
Abstract
Liver condition is hypothesized to be a better condition index for juvenile cod than other somatic condition indices, since cod store most of their excess energy in their liver. It is suggested that population changes in liver condition can be used as a proxy for overwinter mortality. Here, we have collected 0- and 1-group cod from the same cohort in the Barents Sea before and at the end of the winter season. We have taken measurements for somatic- and liver-based condition indices, as well as enumerating nematode endoparasites found in each individual cod. The average wet weight of sampled cod increased by 85% (approx. 0.4% day−1) from 6.0 to 10.1 g from September to February, while the average liver wet weight decreased by 15%. The relative water content of the liver also increased significantly over the winter, resulting in a 42% reduction in average dry liver weight from 0.116 to 0.067 g over the 4–5-month period between samplings. About 30% of the 1-group cod surviving the winter had less than 20% of the liver energy content of same-sized 0-group cod. The liver condition was also 5–10% lower in cod with high nematode endoparasite load than the cod without nematode endoparasites. The significance of the loss of liver energy content is discussed in relation to the marked seasonality combined with climate change and the challenges this imposes on the survival of 0-group cod in high-latitude environments during their first winter.
Similar content being viewed by others
Introduction
Barents Sea (BS) cod (Gadus morhua) is a demersal fish species that typically spawn between March and May along the coast of Norway between 62°N and 71°N (Suthers and Sundby 1993; Sundby and Nakken 2008). Eggs and larvae are carried north- and eastward by the Norwegian Atlantic Current and the Norwegian Coastal Current (Bergstad et al. 1987; Skagseth et al. 2008). The larvae initially feed on copepod nauplii and copepodites, with Calanus finmarchicus as one of the most important prey items (Helle 1994; Pedersen and Fossheim 2008). During the larval stage, most of the surplus energy is allocated to rapid growth with little energy being stored as reserves (Fuiman 2002; Jordaan and Brown 2003; Folkvord 2005).
Metamorphosis from larvae to juvenile in BS cod occurs 2–3 months after hatching and juvenile 0-group cod remain in the pelagic zone until September–October when they will start settling (Dingsør 2005; Ottersen et al. 2014). Since the larval and post-larval stages are strongly affected by currents, distribution in the Barents Sea can be as much as 500–800 nm north and east from the spawning grounds (Bergstad et al. 1987). Timing of settlement, as well as body size, has been shown to be correlated to the post-settlement survival in cod and other species (Post and Evans 1989; Salvanes et al. 1994; Juanes 2007). The settled BS cod juveniles experience high predation pressure from several species, with older conspecifics being a major mortality source (Dolgov et al. 2011; Yaragina et al. 2009). Although their most important prey will still be euphausiids after settling, they will switch over to becoming more generalists in their diet (Dalpadado and Bogstad 2004). The juveniles will, unlike the larvae, store a notable fraction of the energy obtained, and the excess energy in the liver is almost exclusively stored in the form of lipids (Black and Love 1986).
Seasonality is characterized by annual partitioning into periods of growth and non-growth (Conover 1992; Varpe 2017). For cod, summer and autumn are regarded as important periods of growth, and juvenile cod condition and growth will typically be higher in summer and autumn, than in winter and spring (Hawkins et al. 1985). The winter period is characterized by both lower food availability and lower temperatures but can serve as a period of reduced predation due to shorter day length hours available for visual predation and lowered predator consumption rates. Older conspecifics are often not found in the coldest areas (Ciannelli et al. 2007), and young cod residing in relatively colder areas may thus benefit from reduced predation as well as reduced energy expenditure over the winter period. It has been shown in experimental studies that the over-winter survival of 0-group cod can be significantly affected by the body size (Gotceitas et al. 1999; Geissinger et al. 2021). Field studies on western Atlantic cod juveniles further suggest that pre-winter conditions and winter duration may significantly affect winter survival (Geissinger et al. 2022, 2023). There is thus an expected trade-off between pre-winter growth and conditions where higher growth may reduce size-dependent predation while higher conditions may reduce the risk of starvation (Jørgensen et al. 2014). Given the challenges a long dark winter period at the high (> 70 N) latitudes impose on the BS cod during the first winter, a closer examination of the condition of 0-group cod prior to their first winter is warranted. Also, the ongoing climate change with increasing temperatures, especially at these latitudes, represents an added metabolic challenge for species which may have limited energy reserves to survive the dark winter period.
Extensive survey efforts have been carried out to collect length and weight data of young cod in the Barents Sea (e.g., Eriksen et al. 2018; Skjoldal et al. 2022), but few recent attempts have been made to assess the somatic condition of young cod in this area (but see examples of older Russian contributions in Eriksen and Prozorkevich 2011). Since cod mainly store their excess energy as lipids in their liver (Black and Love 1986; Lambert and Dutil 1997a; Grant and Brown 1999), the liver condition index of 0-group cod is hypothesized to be a particularly good proxy for starvation-related mortality risk over the upcoming winter. Taking part in standardized fisheries surveys, we follow the 2018 cohort of BS cod, comparing both somatic and liver conditions of 0-group cod prior to the onset of their first winter with the condition of surviving cod 4–5 months later. Since numerous accounts of nematode endoparasites were found on the young cod in the study material, the effect of their presence on the size and condition of cod was also evaluated.
Materials and methods
Sampling and morphological measurements
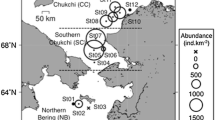
The cod in this study was sampled on two ecosystem survey cruises carried out by the Institute of Marine Research (IMR) with pre-set stations in the Barents Sea (Fig. 1). Our sampling targeted 0-group cod in the first survey cruise, the Barents Sea Ecosystem Survey (BESS, Eriksen et al. 2018), in the period between 19.09.2018 and 26.09.2018. In the second survey cruise, the winter survey, our sampling targeted 1-group cod belonging to the same 2018 cohort as the previously sampled 0-group cod, in the period between 03.02.2019 and 10.02.2019.

Map of the Barents Sea, showing sampling sites for 0-group (green circles) and 1-group (red triangles) Barents Sea cod with “Number” denoting the number of individuals sampled at each site
During the autumn ecosystem survey, the fish were collected using a pelagic Harstad trawl and a Campelen 1800 bottom trawl. Since the young cod will have settled by the time of the winter cruise, only the Campelen 1800 bottom trawl was used for this survey. The Campelen 1800 trawl, has a codend stretched mesh size of 22 mm. The Harstad trawl has a net circumference of 313 m, and a codend with 10 mm stretched mesh size (Irgens et al. 2017). Both trawls were towed at a speed was approximately 3.0 m/s. The standard IMR procedure for pelagic trawling at each station is to first tow for 10 min with headlines at 0 m and then 10 min with headlines at 20 and 40 m depth. Environmental variables were collected at the predetermined stations during both cruises using a Sea-bird SBE 9 CTD. Relatively minor differences were observed between the juvenile cod sampled with pelagic and bottom trawl during the BESS cruise (Suppl. Table 1), and the combined data were used from this cruise in the following analyses (Suppl. Table 2).
