
Abstract
Understanding the aggregation and habitat use patterns of a species can aid the formulation and improved design of management strategies aiming to conserve vulnerable populations. We used photo-identification techniques and a novel remote underwater camera system to examine the population sizes, patterns of residency and habitat use of oceanic (Mobula birostris) and reef (Mobula alfredi) manta rays in Seychelles (5.42°S; 53.30°E) between July 2006 and December 2018. Sightings of M. birostris were infrequent (n = 5), suggesting that if aggregation areas for this species exist, they occur outside of the boundary of our study. A total of 236 individual M. alfredi were identified across all surveys, 66.5% of which were sighted at D’Arros Island (Amirante Group) and 22.5% at St. François Atoll (Alphonse Group). Males and females were evenly represented within the identified population. M. alfredi visited a cleaning station at D’Arros Island less frequently during dawn and dusk than at midday, likely due to the adoption of a crepuscular foraging strategy. The remote and isolated nature of the Amirante and Alphonse Group aggregation areas, coupled with the lack of a targeted mobulid fishery in Seychelles, suggests that with appropriate regulations and monitoring, the marine protected areas gazetted within these two groups will benefit the conservation of M. alfredi in Seychelles.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Mobulid rays (family Mobulidae; Couturier et al. 2012; Hosegood et al. 2020) are medium-to-large (1.1–6.8 m disc widths; White et al. 2018) filter-feeding elasmobranchs that often predictably aggregate in coastal waters or at oceanic islands and seamounts in tropical and temperate locations around the world (Couturier et al. 2012; Stevens et al. 2019). There is a general consensus that populations of mobulids are now declining on a global scale (Croll et al. 2016; Lawson et al. 2017; Stewart et al. 2018; Strike et al. 2022). Fisheries targeting mobulids that supply their gill plates to the market for traditional Asian medicines have hastened this trend (Croll et al. 2016; O’Malley et al. 2017; Rohner et al. 2017), although bycatch, accidental entanglement and boat strikes are also sources of mortality (Croll et al. 2016; Germanov et al. 2019; McGregor et al. 2019). The conservative life history strategy of these species, including low fecundity, late maturation and long gestation periods (Couturier et al. 2012), prolongs the period of time required for populations to recover from the loss of individuals. This highlights the need to identify and understand the role that critical habitats play in the life history of mobulid rays (Stewart et al. 2018).
Manta rays are the largest of the mobulids (Stewart et al. 2018; Stevens et al. 2019). The oceanic manta ray (Mobula birostris) attains a maximum disc width of 6.8 m and is often observed at oceanic islands and along tropical and temperate coastlines, whereas reef manta rays (Mobula alfredi) have smaller disc widths (maximum 5.5 m) and frequently aggregate along tropical coastlines and at coral atolls (Marshall et al. 2009; Kashiwagi et al. 2011; Armstrong et al. 2020; Fonseca-Ponce et al. 2022). Oceanic and reef manta rays are, respectively, listed as ‘Endangered’ and ‘Vulnerable’ on the International Union for the Convention of Nature’s Red List (Marshall et al. 2018, 2022), on Appendix II of the Convention on International Trade in Endangered Species, and on Appendices I and II of the Convention on the Conservation of Migratory Species.
Aggregations of manta rays are thought to be driven by food availability (Couturier et al. 2012) and the presence of cleaning stations (Couturier et al. 2018; Peel et al. 2020) that provide an opportunity for individuals to have external parasites removed from their bodies by resident reef fishes (Potts 1973; Armstrong et al. 2021). Cleaning stations also facilitate social interactions between individuals, including courtship and mating behaviours (Deakos et al. 2011; Stevens et al. 2018). The significance of these stations is further demonstrated by the repeat visits of the same individuals over prolonged periods of time (years), despite their ability to routinely travel large distances (10s–100s km; Germanov and Marshall 2014; Couturier et al. 2018; Andrzejaczek et al. 2020). Sighting predictability of manta rays at these aggregation sites has greatly benefited global efforts to determine the drivers of their behaviour and movement in recent decades, as sampling can take advantage of aggregation events (Deakos et al. 2011; Jaine et al. 2012; Peel et al. 2019a). The information collected during targeted surveys is particularly valuable in dispersed, archipelagic reef systems such as Seychelles, where it can be challenging to obtain continuous access to remote study sites throughout the year due to environmental (e.g. monsoonal seasons) and logistical (e.g. limited or lack of infrastructure on islands) factors.
The presence of unique patterns of pigmentation on the ventral surface of individual manta rays allows photo-identification for the differentiation of individuals to be used to assess the size and distributions of populations at aggregation sites (Deakos et al. 2011; Marshall et al. 2011; Stevens et al. 2018; Germanov et al. 2019) over long periods of time (> 20 years; Marshall and Pierce 2012; Couturier et al. 2014; Stevens 2016; Marie 2022). Environmental data can also be collected simultaneously with sighting records to determine potential drivers of movements (O’Shea et al. 2010; Rohner et al. 2013; Couturier et al. 2018; Harris and Stevens 2021). In addition, photo-identification photographs can be collected by citizen scientists to facilitate sampling across large spatial scales (Town et al. 2013; Couturier et al. 2014; Germanov and Marshall 2014; Armstrong et al. 2019), providing insight into the broader (> 100 km) patterns of movement of individuals and their habitat range. Collectively, these data can expand our understanding of manta ray populations and inform conservation and management strategies aimed at protecting them (Nicholson-Jack et al. 2021; Strike et al. 2022).
Despite facing increasing pressures from targeted and small-scale fisheries in the Western Indian Ocean (Temple et al. 2019), little is currently known about manta ray populations in Seychelles, which aggregate around remote islands within the archipelago. Such isolated aggregations present an opportunity to not only examine the biology and ecology of manta rays in the absence of significant human impact, but also inform management and conservation decisions in a proactive, rather than reactive, manner. Considering this, we used photo-identification techniques and a remote underwater monitoring system to examine population size and the patterns and potential drivers of residency for M. birostris and M. alfredi in Seychelles. Primary efforts were focussed on the two aggregation areas identified for M. alfredi at D’Arros Island and St. Joseph Atoll (collectively referred to as D’Arros Island) and an aggregation area at St. François Atoll. Specifically, we aimed to (1) use photo-identification data collected opportunistically throughout Seychelles, and during targeted surveys at D’Arros Island and St. François Atoll, to examine the patterns of residency and movement of manta rays throughout the archipelago; (2) assess the population composition (i.e. sex and life stage ratios) and habitat use patterns of manta rays at these aggregation areas and (3) use a novel remote underwater camera system and environmental data to investigate the influence of environmental and temporal factors on patterns on visits to a cleaning station located to the north of D’Arros Island. Finally, we aimed to examine the significance of the aggregation sites at D’Arros Island and St. François Atoll to these globally ‘Vulnerable’ species.
Methods
Study site
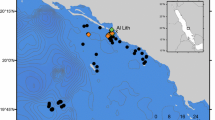
Located in the Western Indian Ocean, Seychelles is a remote archipelago comprised of 115 tropical islands (Fig. 1A). These islands cover a total land mass of approximately 452 km2, which encompasses just 0.03% of the Exclusive Economic Zone of the country. The islands of Seychelles form six island groups (Aldabra, Alphonse, Amirante, Farquhar, Inner and Southern Coral) based on their geography and geological composition. The populous Inner Island Group includes 41 granitic islands that are situated upon the Mahé Plateau. All remaining islands constitute the coralline Outer Islands and are further grouped based on their location and proximity to one another. Confirmed and incidental sightings of manta rays have been recorded throughout the archipelago; however, only two reliable aggregation areas have been identified to date; the first, at D’Arros Island (Amirate Group; Fig. 1B) and the second at St. François Atoll (Alphonse Group; Fig. 1D). These areas represent the focal survey areas within the wider Seychelles study area, with the cleaning station at D’Arros Island (Fig. 1C) selected for a targeted investigation into the patterns driving the repeated visitation to such aggregation sites by M. alfredi.

