-populations (e.g. Wikelski et al. 2003), and among-individuals (Réale et al. 2010). Individuals, populations, or species towards the fast end of the pace-of-life (POL) continuum are predicted to have a fast growth rate and invest heavily into early reproduction, resulting in a short lifespan (e.g. Stearns 1983; Promislow and Harvey 1990; Blackburn 1991; Owens and Bennett 1995). In addition, the fast POL strategy is predicted to have weakened immune responses and low physiological as well as behavioral stress responses (Réale et al. 2010). At the same time, individuals following a fast strategy should be aggressive, bold, and explorative, while also exhibiting low levels of parental care (Réale et al. 2010). The opposite trait expressions are expected for the slow end of the POL-continuum. Trait combinations following these predictions have been found between species (Ricklefs 1974; Tieleman et al. 2006; Wiersma et al. 2007; Speakman 2008; Williams et al. 2010; Jimenez et al. 2014) and even among-individuals within populations (e.g. Boon et al. 2007; Dammhahn 2012; Niemelä et al. 2013; Montiglio et al. 2014; Jacques-Hamilton et al. 2017). Nonetheless, these predictions have not been supported across studies (Závorka et al. 2015; Kim and Velando 2016). One explanation for this discrepancy might be that important systematic sources of variation, such as environment (Salzman et al. 2018, topical collection on Pace-of-life syndromes), ecology (Montiglio et al. 2018, topical collection on pace-of-life syndromes), or age- and sex-specific variation (Hämäläinen et al. 2018, Immonen et al. 2018; both in topical collection on Pace-of-life syndromes) have not often been considered.
Due to differences in the potential rate of reproduction, natural and sexual selection are expected to act differently on males and females, driving the evolution of sexual dimorphism (Bateman 1948; Parker 1979; Hedrick and Temeles 1989; Stearns 1992; Andersson 1994; Roff 2002; Fairbairn et al. 2007). Indeed, life-history traits, such as size at birth, growth and maturation rate, reproductive lifespan, and mortality rate commonly differ between the sexes across the animal kingdom (Clutton-Brock and Vincent 1991; Promislow 2003; Fairbairn et al. 2007; Bonduriansky et al. 2008; Maklakov and Lummaa 2013; Adler and Bonduriansky 2014). Assuming that physiological and behavioral traits are aligned with life-history traits on a general POL-continuum, we might predict sex-specific differences in the POL (i.e., sex-specific differences in trait means across all trait categories). Sex differences in POL may be an important driver generating individual-level variation in POLS (i.e., sex-specific trait correlations within populations; e.g., Hämäläinen et al. 2018, Immonen et al. 2018; both in topical collection on pace-of-life syndromes). Indeed, males and females of many species differ in their physiology (e.g., immunology, (Lee 2006; Restif and Amos 2010), metabolism (Nagy 1987; Blaxter 1989; Rønning et al. 2016), hormone profiles (Nelson 2011), and behavior (Smith and Blumstein 2008; Schuett et al. 2010), but the directionality of these differences within and across trait categories has not been formally tested. Fundamentally, differences between males and females in average trait expression may stem from sex-specific reproductive tactics caused by anisogamy (e.g., Bateman 1948; Maynard Smith 1958, 1982; Trivers 1972; Lehtonen et al. 2016; but see Kokko and Jennions 2008). Assuming anisogamy as a naïve null hypothesis would predict that males should be the overall faster sex in life-history, physiology, and behavioral traits. Importantly, however, sex-specific natural selection, as well as sexual selection, may act differently on trait expression and trait associations between the sexes (Arnqvist and Rowe 2005) and could therefore override the faster male POL predicted by anisogamy.
Environmental and social characteristics can generate or alter sex-specific selection that might enhance, mitigate, or even reverse differences dictated by anisogamy in the POL between the sexes (Hämäläinen et al. 2018, topical collection on Pace-of-life syndromes). Breeding biology, such as different mating systems and breeding strategies (semelparity and iteroparity), can affect the intensity of sexual selection (Andersson 1994; Bonnet 2011; Kralj-Fišer et al. 2011; Fisher et al. 2013; Janicke et al. 2016), and thus, the existence of sex-specific POL and POLS. For example, we might expect fast males in polygynous systems, fast females in polyandrous systems, and little differences between the sexes in monogamous systems since the intensity of the sex-specific selection varies across these different mating systems. The environment in which the organisms are studied (i.e., natural habitat or laboratory) can have an effect on selection, e.g., selection can be relaxed or modified in laboratory conditions (Frankham et al. 1986; Pelletier et al. 2009). Such effects may be sex-specific since (i) estimates of life-history are expected to be highly sensitive to environmental variation (absent in laboratory environments) (e.g., Kawasaki et al. 2008), and (ii) extrinsic mortality and mate competition may be lower in laboratory environments (Bonduriansky et al. 2008), potentially leading to differences in the expression of sex-specific POL and POLS between studies conducted in laboratory and wild environment.
Another source of systematic variation that might have led to contradictory evidence for POLS could be consistent differences in sex-specific variances across trait categories. Hence, in addition to mean differences, these may be important for interpreting sex-specific POL and POLS. Differences in phenotypic variance might indicate past episodes of sex-specific selection or be the result of stronger condition dependence (e.g., in the sex with a faster growth rate or larger body size; Rowe and Houle 1996; Råberg et al. 2005). Larger variances in one sex may also imply greater opportunities for sexual and natural selection which can facilitate the evolution of sex-specific POLS (Hämäläinen et al. 2018, topical collection on Pace-of-life syndromes). It has already been well established throughout the animal kingdom that males on average exhibit a higher variance in reproductive success (Janicke et al. 2016); however, much less is known in other traits. Some results indicate that males exhibit a higher phenotypic variance in morphological and behavioral traits (Wyman and Rowe 2014); however, a systematic review across POLS trait categories has not yet been conducted.
Here, we test whether the sexes generally differ in means and variances of traits that are predicted to fall along a fast-slow POL-continuum using meta-analytic methods. Given that data on sex-specific covariance structures across traits, a requirement for testing sex-specific POLS, is very scarce, we decided to focus on testing for sex-differences in trait categories, which are hypothesized to be part of POLS. Specifically, we tested the following questions: (1) Do males generally show a faster POL in adult life-history (e.g., lifespan), developmental life-history (e.g., developmental time or growth rate), physiological and behavioral traits, as predicted by anisogamy? (2) Do effects depend on the breeding strategy, mating system, or the study environment, as predicted by the literature? (3) Do the sexes generally, or for specific trait categories, differ in the amount of phenotypic variance?