A total of 761 individuals of 1-group, and 726 individuals of 0-group, were sampled to be used in the main analyses. Of these, 195 were used to test for length shrinkage effects of frozen specimens. The fish were length measured on board before being frozen at −18 °C in plastic bags and stored for subsequent analysis in the laboratory together with the remainder of the material. Additional fish subsequently identified as 2-group fish are not included in the main analyses (see section on cohort identification). A maximum of 48 fish per station were measured for fish wet weight, WW (± 0.01 g) and total length, TL (± 1 mm). Both sagittae (hereafter otoliths) were extracted from sampled cod and stored in labelled NUNC containers for subsequent analyses.
Livers were extracted from all fish and placed in numbered and pre-weighed ceramic containers. Liver wet weight (LW), liver dry weight (LDW), and liver ash weight (LAW) were measured on a scale with an increment of 1 × 10–5 g. After LW was measured, LDW was obtained by drying the liver at 65 °C for 16 to 18 h. The liver dry weight (LDW) was obtained by subtracting the weight of the pre-weighed ceramic container from the weight of the container containing the dried liver. For 85 1-group cod, liver ash weight (LAW) was also obtained. This was done by placing the ceramic containers with dried fish livers into an incinerator at 550 °C for 16 to 18 h. The LAW after incineration was obtained by subtracting the weight of the pre-weighed ceramic containers from the containers containing the liver ash.
Cohort identification
During the BESS autumn survey, a minor length overlap was observed between the largest 0-group and the small 1-group (around 110–130 mm). The assignment of individual fish to the actual cohort was based on direct visual observation of otolith morphology as described in Irgens et al. (2017), where the 0-group was shown to have a markedly higher outer crenulation (higher circumference on an area basis). Cohort determination of young cod sampled during the winter cruise could not unequivocally be accomplished based on this criteria, and otoliths of fish in the overlapping size range (135–150 mm) were therefor weighed and sectioned for determination of age. The otoliths were embedded in epoxy resin and sectioned medially through the core with a Buehler Isomet low-speed saw. The 500 µm section was polished down to approx. 250 µm and photographed using a Nikon DS-Fi2 camera attached to a Leica M125 stereomicroscope for determination of inner annuli in the cod otoliths. The procedure provided an exact cohort assignment of all the young cod from the winter cruise where a relatively higher otolith weight at a given otolith area also matched the occurrence of inner annuli in the 2-group cod (Suppl. Figure 1).
Morphometric indices and statistical analysis
Statistical analyses and graphical presentations were done using RStudio (RStudio Team 2020) with R version 4.2.1. and Statistica (TIBCO Software Inc. 2017). The level of significance was set to α = 0.05. Linear mixed-effects models (nlme R-package) were used to test for differences in morphometric relations between sampled 0- and 1-group cod (Zuur and Ieno 2016). The sampling station was set as a random factor in all mixed effects models. Size variables were generally log transformed to stabilize the variance structure. The initial mixed-effects models were typically on the form:
lme(Condtion or Size measure 1 ~ Size measure 2 * Age group, random = ~ 1 | Station),
where non-significant interaction terms were subsequently removed based in favor of models with the lowest AIC (Zuur and Ieno 2016). Final models are listed in Suppl. Table 3 with condition or size measure 1 representing the more responsive morphological entity, and size measure 2 representing the less responsive morphological entity (Ferron and Leggett 1994).
Since the linear mixed effects model between fish length and weight confirmed that the fish did not have isometric growth (see “Results” section), a relative condition factor was used as a somatic condition factor. To obtain the relative condition factor, the length–weight relationship was first estimated using equation Eq. (1).
where ai is cohort-specific intercept and b is the common slope of both the 0- and 1-group cod from the mixed effects model. The relative condition factor (RCF) was estimated by Eq. (2):
where WWo represents the observed weight of the fish, and WWe is the estimated weight based on observed length, the common slope b, and a common intercept a (set to that of 0-group).
The hepatosomatic index (HSI), corresponding to the liver condition index defined by Yaragina and Marshall (2000), and the fatness index by Ponomarenko (1965) were created (Eq. 3):
where liver wet weight (LW) and fish wet weight (WW) are in grams.
A modified version of the liver condition index was created using the dry liver weight instead of liver wet weight (Eq. 4):
where LDW is liver dry weight in grams. Standard deviation and skewness were used to describe the distributions of the morphometric measures. The total amount of liver energy of an individual cod was estimated using the weight-specific liver energy content (LEC) relationship derived by Lambert and Dutil (1997a) multiplied by liver wet weight (Eq. 5):
where LEC is in kJ/g liver wet weight and LWC is liver water content (%).
Simplified tests of regional effects were carried out by dividing the sampling area into a western and an eastern region at a longitude of 25°E, as there was approximately the same number of stations between east (n = 20) and west (n = 22). Mixed-effects models were applied separately for each age cohort to test if there was any difference in the relationship between somatic- and liver-based condition measures between the two geographical regions.
Frequent occurrences of the nematode endoparasite Hysterothylacium aduncum were found inside several cavities (mostly in the abdomen) of the fish. The rate of parasite infestation per fish was categorized into three groups: none, some (1–3), and several (four and more). A log-linear model was used to test for regional and fish age effects on the frequency of infestation rates (TIBCO Software Inc. 2017).
Results
Separation of 1- and 2-group cod sampled during the winter survey
An individual separation of the largest 1-group and smallest 2-group cod in the overlapping size range during the winter survey was done by means of otolith-based aging. The 2-group cod had clearly visible inner annuli in addition to the translucent zone being formed at the outer edge of the otolith (Suppl. Fig. 1a). The results were confirmed by contrasting otolith weight relative to fish weight, with the older slower growing 2-group consistently having heavier otoliths at a given fish length than the faster growing 1-group cod (Suppl. Fig. 1b).
Fish length and weight distribution
The 0-group cod caught in September had a left tailed distribution (skewness = −0.866) with sizes ranging from 51 to 116 mm while the 1-group caught in February had a more symmetrical length distribution (skewness = 0.157) ranging from 80 to 150 mm (Fig. 2a). The average length increased from 92.2 to 116.4 mm. The corresponding increase in average weight was from 6.0 to 10.1 g (Fig. 2b). The 195 individuals that TL was measured fresh onboard the ship had an average length before freezing of 114.3 mm, and the average length two months later after freezing was 112.1 mm, corresponding to a shrinkage of 1.9% (paired t-test, t = 32.59, p < 0.001).