The six Island Groups of Seychelles (A; EEZ in the top left inset). D’Arros Island and St. Joseph Atoll (St. J; B) are located within the Amirante Group. Manta ray photo-identification surveys conducted at D’Arros Island (C) targeted nine sites during surface-based searches (white circles), while dive-based searches were conducted at the manta ray cleaning station to the north of the island (black diamond). Manta ray surveys were also conducted at St. François Atoll (D). ×, position of the Victoria Fish Market on Mahé Island. Drone images of D’Arros and St. J courtesy and copyright of Drone Adventures for the Save Our Seas Foundation
Data collection
Photographs of the unique pigmentation patterns on the ventral surface of manta rays were used to identify individuals throughout Seychelles (Marshall et al. 2011; Marshall and Pierce 2012). A sighting for an individual manta ray was defined as a confirmed photo-identification of that individual on a given day at a single location, and, as such, all sighting records included a clear identification photograph, location and GPS position (where possible), as well as date and time information. Species identification was based on the physical characteristics described by Marshall et al. (2009), typically by the distribution and density of pigmentation on the ventral surface, the colouration of the pectoral fin margin and the shape of the dorsal shoulder patches. Assignment of individuals to a colour morph type—common (i.e. chevron), black (i.e. melanistic) or white (i.e. leucistic)—also followed Marshall et al. (2009). Where possible, the presence (male) or absence (female) of claspers was used to determine the sex of individuals (Marshall and Bennett 2010). The disc width (DW; m) of each individual was estimated visually by experienced field staff to the nearest 0.1 m (m) and used as a proxy to assess the maturity status of M. alfredi based on the following metrics: juvenile (≤ 2.4 m), sub-adult (male, 2.5–2.8 m; female, 2.5–3.1 m) or adult (male, ≥ 2.9 m; female, ≥ 3.2 m) (Stevens 2016). Disc width estimates were initially made independently in-water by the field team before a final estimate was agreed upon during image processing. The presence of mating scars, fresh mating wounds and pregnancy bulges on females, and the degree of calcification of the claspers of males, were also considered during the assignment of maturity status, as these characteristics are indicative of sexually active individuals (Marshall and Bennett 2010; Stevens et al. 2018). In addition, the size of pregnancy bulges in females was used to estimate stage of gestation (Marshall and Bennett 2010). Physical injuries were also noted during sightings, but were not used as a primary characteristic to identify individuals given the rapid healing rates of manta rays (McGregor et al. 2019). Likely sources of injuries were categorised as natural (e.g. shark bite to a pectoral fin) or anthropogenic (e.g. fishing line scarring around a cephalic fin; Fig. 2), where possible.

Injury types recorded on reef manta rays (Mobula alfredi) in Seychelles. C: cephalic fin injury, D dorsal fin injury, G gill infection, P pectoral fin injury, T tail injury
The behaviour of individuals during sighting events was noted wherever possible. Four primary behaviours were recognised: feeding, courtship, cleaning and cruising. Feeding occurred when individual manta rays had unfurled cephalic fins, while swimming with the mouth open. When feeding, individuals often formed ‘chains’, where individuals swam in a single file one behind another. They also occasionally performed backward somersaults through the water column when feeding (Stevens et al. 2019), and this behaviour was noted when observed. The movements of individuals involved in courtship were notably faster than those involved in feeding. Courtship was characterised by one or more males actively pursuing a single female, while not engaged in feeding, cleaning or cruising behaviours (Stevens et al. 2018). Cleaning behaviour occurred when manta rays reduced their speed to linger over sections of reef, where cleaner fishes removed external parasites from their skin. Individuals often unfurled their cephalic fins during this process. Behaviour was classified as cruising when manta rays swam through the water column with their cephalic fins furled and their mouths closed. Where multiple behaviours occurred during a sighting, the behaviour that was observed to occupy the majority of the encounter was reported.
Survey frequency
Identification photographs of manta rays were collected across three spatio-temporal scales in Seychelles. First, sighting records of manta rays were collected opportunistically by the authors, collaborators and citizens throughout the archipelago between 2006 and 2018 to identify key aggregation areas for these species and to assess the range of individual movements among islands and reefs. Opportunistic records were sourced either from archival photographs or newly captured images submitted to the authors by citizens aware of the Seychelles Manta Ray Project (https://www.mantatrust.org/seychelles).
Second, intensive surveying was conducted at the two identified aggregation areas for reef manta rays. Three intensive surveying periods were conducted at D’Arros Island by the authors from the Save Our Seas Foundation—D’Arros Research Centre and the Manta Trust during November 2013, 2016 and 2017 (Table 1). Surveys were conducted at all times during the day (range 07:13–18:00) by between two and five people, with surface-based searches averaging 85 min (range 15–260 min), and manta dives averaging 56 min (range 16–162 min). Collected photo-identification data were used to examine the number of unique individuals visiting the area and their level of residency at this location. Observations of manta behaviour were also recorded at ten sites around D’Arros Island (Fig. 1C) during surface-based and dive-based searches to examine patterns of habitat use. A week-long pilot study was also completed by the authors from the Island Conservation Society and the Manta Trust at St. François Atoll (2–9 December 2017; Fig. 1D; Table 1) to investigate the occurrence of manta rays at this location and to examine their use of the lagoon within the Atoll. These latter surveys were conducted by teams of two to five people throughout the day (range 07:05–16:45), with surface-based searches averaging 414 min (range 105–563 min) and manta dives averaging 38 min (range 20–58 min).
Finally, a remote underwater camera system (hereafter, MantaCam; described below) was used to continuously monitor manta ray visits to a cleaning station at D’Arros Island over a 2-month period (29 September to 27 November 2017). Data collected by MantaCam were used to identify individuals visiting the cleaning station, monitor the frequency of manta visits and examine the behaviour of individuals at this site.
The total combined count of identified individuals across the three spatio-temporal scales of this study was used to provide a minimum estimate of population size for manta rays in Seychelles.
MantaCam
MantaCam was used to continuously monitor manta ray activity at a cleaning station located to the north of D’Arros Island (Figs. 1, 3) and to collect identification photographs at this site in the absence of human presence (i.e. divers). The camera system consisted of a single GoPro Hero4 (GoPro, California, US) with a Blink time-lapse controller, which was attached by a USB splitter cable to two Voltaic LiPo 44 Wh batteries with an ‘always-on’ feature. The camera and batteries were housed within a 17 × 29-cm PVC cylinder, which was sealed with a 2.3-cm-thick acrylic lid with an O-ring and metal latches. The time lapse for the GoPro was set to take an image every 10 s, allowing for deployment periods of approximately 48 h prior to battery depletion. Constant monitoring of the cleaning station was made possible using two MantaCam systems, whereby one camera was deployed as the other was retrieved. This cycle was repeated during October and November 2017.

MantaCam (A; black arrows) was deployed on the northern edge of the manta ray cleaning station (B; white arrow) at D’Arros Island, Seychelles, to remotely monitor manta ray visits to this location and to capture identification photographs. N north, S south
MantaCam was deployed on the northern edge of the cleaning station facing southwards (Fig. 3). The camera was positioned at an upward angle from the sea floor (~ 35°) to capture an image of the water directly above the cleaning station and towards the surface of the water column. All images captured during daylight hours were analysed, and the number of photographs containing any part of a manta ray was counted and used to record visits. The frequency and duration of manta ray visits to the cleaning station were measured using the timestamps on collected photographs. A visit was deemed to begin with the appearance of any part of a manta ray in a photograph, and end 10 min after the last photograph of any part of a manta ray was captured and all individuals were assumed to have left the vicinity of the station. When MantaCam was able to capture an image of the ventral surface of a manta ray, the individual was identified, and the sighting recorded alongside those from the other surveys of this study.
A current meter and a CTD probe (Valeport; Devon, United Kingdom) were deployed approximately 15 m to the north of the cleaning station at D’Arros Island and within 10 m of MantaCam at a depth of approximately 20 m to examine possible environmental drivers of manta ray visits to this site. Current speed (m s−1) and temperature (°C) records were collected every 15 min while the logger was deployed (07–12 October, and 02–28 November 2017) and data averaged over each hour of MantaCam deployment. Tidal data were also collected at D’Arros Island and used to calculate tidal range (m) and time relative to high tide (hours; h) at the cleaning station. Measures of tidal height were calculated using a tidal model built with the Oregon State University Tidal Model Driver (Egbert and Erofeeva 2002; J. Lea, pers. comm.). The tidal model was based on the tidal harmonics recorded at St. Joseph Atoll, and ground-truthed using in situ tidal measurements using a U20-001 Water Level Data Logger (Hobo, Bourne, MA). The model predicted tidal height in metres every 10 min, and these data were also averaged over each hour that MantaCam was deployed. All aforementioned environmental and temporal variables have been shown to influence manta ray visitation rates and/or movement patterns in previous studies (Jaine et al. 2012; Rohner et al. 2013; Couturier et al. 2018) and were included here to facilitate inter-population comparisons.
Statistical analyses
Sighting frequency
Photo-identification and sighting data were managed using an online database, through which summary statistics relating to individual sighting counts were derived. All other analyses used R (version 3.4.1; R Core Team 2017). Chi-squared tests were used to determine whether the proportion of males and females, and/or the three maturity classes, differed significantly across the three spatio-temporal scales considered in this study.
Visits to the cleaning station
A generalised additive mixed model (GAMM) with a binomial error structure was constructed using the package mgcv (Wood and Wood 2015) to examine the influence of current speed, water temperature, hour of day, tidal range and time to high tide on the occurrence of manta ray visits to the cleaning station (Table 2). Manta ray visits were analysed on an hourly basis across MantaCam deployments over 26 survey days (02–27 November 2017). Each hour per day (05:00–18:00) was assigned either a ‘1’ for a visit occurring or a ‘0’ for no recorded visit, generating a binomial dataset that was used as the response variable within the GAMM. Given the restriction of MantaCam data collection to daylight hours and the relatively short (< 1 month) monitoring period of this study, a tensor product smooth was fitted for hour of day and survey day number (numbered from 1 to 26) to consider potential interactions occurring between the two. Hourly environmental data were then aligned with hourly visit data, where possible, and fitted into GAMMs using thin plate splines (k = 5). This produced a dataset of 277 complete observations of the full suite of variables that were included in the final GAMM. No strong correlation was found to be present between any tested predictors (r < 0.8; Sleeman et al. 2010), and an AR(1) correlation structure was used to account for temporal autocorrelation occurring in the data (Zuur et al. 2009).