Methods
Data collection
In our core search, we collected sex and population-specific phenotypic means and variances of POLS-related traits from published papers citing one or both of the seminal POLS papers by (Ricklefs and Wikelski 2002; Réale et al. 2010) until October 2016. To increase the sample size, we combined these studies with two existing databases collected to explicitly test for sex differences in homologous traits. “Database 1” was constructed to investigate differences in behavioral syndromes between males and females (AG et al. unpublished). This database consists of papers that cited three key reviews in the field of animal personality and behavioral syndrome research (Dall et al. 2004; Sih et al. 2004; Schuett et al. 2010), between the years 2010–2013. “Database 2” was originally constructed to compare male- and female-specific genetic (co)variance–matrices (MT et al. unpublished). Here, four main sources were used: papers cited in Poissant et al. (2009), papers cited in Wyman and Rowe (2014), and searches in Web of Science™ Core Collection and Biosis Previews® on the Web of Science platform. For these searches, the following search topics were used: “genetic correlation*” AND *sex*, refined by science categories: ecology OR genetics heredity OR evolutionary biology (for Web of Science™ Core Collection) and genetics OR evolution and adaptation (for Biosis Previews®), and restricting the search to papers published in 2008–2015.
We collected traits falling into three main POLS-trait categories: life-history and behavioral and physiological traits (Réale et al. 2010). Life-history traits were further divided into (1) developmental life-history traits and (2) adult life-history traits, because their importance for POLS might differ. Developmental life-history traits are life-history traits which are measured in immature, not yet reproducing individuals (growth rates and developmental times), while traits categorized as adult life-history are measured in sexually mature individuals (e.g., lifespan and age at first reproduction). We did not consider morphology, since the POLS hypothesis makes no predictions regarding such traits. A collection of the type of traits under each trait category and their directionality according to the POL-continuum is provided in Table S1.
From the databases, we excluded papers that: (1) were not empirical studies (e.g., reviews without data), (2) only included one sex in the analysis or where sexes were treated differently (e.g., subject to different experimental treatments), (3) involved an organism that could change sex (e.g., hermaphrodites), (4) studied plants or humans, and (5) only presented model outputs or reported summary statistics that were not convertible to mean and standard deviation. Furthermore, traits that were not on ratio scale or traits that were not convertible to raw scale had to be excluded. Principal component scores are composite variables composed of traits that might be associated with the POL-continuum in different directions and hence their interpretation may be difficult so these were also excluded. Database 1 and 2 also had some specific exclusion criteria (see Figs. S2–S3).
After removal of duplicates, we had a total of 1431 papers in the three databases (core database 702 papers, database 1 520 papers, database 2 227 papers). Exclusion according to the above-mentioned criteria left us with 109 papers, containing 423 trait values with mean and variance estimates for both sexes (Figs S1-S3). After further scrutinizing the data, 55 estimates from 19 studies were removed because of missing sample sizes, scale issues, or because the trait could not be placed along the fast-slow POL continuum (e.g., lifetime reproductive success). Furthermore, we excluded 10 additional estimates (e.g., song frequency) that were not related to the POLS hypothesis. Finally, we excluded 43 estimates to avoid pseudoreplication as their levels of dependency (see section below) would have been too complex to include in our models, leaving us with 90 papers and 315 trait estimates in our POL database (Fig. S4).
Effect size statistics
We calculated two different effect size statistics along with their sampling error variance: (1) the log response ratio (lnRR) (Hedges et al. 1999) and (2) log coefficient of variation ratio (lnCVR) (Nakagawa et al. 2015). The lnRR is a ratio between group means, which is free of the effects of group standard deviations (Osenberg et al. 1997; Hillebrand 2008). The lnCVR is the natural logarithm of the ratio between the coefficients of variation, from two groups, enabling a formal comparison of the variability of these groups. We choose to use lnCVR for comparing differences in variance between males and females because it accounts for corresponding changes in the mean; any change in variance could be explained simply by differences in the mean because of the strong mean–variance relationship we observed (Fig. S5; Nakagawa et al. 2015). In our case, our effect sizes were the ratios of the mean (lnRR) and coefficient of variation (CV) (lnCVR) between males and females. Sample sizes and standard deviations for male and female samples do not affect the calculation of effect sizes but are used to calculate each effect size sampling variance (Hedges et al. 1999; Nakagawa et al. 2015). Importantly, for lnRR, we categorized traits as falling along the fast–slow POL continuum in line with predictions from Réale et al. (2010) (see Table S1). Traits for which the directionality was not immediately clear (e.g., walking activity or scale score) were considered only if the authors of the original study indicated the relevance and directionality of the specific trait for the POLS hypothesis. The meaning of larger values for some traits can imply a slow POL (e.g., increased longevity), whereas others (e.g., growth rate or weight gain) would imply a fast POL. As such, we scrutinized individual traits (see Table S1) and inverted directionality of effect magnitude depending on whether higher values of a trait were considered to fall at the fast or slow end of the POL-continuum. In contrast, directionality for lnCVR is independent from the mean and so it was not necessary to invert effect sizes. For both effect sizes, positive estimates indicate faster female POL (lnRR) or larger female variance (lnCVR), whereas negative effect size estimates indicate faster male POL or larger variance in males.
Impact of mating system, breeding strategy, study environment on POL
Based on predictions in Hämäläinen et al. (2018, topical collection on pace-of-life syndromes), we selected three moderators for which we have sufficient data available that might affect the strength and direction of sex differences in life history and physiological and behavioral traits. We categorized species into different breeding strategies (iteroparous or semelparous) and mating systems (monogamy, polygyny, or promiscuity—only one of the studies had a polyandrous system and was therefore excluded from all analyses). We used social monogamy as a proxy for monogamy, because the majority of studies do not report the genetic mating system. We also categorized the study environment in which the study was conducted (“lab” for populations that have spent at least five generations in a laboratory environment and “wild” for populations studied in the wild or at most for five generations in the laboratory, see Charmantier and Garant (2005) meta-analysis with identical categorization).
When possible, we took the above information for each specific population (i.e., study), but in cases where this was not possible, we used general species information. To extract the species moderator category when it was not available, we searched Web of Knowledge and Google Scholar by combining the moderator term with the species scientific name (e.g., “mating system” AND “Parus major”).
Meta-analytic modeling
We analyzed lnRR and lnCVR using multi-level meta-analytic (MLMA—“intercept only” model) and multi-level meta-regression (MLMR—“predictor” model). Models were run in metafor (Viechtbauer 2010) and MCMCglmm (Hadfield 2010) to compare results and because Bayesian approaches provide greater flexibility in generating uncertainty around estimates. All models accounted for effect size sampling error variance along with study, species, and phylogenetic non-independence (Hadfield and Nakagawa 2010; Nakagawa and Santos 2012; Noble et al. 2017). We also estimated a residual variance in our MLMA models by including an observation-level random effect to calculate heterogeneity measures as described by Nakagawa and Santos (2012), as metafor does not estimate a residual variance by default.
To account for phylogenetic dependencies, we estimated phylogenetic relationships among species using the Open Tree of Life database (https://tree.opentreeoflife.org/; Hinchcliff et al. 2015), which constructs tree topology by synthesizing existing phylogenetic information, using taxonomic relationships when no phylogenetic data is available. From our tree topology (Fig. S6), we estimated a correlation matrix from branch lengths derived using Grafen’s method (Grafen 1989), which is akin to assuming the Brownian model of evolution. Branch lengths were estimated assuming node heights were raised to the power of 0.5. Importantly, Grafen’s method does not account for evolutionary divergence between taxa; however, it does provide a rough estimate of phylogenetic relationships by accounting for phylogenetic topology.