Distribution of A total length and B wet weight of 0- and 1-group cod of the 2018 cohort (n = 726 and n = 761)
Somatic relations and condition measures
The length–weight relation was allometric for both 0- and 1-group cod with a common exponent of 3.15 (lme, plength × age group interaction > 0.16, lme, pslope = 3 < 0.001). On average, the 0-group cod were 23% heavier at a given length than 1-group cod (lme, page group < < 0.001, Fig. 3a, Suppl. Table 3), and the residual error was 0.065. The relative condition (RCF) of 0-group cod was 1.00 in 0-group cod compared to 0.82 in 1-group cod (lme, page group < < 0.001, Suppl. Table 3).

A Regressions of fish wet weight (g) versus fish total length (mm), and B liver wet weight (g) versus fish weight (g) on a log–log scale for 0- and 1-group cod of the 2018 cohort
The liver wet weight versus fish weight relation was allometric and best described by a power function with a common slope of 1.31 (lme, pslope interaction > 0.05, pslope = 1 < 0.001, Suppl. Table 3).
On average, the 0-group cod had a 252% heavier wet liver weight at a given fish weight than 1-group cod (lme, page group << 0.001, Fig. 3b, Suppl. Table 3), with a residual error of 0.344. Corresponding analysis when using dry liver weight instead of wet liver weight yielded a common slope of 1.20 (lme, pslope interaction > 0.17, lme, pslope = 1 < 0.001, Suppl. Table 3). On average, the 0-group cod had a 356% heavier dry liver weight at a given fish weight than 1-group cod (lme, page group << 0.001, Suppl, Fig. 2 Suppl. Table 3), with a residual error of 0.439.
The hepatosomatic index was higher in 0- than 1-group cod and overall averaged 3.20 vs 1.57 (lme, page group << 0.001, Fig. 4a, Suppl. Table 3). The same was the case when using dry liver weight (DHSI), averaging 1.91 vs 0.65 (lme, page group << 0.001, Suppl. Table 3). The DSHI values were markedly right tailed (skewness = 2.15), with a notable proportion of the lower values piling up towards an apparent lower threshold around 0.4 (Fig. 4b). There was a great spread in HSI relative to fish length for both 0- and 1-group, although small fish within each cohort typically had low HSI (lme, plength < 0.008, Fig. 5, Suppl. Table 3). This also included the additional large fish from the winter survey that was shown by otolith analysis to be relatively small 2-group cod. These fish partly overlapped in size and HSI with some of the largest 1-group cod from the same survey (Fig. 5).

Distribution of A liver condition index, HSI (%) and B dry liver condition index, DHSI (%) of 0- and 1-group cod of the 2018 cohort (n = 726 and n = 761)

Liver condition index (HSI, %) versus fish total length (mm) for 0-group cod from the autumn BESS survey, and 1- and 2-group cod from the winter survey
Liver composition and energy content
The average water content in the liver was approx. 40 and 60% in 0- and 1-group cod livers, respectively (Suppl. Fig. 3), declining with HSI within each age group (lme, pHSI < 0.001, lme, page group < 0.001, Suppl. Table 3). The liver ash content was approx. 5% of liver wet weight in 1-group cod and declined with increasing HSI (Suppl. Fig. 4). The estimated total liver energy estimated using the individual water content and liver weight values revealed a significantly higher size-specific energy content in 0-group cod livers compared to 1-group cod livers (lme, page group < < 0.001, Fig. 6, Suppl. Table 3). About 30% of the 1-group cod had less than 20% of the average liver energy of similar-sized 0-group cod, and 2% had less than 10%.

Regressions of the total amount of liver energy (LE) versus fish wet weight (WW) of 0- and 1-group cod of the 2018 cohort. Lines corresponding to 10 and 20% of size-specific 0-group liver energy are indicated. Regression lines for 0-group and 1-group, respectively, are: log(LE) = −0.8282 + 1.182*log(WW), and log(LE) = −2.1596 + 1.182*log(WW)
Geographical differences in fish condition and parasite infestation
Both 0- and 1-group cod had tended to have higher liver-based condition indices in the western part of the sampled area than in the eastern part, e.g., dry liver condition index (lme, pregion < 0.051, Suppl. Table 3). This regional trend was not apparent when using relative somatic condition (lme, pregion > 0.6, Suppl. Table 3). When contrasting the western stations versus the eastern sampling stations, a higher proportion of infested juveniles were found in the eastern stations, both among 0-group and 1-group cod (log-linear model, p < 0.05, Suppl. Fig. 5). A slight increase in the degree of infestation was found from 0- to 1-group cod (Suppl. Fig. 5). Both 0- and 1-group cod with the highest parasite load had a 5–10% relative lower HSI than those with few or no visible endoparasites (lme, page group < 0.03, Fig. 7, Suppl. Table 3). This effect was not apparent when contrasting the against RCF (lme, page group > 0.3).

Liver condition index, HSI (%) of 0- and 1-group cod at three different grades of parasite infestation (None = 0 parasites, some parasites = 1–3 parasites, many parasites = 4 or more parasites).
Discussion
The condition of juvenile cod decreased significantly over the winter months in the Barents Sea, and this trend was most clearly seen in liver-based condition indices. On average only a third of the size-specific liver energy content in 0-group cod in September was left in the 1-group cod in February, pointing at the metabolic challenges the young are facing during the first winter period. Not only did the condition decline over the winter months, but a larger variability in relative liver weight was evident after the winter, suggesting differences in feeding success and/or metabolic efficiency of the surviving cod.
Seasonal variation in condition
Variation in seasonal condition was evident among 0- and 1-group cod, and this is in line with observations from earlier studies on cod (e.g., Hawkins et al. 1985; Lambert and Dutil 1997b; Grant and Brown 1999) and other species such as polar cod in high latitude environments (Aune et al. 2021). The total reduction in condition over the winter season could not accurately be estimated with only two cruises, however, with one carried out at around autumnal equinox, and the other one month prior to the vernal equinox, respectively. A continued feeding and build-up of reserves may have taken place after the first cruise, while a further depletion of energy reserves may still have taken place after the second cruise. The seasonal variation in conditions of older Barents Sea cod is highly dependent on the availability of capelin (Yaragina and Marshall 2000) and may reveal different seasonal patterns in condition than for young cod in the area. Maximum liver condition in young cod from the North-western Atlantic has been shown to occur in September–October (Grant and Brown 1999). Other studies on juvenile and adult cod have observed minimum condition around April–May (e.g., Eliassen and Vahl 1982; Hawkins et al. 1985; Lambert and Dutil 1997b). Seasonal minimum liver condition of the closely related polar cod in the eastern Barents Sea was observed in March, while the maximum levels in were observed in September (Aune et al. 2021). A further reduction in individual seasonal conditions over the winter is expected if there has been selective mortality of smaller low-condition individuals in the time interval between the two cruises. Results from studies on cod and other species have shown that the proportional weight loss under low food or starving conditions is larger in smaller individuals, and typically accompanied by higher mortality (Post and Evans 1989; Folkvord 1991). However, the liver condition (HSI) decreased in larger western Atlantic young cod while it increased in smaller cod over the winter when provided low experimental feeding rations (Geissinger et al 2021). As in our study, this can to some extent be confined by condition or size-selective mortality, and the change in seasonal condition is thus expected to have been larger than observed between the two cruises in our study.