All potential combinations of predictor variables were considered during the model selection process. Akaike’s information criterion corrected for sample size (AICc) and AICc weight (wAICc) were used to select the highest-ranking model. The values of wAICc were able to vary from 0 (no support) to 1 (complete support) for each model (Ferreira et al. 2017). Models within 2 AICc units of each other were considered to be equally ranked. When the model with the highest support was equally ranked with other candidate models, the model containing the lowest number of explanatory variables (i.e. the most parsimonious) was selected to be most appropriate for the data. The percentage of deviance explained by models (%DE) was used as a measure of goodness of fit. The function ‘vis.gam’ from the package mgcv was used to visualise the summed effects of the tensor product smooth included in the final GAMM. Contour values within this plot indicated the predicted magnitude of the effect of the combined variables on manta ray visits to the cleaning station. Summed effect plots were then compared to cumulative histograms of manta ray visits to the cleaning station throughout the full monitoring period (29 September to 27 November 2017) for further examination of visitation patterns.
Results
Overview
A total of 1582 sightings of 241 individual manta rays were reported throughout Seychelles between July 2006 and December 2018 across the three spatio-temporal scales of this study. Of these 241, only five individuals were M. birostris (Fig. S1) and the remaining 236 were M. alfredi (Table 3; Fig. 4). No melanistic or leucistic individuals of either species were sighted. Given the disproportionate amount of survey effort at D’Arros Island (total = 193.5 h) relative to the remainder of Seychelles (St. François Atoll = 53.5 h; rest of Republic = opportunistic sightings only), sightings are hereafter discussed separately at the ‘Alphonse and other Island Group’ and ‘D’Arros Island’ scales.

Discovery curve of reef manta ray (M. alfredi) sightings across all of Seychelles (green circles), at D’Arros Island and St. Joseph Atoll (Amirante Group; blue diamonds) and at St. François Atoll (Alphonse Group; orange triangles). Light blue shading indicates timing of three November surveys at D’Arros Island. Orange shading indicates timing of the single survey at St. François Atoll (survey details in Table 1) (colour figure online)
Alphonse and other Island Groups
Oceanic manta rays
Only five sightings of M. birostris were recorded throughout the study period (0.3% of total). All sightings of M. birostris were reported in the Inner Island Group of Seychelles, and none of the five identified individuals (three female, one male and one unknown sex) that were cruising at the time of each encounter were resighted (Fig. 5).

Summary of the distribution of oceanic (Mobula birostris; white values) and reef (M. alfredi; black values) manta ray sightings throughout the Island Groups of Seychelles. For M. birostris, value indicates both number of individuals and number of sightings recorded during this study at the two Inner Island locations. For M. alfredi, bold value indicates number of individuals sighted, and value in parentheses indicates the number of sightings recorded during this study. ‘⋆’ symbol is the position of the main study site; D’Arros Island and St. Joseph Atoll. ‘▼’ symbol indicates the position of St. François Atoll (St. F)
Reef manta rays
Sighting summary
Excluding data from D’Arros Island, a total of 160 sightings of 98 individual M. alfredi were recorded across Seychelles, with the Southern Coral Group being the only Island Group where no sightings were reported (Fig. 5). Sightings of M. alfredi within the Aldabra and Farquhar Groups comprised just 1.7% and 2.1% of the identified population, respectively, and none of the individuals (n = 4 and 5, respectively) in either region were resighted. Of the 11 (4.7%) individuals sighted in the Inner Island Group, only one was resighted and this occurred within 24 h of the first encounter. Excluding those manta rays recorded at D’Arros Island, an additional 22 sightings of M. alfredi were reported in the Amirante Group at Desroches and Poivre Islands. The remaining sighting records for 53 (22.5%) individuals were collected at St. François Atoll (Alphonse Group), of which 33 (62.3%) were resighted. Collectively, the 236 individual M. alfredi identified throughout Seychelles serve as a minimum estimate for the total population size of this archipelago as of December 2018.
The largest aggregation of M. alfredi occurred at St. François Atoll (Alphonse Group) on 19 December 2018, where approximately 40 individuals were observed feeding at the surface. The second largest event occurred at D’Arros Island (Amirante Group) on 4 November 2017 when a group of 25 individuals were observed. The pilot study conducted within the Alphonse Group in December 2017 over 8 field days contributed 43 confirmed sightings of 29 individual M. alfredi to the Seychelles records. Eighteen of these individuals were new to the database at the time of the encounters (Fig. 4). In addition, three incidental sightings of M. alfredi within the St. François Atoll lagoon system were noted. Poor water visibility (< 0.5 m) prohibited the collection of in-water identification images from this location, where observations of up to 15 large M. alfredi (DW > 2.5 m) swimming in the main lagoon channel and a small individual (DW ~ 1 m) feeding within the northernmost lagoon were reported from the field team onboard the survey vessel. The discovery curves for the Alphonse Group, D’Arros Island and Seychelles as a whole, have not yet reached an asymptote.
Sex and maturity
Male and female M. alfredi sighted at locations other than D’Arros Island were encountered at similar frequencies (n = 73 and 79 sightings, respectively; χ2 = 0.24, df = 1, P = 0.63). They also remained almost equally represented within the identified population (n = 43 and 48 individuals, respectively: χ2 = 0.27, df = 1, P = 0.60). Seven individuals (3.8%) were unable to be assigned a sex based on the available photographs (Fig. S2). Mature (n = 36, 36.8%) and sub-adult (n = 40, 40.8%) individuals continued to account for most of the identified M. alfredi population, with a significantly lower proportion of juveniles (n = 16, 16.3%) reported (χ2 = 10.78, df = 2, P = 0.005). The same was true when males (n = 43), whose maturity status is estimated based on the length of the claspers, were considered alone, with juvenile males (n = 5; 11.6%) accounting for less of the identified M. alfredi population (χ2 = 11.35, df = 2, P = 0.003) than sub-adult (n = 15; 34.9%) or adult (n = 23; 53.5%) males. Maturity status could not be assigned to six individuals (6.1%).
Sighting frequencies for all individuals throughout Seychelles varied from a single sighting to a maximum of 61 re-sightings of an adult male between July 2012 and November 2018 at D’Arros Island (Fig. S3; Table S1). The most frequently sighted female was a sub-adult sighted 47 times between December 2015 and November 2017 at D’Arros Island. Of the top 10 most re-sighted individuals in this study, seven were males (Fig. S3; Table S1). The longest time between two sightings of the same individual was 2808 days (7.75 years), where a juvenile male was first sighted at D’Arros Island on 9 August 2010 and next resighted on 17 April 2018 at Poivre Island of the Amirante Group. An updated maturity status was not determined during the latter sighting.
Behaviour
For M. alfredi sightings recorded away from D’Arros Island (n = 160), surface-feeding behaviour was reported most frequently (n = 135, 84.4%). Only five observations of somersault feeding were recorded, all of which occurred at St. François Atoll (Alphonse Group; 3.1%), including within the large feeding aggregation of 40 individuals sighted in December 2018. In addition, up to 15 individual M. alfredi were observed to surface-feed within the narrow (< 100 m wide), main channel of the St. François Lagoon on multiple occasions. On both rising and falling tides, individuals were noted to orient themselves against the prevailing current direction and maintained their position in the fast-flowing water. Cruising behaviour was observed during 17 of the 160 sightings (10.6%), and cleaning behaviour was reported eight times (5.0%). As of December 2018, no records of courtship behaviour (i.e. males pursuing females) currently exist for M. alfredi outside of the Amirante Group (described below).
Movements between Island Groups
Three individuals were recorded to move from the Amirante Group to the Alphonse Group. All three of these individuals travelled a minimum of 198 ± 2 km from D’Arros Island to St. François Atoll (straight line distance between sites), over time frames of 14–1624 days (Fig. 5). Return journeys were not observed in any of these cases, and no additional movements of individuals between other Island Groups were recorded in this study.
Injuries and anthropogenic impacts
Thirty-eight (37.8%) of the 98 individual M. alfredi sighted elsewhere than D’Arros Island displayed an injury to their body. Males (n = 20, 47%) and females (n = 16, 33.3%) displayed similar extents of injury. More adults (17 of 36 individuals, 47.2%) and sub-adults (15 of 40 individuals, 16.7%) displayed injuries than those of juveniles (10 of 35 individuals, 28.6%) (χ2 = 8.17, df = 1, P = 0.017 and χ2 = 8.05, df = 1, P = 0.005, respectively). There was no significant difference in injury rates between sub-adults and adults (χ2 = 0.125, df = 1, P = 0.724). The occurrence and cause (i.e. natural or anthropogenic) of injuries reported for these M. alfredi were similar to those reported for individuals at D’Arros Island (described below; Table S2). Injuries to the pectoral fins were most common (present on 32.7% of individuals sighted away from D’Arros), with the majority (75.7% of affected individuals) being natural in cause. Anthropogenic impacts were most frequently associated with cephalic fin injuries (5.4% of injured individuals).
Four deceased M. alfredi were reported in the Victoria Fish Market on Mahé Island (Inner Island Group) by collaborators during this study (Fig. 6). Landings occurred in 2015 (n = 2), 2016 (n = 1) and 2018 (n = 1) and included either finned individuals (n = 2) or pieces of pectoral fin (n = 2). In addition, two lethal entanglements of M. alfredi were reported from the Inner Islands: one involving a discarded fishing line (March 2009), and the other, a mooring line (December 2017).