While metafor uses frequentist (i.e., likelihood) approaches to estimate parameters, MCMCglmm is Bayesian, and requires one to explicitly define prior probability distributions for parameters in the model. As such, we used inverse-Wishart priors (assuming V = 1 and nu = 0.002) on all random effects and uniform priors on fixed effects. MCMC approaches estimate parameters through a process similar to stochastic simulation, where parameters in the model are estimated and updated through an iterative sampling process using Bayes theorem. This requires the model to be run iteratively to generate a posterior probability distribution for each parameter. To accomplish this, we ran MCMCglmm models for a total of 500,000 iterations, using a burning period of 13,000 iterations. We thinned the MCMC chain by sampling every 100 iterations and ensured that we had an effective sample size for our posterior distribution of greater than 1000. We ensured that a thinning interval of 100 was sufficient by checking the autocorrelation between sampling lags to ensure that values were less than r = 0.10. In all cases, results from MCMCglmm closely matched results from metaphor (results not shown). We present model estimates along with their 95% confidence/credible intervals (CIs) throughout, interpreting estimates with CI’s excluding zero as statistically significant. Overall effects of interest were then back-transformed to percent increases for each sex where appropriate (Nakagawa and Schielzeth 2016). Below, we describe the specific models used to answer our three questions in detail.
Models 1 and 2: testing for overall faster and more variable males
To test for existence of overall faster POL in males (model 1), we estimated intercept-only MLMA model using lnRR as our response variable as follows:
where u is the overall mean lnRR; sk[i] is the study-specific random effect, k, applied to effect size, i; ap[i] is the phylogenetic random effect for species, j, applied to effect size, i; spj[i] is the species-specific effect, k, applied to effect size, i, and accounts for repeated species in the dataset. In contrast, ei is the observation-level or residual variance for effect size, i, and mi is the effect-size-specific sampling variance, which is known (i.e., inverse sampling variance for each effect size). In our model, all random effects are assumed to be normally distributed with a mean 0 and variance estimated from the model (i.e., ~N(0, \( {\sigma}_l^2\boldsymbol{I} \)), where \( {\sigma}_l^2 \) is any of the estimated variance parameters—species identity, study identity, phylogeny, or residual variance and I an identity matrix).
To test whether males have increased variance overall compared to females (model 2), independent of any changes in the mean between males and females, we modeled lnCVR as follows:
where u is the overall mean lnCVR and all other variables are defined the same as above.
Using our MLMA models (model 1 and 2), we calculated total heterogeneity (I2tot) as the proportion of variance among effect sizes after excluding the total sampling error variance (Higgins and Thompson 2002). We also estimated the proportion of variance explained by study (I2stdy) and phylogeny (I2phy—representing a phylogenetic signal) as a function of total variance (excluding sampling variance) (Nakagawa and Santos 2012). We calculated heterogeneity estimates from metafor (although estimates were similar from MCMCglmm: results not shown). To generate confidence intervals on our heterogeneity estimates, we used parametric bootstrapping (n simulations = 1500) using variance estimates derived from our MLMA models.
Our data also contained additional sources of non-independence at the within-study level (Nakagawa et al. 2017; Noble et al. 2017). For example, in some studies, different traits were measured on the same individuals (individual-level dependency) at the same time, whereas in other studies, effect sizes were derived from the same individuals measured at different time points (temporal dependency). To test if these additional levels of dependency influenced our results, we modified the identity matrix to a “dependency” matrix, D, such that ei ~N(0, \( {\sigma}_e^2\mathbf{D} \)), by including correlations at the residual (effect-size) level. Here, \( {\sigma}_e^2 \) is the residual variance and D is a matrix where off-diagonals for correlated effect sizes are assumed to be 0.5 (Noble et al. 2017). We assumed a medium-strength, positive correlation between dependent effect sizes, because the actual correlation was not known. In all cases, including D did not impact results from our MLMA models (results not shown), and therefore, we present models without correlation matrices.
Models 3–8: effects of mating system, breeding strategy, and study environment
We tested whether sexes differ in the direction of POL between POLS-related trait categories (adult life-history, developmental life-history, physiology and behavior) and whether the moderators affect the strength and direction of sex-specific POL.
We used MLMR models, which accounted for sampling variance, study, and phylogenetic non-independence in metafor and MCMCglmm. A full model approach was used (included all main effects for each moderator) including (1) trait category (4 levels: adult life-history, developmental life-history, physiology and behavior), (2) the mating system (3 levels: polygyny, promiscuity and monogamy), (3) breeding strategy (2 levels: iteroparity and semelparity), and (4) study environment (2 levels: wild and lab). We only estimated main effects, and not interactions, in our models, given the unbalanced number of trait estimates in many of the categories of the input variables, which would lead to spurious effect sizes. The model was structured as follows:
where ESi is the effect size, either lnRR or lnCVR; Χ is the design matrix; and β is a vector of estimated coefficients for each of the moderator levels described above. All other variables are described above (see the “Models 1 and 2: testing for overall faster and more variable males” section).
To test the robustness of our results, we again modified the identity matrix to include an assumed correlation (r = 0.5—as above) between dependent effect sizes (i.e., replacing I with D, such that ei ~N(0, \( {\sigma}_e^2\boldsymbol{D} \)), see the “Models 1 and 2: testing for overall faster and more variable males” section). However, models ignoring this dependency structure were much better supported (difference between model without and with residual correlation matrix: lnRR-∆AICc = 301.03 for both temporal and individual-level dependency in MLMR models; lnCVR-∆AICc = 20.77 for both temporal and individual-level dependencies). Therefore, we present models assuming effect sizes within studies were independent.
We generated marginal (unconditional) mean estimates, which gives us the mean response for each moderator level, adjusted for any other moderator level in the model. Unconditional means for a given level of a categorical variable (e.g., the level “wild” in the two-level categorical moderator “study environment”) are calculated by averaging (weighted average) the means of this level (i.e., wild) across different combinations of other variables in the model (e.g., each level of trait category: developmental life-history, “adult life-history,” “behavior,” and “physiology”). To accomplish this, and generate credible intervals for these means, we used the posterior distribution from MCMCglmm.
It is not yet confirmed whether all subclasses of behavioral or physiological traits are part of the POLS framework. To exclude the possibility that potential effects in trait categories were masked by different or lack of POL-directionality, we also conducted subset analyses [for both lnRR (behavior—model 5, physiology—model 6) and lnCVR (behavior—model 7, physiology—model 8)] using just behavioral and physiological traits. In the first subset analysis, we categorized behavioral traits into measures of “activity,” “aggression,” “boldness,” “exploration,” “parenting,” and “stress-coping” following Réale et al. (2007) (see Table S1). For the second subset analysis, we categorized physiological traits into four categories: immunology, baseline hormone measurement (hereafter “baseline”), and hormones after a stressor was applied (hereafter “stressed”), and “other” for physiology traits not falling into any of the other categories, following Réale et al. (2010).