Sensitivity differences of condition indices
The liver-based condition indices changed relatively more over the winter season than length–weight-based condition indices and are thus more sensitive to the winter-induced changes in environmental conditions. While there was only a 23% change in wet weight-at-length, the corresponding change in dry- and wet liver weight-at-length was over 250 and 350%, respectively. Further, liver-based condition indices detected regional differences in conditions in this study while other whole-body condition indices failed to do so. This is not surprising since the liver is the main energy storage in cod and several other gadoids (Black and Love 1986; Lambert and Dutil 1997a), and liver energy has been shown in experimental studies to be preferentially utilized during starvation (Black and Love 1986; Hemre et al. 1993). Liver-based condition indices have also been shown to be good proxies of energy density in other species (Wuenschel et al. 2019). Muscle protein which is the other main internally available energy source in cod, is preserved relative to liver lipids. This is expected as a reduction in muscle protein will also have additional negative effects related to reduced swimming capacity (Lambert and Dutil 1997a; Martinez et al. 2004). Muscle constitutes a higher mass than liver in fish, and in adult Atlantic cod around 50–60% of total mass will be composed of muscle tissue (Lambert and Dutil 1997a), while the liver in wild-caught fish as in this study typically accounts for less than 6%. In Pacific cod (Gadus macrocephalus) juveniles, the muscle content constituted up to 70% and the liver typically around 4% of body mass (Copeman et al. 2017). Still, the overall energy density is much higher in the cod liver than in the muscle tissue (approx. 3.5 to 6 times higher, Lambert and Dutil 1997a), with lower energy density being associated with higher water content in both tissues. For liver-based condition measures, the relative water content decreases slightly from higher to intermediate HSI levels, and only at the lower end will water content increase progressively as lipids are replaced (Lambert and Dutil 1997a). This was accompanied by an increase in relative liver ash weight as has also been noted in relative whole body ash content in cod (Grant and Brown 1999). In juvenile Pacific cod, the lipid density among fish reared in the range from 0 to 9 °C was 30–70 times higher in the liver than muscle, yielding in total 7.7–19.2 times higher lipid storage energy (triacyl glycerols, TAG) in the liver compared to muscle tissue (Copeman et al. 2017).
The choice of condition measure to be used in ecological studies will depend on the questions asked and the latency and dynamics of the condition measure itself (Ferron and Leggett 1994, Wuenschel et al. 2019). In the case of seasonal condition changes in cod at high latitudes, the liver-based condition indices are deemed to be highly applicable and responsive. As a measure of available energy at the 0-group stage, wet-weight-based liver measures may suffice, while dry liver-based conditions may be more informative for the early 1-group stage when energy reserves are low and approaching a critical level for survival. As seen in our study the dry liver condition index (DSHI) showed the most marked reduction over the time-period between the cruises, and a dry liver condition index is expected to be a better condition measure than a wet liver condition index as it is more closely related to available energy reserves (Lambert and Dutil 1997a). Further, the apparent lower DSHI threshold of around 0.25 may serve as a practical measure of young cod in a critically low condition. Even if it amounts to a higher sampling effort, we encourage the implementation of dry liver measures as a highly relevant condition measure of young Barents Sea cod.
Energetics and winter mortality
A noticeable fraction of the 1-group cod was in a poor energetic state and likely in a starving condition. Estimations of liver-based energy content suggested that on average more than two-thirds of the available liver energy was spent between the sampling periods, and that 2% of the 1-group cod had less than 10% average liver energy of similar-sized 0-group cod. High winter mortality is suggested to be one of the main drivers behind observed countergradient variations in growth rates, where fish from higher latitudes with shorter growing seasons are adapted to higher growth capacity to reach a minimum size prior to their first winter (Conover 1992). As larger fish typically have larger energy reserves and at the same time have lower size-specific metabolic costs (Edwards et al. 1972; Schultz and Conover 1999), this may be important to cope with the winter season with less feeding opportunities (Post and Evans 1989; Hurst and Conover 1998). An experimental study on cod from Newfoundland showed that larger 0-group cod had lower total mortality than smaller cod over a six-month period (42 vs 86%) at both food levels offered (0.25 and 1% of body weight day−1) (Gotceitas et al. 1999). The growth rate of the surviving smallest individuals was higher than that of the larger cod, in line with a higher energetic turnover in these fish. This is in contrast to a more recent study with cod from Newfoundland by Geissinger et al. (2021) where the low food rations (less than 0.5% of body weight) over the winter yielded relatively low mortality for both small and large cod juveniles. A study of the lipid levels of the gadid walleye pollack, Theragra chalcgramma, also revealed that smaller individuals generally had relatively lower lipid levels which was linked to condition-dependent mortality during the winter (Sogard and Olla 2000). A combined laboratory and field study on age-0 Pacific herring (Clupea pallasi) found that the whole body energy content found in field-caught specimens towards the end of the winter, matched those found in specimens starved in the laboratory for a 122-day period over the winter (Paul and Paul 1998). The inferior feeding conditions during the winter season in high latitudes are related both to the lower productivity during this period, but also due to the reduced visual prey detection rates caused by low light intensities during winter darkness (Varpe and Fiksen 2010). Fish species at high latitudes may thus face increasing energy deficit in a light-limiting environment during winter if related metabolic costs increase with increasing temperature.