© John Nevill) and lethal entanglements (bottom; © Marco Christensen) of reef manta rays (M. alfredi) have been reported in the Inner Island Group of Seychelles
Landings (top;
D’Arros Island
Population size and composition
Collective survey efforts at D’Arros Island identified a total of 157 individual M. alfredi. Of these individuals, 150 were first sighted at this location (63.6% of Seychelles total) and 122 (78.0% of D’Arros total) have been resighted on at least one occasion.
Male M. alfredi were sighted more frequently than females across surveys conducted at D’Arros Island (n = 824 and 591, respectively; χ2 = 38.37, df = 1, P < 0.001). Male (n = 79, 50.3%) and female (n = 76, 48.4%) individuals were almost equally represented within the population (χ2, = 0.06, df = 1, P = 0.81), and only two individuals (1.3%) were unable to be assigned a sex (Fig. S2). Mature (n = 75, 47.8%) and sub-adult (n = 62, 39.5%) individuals represented the majority of the M. alfredi sightings recorded at D’Arros Island, with a significantly lower number of juveniles (n = 20, 12.7%) identified by all survey methods at this location (χ2 = 31.58, df = 2, P < 0.001). The same was true when males (n = 79) were considered alone, with juvenile males (n = 8; 10.1%) accounting for less of the identified M. alfredi population (χ2 = 22.35, df = 2, P < 0.001) than sub-adult (n = 29; 36.7%) or adult (n = 42; 53.2%) males. All individuals at D’Arros Island were assigned a maturity status.
The relative abundance of different maturity classes (juvenile, sub-adult and adult) did not differ by sex at D’Arros Island (χ2, P > 0.05). In addition, there was no significant difference in resighting rates between identified males (n = 66, 83.5%) or females (n = 76, 73.7%; χ2 = 0.04, df = 1, P > 0.43), or among juveniles (n = 12, 66.7%), sub-adults (n = 47, 75.8%) and adults (n = 63, 81.8%; χ2 = 1.55, df = 2, P > 0.46).
Size distribution
Almost half (44.6%) of the M. alfredi identified at D’Arros Island had disc widths of 2.5–3.0 m, with an average disc width of 2.9 ± 0.5 m (Fig. S4). Six individuals had disc widths < 2 m, the majority of them being males (n = 4, 66.7%).The smallest individual, whose sex could not be determined, was estimated to have a disc width of 1.5 m. Females had a mean disc width significantly larger than that of males (mean = 3.06 and 2.82 m, respectively; t = − 3.52, df = 121.71, P < 0.001). A mature female was the only individual to exceed a disc width of 4.0 m.
Patterns of habitat use
Surface-feeding accounted for 53.6% of the 1391 behavioural observations of M. alfredi at D’Arros Island and was recorded at all sites (Fig. 7). Feeding behaviour was observed most frequently at sites towards the northeast of D’Arros Island, with surface-feeding observed during 96.7, 97 and 100% of sightings of M. alfredi at the northern, northeastern and eastern sites, respectively. Courtship behaviour was observed 22 times at the cleaning station to the north of D’Arros Island, accounting for 3.7% of the total behavioural observations recorded at this site. Cleaning behaviours were observed 545 times (39.7%) along the northern shoreline of D’Arros Island, most frequently at the cleaning station (n = 534). Cleaning behaviour was only reported once at Airstrip North and eight times at Jetty (Fig. 7). Cruising behaviour constituted 5.1% (n = 70) of all behavioural observations at D’Arros Island and was most frequently observed along the western and southern coastlines (range 2.5–16.7% of records across sites).

Proportion of occurrence and distribution of feeding, courtship, cleaning and cruising behaviour by reef manta rays (M. alfredi) at 10 sites around D’Arros Island, Seychelles. Values within pie charts indicate the total number of behavioural records collected at each site. Eastern survey sites located within the channel between D’Arros Island and St. Joseph Atoll (St. J). Colour allocations to behaviours are described in figure legend. Drone images of D’Arros and St. J courtesy and copyright of Drone Adventures for the Save Our Seas Foundation
Injuries and anthropogenic impacts
Almost half (n = 76; 48.4%) of the individual M. alfredi sighted at D’Arros Island had an injury to their body. Males (n = 35, 44.0%) and females (n = 40, 52.6%) displayed similar extents of injury. Adults (41 of 75 individuals, 54.7%) were more frequently injured than juveniles (6 of 20 individuals, 30.0%; χ2 = 7.15, df = 1, P = 0.01), but there was no significant difference in injury rates between sub-adults (29 of 62 individuals, 46.8%) and either juveniles or adults (χ2 = 3.68, df = 1, P = 0.06 and χ2 = 0.60, df = 1, P = 0.44, respectively).
Injuries to pectoral fins were most common, followed by injuries to the tail and/or dorsal fin, gills and cephalic fins (43.3%, 13.4%, 5.1% and 3.8% of those injured, respectively; Table 4). Natural causes were responsible for most injuries to these individuals (76.3%), with anthropogenic threats (namely, damage caused by fishing line) accounting for 3.9% of injuries. Injuries from a combination of natural and anthropogenic causes were observed in four individuals (5.3%), whereas the likely causes of injury were unknown in 11 individuals (14.5%). Anthropogenic causes were most frequently associated with injuries to cephalic fins.
Cleaning station
Sighting summary
MantaCam captured 203,277 photographs during the 599.5 h that it was deployed and active at the D’Arros cleaning station, of which 3146 (1.5%) contained images of M. alfredi. These latter images contributed 341 confirmed sightings of 83 individuals to the Seychelles database (21.6% of total), of which seven were new. No M. birostris were recorded at the cleaning station by MantaCam. The largest number of photographs containing M. alfredi collected during a single day was 174 on 10 November 2017. The maximum number of individuals estimated to be photographed by MantaCam in a single day was 18 (27 November 2017). Of these, 15 were individually identified and three were unidentifiable.
Males (n = 217, 63.6%) were sighted more frequently at the cleaning station than females (n = 124, 36.4%; χ2 = 25.4, df = 1, P < 0.001). In addition, the cleaning station was more frequently visited by adult and sub-adult individuals, than by juveniles (53.1%, 38.7% and 8.2%, respectively; χ2 = 31.5, df = 2, P < 0.001).
Visits of M. alfredi to the cleaning station lasted an average of 4.92 ± 6.84 (S.D.) min, although visits lasting 1 min occurred most frequently. MantaCam recorded a total of 392 visits by M. alfredi to the cleaning station during the full deployment period (Fig. S5). The largest number of individuals recorded at the cleaning station at the same time was seven (26 November 2017), and the highest number of visits that were recorded in a single day was 14 by 13 individuals (17 November 2017).
Behaviour at the cleaning station
Cleaning dominated behavioural records at the station (90.7% of observations), although feeding, courtship and cruising were also reported (3.1%, 3.7% and 2.5%, respectively). Courtship had not been observed at this site prior to the deployment of MantaCam in 2017. Images collected by MantaCam were used to contribute 12 additional courtship events to the existing database of records at D’Arros Island (Fig. S6), where only four observations of courtship had been recorded during previous years. In some instances, identification images were captured during courtship events (Fig. S6A), allowing identification of participants. The largest number of individuals observed engaging with each other in a single courtship event was four (Fig. S6B), although events involving only two (n = 9) or three (n = 2) individuals were more common.
MantaCam captured images of one pregnant female (DW = 3.6 m) on 12 October 2017 (Fig. S6C). It is estimated that this individual was in the fourth trimester of a 12-month pregnancy based on the extent of the ventral stomach bulge (Stevens 2016). In total, four pregnant females were identified throughout Seychelles during this study, and an additional 16 individuals were seen with mating scars on the tip of their left pectoral fin. These individuals accounted for 17.4% of the population of female M. alfredi identified in Seychelles as of December 2018.
Patterns of visits
The top-ranked GAMM explained 14.8% of the variation in the visitation data and included only the tensor product spline of time of day and day of survey as a driver of manta visits to the cleaning station (Table 5). Current speed, water temperature, tidal range and time to high tide did not have a significant effect on the frequency of visits. Visits by M. alfredi to the cleaning station varied significantly with the hour of the day and were more likely to occur during the middle of the day (10:00–15:00) than in the hours following dawn or preceding dusk (Fig. 8A). This pattern of visitation was most consistent during the first 2 weeks of November 2017, with a greater amount of variation observed in the visit times recorded during the last 2 weeks of the month (Fig. 8A). The same pattern was also observed when M. alfredi visits were considered cumulatively per hour over all MantaCam deployments (29 September to 27 November 2017), where the majority of visits occurred between 10:00 and 14:00 (Fig. 7B).