Publication bias
We explored evidence for publication bias in lnRR and lnCVR—often resulting from non-significant results not being published (Nakagawa et al. 2017)—by exploring funnel plots of precision (inverse of sampling error variance) versus meta-analytic residuals from intercept-only MLMA. “Meta-analytic residuals” (sensu Nakagawa and Santos 2012) were calculated using MCMCglmm (Hadfield 2010) and were conditioned on the random effects accounting for non-independence. Symmetrical funnel plots suggest weak evidence for publication bias; however, funnel plots are often insufficient on their own (Nakagawa et al. 2017). Egger’s regression, in contrast, is a statistical test for funnel plot asymmetry (Egger et al. 1997). It explicitly tests the null hypothesis that the intercept of a regression between effect size residuals (i.e., meta-analytic residuals) standardized by the sampling error against precision (i.e., inverse sampling error variance) is equal to zero (Nakagawa and Santos 2012). A significant intercept is suggestive of publication bias (Egger et al. 1997). While we conducted Egger’s regression with both lnRR and lnCVR, it is important to recognize that there is no clear theoretical reason why lnCVR would be impacted by publication bias, and the utility of methods for assessing publication bias with variance measures has been questioned previously (Senior et al. 2015).
Results
Overall, we collected n = 315 trait mean estimates and standard deviations across 90 studies. Most estimates were derived from physiological traits (n = 157), followed by behavioral (n = 96), developmental life-history (n = 41), and adult life-history traits (n = 21). Traits were measured across 69 unique species, with the majority of estimates coming from birds (n = 133), followed by insects (n = 73), mammals (n = 57), fish (n = 27), and amphibians and reptiles (n = 25). Importantly, most estimates came from systems studied in the wild (lab: n = 123; wild: n = 192).
Models 1 and 2: testing for overall faster and more variable males
The overall effect for lnRR (est. = − 0.0498; 95% CI = − 0.1514; 0.0517) and lnCVR (est. = 0.0409; 95% CI = − 0.0054; 0.0873) were both small and not significantly different from zero. Nonetheless, total heterogeneity was high (lnRR: I2tot = 0.9990, 95% CI = 0.9988; 0.9991; lnCVR: I2tot = 0.9856, 95% CI = 0.9835; 0.9875). We also observed significant study level heterogeneity for both lnRR and lnCVR (lnRR: I2stdy = 0.5080, 95% CI = 0.4277; 0.5871; lnCVR: I2stdy = 0.1129, 95% CI = 0.0827; 0.1486), even though between-study differences explained little variance for lnCVR. Sub-group analyses for each trait category separately (same MLMA model structure as in Model 1 and 2) also suggested moderate to high between-study variance for developmental life-history and physiological traits, whereas variance among effects in behavioral and life-history traits were primarily driven by within-study effects (Table S2). We only observed a significant, albeit small, phylogenetic signal for lnRR (I2phy = 0.0719, 95% CI = 0.0485; 0.0990), whereas we did not observe a phylogenetic signal for lnCVR (I2phy = 0).
Model 3–8: effects of mating system, breeding strategy, and study environment
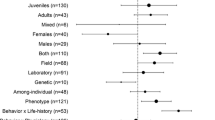
Interestingly, males tended to exhibit a faster POL in polygynous mating systems (Fig. 1a—lnRR = − 0.21, 95% CI = − 0.38; − 0.03; see also Table S3 for full model contrast results). This was true for developmental life-history, physiology, and behavior, but not adult life-history, whereas a significant effect did not exist in promiscuous and monogamous mating systems (Fig. 2). Neither mating system nor breeding strategy moderated differences in variance (lnCVR) between the sexes (Fig. 1b, Table S3). Breeding strategy (semelparity/iteroparity) did not moderate the effect sizes (lnRR and lnCVR) (Fig 1a, b).

Marginal mean estimates for a lnRR and b lnCVR. Point estimates and 95% credible intervals are provided. Sample size (N) is provided for each level of moderators. Positive values indicate faster POL in females

Predicted mean effect size from MLMR model (lnRR) across mating systems and study environment. Means are predicted for an “iteroparous” species. Point symbols correspond to the trait type (square = adult life-history; circle = developmental life-history; star = physiology and diamond = behavior). Point estimates and 95% confidence intervals are provided
Studies conducted in the wild had a significant negative lnRR (− 0.135, 95% CI = − 0.272; − 0.02), with males exhibiting a 12% overall faster POL compared to females. This suggests that males in the wild are more likely to exhibit faster POL than females, across averaged trait categories, mating systems, and breeding strategies (Fig. 1a). We did not observe any difference in variance (lnCVR) between wild and laboratory-based studies (Fig. 1b, Table S3).
Accounting for the trait categories and the three moderators, average lnRR showed a small, statistically non-significant, negative effect for behavioral, developmental life-history, and physiological traits (Fig. 1a, for the full model with non-marginalized coefficients; see Table S3), with males exhibiting a 6.22% (behavior) to 9.29% (developmental life-history) faster POL compared to females. The opposite was true of adult life-history traits, which were primarily composed of lifespan (14/21 effect sizes). Here, females exhibited, on average, a 21% faster POL (lnRR = 0.1948, 95% CI = − 0.0096; 0.4164) (Fig. 1a, Table S3). While females appeared to be more variable than males (lnCVR) across traits, these effects were quite small and 95% CI’s overlapped heavily with zero (Fig. 1b, Table S3).
When considering the study environments separately, the trait categories developmental life-history, behavior, and physiology were always clustered, while adult life-history deviates from this pattern (Fig. 2). Furthermore, polygynous mating systems were more likely to have males with traits (at least for developmental life-history, behavior, and physiology) that follow a faster POL, while the effect was weaker (but going in the same direction) in monogamous or promiscuous mating systems. Importantly, this pattern only holds for systems in the wild (Fig. 2). In the lab environment, females exhibit faster life-history across all mating systems.
In the models testing directional heterogeneity in the trait subclasses of behavior and physiology, we generally found no sex-specific differences except for parental care in lnRR (models 5 + 6; Fig. S7A-B, Table 1). Females showed a higher level of parental care, indicative of a slower POL, compared to males (Fig. S7A—lnRR = 0.56, 95% CI = 0.131; 0.960). Trait subclasses showed no effect for lnCVR (Models 7 + 8; Fig. S8A-B, Table 1).
Publication bias
We found no evidence for publication bias for lnRR and lnCVR given that meta-analytic residuals showed fairly symmetrical funnel plots (Fig. 3a, b). This was further supported by Egger’s regression with the intercepts not differing significantly from zero (lnRR: df = 313, t = − 0.861, p = 0.39; lnCVR: df = 313, t = − 1.86, p = 0.064).

Funnel plots of precision (inverse of sampling standard error) as a function of residuals from meta-analytic models for a lnRR and b lnCVR. Red vertical line indicates zero effect
Discussion
We provide the first quantitative overview testing for differences in the pace-of-life (POL) between the sexes across traits involved in POLS (i.e., adult life-history, developmental life-history, physiology and behavior). Although not exhaustive, our database is based on an unbiased sample of POLS-related studies. Our meta-analysis did not provide general support for overall faster POL in males (i.e., the anisogamy hypothesis), but males exhibit faster POL in polygynous study systems in the wild, at least for developmental life-history and physiological and behavioral traits.