Growth and mortality trade offs
A large variation in individual liver-based conditions was observed among 0- and 1-group in this study and an added effort was made by the use of otoliths to clarify the year-class and cohort origin of fish compared between survey periods (Irgens et al. 2017). Normally, the relatively minor size overlap will not represent a problem in the quantitative estimation of proportions of cod being either 0, 1, or 2-group, but in terms of individual condition, it is important to determine if fish in the overlapping size ranges are among the largest of the youngest cohort, or the smallest of the older. The emerging pattern of size-dependent condition was at first somewhat surprising in that some of the larger 0- and 1 group cod had a low liver condition, which was also recently observed in an experimental study on young cod from the western Atlantic (Geissinger et al. 2021). As expected, the smallest individuals in either age group in our study had relatively low conditions. These results point to the likely growth-mortality trade-offs young cod are facing during the early stages (Jørgensen et al. 2014). The cod need to balance the benefits of feeding to secure high growth and the risks of not being vigilant towards predators, and during the larval stages surviving cod larvae in the field seem to be growing close to their size-dependent capacity (Folkvord 2005), possibly due to higher size selective morality of relatively smaller individuals. Unlike cod larvae, the juvenile will accumulate higher amounts of lipids in their liver, providing them with a better capacity to cope with starvation-induced mortality (Folkvord 1991; Jordaan and Brown 2003). The strategy of prioritizing growth among high-latitude populations relative to lower latitude populations is well documented (Conover 1992; Lankford et al. 2001) and is in part driven by the need to obtain sufficiently large size to cope with the upcoming first winter. Unlike the case for low latitude species, such as for example. dwarf perch, Micrometrus minimus (Schultz 1993), the marked seasonality and limited period with sufficient light and production, provides limited possibilities to adjust by fitness/size of offspring by simply shifting the timing of reproduction (Varpe 2017). The low condition even in some of the largest individuals of a cohort in this study may indicate past tradeoffs in these fish in favor of growth and potential predator avoidance rather than energy accumulation to reduce starvation risk. A similar finding was seen among young-of -the-year herring (Clupea pallasi) in Prince William Sound, Alaska, where herring smaller than 75 mm prioritized growth over lipid storage (Sewall et al. 2019). The success of these trade-offs will naturally depend on the regional predation pressure as well as energy availability and consumption over the winter.
Regional differences and effects of climate
The regional difference in cod condition was relatively small over the studied area and only significant when based on dry-liver content. For both age groups, the condition was higher in the western part which could be linked to generally higher temperatures due to the proximity of the warmer inflowing Atlantic water (Suppl. Fig. 6, Ottersen 2010). Spatial differences in condition may thus be minor as also found for the closely related 0-group Pacific cod (Abookire et al. 2022), even though annual variations in condition were significantly correlated with seasonal temperatures in their study. It should be noted, however, that the samples in our study only originated from the north-western part of the Barents Sea. Results from earlier studies did not document any apparent regional association between higher prey concentrations and the size of young cod, while the abundance of juveniles was higher when plankton biomass was higher (Helle 1994). Larger length-at-age has previously been documented to be associated with higher overall ambient temperature conditions in the entire Barents Sea at the annual level (e.g., Ottersen and Loeng 2000) and in sub-areas in the Barents Sea (e.g., Ponomarenko 1965). However, the link between temperature, food levels, and the size and abundance of 0-group cod may be more complex as a higher initial inflow of Atlantic water containing plankton and young cod may result in a more easterly distribution of larger and possibly older 0-group cod, eventually inhabiting colder temperature conditions (Helle 1994). Survival and size of early cod juveniles are suggested to be affected by the co-occurrence of young cod and plankton, both being a result of the higher influx of plankton-rich Atlantic water to the Barents Sea (Helle and Pennington 1999; Ottersen et al. 2002). Further analysis of spatial distribution and abundance of cod indicated that both density and fish length were reduced at the highest temperatures compared to those at mean core thermal habitats (Eriksen et al. 2012), suggesting that there is an upper limit to the growth benefits of climate-induced elevated temperatures in the region. As experimental studies with 8 g cod being fed in excess yielded higher somatic conditions at higher temperatures (Árnason et al. 2009), this suggests that the benefits of overall increased plankton availability at the 0-group stage are not sufficient to compensate for elevated metabolic costs at higher temperatures. This effect is likely to be exacerbated with general warming over the winter months when prey availability, either in numbers or visibility, is insufficient to compensate for the basic metabolic costs of young cod. Temperature-related differences in the condition of early juveniles were also noted for YOY herring, where the highest lipid stores were observed in the coldest years, rather than the years with the most preferable diet availability (Sewall et al. 2019). At high latitudes, an increased temperature-driven prey search capability will not necessarily suffice if limited prey detection at unaltered winter light levels remains the main limiting factor. It is thus expected that the nutritional challenges imposed on the young cod, and the influence of pre-winter condition, are expected to more important under future climate change.
A regional difference in the occurrence of the endoparasite Hysterothylacium aduncum was found, and individuals with higher parasite load had lower liver conditions than those with none or few parasites. High infestation rates of H. aduncum have also been found in post-larval cod in the North Sea, where infestation rates were typically higher in larger and presumably older individuals (Skovgaard et al. 2011). The intermediate hosts of H. aduncum are copepods and other marine invertebrates, and the observed difference in parasite infestation in young cod in pour study is thus expected to represent either a higher spatial overlap with the intermediate hosts or a longer temporal exposure due to older age of 0-group cod in the eastern part of the Barents Sea. A higher feeding motivation of an infested host due to parasite–host interactions can potentially induce a higher growth and condition of infested individuals (Barber et al. 2000). This does not seem to be the case in our study, suggesting that the infestation of parasites resulted in a net energy loss compared to non-infested specimens. The observed differences in condition are minimum estimates, however, as parasite weight was included in the total weight of the infested fish (Lagrue and Poulin 2015). In any case, the reduced condition of infested vs non-infested specimens was relatively minor compared to the seasonal changes in condition and are thus only expected to have a direct fitness impact under otherwise marginal situations.
Recruitment aspects and future perspectives
The mortality of young cod during their first winter is expected to be high and variable due to variations in prior feeding conditions and predation pressure (Bogstad et al. 2016). Although there is a notable correlation between abundance at post-larval stages and subsequent 0-group stages (Sundby et al. 1989), the age of recruitment in Barents Sea cod is typically set at age 3 in fisheries assessment (e.g., Bogstad et al. 1994). At this point, the strength of the year-class is presumed to be relatively fixed, but there is a potential to utilize biological information from earlier stages to reduce prediction uncertainty in the assessment of the stock by providing earlier reliable measures of recruitment (Ulltang 1996). Given the observed temporal covariation in 0-group length of related fish species in the Barents Sea, large-scale climatic (e.g., temperature) or productivity (e.g., food availability) processes are likely important (Skjoldal et al. 2022). The extent of first winter mortality has been shown to greatly influence recruitment and year-class strength in striped bass (Morone saxatilis) (Hurst and Conover 1998). In their study, selective mortality of smaller individuals was only apparent in some of the years studied, and this was suggested to be linked to low temperatures limiting subsequent growth. These observations were based on length measures, however, and estimates of size-specific energy reserves of the overwintering fish were not provided. Field studies on Atlantic cod off Newfoundland demonstrated that pre-winter condition was important for survival when the winter conditions were prolonged, while contrary to expectation, smaller fish from late-arriving pulses survived better than those from earlier pulses when winter conditions were of shorter duration (Geissinger et al. 2023). Autumn lipid accumulation and the first winter conditions were also shown to be critical for the overall survival and first-year recruitment success of largemouth bass (Micropterus salmoides) (Ludsin and DeVries 1997). In line with Heintz et al. (2013) who found a positive correlation between total energy and recruitment of 0-group pollack in the eastern Bering Sea, we thus argue that the use of qualitative aspects of 0- and 1-groups cod beyond quantitative abundance and length estimates may provide such valuable additional information, especially in the light of expected higher first winter mortality. Given future changes and uncertainties in cod population dynamics following global warming (Kjesbu et al. 2023), it is essential for future stock assessments to obtain informative and relevant biological information to document expected climate effects on all life stages. As noted by Hurst (2007), few stock assessments and management regimes take explicitly into account variability in winter mortality. We have reason to believe that the first overwinter mortality will be increasingly challenging for high-latitude populations such as the Barents Sea cod, resulting in more variable recruitment than before. The added temperature-mediated increase in metabolism due to climate change combined with the unaltered light-limited feeding opportunities during winter darkness may also infer a more severe challenge for the Barents Sea cod than for other cod populations at lower latitudes where light-limited feeding opportunities may be less of a problem. Adding additional informative qualitative information about the young cod, such as liver condition measures, either on a wet- or dry basis may thus be warranted and justified to ensure future high-quality recruitment predictions and stock assessments.