Frequency of visits by reef manta rays (M. alfredi) to a cleaning station at D’Arros Island, Seychelles, recorded using a remote camera system (‘MantaCam’), relative to time of day (h) and survey day (2–27 November 2017; numbered 1–26; A). Contour lines and associated values indicate the magnitude of summed effects of both variables on the occurrence of manta rays at this site, ranging from low (red), to average (yellow), to high (white). Cumulative number of manta visits to the cleaning station across all MantaCam deployments are also presented (29 September (yellow) to 27 November 2017 (purple); B)
Discussion
Sightings of manta rays in Seychelles recorded using photo-identification techniques between 2006 and 2018 were dominated by a relatively small, semi-resident, population of 236 individual M. alfredi. Only five M. birostris were sighted, and always within the Inner Island group of Seychelles. The combination of systematic survey efforts and opportunistic capture of photo-identification photographs by citizen scientists used in this study allowed for the first description of the population sizes of manta rays in Seychelles, the connectivity of individuals between Island Groups of the archipelago and examination of residency and habitat use of M. alfredi at and around key aggregation areas. In addition, the use of a remote underwater monitoring system combined with environmental data allowed us to examine the drivers of visits by M. alfredi to a cleaning station at D’Arros Island. This study highlights the value in combining data captured opportunistically by citizen scientists with that of routine surveys—particularly across vast and remote study areas—and provides important information regarding the conservation needs of these globally threatened species in a remote region of the Western Indian Ocean.
Amirante, Alphonse and other Island Groups
The population of M. alfredi identified across the islands of Seychelles as of December 2018 is relatively small at 236 individuals, with the majority sighted within the Amirante and Alphonse Groups (n = 216, 91.5%), at D’Arros Island and St. François Atoll, respectively. Similar population sizes for M. alfredi have been observed in the archipelagic waters of Hawaiʻi and the Yaeyama Islands of Japan (290 and 300 individuals, respectively; Homma et al. 1999; Deakos et al. 2011). In contrast, a population of 4247 individuals has been identified in the Maldives, located 2200 km to the northeast of Seychelles (Stevens et al. 2018). To a large extent, these differences in population size likely reflect the variable spread of human populations (i.e. observers) and available monitoring resources within Seychelles and the Maldives. In the former, many Island Groups are remote and difficult to access (e.g. Aldabra), whereas in the latter, all atolls have human settlements. In addition, there has been a targeted manta research programme operating within the Maldives year-round for over a decade and the economy of the country is based, in large part, on marine ecotourism (Zimmerhackel et al. 2018, 2019), creating an incentive to locate aggregations of manta rays (Anderson et al. 2011; O’Malley et al. 2013; Ward-Paige et al. 2013; Harris et al. 2020). The lack of human access to other remote Island Groups in Seychelles, where manta rays are known to occur, and the connectivity documented between the D’Arros Island and St. François Atoll aggregation sites, mean that our study has recorded only a fraction of the population of M. alfredi likely to exist in the Seychelles archipelago as a whole. It is also likely that the aggregations in the Amirante and Alphonse Groups, where our sampling effort was focussed, are larger than estimated here, as discovery curves did not reach an asymptote (Couturier et al. 2011; Deakos et al. 2011).
The few sightings of M. birostris in comparison to M. alfredi is similar to other localities, including Hawaiʻi, French Polynesia, Australia and Fiji (Clark 2010; Carpentier et al. 2019; Armstrong et al. 2020; Gordon and Vierus 2022). This could be due to the preference of M. birostris to frequent oceanic environments and their transient use of tropical coral reefs throughout much of their range (Carpentier et al. 2019; Armstrong et al. 2020; Gordon and Vierus 2022). Conversely, these coral reef habitats are where M. alfredi tend to aggregate and display high levels of site fidelity (including in Seychelles); sites where increased numbers of human observers also tend to be present (Couturier et al. 2012; Stewart et al. 2016, 2018; Armstrong et al. 2019; Peel et al. 2019a). For the most part in Seychelles, sightings of M. birostris appear to be serendipitous events, where observers crossed paths with animals transiting coastal environments, as was the case with all M. birostris sightings reported here. Should aggregation areas for the species exist in Seychelles, they likely occur outside of the boundary of our study.
Individual M. alfredi moved occasionally between the Amirante and Alphonse Groups (~ 200 km) and not between any other Island Groups in Seychelles (distances ranging ~ 150–1100 km). Although our study was limited by the small size of the photo-identification database from Alphonse, this conclusion was supported by the results of passive acoustic (Peel et al. 2020) and satellite telemetry (Peel et al. 2019a) studies, which also indicate that this species displays restricted patterns of movement within Seychelles as a whole. Deep water (> 1000 m) occurs between the Island Groups of Seychelles and is thought to act as a barrier to the movements of M. alfredi (Deakos et al. 2011), despite individuals displaying the ability to undertake large-scale movements in locations including Japan, Indonesia and eastern Australia (> 300 km; Homma et al. 1999; Germanov and Marshall 2014; Jaine et al. 2014; Armstrong et al. 2019). It has been hypothesised that deep water may increase the threat of predation (Marshall and Bennett 2010; Deakos et al. 2011) and reduce the likelihood of encountering prey in offshore locations (Gove et al. 2016). It is also possible, however, that sufficient food availability and mating opportunities afforded to M. alfredi within these Island Groups reduce the need for animals to transit the relatively large distances between aggregation sites (Couturier et al. 2018), resulting in the site fidelity observed here. Equally, for the neighbouring Amirante and Alphonse Groups, the shallow waters of the Amirantes Bank (< 40 m) may temper the barrier imposed by the deep water associated with their final stage of transit to the Alphonse Group from D’Arros Island, reducing the distance spent in potentially riskier waters from 200 km to < 100 km and increasing resource availability for individuals visiting these two sites. Continued monitoring of M. alfredi aggregations across the archipelago via photo-identification, satellite telemetry and genetic analyses will be required to confirm the full extent of population connectivity exhibited among the Island Groups of Seychelles.
D’Arros Island
The resighting of 78% of individuals identified at D’Arros Island on at least one occasion and the maximum resighting frequency of 61 occasions reflect their fidelity to this aggregation site and demonstrate the continued significance of this location to M. alfredi in Seychelles. D’Arros Island may provide critical habitat to M. alfredi on the shallow Amirantes Bank (< 40 m; Stoddart et al. 1979) and as well as refuge for individuals across the extensive reef flats (Couturier et al. 2014; McCauley et al. 2014; Germanov et al. 2019; Peel et al. 2019a). In addition, the presence of the cleaning station may facilitate important social interactions among individuals at this location (Deakos et al. 2011; Stevens et al. 2018; Germanov et al. 2019). It is also possible that the unique bathymetry of the relatively deep St. Joseph Channel (60 m; Stoddart et al. 1979) between D’Arros Island and St. Joseph Atoll enhances zooplankton aggregation at this location, allowing for cost-efficient foraging (Anderson et al. 2011; Armstrong et al. 2016). Regardless of the mechanisms driving the observed residency patterns, the prolonged and frequent visits of M. alfredi to this location highlight its significance to this population.
Females and males were almost equally represented at D’Arros Island. Equal sex ratios have been reported for M. alfredi populations in Japan, Hawaiʻi and the Maldives (Homma et al. 1999; Deakos et al. 2011; Stevens et al. 2018), although higher proportions of males are observed in Indonesia (1.42:1; Germanov et al. 2019) and higher proportions of females are observed at Lady Elliot Island on the Great Barrier Reef (1.32:1; Couturier et al. 2011). The largest female bias has been reported in Mozambique (3.55:1; Marshall et al. 2011); however, it is likely that these differences in demographic ratios are being driven by varied survey methodologies and timing across manta aggregation areas. For example, in the Maldives, male and female M. alfredi were represented equally across the whole population; however, a female bias was reported when only sightings at cleaning stations were considered (Stevens 2016). During periods of courtship and mating, this shifted towards a male bias (Stevens 2016). In addition, while there was no sex bias in the number of male and female M. alfredi reported at Maldivian feeding areas overall, female individuals were sighted more frequently and tended to be bolder than males, increasing the likelihood of missed sighting opportunities for males during feeding aggregations given their lower likelihood of returning to the site (Stevens 2016). Collectively, these findings emphasise the importance of year-round, multi-site sampling for studies of M. alfredi population dynamics and highlight the value of remote systems such as MantaCam in capturing the sighting data required to fully resolve the composition and habitat use patterns of populations.
Adult and sub-adult M. alfredi comprised the majority of the population at D’Arros Island and St. Joseph Atoll. Juveniles were observed infrequently throughout the wider region of Seychelles, with the smallest individual (DW ~ 1.5 m) being sighted within the lagoon of the St. François Atoll. Although this is a single sighting, lagoon habitats are thought to provide refuges and food sources for young M. alfredi (McCauley et al. 2014; Andrzejaczek et al. 2020), so this region may function as a pupping area for the species in the Alphonse Group. In addition, the observation of large numbers of individual M. alfredi feeding within the main channel of the St François Atoll lagoon suggests that this locality may provide significant foraging opportunities to individuals in the Alphonse Group. Further sampling at St. François Atoll will be required to confirm or refute the hypothesis that this location serves as a nursery area for M. alfredi and to quantify the importance of the lagoon system to the foraging ecology of the species.