While males often have the potential for a higher reproductive rate than females, our results show that they do not generally exhibit a faster POL in adult and developmental life-history and behavioral and physiological traits. These findings suggest that anisogamy is, as expected, not the only driving force for sex-specific POL and hence potentially also not for POLS. Our results do not suggest that the general predictions about POL differences at the individual level (Réale et al. 2010) can be translated to the between-sex level without considering the social and environmental characteristics of each population or species. Indeed, social and environmental characteristics have also been shown to influence POLS at the among-individual level (Salzman et al. 2018; Montiglio et al. 2018; Royauté et al. 2018; all in topical collection on pace-of-life syndromes).
When looking at patterns in specific POLS-trait categories, we found that behavior, physiology, and developmental life-history cluster together along the slow-fast POL continuum, while adult life-history does not. Faster male development is predicted to increase male resource holding potential, given that sexual selection often acts more strongly on males compared to females (Bateman 1948; Trivers 1972; Janicke et al. 2016; Lehtonen et al. 2016). Here, we find that it does not appear to be a general feature across species in our data. However, males in polygynous mating systems and in studies conducted in the wild have a faster developmental life-history than females (see next section). On the other hand, we also show that females overall had a faster POL in adult life-history (i.e., shorter life). The pace of developmental life-history going against the pace of adult life-history between the sexes contradicts population and species-level predictions from classical life-history theory, which expects short lifespan to be associated with fast growth and early age at maturity (Stearns 1992; Roff 2002; Bonduriansky et al. 2008; Brooks and Garratt 2017). Differences in lifespan between males and females can be both condition- and taxon-dependent and may result from asymmetric inheritance of sex chromosomes, differences in diet and physiology, maternal effects, sexual conflict, and sex-specific selective pressures (Gemmell et al. 2004; Tower and Arbeitman 2009; Maklakov and Lummaa 2013; Adler and Bonduriansky 2014; De Lisle and Rowe 2015; English and Uller 2016), which may explain this unexpected finding. Of course, differences in adult survival may also arise due to early disappearance of one sex from the population. Adult survival estimates in our data comprise a mixture of different taxonomic groups, breeding strategies, and wild and laboratory studies, which should diminish this problem. Nevertheless, the observed differences in adult life-history between the sexes should be interpreted with care, given the small number of effect sizes in our data. Sex-specific differences in adult life-history may affect sex-specific patterns of trait covariance in the context of POLS; however, more empirical research on the magnitude and extent of sex-specific trait covariance patterns across populations and species is needed to investigate this.
Some evidence indicates that males and females frequently differ in the mean expression of behaviors related to POLS (reviewed in Schuett et al. 2010). Sexual selection resulting in different selection pressures between males and females has been argued to explain these differences between the sexes (Andersson 1994; Schuett et al. 2010). However, we did not find any indication that sex-differences in behavior are a general phenomenon across diverse species. The only significant difference between sexes we find is that females generally show more parental behavior compared to males. This is not surprising given that females usually produce far fewer gametes and hence are expected to invest more into post-fertilization care (Trivers 1972). Although differences in parental care may occur for other reasons than the role in life history trade-offs, parental care behavior has been identified as a key driver in linking individual differences in behavior with reproductive success (Mutzel et al. 2013), which suggests a clear link to POLS. Unexpectedly, we also found no indication for sex differences in physiological traits (baseline and challenged HPA-axis related hormones and immune parameters). Sex differences in anatomy and function of HPA-axis regulatory mechanisms are well documented for mammals and stem from sexual dimorphism in the central nervous system affecting physiology and behavior through endocrinology (Rhodes and Rubin 1999). Individual differences in endocrinological processes and traits that are strongly affected by these, e.g. growth rate, have been suggested, and meta-analytically shown, to be the underlying driving forces for consistent individual differences in behavior (Stamps 2007; Biro and Stamps 2008; for meta-analytical approach Niemelä and Dingemanse 2018). In addition, these processes have also been proposed to underlie sex differences in POLS (Immonen et al. 2018). Our results, however, suggest that sex-differences in the HPA-axis related hormone concentrations cannot be generalized across taxa. Sex differences in immunity are also often reported for mammals and birds (Demas and Nelson, 2012; Roved et al. 2016). Although sexual selection affecting the trade-off between reproduction and self-maintenance is assumed to influence male immune responses negatively, we found no sex-differences.
The sex-specific POL predicted by anisogamy can be altered or even overridden by contrasting selection created by, for example, different mating systems or breeding strategies (Hämäläinen et al. 2018, topical collection on pace-of-life syndromes). Indeed, we found that males of polygynous mating systems showed a faster POL than females, as predicted, presumably because sexual and/or natural selection is stronger on males in polygynous species (Greenwood 1980; Wingfield et al. 1990; Clutton-Brock and Isvaran 2007; Brooks and Garratt 2017). On the other hand, social monogamy, often entailing biparental care, is thought to reduce the level of antagonistic selection and conflict between the sexes, leading to more similar life-history optima (Klug et al. 2013). Monogamy is therefore predicted to result in reduced dimorphism of POLS traits, such as lifespan or development (Promislow 1992; Liker and Szekely 2005)—a pattern supported by our results. Also, no sex differences were found in promiscuous mating systems, although we would have expected a more pronounced sexual dimorphism driven by a strong sexual conflict (Hämäläinen et al. 2018, topical collection on pace-of-life syndromes). The sexual conflict should be particularly strong in promiscuous mating systems where males harass females into matings, in opposite to systems where females mate voluntarily, in which case we would expect less dimorphic POLS traits between the sexes. If our database contains a mixture of these two types of promiscuous mating systems, this could explain the lack of sex differences in this group. We therefore advice future studies to specifically look into these differences. Another potential confounding factor might be the presence or absence of parental care. In promiscuous insect systems, where parental care is most often absent, sexual conflict can be mitigated, leading to more monomorphic POL strategies, while in promiscuous mammals and birds, where parental care is female biased, sex differences should be attenuated (Schlicht and Kempenaers 2013; Clutton-Brock 2016). The number of possible breeding attempts per life time combined with promiscuity is also expected to mitigate POL differences between sexes (Hämäläinen et al. 2018, topical collection on Pace-of-life syndromes). In our data, breeding strategy, averaged across all mating systems, did not affect POL in the sexes.
Our results did show a general effect of study environment, with males in the wild having a faster POL. This may be explained by the “robust-male hypothesis” (Bonduriansky et al. 2008), if males in the wild experience higher mortality rates. This hypothesis states that the sex experiencing higher extrinsic mortality in the wild will experience higher condition-dependent selection when brought into benign laboratory conditions, potentially reversing the POL between males and females (Hämäläinen et al. 2014). In artificial environments, trait means might change in an unpredictable way (e.g., individual level behaviors: Niemelä and Dingemanse 2014), because traits measured under laboratory conditions are not under similar constraints or selection, potentially leading to the expression of genetic and phenotypic variation that would have been selected against in the wild (Ghalambor et al. 2007; Schlichting 2008).