Data availability
Datasets generated and analyzed in the study are available from the corresponding author on reasonable request.
References
Abookire AA, Litzow MA, Malick MJ, Laurel BJ (2022) Post-settlement abundance, condition, and survival in a climate-stressed population of Pacific cod. Can J Fish Aquat Sci 79:958–968. https://doi.org/10.1139/cjfas-2021-0224
Árnason T, Björnsson B, Steinarsson A (2009) Allometric growth and condition factor of Atlantic cod (Gadus morhua) fed to satiation: effects of temperature and body weight. J Appl Ichthyol 25:401–406. https://doi.org/10.1111/j.1439-0426.2009.01259.x
Aune M, Raskhozheva E, Andrade H, Augustine S, Bambulyak A, Camus L, Carroll J, Dolgov AV, Hop H, Moiseev D, Renaud PE, Varpe Ø (2021) Distribution and ecology of polar cod (Boreogadus saida) in the eastern Barents Sea: a review of historical literature. Marine Environ Res 166:105262. https://doi.org/10.1016/j.marenvres.2021.105262
Barber I, Hoare D, Krause J (2000) Effects of parasites on fish behaviour: a review and evolutionary perspective. Rev Fish Biol Fish 10:131–165. https://doi.org/10.1023/A:1016658224470
Bergstad OA, Jørgensen T, Dragesund O (1987) Life history and ecology of the gadoid resources of the Barents Sea. Fish Res 5:119–161
Black D, Love RM (1986) The sequential mobilization and restoration of energy reserves in tissues of Atlantic cod during starvation and refeeding. J Comp Physiol B 156:469–479. https://doi.org/10.1007/BF00691032
Bogstad B, Lilly G, Mehl S, Pálsson OK, Stefánsson G (1994) Cannibalism and year-class strength in Atlantic cod (Gadus morhua L.) in the Arcto-boreal ecosystems (Barents Sea, Iceland and Eastern Newfoundland). ICES Mar Sci Symp 198:576–599
Bogstad B, Yaragina NA, Nash RDM (2016) The early life-history dynamics of Northeast Arctic cod: levels of natural mortality and abundance during the first 3 years of life. Can J Fish Aquat Sci 73:246–256. https://doi.org/10.1139/cjfas-2015-0093
Ciannelli L, Dingsør GE, Bogstad B, Ottersen G, Chan K-S, Gjøsæter H, Stiansen JE, Stenseth NC (2007) Spatial anatomy of species survival: effects of predation and climate-driven environmental variability. Ecology 88:635–646. https://doi.org/10.1890/05-2035
Conover DO (1992) Seasonality and the scheduling of life history at different latitudes. J Fish Biol 41:161–178
Copeman LA, Laurel BJ, Spencer M, Sremba A (2017) Temperature impacts on lipid allocation among juvenile gadid species at the Pacific Arctic-Boreal interface: an experimental laboratory approach. Mar Ecol Prog Ser 566:183–198
Dalpadado P, Bogstad B (2004) Diet of juvenile cod (age 0–2) in the Barents Sea in relation to food availability and cod growth. Polar Biol 27:140–154. https://doi.org/10.1007/s00300-003-0561-5
Dingsør GE (2005) Estimating abundance indices from the international 0-group fish survey in the Barents Sea. Fish Res 72:205–218. https://doi.org/10.1016/j.fishres.2004.11.001
Dolgov AV, Bogstad B, Johannesen E, Skern-Mauritzen M (2011) An overview of trophic interactions in the Barents Sea. In: Jakobsen T, Ozhigin VK (eds) The Barents Sea – ecosystem, resources, management. Tapir Academic Press, Trondheim, pp 431–437
Edwards RRC, Finlayson DM, Steele JH (1972) An experimental study of the oxygen consumption, growth, and metabolism of the cod (Gadus morhua L.). J Exp Mar Biol Ecol 8:299–309
Eliassen JE, Vahl O (1982) Seasonal variations in biochemical composition and energy content of liver, gonad and muscle of mature and immature cod, Gadus morhua (L) from Balsfjorden, northern Norway. J Fish Biol 20:707–716. https://doi.org/10.1111/j.1095-8649.1982.tb03981.x
Eriksen E, Prozorkevich DV (2011) 0-group surveys. In: Jakobsen T, Ozhigin VK (eds) The Barents Sea – Ecosystem, resources, management Half a century of Russian-Norwegian cooperation. Tapir Academic Press, Trondheim, pp 557–569
Eriksen E, Ingvaldsen R, Stiansen JE, Johansen GO (2012) Thermal habitat for 0-group fish in the Barents Sea; how climate variability impacts their density, length, and geographic distribution. ICES J Mar Sci 69:870–879. https://doi.org/10.1093/icesjms/fsr210
Eriksen E, Gjøsæter H, Prozorkevich D, Shamray E, Dolgov A, Skern-Mauritzen M, Stiansen JE, Kovalev Y, Sunnanå K (2018) From single species surveys towards monitoring of the Barents Sea ecosystem. Prog Oceanogr 166:4–14. https://doi.org/10.1016/j.pocean.2017.09.007
Ferron A, Leggett WC (1994) An appraisal of condition measures for marine fish larvae. Adv Mar Biol 30:217–303
Folkvord A (1991) Growth, survival and cannibalism of cod juveniles (Gadus morhua L.): effects of feed type, starvation and fish size. Aquaculture 97:41–59. https://doi.org/10.1016/0044-8486(91)90278-F
Folkvord A (2005) Comparison of size-at-age of larval Atlantic cod (Gadus morhua) from different populations based on size- and temperature-dependent growth models. Can J Fish Aquat Sci 62:1037–1052. https://doi.org/10.1139/f05-008
Fuiman LA (2002) Special considerations of fish eggs and larvae. In: Fuiman LA, Werner RG (eds) Fishery science – The unique contributions of early life stages. Blackwell Publishing, Oxford, pp 1–32
Geissinger EA, Gregory RS, Laurel BJ, Snelgrove PVR (2021) Food and initial size influence overwinter survival and condition of a juvenile marine fish (age-0 Atlantic cod). Can J Fish Aquat Sci 78:472–482. https://doi.org/10.1139/cjfas-2020-0142
Geissinger EA, Gregory RS, Laurel BJ, Snelgrove PVR (2022) High site-fidelity and low mortality of juvenile Atlantic cod (Gadus morhua) in subarctic coastal habitat during their first winter. ICES J Mar Sci 79:1408–1418. https://doi.org/10.1093/icesjms/fsac065