Cleaning station
At the D’Arros Island cleaning station specifically, individual M. alfredi were more likely to be sighted during the middle hours of the day than that at dawn or dusk. A similar visitation pattern has been reported for M. alfredi at cleaning stations at other locations around the world, including eastern Australia, Indonesia, Mozambique and the Maldives (Couturier et al. 2018; Setyawan et al. 2018; Venables et al. 2020; Harris and Stevens 2021). This may be due to the combined factors of cleaner fishes at cleaning stations only being active during daylight hours (Potts 1973) and M. alfredi having the ability to repeatedly, and significantly, associate their space use with the distribution of blue-streak cleaner wrasse (Labroides dimidiatus) and hard coral substrate (Armstrong et al. 2021). In addition, such patterns of visitation may be the result of the feeding behaviour of M. alfredi. Stable isotope analyses have revealed that emergent zooplankton comprises a large proportion of the diet of this species at D’Arros Island (38%; Peel et al. 2019b) and in eastern Australia (Couturier et al. 2013). The crepuscular movement of these benthic zooplankton communities may trigger foraging by M. alfredi at dawn and dusk (Armstrong et al. 2016) and subsequently reduce the frequency of visits by individuals to the cleaning station at D’Arros Island during these times. Studies using passive acoustic telemetry further support this hypothesis, with M. alfredi more likely to be detected during the day than at night at D’Arros Island, and elsewhere (Couturier et al. 2018; Setyawan et al. 2018; Peel et al. 2019a). These telemetry data also overcome the restriction of data collection by MantaCam to daylight hours, providing insight into the patterns of movement of M. alfredi throughout the full diel cycle.
Current speed, water temperature, tidal range and time to high tide did not influence the frequency of M. alfredi visits to the cleaning station at D’Arros Island. This was unexpected given the importance of environmental drivers at aggregation sites for M. alfredi on the Great Barrier Reef (O’Shea et al. 2010; Jaine et al. 2012), in the coastal waters of Mozambique (Rohner et al. 2013), in the Maldives (Harris and Stevens 2021) and at D’Arros Island as a whole (Peel et al. 2019a). In addition, in-water observations of M. alfredi at the D’Arros Island cleaning station suggested that current speed may influence how individuals visit this location, with sightings frequently being reported at the onset of, and during a slack tide under minimal current speeds (L. Peel, pers. comm.). It seems likely, however, that the relatively short (approximately 1 month) deployment period of MantaCam contributed to our result. The fortnightly variation observed in M. alfredi visits during this study suggests that tidal cycles may play a role in manta ray visitations to cleaning sites; however, these patterns were unable to be clearly resolved here. Furthermore, the ~ 85% of variance yet to be explained by our model suggests that other factors (e.g. prey distribution and availability) may also be playing a role in how M. alfredi visit and use cleaning stations. Longer MantaCam deployment times may be required to identify important environmental drivers for M. alfredi occurring at tidal and lunar scales (Couturier et al. 2018), to allow for the variation occurring over these timeframes to be better captured and understood.
Despite the short duration of sampling, the deployment of MantaCam at the cleaning station to the north of D’Arros Island quadrupled the number of courtship records for M. alfredi at this location. Previously, courtship behaviour had not been recorded at the D’Arros cleaning station, despite over 80 dive-based manta searches being completed at the site and courtship often being recorded at cleaning stations for this species elsewhere (e.g. the Maldives, Hawai’i; Deakos et al. 2011; Stevens et al. 2018). This finding suggests that the significance of D’Arros Island to the courtship and reproductive behaviours of M. alfredi were underestimated by previous survey efforts. It also highlights the value of using remote monitoring systems at key aggregation sites for M. alfredi to examine patterns of habitat use (O’Shea et al. 2010) and to increase the number of collected identification images. Despite only being deployed for 3 months, sightings from MantaCam accounted for 22% of all sightings in the database for Seychelles at the completion of this study (December 2018). The ability of this camera system to facilitate a rapid accumulation of data over a short time frame underscores its value for establishing baseline population estimates and documenting behaviours of manta rays in remote locations.
Manta ray conservation in Seychelles
The current lack of a targeted mobulid fishery in Seychelles, coupled with the remote nature of the archipelago and aggregation areas, serves to protect local manta ray populations from the pressures of increased human populations, development and marine resource overexploitation that have resulted in declining populations elsewhere (Lawson et al. 2017). It remains important to recognise, however, that instances of mobulid bycatch and entanglement do still occur, particularly around the populated Inner Islands where fishing pressures are greatest, and that anecdotal records indicate that manta rays were historically hunted for their meat in the country (Keynes 1959). In addition, marine debris, including fish aggregation devices and ghost nets, that pose an entanglement risk to manta rays may float into the Seychelles EEZ from elsewhere in the Indian Ocean, further complicating the management of these populations (Weideman et al. 2023). Despite the increased human population, number of water users (i.e. citizen scientists) and public awareness of manta rays in the news and media in Seychelles (L. Peel, pers. obs.), sightings of M. alfredi remain comparatively low in the Inner Islands compared to those recorded across the Outer Islands. Together with the entanglement and bycatch observations presented here, this suggests that the M. alfredi population may have been reduced within the Inner Islands, emphasising the importance of conserving the aggregation areas currently identified at D’Arros Island and St. François Atoll.
The isolated nature of the M. alfredi aggregations identified at the Amirante and Alphonse Groups provides an important opportunity to both conserve manta rays in Seychelles and gather important baseline information about the biology and ecology of these animals in the absence of common anthropogenic disturbances. The increasing impact of small fisheries in the Western Indian Ocean, however, raises concerns about increasing anthropogenic threats to M. alfredi aggregations at D’Arros Island and St. François Atoll (Temple et al. 2019). Should demand for mobulid products increase in the region, it is possible that these predictable aggregations could be readily and rapidly impacted by targeted fisheries (Temple et al. 2019). With a relatively small population in the tens of hundreds and for a species with such a conservative life history, even small increases in mortality from anthropogenic sources could quickly lead to regional extinction (Couturier et al. 2012; Ward-Paige et al. 2013; Rohner et al. 2017).
This concern, alongside the findings presented here for this small, semi-resident population, highlights the importance of the Marine Spatial Plan announced by the Government of Seychelles in 2020 (Government of Seychelles 2020), which includes the designation of a Zone 1 (i.e. High Biodiversity Protection) Marine Protected Area around D’Arros Island and St. Joseph Atoll and a Zone 2 (i.e. Medium Biodiversity Protection and Sustainable Use) Marine Protected Area around St. François Atoll and Alphonse Island. While the level of protection afforded to manta rays within these areas will depend on the regulations incorporated into their management, prohibiting their capture and the implementation of safe ecotourism interaction protocols (Venables et al. 2016; Murray et al. 2020) would provide a significant improvement in conserving a key manta aggregation area within the archipelago. Such preemptive conservation measures would serve not only to protect individuals of a globally vulnerable species from future threat but also to prolong our ability to study this population of M. alfredi, most notably within the other Outer Island Groups in Seychelles, where similar significant, and equally vulnerable, aggregations likely occur but have not yet been surveyed. A better understanding of critical habitat for M. alfredi in these areas could then be used to inform the establishment of management strategies for aggregations of M. alfredi occurring elsewhere in the Western Indian Ocean and beyond.
Data availability
The datasets generated and analysed during the current study are not publicly available because the manta sighting data are owned by a large number of contributors. They are available from the corresponding author upon reasonable request.