We found little evidence for sex-specific effects in phenotypic variances. We could have predicted overall higher variances in males resulting from a number of mechanisms. First, males often are under stronger sexual selection than females, causing a higher variance in reproductive success (Janicke et al. 2016), and potentially therefore also in traits related to reproductive success. Second, males inherit the mitochondrial genome from their mothers, which may contain detrimental alleles that increase variance in male traits (i.e., “Mother’s curse”), particularly in traits related to metabolism (Gemmell et al. 2004; Innocenti et al. 2011). Lastly, stressful environments have been suggested to have important consequences on trait variance (O’Dea et al. 2016), and the sex with higher condition dependence should be more affected by stressful conditions. Males in systems with high male competition (e.g., polygyny) often experience more stressful environments than females and should therefore have a higher variance (e.g., Rowe and Houle 1996). Despite these predictions, we did not find any variance differences between the sexes across any of the trait categories, mating systems, or study environments. If anything, the variances seem to be slightly higher in females. One possible explanation for the lack of a clear pattern in variances could be the mixture of different sex-chromosome systems represented in our data. The sex-chromosome hypothesis states that in species with dosage compensation, the heterogametic sex should show a larger trait variance compared to the homogametic sex (James 1973; Reinhold and Engqvist 2013).
In conclusion, our meta-analysis did not find that males have an overall faster POL, although they were overall faster in polygynous mating systems and in studies conducted in the wild (i.e., more natural environments). This indicates that future studies should take into account the mating system and study environment when trying to understand sex-specific patterns in POL and POLS. Another promising avenue for future research is also to look at how such social and environmental factors interact and affect sex-specific POL and POLS. Surprisingly, we found that adult life-history deviates from all the other trait categories (behavior, physiology and developmental life-history), which all cluster together along the fast-slow POL continuum. In the POLS hypothesis, life history traits play a key role because the framework aims to predict how individuals and sexes mediate the trade-off between current and future reproduction (Dammhahn et al. 2018, topical collection on Pace-of-life syndromes). Thus, life history traits must always be included in studies investigating POLS. Our data clearly show that future studies need to acknowledge that the conclusion about POLS might differ depending on whether the chosen life-history trait represents developmental or adult life-history traits. The lack of differentiation between sexes in the POLS literature, so far, may have been caused by a general focus on females in the classical life-history literature. Our results encourage a more thorough investigation into sex-specific POL, how such differences translate into different pace-of-life syndromes, and the implications of these trait covariances for population ecology and evolution.
References
Adler M, Bonduriansky R (2014) Sexual conflict, life span, and aging. CSH Perspect Biol 6:a017566
Andersson M (1994) Sexual selection. Princeton University Press, Princeton
Arnqvist G, Rowe L (2005) Sexual conflict. Princeton University Press, Princeton
Bateman A (1948) Intra-sexual selection in Drosophila. Heredity 2:349–368
Biro PA, Stamps JA (2008) Are animal personality traits linked to life-history productivity? Trends Ecol Evol 23:361–368
Blackburn T (1991) Evidence for a `fast-slow’continuum of life-history traits among parasitoid hymenoptera. Funct Ecol 5:65–74
Blaxter K (1989) Energy metabolism in animals and man. Cambridge University Press, Cambridge
Bonduriansky R, Maklakov A, Zajitschek F, Brooks R (2008) Sexual selection, sexual conflict and the evolution of ageing and life span. Funct Ecol 22:443–453
Bonnet X (2011) The evolution of semelparity. In: Aldridge RD, Sever DM (eds) Reproductive biology and phylogeny of snakes. CRC Press, Boca Raton, pp 645–672
Boon A, Réale D, Boutin S (2007) The interaction between personality, offspring fitness and food abundance in North American red squirrels. Ecol Lett 10:1094–1104
Brooks R, Garratt M (2017) Life history evolution, reproduction, and the origins of sex dependent aging and longevity. Ann N Y Acad Sci 1389:92–107
Charmantier A, Garant D (2005) Environmental quality and evolutionary potential: lessons from wild populations. Proc R Soc Lond B 272:1415–1425
Clutton-Brock T (2016) Mammal societies. John Wiley & Son Inc., Chichester
Clutton-Brock T, Isvaran K (2007) Sex differences in ageing in natural populations of vertebrates. Proc R Soc Lond B 274:3097–3104
Clutton-Brock T, Vincent A (1991) Sexual selection and the potential reproductive rates of males and females. Nature 351:58–60
Dall SRX, Houston AL, McNamara JM (2004) The behavioural ecology of personality: consistent individual differences from an adaptive perspective. Ecol Lett 7:734–739
Dammhahn M (2012) Are personality differences in a small iteroparous mammal maintained by a life-history trade-off? Proc R Soc Lond B 279:2645–2651
Dammhahn M, Dingemanse NJ, Niemelä PT, Réale D (2018) Pace-of-life syndromes: a framework for the adaptive integration of behaviour, physiology and life-history. Behav Ecol Sociobiol 72:62
De Lisle SP, Rowe L (2015) Ecological character displacement between the sexes. Am Nat 186:693–707
Demas GE, Nelson RJ (2012) Ecoimmunology. In: Oxford. Oxford University Press
Egger M, Smith GD, Schneider M, Minder C (1997) Bias in meta-analysis detected by a simple, graphical test. Brit Med J 315:629–634
English S, Uller T (2016) Does early-life diet affect longevity? A meta-analysis across experimental studies. Biol Lett 12:20160291
Fairbairn D, Blanckenhorn W, Székely T (2007) Sex, size and gender roles: evolutionary studies of sexual size dimorphism. Oxford University Press, Oxford
Fisher D, Dickman C, Jones M, Blomberg S (2013) Sperm competition drives the evolution of suicidal reproduction in mammals. P Natl Acad Sci USA 110:17910–17914
Frankham R, Hemmer H, Ryder OA, Cothran EG, Soulé ME, Murray ND, Snyder M (1986) Selection in captive populations. Zoo Biol 5:127–138
Gemmell NJ, Metcalf VJ, Allendorf FW (2004) Mother’s curse: the effect of mtDNA on individual fitness and population viability. Trends Ecol Evol 19:238–244
Ghalambor CK, McKay JK, Carroll SP, Reznick DN (2007) Adaptive versus non-adaptive phenotypic plasticity and the potential for contemporary adaptation in new environments. Funct Ecol 21:394–407
Grafen A (1989) The phylogenetic regression. Phil Trans R Soc B 326:119–157