Geissinger EA, Bloom CR, Gregory RS, Laurel BJ, Snelgrove PVR (2023) Condition, size, and winter duration affect winter survival probability of juvenile Atlantic cod Gadus morhua in a coastal subarctic ecosystem. Mar Ecol Prog Ser 711:47–60. https://doi.org/10.3354/meps14310
Gotceitas V, Methven DA, Fraser S, Brown JA (1999) Effects of body size and food ration in over-winter survival and growth of age-0 Atlantic cod, Gadus morhua. Environ Biol Fishes 54:413–420
Grant SM, Brown JA (1999) Variation in condition of coastal Newfoundland 0-group Atlantic cod (Gadus morhua): field and laboratory studies using simple condition indices. Mar Biol 133:611–620
Hawkins AD, Soofiani NM, Smith GW (1985) Growth and feeding of juvenile cod (Gadus morhua L.). J Conseil Int Pour L’explor De La Mer 42:11–32
Heintz RA, Siddon EC, Farley EV, Napp JM (2013) Correlation between recruitment and fall condition of age-0 pollock (Theragra chalcogramma) from the eastern Bering Sea under varying climate conditions. Deep Sea Res Part II Top Stud Oceanogr 94:150–156. https://doi.org/10.1016/j.dsr2.2013.04.006
Helle K (1994) Distribution of early juvenile Arcto-Norwegian cod (Gadus morhua L.) in relation to food abundance and water mass properties. ICES Mar Sci Symp 198:440–448
Helle K, Pennington M (1999) The relation of the spatial distribution of early juvenile cod (Gadus morhua L.) in the Barents Sea to zooplankton density and water flux during the period 1978–1984. ICES J Mar Sci 56:15–27. https://doi.org/10.1006/jmsc.1998.0427
Hemre G-I, Karlsen Ø, Lehmann G, Holm JC, Lie Ø (1993) Utilization of protein, fat and glycogen in cod (Gadus morhua) during starvation. Fisk Skr Serie Ernær 6:1–9
Hurst TP (2007) Causes and consequences of winter mortality in fishes. J Fish Biol 71:315–345. https://doi.org/10.1111/j.1095-8649.2007.01596.x
Hurst TP, Conover DO (1998) Winter mortality of young-of-the-year Hudson River striped bass (Morone saxatilis): size-dependent patterns and effects on recruitment. Can J Fish Aquat Sci 55:1122–1130. https://doi.org/10.1139/f98-017
Irgens C, Kjesbu OS, Folkvord A (2017) Ontogenetic development of otolith shape during settlement of juvenile Barents Sea cod (Gadus morhua). ICES J Mar Sci 74:2389–2397. https://doi.org/10.1093/icesjms/fsx088
Jordaan A, Brown JA (2003) The risk of running on empty: the influence of age on starvation and gut fullness in larval Atlantic cod (Gadus morhua). Can J Fish Aquat Sci 60:1289–1298
Jørgensen C, Opdal AF, Fiksen Ø (2014) Can behavioural ecology unite hypotheses for fish recruitment? ICES J Mar Sci 71:909–917. https://doi.org/10.1093/icesjms/fst083
Juanes F (2007) Role of habitat in mediating mortality during the post-settlement transition phase of temperate marine fishes. J Fish Biol 70:661–677. https://doi.org/10.1111/j.1095-8649.2007.01394.x
Kjesbu OS, Alix M, Sandø AB, Strand E, Wright PJ, Johns DG, Thorsen A, Marshall CT, Bakkeplass KG, Vikebø FB, Skuggedal Myksvoll M, Ottersen G, Allan BJM, Fossheim M, Stiansen JE, Huse G, Sundby S (2023) Latitudinally distinct stocks of Atlantic cod face fundamentally different biophysical challenges under on-going climate change. Fish Fish 24:297–320. https://doi.org/10.1111/faf.12728
Lagrue C, Poulin R (2015) Measuring fish body condition with or without parasites: does it matter? J Fish Biol 87:836–847. https://doi.org/10.1111/jfb.12749
Lambert Y, Dutil JD (1997a) Can simple condition indices be used to monitor and quantify seasonal changes in the energy reserves of cod (Gadus morhua)? Can J Fish Aquat Sci 54:104–112. https://doi.org/10.1139/f96-149
Lambert Y, Dutil JD (1997b) Condition and energy reserves of Atlantic cod (Gadus morhua) during the collapse of the northern Gulf of St. Lawrence stock. Can J Fish Aquat Sci 54:2388–2400. https://doi.org/10.1139/f97-145
Lankford TE, Billerbeck JM, Conover DO (2001) Evolution of intrinsic growth and energy acquisition rates. II. Trade-offs with vulnerability to predation in Menidia menidia. Evolution 55:1873–1881
Ludsin SA, DeVries DR (1997) First-year recruitment of largemouth bass: the interdependency of early life stages. Ecol Appl 7:1024–1038
Martinez M, Bedard M, Dutil JD, Guderley H (2004) Does condition of Atlantic cod (Gadus morhua) have a greater impact upon swimming performance at Ucrit or sprint speeds? J Exp Biol 207:2979–2990. https://doi.org/10.1242/jeb.01142
Ottersen G (2010) A digital temperature atlas for the Norwegian Sea. ICES J Mar Sci 67:1525–1537. https://doi.org/10.1093/icesjms/fsq099
Ottersen G, Loeng H (2000) Covariability in early growth and year-class strength of Barents Sea cod, haddock, and herring: the environmental link. ICES J Mar Sci 57:339–348
Ottersen G, Helle K, Bogstad B (2002) Do abiotic mechanisms determine interannual variability in length-at-age of juvenile Arcto-Norwegian cod? Can J Fish Aquat Sci 59:57–65
Ottersen G, Bogstad B, Yaragina NA, Stige LC, Vikebø FB, Dalpadado P (2014) A review of early life history dynamics of Barents Sea cod (Gadus morhua). ICES J Mar Sci 71:2064–2087. https://doi.org/10.1093/icesjms/fsu037
Paul AJ, Paul JM (1998) Comparisons of whole body energy content of captive fasting age zero Alaskan Pacific herring (Clupea pallasi Valenciennes) and cohorts over-wintering in nature. J Exp Marine Biol Ecol 226:75–86. https://doi.org/10.1016/S0022-0981(97)00241-4
Pedersen T, Fossheim M (2008) Diet of 0-group stages of capelin (Mallotus villosus), herring (Clupea harengus) and cod (Gadus morhua) during spring and summer in the Barents Sea. Mar Biol 153:1037–1046. https://doi.org/10.1007/s00227-007-0875-x