References
Anderson RC, Adam MS, Goes JI (2011) From monsoons to mantas: seasonal distribution of Manta alfredi in the Maldives. Fish Oceanogr 20:104–113. https://doi.org/10.1111/j.1365-2419.2011.00571.x
Andrzejaczek S, Chapple TK, Curnick DJ, Carlisle AB, Castleton M, Jacoby DMP, Peel LR, Schallert RJ, Tickler DM, Block BA (2020) Individual variation in residency and regional movements of reef manta rays Mobula alfredi in a large marine protected area. Mar Ecol Prog Ser 639:137–153. https://doi.org/10.3354/meps13270
Armstrong AO, Armstrong AJ, Jaine FRA, Couturier LIE, Fiora K, Uribe-Palomino J, Weeks SJ, Townsend KA, Bennett MB, Richardson AJ (2016) Prey density threshold and tidal influence on reef manta ray foraging at an aggregation site on the Great Barrier Reef. PLoS ONE 11:1–18. https://doi.org/10.1371/journal.pone.0153393
Armstrong AO, Armstrong AJ, Bennett MB, Richardson AJ, Townsend KA, Dudgeon CL (2019) Photographic identification and citizen science combine to reveal long distance movements of individual reef manta rays Mobula alfredi along Australia’s east coast. Mar Biodivers Rec 12:10–15. https://doi.org/10.1186/s41200-019-0173-6
Armstrong AJ, Armstrong AO, Bennett MB, McGregor F, Abrantes KG, Barnett A, Richardson AJ, Townsend KA, Dudgeon CL (2020) The geographic distribution of reef and oceanic manta rays (Mobula alfredi and Mobula birostris) in Australian coastal waters. J Fish Biol 96:835–840. https://doi.org/10.1111/jfb.14256
Armstrong AO, Armstrong AJ, Bennett MB, Richardson AJ, Townsend KA, Everett JD, Hays GC, Pederson H, Dudgeon CL (2021) Mutualism promotes site selection in a large marine planktivore. Ecol Evol 11:5606–5623
Carpentier AS, Berthe C, Ender I, Jaine FRA, Mourier J, Stevens G, De Rosemont M, Clua E (2019) Preliminary insights into the population characteristics and distribution of reef (Mobula alfredi) and oceanic (M. birostris) manta rays in French Polynesia. Coral Reefs 38:1197–1210
Clark TB (2010) Abundance, home range, and movement patterns of manta rays (Manta Alfredi, M. birostris) in Hawai’i. p 149
Couturier LIE, Jaine FRA, Townsend KA, Weeks SJ, Richardson AJ, Bennett MB (2011) Distribution, site affinity and regional movements of the manta ray, Manta alfredi (Krefft, 1868), along the east coast of Australia. Mar Freshw Res 62:628–637
Couturier LIE, Marshall AD, Jaine FRA, Kashiwagi T, Pierce SJ, Townsend KA, Weeks SJ, Bennett MB, Richardson AJ (2012) Biology, ecology and conservation of the Mobulidae. J Fish Biol 80:1075–1119. https://doi.org/10.1111/j.1095-8649.2012.03264.x
Couturier LIE, Rohner CA, Richardson AJ, Marshall AD, Jaine FRA, Bennett MB, Townsend KA, Weeks SJ, Nichols PD (2013) Stable isotope and signature fatty acid analyses suggest reef manta rays feed on demersal zooplankton. PLoS ONE 8:e77152
Couturier LIE, Dudgeon CL, Pollock KH, Jaine FRA, Bennett MB, Townsend KA, Weeks SJ, Richardson AJ (2014) Population dynamics of the reef manta ray Manta alfredi in eastern Australia. Coral Reefs 33:329–342. https://doi.org/10.1007/s00338-014-1126-5
Couturier LIE, Newman P, Jaine FRA, Bennett MB, Venables WN, Cagua EF, Townsend KA, Weeks SJ, Richardson AJ (2018) Variation in occupancy and habitat use of Mobula alfredi at a major aggregation site. Mar Ecol Prog Ser 599:125–145. https://doi.org/10.3354/meps12610
Croll DA, Dewar H, Dulvy NK, Fernando D, Francis MP, Galván-Magaña F, Hall M, Heinrichs S, Marshall A, Mccauley D (2016) Vulnerabilities and fisheries impacts: the uncertain future of manta and devil rays. Aquat Conserv Mar Freshw Ecosyst 26:562–575
Deakos MH, Baker JD, Bejder L (2011) Characteristics of a manta ray Manta alfredi population off Maui, Hawaii, and implications for management. Mar Ecol Prog Ser 429:245–260. https://doi.org/10.3354/meps09085
Egbert GD, Erofeeva SY (2002) Efficient inverse modeling of barotropic ocean tides. J Atmos Ocean Technol 19:183–204
Ferreira LC, Thums M, Heithaus MR, Barnett A, Abrantes KG, Holmes BJ, Zamora LM, Frisch AJ, Pepperell JG, Burkholder D (2017) The trophic role of a large marine predator, the tiger shark Galeocerdo cuvier. Sci Rep 7:1–14
Fonseca-Ponce IA, Zavala-Jiménez AA, Aburto-Oropeza O, Maldonado-Gasca A, Galván-Magaña F, González-Armas R, Stewart JD (2022) Physical and environmental drivers of oceanic manta ray Mobula birostris sightings at an aggregation site in Bahía de Banderas, Mexico. Mar Ecol Prog Ser 694:133–148
Germanov ES, Marshall AD (2014) Running the Gauntlet: regional movement patterns of Manta alfredi through a complex of parks and fisheries. PLoS ONE 9:1–9. https://doi.org/10.1371/journal.pone.0110071
Germanov ES, Bejder L, Chabanne DBH, Dharmadi D, Hendrawan IG, Marshall AD, Pierce SJ, van Keulen M, Loneragan NR (2019) Contrasting habitat use and population dynamics of reef manta rays within the Nusa Penida marine protected area, Indonesia. Front Mar Sci 6:215
Gordon L, Vierus T (2022) First photographic evidence of oceanic manta rays (Mobula birostris) at two locations in the Fiji islands. PeerJ 10:e13883
Gove JM, McManus MA, Neuheimer AB, Polovina JJ, Drazen JC, Smith CR, Merrifield MA, Friedlander AM, Ehses JS, Young CW (2016) Near-island biological hotspots in barren ocean basins. Nat Commun 7:1–8
Government of Seychelles (2020) Seychelles marine spatial plan policy. Ministry of Environment Energy and Climate Change, Seychelles. p 24
Harris JL, Stevens GMW (2021) Environmental drivers of reef manta ray (Mobula alfredi) visitation patterns to key aggregation habitats in the Maldives. PLoS ONE 16:e0252470
Harris JL, McGregor PK, Oates Y, Stevens GMW (2020) Gone with the wind: seasonal distribution and habitat use by the reef manta ray (Mobula alfredi) in the Maldives, implications for conservation. Aquat Conserv Mar Freshw Ecosyst 30:1649–1664
Homma K, Maruyama T, Itoh T, Ishihara H, Uchida S (1999) Biology of the manta ray, Manta birostris Walbaum, in the Indo-Pacific. In: Seret B, Sire J (eds) In Indo-Pacific fish biology: Proceedings of the 5th Indo-Pacific Fisheries Conference, Noumea, 1997. Ichthyological Society of France, Paris, pp 209–216
Hosegood J, Humble E, Ogden R, de Bruyn M, Creer S, Stevens GMW, Abudaya M, Bassos-Hull K, Bonfil R, Fernando D, Foote AD, Hipperson H, Jabado RW, Kaden J, Moazzam M, Peel LR, Pollett S, Ponzo A, Poortvliet M, Salah J, Senn H, Stewart JD, Wintner S, Carvalho G (2020) Phylogenomics and species delimitation for effective conservation of manta and devil rays. Mol Ecol 29:4783–4796. https://doi.org/10.1111/mec.15683
Jaine FRA, Couturier LIE, Weeks SJ, Townsend KA, Bennett MB, Fiora K, Richardson AJ (2012) When giants turn up: sighting trends, environmental influences and habitat use of the manta ray Manta alfredi at a coral reef. PLoS ONE 7(10):e46170
Jaine FRA, Rohner CA, Weeks SJ, Couturier LIE, Bennett MB, Townsend KA, Richardson AJ (2014) Movements and habitat use of reef manta rays off eastern Australia: offshore excursions, deep diving and eddy affinity revealed by satellite telemetry. Mar Ecol Prog Ser 510:73–86. https://doi.org/10.3354/meps10910
Kashiwagi T, Marshall AD, Bennett MB, Ovenden JR (2011) Habitat segregation and mosaic sympatry of the two species of manta ray in the Indian and Pacific Oceans: Manta alfredi and M. birostris. Mar Biodivers Rec 4:e53
Keynes Q (1959) Seychelles, tropic Isles of Eden. Natl Geogr Mag 116:670–695
Lawson JM, Fordham SV, O’Malley MP, Davidson LNK, Walls RHL, Heupel MR, Stevens G, Fernando D, Budziak A, Simpfendorfer CA (2017) Sympathy for the devil: a conservation strategy for devil and manta rays. PeerJ 5:e3027
Marie J (2022) World’s oldest known manta ray celebrates more than 40 years on Great Barrier Reef. ABC News
Marshall AD, Bennett MB (2010) The frequency and effect of shark-inflicted bite injuries to the reef manta ray Manta alfredi. Afr J Mar Sci 32:573–580
Marshall AD, Pierce SJ (2012) The use and abuse of photographic identification in sharks and rays. J Fish Biol 80:1361–1379
Marshall AD, Compagno LJV, Bennett MB (2009) Redescription of the genus Manta with resurrection of Manta alfredi (Krefft, 1868) (Chondrichthyes; Myliobatoidei; Mobulidae). Zootaxa 2301(1):1–28. https://doi.org/10.11646/zootaxa.2301.1.1
Marshall AD, Dudgeon CL, Bennett MB (2011) Size and structure of a photographically identified population of manta rays Manta alfredi in southern Mozambique. Mar Biol 158:1111–1124. https://doi.org/10.1007/s00227-011-1634-6
Marshall AD, Kashiwagi T, Bennett MB, Deakos M, Stevens G, McGregor F, Clark T, Ishihara H, Sato K (2018) Mobula alfredi (amended version of 2011 assessment). The IUCN red list of threatened species 2018: e. T195459A126665723
Marshall A, Barreto R, Carlson J, Fernando D, Fordham S, Francis MP, Derrick D, Herman K, Jabado RW, Liu KM, Rigby CL, Romanov E (2022) Mobula birostris (amended version of 2020 assessment). The IUCN red list of threatened species 2022: e.T198921A214397182
McCauley DJ, DeSalles PA, Young HS, Papastamatiou YP, Caselle JE, Deakos MH, Gardner J, Garton DW, Collen JD, Micheli F (2014) Reliance of mobile species on sensitive habitats: a case study of manta rays (Manta alfredi) and lagoons. Mar Biol 161:1987–1998