Greenwood P (1980) Mating systems, philopatry and dispersal in birds and mammals. Anim Behav 28:1140–1162
Hadfield JD (2010) MCMC methods for multi-response generalized linear mixed models: the MCMCglmm R package. J Stat Softw 33:1–22
Hadfield JD, Nakagawa S (2010) General quantitative genetic methods for comparative biology: phylogenies, taxonomies and multi-trait models for continuous and categorical characters. J Evol Biol 23:494–508
Hämäläinen A, Dammhahn M, Aujard F, Eberle M, Hardy I, Kappeler PM, Perret M, Schliehe-Diecks S, Kraus C (2014) Senescence or selective disappearance? Age trajectories of body mass in wild and captive populations of a small-bodied primate. Proc R Soc B 281:20140830
Hämäläinen A, Immonen E, Tarka M, Schuett W (2018) Evolution of sex-specific pace-of-life syndromes: causes and consequences. Behav Ecol Sociobiol 72:50
Hedges LV, Gurevitch J, Curtis PS (1999) The meta-analysis of response ratios in experimental ecology. Ecology 80:1150–1156
Hedrick A, Temeles E (1989) The evolution of sexual dimorphism in animals: hypotheses and tests. Trends Ecol Evol 4:136–138
Higgins JPT, Thompson SG (2002) Quantifying heterogeneity in a meta-analysis. Stat Med 12:1539–1558
Hillebrand H (2008) Meta-analysis in ecology. Encyclopedia of Life Sciences, Wiley, Chichester
Hinchcliff CE, Smith SA, Allman JF et al (2015) Synthesis of phylogeny and taxonomy into a comprehensive tree of life. P Natl Acad Sci USA 112:12764–12769
Immonen E, Hämäläinen A, Schuett W, Tarka M (2018) Evolution of sex-specific pace-of-life syndromes: genetic architecture and physiological mechanisms. Behav Ecol Sociobiol 72:60
Innocenti P, Morrow EH, Dowling D (2011) Experimental evidence supports a sex-specific selective sieve in mitochondrial genome evolution. Science 332:845–848
Jacques-Hamilton R, Hall M, Buttemer W, Matson KD, Gonçalves da Silva A, Mulder RA, Peters A (2017) Personality and innate immune defenses in a wild bird: evidence for the pace-of-life hypothesis. Horm Behav 28:31–40
James JW (1973) Covariances between relatives due to sex-linked genes. Biometrics 29:584–588
Janicke T, Haderer IK, Lajeunesse MJ, Anthes N (2016) Darwinian sex roles confirmed across the animal kingdom. Sci Adv 2:e1500983
Jimenez A, Cooper-Mullin C, Calhoon E, Williams J (2014) Physiological underpinnings associated with differences in pace of life and metabolic rate in north temperate and neotropical birds. J Comp Physiol B 184:545–561
Kawasaki N, Brassil C, Brooks R, Bonduriansky R (2008) Environmental effects on the expression of life span and aging: an extreme contrast between wild and captive cohorts of Telostylinus angusticollis (Diptera: Neriidae). Am Nat 172:346–357
Kim S-Y, Velando A (2016) Unsociable juvenile male three-spined sticklebacks grow more attractive. Behav Ecol Sociobiol 70:975–980
Klug H, Bonsall MB, Alonzo SH (2013) Sex differences in life history drive evolutionary transitions among maternal, paternal, and bi-parental care. Ecol Evol 3:792–806
Kokko H, Jennions M (2008) Parental investment, sexual selection and sex ratios. Evol Biol 21:919–948
Kralj-Fišer S, Gregoric M, Zhang S, Li D, Kuntner M (2011) Eunuchs are better fighters. Anim Behav 81:933–939
Lee K (2006) Linking immune defenses and life history at the levels of the individual and the species. Integr Comp Biol 46:1000–1015
Lehtonen J, Parker G, Schärer L (2016) Why anisogamy drives ancestral sex roles. Evolution 70:1129–1135
Liker A, Székely T (2005) Mortality costs of sexual selection and parental care in natural populations of birds. Evolution 59:890–897
Maklakov A, Lummaa V (2013) Evolution of sex differences in lifespan and aging: causes and constraints. Bioessays 35:717–724
Maynard Smith J (1958) Prolongation of the life of Drosophila subobscura by a brief exposure of adults to a high temperature. Nature 181:496–497
Maynard Smith J (1982) Evolution and the theory of games. Cambridge University Press, Cambridge
Montiglio P-O, Garant D, Pelletier F, Réale D (2014) Intra-individual variability in fecal cortisol metabolites varies with lifetime exploration and reproductive life history in eastern chipmunks (Tamias striatus). Behav Ecol Sociobiol 69:1–11
Montiglio P-O, Dammhahn M, Dubuc Messier G, Réale D (2018) The pace-of-life syndrome revisited: the role of ecological conditions and natural history on the slow-fast continuum. Behav Ecol Sociobiol
Mutzel A, Dingemanse NJ, Araya-Ajoy YG, Kempenaers B (2013) Parental provisioning behaviour plays a key role in linking personality with reproductive success. Proc R Soc B 280:20131019
Nagy K (1987) Field metabolic rate and food requirement scaling in mammals and birds. Ecol Monogr 57:111–128
Nakagawa S, Santos ESA (2012) Methodological issues and advances in biological meta-analysis. Evol Ecol 26:1253–1274
Nakagawa S, Schielzeth H (2016) Coefficient of determination R2 and intra-class correlation coefficient ICC from generalized linear mixed-effects models revisited and expanded. J R Soc Interface 14:20170213
Nakagawa S, Poulin R, Mengersen K, Reinhold K, Engqvist L, Lagisz M, Senior AM (2015) Meta-analysis of variation: ecological and evolutionary applications and beyond. Methods Ecol Evol 6:143–152
Nakagawa S, Noble DWA, Senior AM, Lagisz M (2017) Meta-evaluation of meta-analysis: ten appraisal questions for biologists. BMC Biol 15:18
Nelson RJ (2011) An introduction to behavioral endocrinology. Sinauer Associates, Sunderland
Niemelä PT, Dingemanse NJ (2014) Artificial environments and the study of ‘adaptive’ personalities. Trends Ecol Evol 29:245–247
Niemelä PT, Dingemanse NJ (2018) Meta-analysis reveals weak associations between intrinsic state and personality. Proc R Soc B 285:20172823
Niemelä PT, Dingemanse NJ, Alioravainen N, Vainikka A, Kortet R (2013) Personality pace-of-life hypothesis: testing genetic associations among personality and life history. Behav Ecol 24:35–941
Noble DWA, Lagisz M, O’Dea RE, Nakagawa S (2017) Non-independence and sensitivity analyses in ecological and evolutionary meta-analyses. Mol Ecol 26:2410–2425
O’Dea RE, Noble DWA, Johnson SL, Hesselson D, Nakagawa S (2016) The role of non-genetic inheritance in evolutionary rescue: epigenetic buffering, heritable bet hedging and epigenetic traps. Environ Epigenet 2:dvv014
Osenberg C, Sarnelle O, Cooper S (1997) Effect size in ecological experiments: the application of biological models in meta-analysis. Am Nat 150:798–812
Owens I, Bennett P (1995) Ancient ecological diversification explains life-history variation among living birds. Proc R Soc Lond B 261:227–232
Parker G (1979) Sexual selection and sexual conflict. In: Blum MS, Blum NA (eds) Sexual selection and reproductive competition in insects. Academic Press, New York, pp 123–166
Pelletier F, Réale D, Watters J, Boakes EH, Garant D (2009) Value of captive populations for quantitative genetics research. Trends Ecol Evol 24:263–270