Ponomarenko IJ (1965) Comparative characteristics of some biological indices of the bottom stages of 0-group cod belonging to the 1956, 1958, 1959, 1960 and 1961 year-classes. ICNAF Spec Publ 6:349–354
Post JR, Evans DO (1989) Size-dependent overwinter mortality of young-of-the-year yellow perch (Perca flavescens): laboratory, in situ enclosure, and field experiments. Can J Fish Aquat Sci 46:1958–1968
Rstudio Team (2020) Rstudio: integrated development for R. PBC, Boston
Salvanes AGV, Giske J, Nordeide JT (1994) Life-history approach to habitat shifts for coastal cod, Gadus morhua L. Aquac Fish Manage 25:215–228
Schultz ET (1993) The effect of birth date on fitness of female dwarf perch, Micrometrus minimus (Perciformes, Embiotocidae). Evolution 47:520–539. https://doi.org/10.2307/2410069
Schultz ET, Conover DO (1999) The allometry of energy reserve depletion: test of a mechanism for size-dependent winter mortality. Oecologia 119:474–483. https://doi.org/10.1007/s004420050810
Sewall F, Norcross B, Vollenweider J, Heintz R (2019) Growth, energy storage, and feeding patterns reveal winter mortality risks for juvenile Pacific herring in Prince William Sound, Alaska, USA. Mar Ecol Prog Ser 623:195–208
Skagseth Ø, Furevik T, Ingvaldsen R, Loeng H, Mork KA, Orvik KA, Ozhigin V (2008) Volume and heat transports to the arctic ocean via the Norwegian and Barents Seas. In: Dickson RR, Meincke J, Rhines P (eds) Arctic-Subarctic ocean fluxes. Springer, Dordrecht
Skjoldal HR, Eriksen E, Gjøsæter H, Skagseth Ø, Prozorkevich D, Lien VS (2022) Recruitment variability of fish stocks in the Barents Sea: Spatial and temporal variation in 0-group fish length of six commercial species during recent decades of warming (1980–2017). Prog Oceanogr 206:102845. https://doi.org/10.1016/j.pocean.2022.102845
Skovgaard A, Bahlool QZM, Munk P, Berge T, Buchmann K (2011) Infection of North Sea cod, Gadus morhua L., larvae with the parasitic nematode Hysterothylacium aduncum Rudolphi. J Plankton Res 33:1311–1316. https://doi.org/10.1093/plankt/fbr027
Sogard SM, Olla BL (2000) Endurance of simulated winter conditions by age-0 walleye pollock: effects of body size, water temperature and energy stores. J Fish Biol 56:1–21. https://doi.org/10.1111/j.1095-8649.2000.tb02083.x
Sundby S, Nakken O (2008) Spatial shifts in spawning habitats of Arcto-Norwegian cod related to multidecadal climate oscillations and climate change. ICES J Mar Sci 65:953–962. https://doi.org/10.1093/icesjms/fsn085
Sundby S, Bjørke H, Soldal AV, Olsen S (1989) Mortality rates during the early life stages and year-class strength of northeast Arctic cod (Gadus morhua L.). Rapp P-V Réun Cons Int Pour L’explor Mer 191:351–358
Suthers IM, Sundby S (1993) Dispersal and growth of pelagic juvenile Arcto-Norwegian cod (Gadus morhua), inferred from otolith microstructure and water temperature. ICES J Mar Sci 50:261–270
TIBCO_Software_Inc. (2017) Statistica (data analysis software system), version 13, http://statistica.io
Ulltang Ø (1996) Stock assessment and biological knowledge: can prediction be reduced? ICES J Mar Sci 53:659–675
Varpe Ø (2017) Life history adaptations to seasonality. Integr Comp Biol 57:943–960. https://doi.org/10.1093/icb/icx123
Varpe Ø, Fiksen Ø (2010) Seasonal plankton-fish interactions: light regime, prey phenology, and herring foraging. Ecology 91:311–318. https://doi.org/10.1890/08-1817.1
Wuenschel MJ, McElroy WD, Oliveira K, McBride RS (2019) Measuring fish condition: an evaluation of new and old metrics for three species with contrasting life histories. Can J Fish Aquat Sci 76:886–903. https://doi.org/10.1139/cjfas-2018-0076
Yaragina NA, Marshall CT (2000) Trophic influences on interannual and seasonal variation in the liver condition index of Northeast Arctic cod (Gadus morhua). ICES J Mar Sci 57:42–55. https://doi.org/10.1006/jmsc.1999.0493
Yaragina N, Bogstad B, Kovalev Y (2009) Variability in cannibalism in Northeast Arctic cod (Gadus morhua) during the period 1947–2006. Mar Biol Res 5:75–85. https://doi.org/10.1080/17451000802512739
Zuur AF, Ieno EN (2016) A protocol for conducting and presenting results of regression-type analyses. Methods Ecol Evol 7:636–645. https://doi.org/10.1111/2041-210x.12577
Acknowledgements
The help of Frank Midtøy (UiB) and vessel crew (IMR) during the cruises, Christel Krossøy (UiB) and Julie Skadal (UiB) during laboratory analyses, and Egil Karlsbakk (UiB) for guidance during identification of nematodes, is greatly appreciated. The research was covered by the University of Bergen, and the Institute of Marine Research.
Funding
Open access funding provided by University of Bergen (incl Haukeland University Hospital).
Author information
Authors and Affiliations
Contributions
ØL: laboratory analysis, data analysis, writing, review, and editing, EE: conceptualization, data analysis, writing review, and editing, AF: conceptualization, data analysis, writing, review, and editing.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflict of interest.
Ethical approval
No ethics approval was required for this study.
Additional information
Responsible Editor: Scott Hamilton.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Langnes, Ø., Eriksen, E. & Folkvord, A. Liver condition of 0 and 1-group cod (Gadus morhua) in the Barents Sea. Mar Biol 171, 41 (2024). https://doi.org/10.1007/s00227-023-04354-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00227-023-04354-6