McGregor F, Richardson AJ, Armstrong AJ, Armstrong AO, Dudgeon CL (2019) Rapid wound healing in a reef manta ray masks the extent of vessel strike. PLoS ONE 14:e0225681
Murray A, Garrud E, Ender I, Lee-Brooks K, Atkins R, Lynam R, Arnold K, Roberts C, Hawkins J, Stevens G (2020) Protecting the million-dollar mantas; creating an evidence-based code of conduct for manta ray tourism interactions. J Ecotourism 19:132–147
Nicholson-Jack AE, Harris JL, Ballard K, Turner KME, Stevens GMW (2021) A hitchhiker guide to manta rays: patterns of association between Mobula alfredi, M. birostris, their symbionts, and other fishes in the Maldives. PLoS ONE 16:e0253704
O’Malley MP, Lee-Brooks K, Medd HB (2013) The global economic impact of manta ray watching tourism. PLoS ONE 8:e65051
O’Malley MP, Townsend KA, Hilton P, Heinrichs S, Stewart JD (2017) Characterization of the trade in manta and devil ray gill plates in China and South-east Asia through trader surveys. Aquat Conserv Mar Freshw Ecosyst 27:394–413
O’Shea OR, Kingsford MJ, Seymour J (2010) Tide-related periodicity of manta rays and sharks to cleaning stations on a coral reef. Mar Freshw Res 61:65–73
Peel LR, Stevens GMW, Daly R, Daly CAK, Lea JSE, Clarke CR, Collin SP, Meekan MG (2019a) Movement and residency patterns of reef manta rays Mobula alfredi in the Amirante Islands, Seychelles. Mar Ecol Prog Ser 621:169–184. https://doi.org/10.3354/meps12995
Peel LR, Daly R, Keating Daly CA, Stevens GMW, Collin SP, Meekan MG (2019b) Stable isotope analyses reveal unique trophic role of reef manta rays (Mobula alfredi) at a remote coral reef. R Soc Open Sci 6(9):190599. https://doi.org/10.1098/rsos.190599
Peel LR, Stevens GMW, Daly R, Keating Daly CA, Collin SP, Nogués J, Meekan MG (2020) Regional movements of reef manta rays (Mobula alfredi) in Seychelles waters. Front Mar Sci 7:1–17. https://doi.org/10.3389/fmars.2020.00558
Potts GW (1973) The ethology of Labroides dimidiatus (cuv. & val.) (Labridae, Pisces) on Aldabra. Anim Behav 21:250–291
R Core Team (2017) R: a language and environment for statistical computing
Rohner CA, Pierce SJ, Marshall AD, Weeks SJ, Bennett MB, Richardson AJ (2013) Trends in sightings and environmental influences on a coastal aggregation of manta rays and whale sharks. Mar Ecol Prog Ser 482:153–168
Rohner CA, Flam AL, Pierce SJ, Marshall AD (2017) Steep declines in sightings of manta rays and devilrays (Mobulidae) in southern Mozambique. PeerJ Prepr 5:e3051v1
Setyawan E, Sianipar AB, Erdmann MV, Fischer AM, Haddy JA, Beale CS, Lewis SA, Mambrasar R (2018) Site fidelity and movement patterns of reef manta rays (Mobula alfredi: Mobulidae) using passive acoustic telemetry in northern Raja Ampat, Indonesia. Nat Conserv Res 3:17–31
Sleeman JC, Meekan MG, Fitzpatrick BJ, Steinberg CR, Ancel R, Bradshaw CJA (2010) Oceanographic and atmospheric phenomena influence the abundance of whale sharks at Ningaloo Reef, Western Australia. J Exp Mar Bio Ecol 382:77–81
Stevens GMW (2016) Conservation and population ecology of manta rays in the Maldives. PhD thesis, University of York, Heslington
Stevens GMW, Hawkins JP, Roberts CM (2018) Courtship and mating behaviour of manta rays Mobula alfredi and M. birostris in the Maldives. J Fish Biol 93:344–359
Stevens G, Fernando D, Dando M, Di Sciara GN (2019) Guide to the Manta and Devil Rays of the World. In: Guide to the Manta and Devil Rays of the World. Princeton University Press,
Stewart JD, Beale CS, Fernando D, Sianipar AB, Burton RS, Semmens BX, Aburto-Oropeza O (2016) Spatial ecology and conservation of Manta birostris in the Indo-Pacific. Biol Conserv 200:178–183
Stewart JD, Jaine FRA, Armstrong AJ, Armstrong AO, Bennett MB, Burgess KB, Couturier LIE, Croll DA, Cronin MR, Deakos MH, Dudgeon CL, Fernando D, Froman N, Germanov ES, Hall MA, Hinojosa-Alvarez S, Hosegood JE, Kashiwagi T, Laglbauer BJL, Lezama-Ochoa N, Marshall AD, McGregor F, di Sciara GN, Palacios MD, Peel LR, Richardson AJ, Rubin RD, Townsend KA, Venables SK, Stevens GMW (2018) Research priorities to support effective Manta and Devil Ray conservation. Front Mar Sci 5:1–27. https://doi.org/10.3389/fmars.2018.00314
Stoddart DR, Coe MJ, Fosberg FR (1979) D’Arros and St. Joseph, Amirante Islands
Strike EM, Harris JL, Ballard KL, Hawkins JP, Crockett J, Stevens GMW (2022) Sublethal injuries and physical abnormalities in Maldives manta rays, Mobula alfredi and M. birostris. Front Mar Sci 270
Temple AJ, Wambiji N, Poonian CNS, Jiddawi N, Stead SM, Kiszka JJ, Berggren P (2019) Marine megafauna catch in southwestern Indian Ocean small-scale fisheries from landings data. Biol Conserv 230:113–121
Town C, Marshall A, Sethasathien N (2013) Manta Matcher: automated photographic identification of manta rays using keypoint features. Ecol Evol 3:1902–1914
Venables S, McGregor F, Brain L, van Keulen M (2016) Manta ray tourism management, precautionary strategies for a growing industry: a case study from the Ningaloo Marine Park, Western Australia. Pacific Conserv Biol 22:295–300
Venables SK, van Duinkerken DI, Rohner CA, Marshall AD (2020) Habitat use and movement patterns of reef manta rays Mobula alfredi in southern Mozambique. Mar Ecol Prog Ser 634:99–114. https://doi.org/10.3354/meps13178
Ward-Paige CA, Davis B, Worm B (2013) Global population trends and human use patterns of Manta and Mobula rays. PLoS ONE 8:e74835
Weideman EA, Perold V, Donnarumma V, Suaria G, Ryan PG (2023) Proximity to coast and major rivers influence the density of floating microplastics and other litter in east African coastal waters. Mar Pollut Bull 188:114644
White WT, Corrigan S, Yang LEI, Henderson AC, Bazinet AL, Swofford DL, Naylor GJP (2018) Phylogeny of the manta and devil rays (Chondrichthyes: Mobulidae), with an updated taxonomic arrangement for the family. Zool J Linn Soc 182:50–75
Wood S, Wood MS (2015) Package ‘mgcv.’ R Package Version 1 1:729
Zimmerhackel JS, Rogers AA, Meekan MG, Ali K, Pannell DJ, Kragt ME (2018) How shark conservation in the Maldives affects demand for dive tourism. Tour Manag 69:263–271
Zimmerhackel JS, Kragt ME, Rogers AA, Ali K, Meekan MG (2019) Evidence of increased economic benefits from shark-diving tourism in the Maldives. Mar Policy 100:21–26
Zuur AF, Ieno EN, Walker NJ, Saveliev AA, Smith GM (2009) Mixed effects models and extensions in ecology with R. Springer, New York
Acknowledgements
The authors would like to thank The Alphonse Foundation and The University of Western Australia’s Postgraduate Students Association for providing additional funding to support the research at St. François Atoll, as well as Rainer von Brandis, Christopher Boyes and Grace Phillips for their assistance in establishing the manta database at D’Arros Island. In addition, the authors thank Justin Blake, Luke Gordon, Ariadna Fernández, Christopher Narty and Lucy Martin for their invaluable assistance in the field; the ICS staff who have contributed identification photographs throughout the Outer Islands; John Nevill and Environment Seychelles for providing data on mantas encountered in the Mahé Fish market and the citizens of Seychelles who have contributed their valuable sighting records to the Seychelles Manta Ray Project. The authors also thank the editor and two anonymous reviewers for their valuable feedback on an earlier version of this manuscript.
Funding
Open Access funding enabled and organized by CAUL and its Member Institutions. This study was funded by the Save Our Seas Foundation (SOSF) and supported by The Manta Trust, The University of Western Australia, and the Australian Institute of Marine Science. Field work was supported by the SOSF-D’Arros Research Centre, Island Conservation Society (ICS), The Alphonse Foundation, Islands Development Company, Blue Safari Seychelles and the UWA Postgraduate Student’s Association.
Author information
Authors and Affiliations
Contributions
LP, RD, GS, SC and MM contributed to the study conception and design. Material preparation and data collection were performed by LP, RD, CK, GS and JN. Data analysis was completed by LP. The first draft of the manuscript was written by LP as part of a PhD thesis, and all authors commented on subsequent versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no competing interests.
Ethics approval
All research was conducted in accordance with the approval of The University of Western Australia’s Animal Ethics Committee (Permit RA/100/3/1480) and permissions from The Seychelles Bureau of Standards (A1057) and The Seychelles Ministry for Environment, Energy and Climate Change.
Additional information
Responsible Editor: J. Carlson.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Peel, L.R., Meekan, M.G., Daly, R. et al. Remote hideaways: first insights into the population sizes, habitat use and residency of manta rays at aggregation areas in Seychelles. Mar Biol 171, 83 (2024). https://doi.org/10.1007/s00227-024-04405-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00227-024-04405-6