Poissant J, Wilson AJ, Coltman DW (2009) Sex-specific genetic variance and the evolution of sexual dimorphism: a systematic review of cross-sex genetic correlations. Evolution 64:97–107
Promislow D (1992) Costs of sexual selection in natural populations of mammals. Proc R Soc Lond B 247:203–210
Promislow D (2003) Mate choice, sexual conflict, and evolution of senescence. Behav Genet 33:191–201
Promislow D, Harvey P (1990) Living fast and dying young: a comparative analysis of life-history variation among mammals. J Zool 220:417–437
Råberg L, Stjernman M, Nilsson J-A (2005) Sex and environmental sensitivity in blue tit nestlings. Oecologia 145:496–503
Réale D, Reader S, Sol D, Mcdougall PT, Dingemanse NJ (2007) Integrating animal temperament within ecology and evolution. Biol Rev 82:291–318
Réale D, Garant D, Murray M, Humphries MM, Bergeron P, Careau V, Montiglio P-O (2010) Personality and the emergence of the pace-of-life syndrome concept at the population level. Phil Trans R Soc B 365:4051–4063
Reinhold K, Engqvist L (2013) The variability is in the sex chromosomes. Evolution 67:3662–3668
Restif O, Amos W (2010) The evolution of sex-specific immune defences. Proc R Soc Lond B 277:2247–2255
Rhodes ME, Rubin RT (1999) Functional sex differences (‘sexual diergism’) of central nervous system cholinergic systems, vasopressin , and hypothalamic – pituitary – adrenal axis activity in mammals?: a selective review. Brain Res Rev 30:135–152
Ricklefs R (1974) Energetics of reproduction in birds. In: Paynter RA Jr (ed) Avian energetics. Proc. Nuttall Ornithological Club, No. 15, Cambridge, MA, pp 152–292
Ricklefs R, Wikelski M (2002) The physiology/life-history nexus. Trends Ecol Evol 17:462–468
Roff D (2002) Life history evolution. Sinauer Associates Inc., Sunderland
Rønning B, Broggi J, Bech C, Moe B, Ringsby TH, Pärn H, Hagen IJ, Sæther BE, Jensen H (2016) Is basal metabolic rate associated with recruit production and survival in free-living house sparrows? Funct Ecol 30:1140–1148
Roved J, Westerdahl H, Hasselquist D (2016) Sex differences in immune responses: hormonal effects, antagonistic selection, and evolutionary consequences. Horm Behav 88:95–105
Rowe L, Houle D (1996) The lek paradox and the capture of genetic variance by condition dependent traits. Proc R Soc Lond B 263:1415–1421
Royauté R, Berdal MA, Garrison CR, Dochtermann NA (2018) Paceless life? A meta-analysis of the pace-of-life syndrome hypothesis. Behav Ecol Sociobiol 72:64
Salzman TC, McLaughlin AL, Westneat DF, Crowley PH (2018) Energetic trade-offs and feedbacks between behavior and metabolism influence correlations between pace-of-life attributes. Behav Ecol Sociobiol 72:54
Schlicht E, Kempenaers B (2013) Effects of social and extra-pair mating on sexual selection in blue tits (Cyanistes caeruleus). Evolution 67:1420–1434
Schlichting CD (2008) Hidden reaction norms, cryptic genetic variation, and evolvability. Ann N Y Acad Sci 1133:187–203
Schuett W, Tregenza T, Dall SRX (2010) Sexual selection and animal personality. Biol Rev 85:217–246
Senior AM, Nakagawa S, Lihoreau M, Simpson SJ, Raubenheimer D (2015) An overlooked consequence of dietary mixing: a varied diet reduces interindividual variance in fitness. Am Nat 186:649–659
Sih A, Bell A, Johnson JC (2004) Behavioral syndromes: an ecological and evolutionary overview. Trends Ecol Evol 19:372–378
Smith BR, Blumstein DT (2008) Fitness consequences of personality: a meta-analysis. Behav Ecol 19:448–455
Speakman J (2008) The physiological costs of reproduction in small mammals. Phil Trans R Soc B 363:375–398
Stamps JA (2007) Growth-mortality tradeoffs and “personality traits” in animals. Ecol Lett 10:355–363
Stearns S (1983) The influence of size and phylogeny on patterns of covariation among life-history traits in the mammals. Oikos 41:173–187
Stearns S (1992) The evolution of life histories. Oxford University Press, Oxford
Tieleman B, Dijkstra T, Lasky J, Mauck RA, Visser GH, Williams JB (2006) Physiological and behavioural correlates of life-history variation: a comparison between tropical and temperate zone house wrens. Funct Ecol 20:491–499
Tower J, Arbeitman M (2009) The genetics of gender and life span. J Biol 8(38):38
Trivers R (1972) Sexual selection and the descent of man. Aldine de Gruyter, New York
Viechtbauer W (2010) Conducting meta-analyses in R with the metafor package. J Stat Softw 36:1–48
Wiersma P, Muñoz-Garcia A, Walker A, Williams J (2007) Tropical birds have a slow pace of life. P Natl Acad Sci USA 104:9340–9345
Wikelski M, Spinney L, Schelsky W, Scheuerlein A, Gwinner E (2003) Slow pace of life in tropical sedentary birds: a common-garden experiment on four stonechat populations from different latitudes. Proc R Soc Lond B 270:2383–2388
Williams J, Miller R, Harper J, Wiersma P (2010) Functional linkages for the pace of life, life-history, and environment in birds. Integr Comp Biol 50:855–868
Wingfield J, Hegner R, Dufty AJ, Ball G (1990) The “challenge hypothesis”: theoretical implications for patterns of testosterone secretion, mating systems, and breeding strategies. Am Nat 136:829–846
Wyman MJ, Rowe L (2014) Male bias in distributions of additive genetic, residual, and phenotypic variances of shared traits. Am Nat 184:326–337
Závorka L, Aldvén D, Näslund J, Höjesjö J, Johsson JI (2015) Linking lab activity with growth and movement in the wild: explaining pace-of-life in a trout stream. Behav Ecol 26:877–884
Acknowledgments
We thank the organizers and participants of the POLS workshop for stimulating discussions. We also thank Jon Wright, Irja Ratikainen, Elina Immonen, Anni Hämäläinen, and Wiebke Schuett for the fruitful discussions and comments on an early version of the manuscript. We also thank two anonymous reviewers and the editor for helpful comments that improved the manuscript.
Funding
The authors thank the VW Foundation for funding the two workshops “Towards a general theory of the pace-of-life syndrome” which inspired this study. MT was funded by the Research Council of Norway (SFF-III 223257). AG was supported by a Leopoldina Postdoc Scholarship (LPDS 2015-04). DWAN was supported by an ARC Discovery Early Career Research Award (DECRA) (DE150101774) and SN an ARC Future Fellowship (FT130100268).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Communicated by D. Réale
This article is a contribution to the Topical Collection Pace-of-life syndromes: a framework for the adaptive integration of behaviour, physiology, and life-history
Guest Editors: Melanie Dammhahn, Niels J. Dingemanse, Petri T. Niemelä, Denis Réale
Electronic supplementary material
ESM 1
(DOCX 1136 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Tarka, M., Guenther, A., Niemelä, P.T. et al. Sex differences in life history, behavior, and physiology along a slow-fast continuum: a meta-analysis. Behav Ecol Sociobiol 72, 132 (2018). https://doi.org/10.1007/s00265-018-2534-2
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00265-018-2534-2