
Abstract
The oceanic manta ray (Mobula birostris) is a circumglobally distributed species, but little to no information exists for many parts of their range. The eastern central Atlantic Ocean is a particularly data-deficient area, where manta ray presence has seldom been reported and regional descriptions of the distribution, movement, and connectivity of populations are unavailable. This study makes use of satellite telemetry, coupled with ecological niche modeling, to provide the first description an oceanic manta ray population in the region based on observations from Cabo Verde, and describes the most likely distribution of the species in the region. Repeated surveys between 2015 and 2019 and data from 11 satellite-tracked individuals revealed a reliable and continued presence of manta rays in the archipelago, although the number of sightings was considerably lower than those reported for some of the major aggregation sites for the species and no individual was ever re-sighted. Ecological niche modeling indicates suitable habitat for manta rays exists in Cabo Verde for most of the year, though in considerably higher extent during the summer months. The model furthermore suggests possible seasonal connections between the archipelago and the wider Gulf of Guinea. The role of Cabo Verde in the life-history of oceanic manta rays in the region remains to be elucidated, though the low abundance and re-sighting frequency, coupled with modeling suggesting expanding and contracting suitable habitat, suggests that Cabo Verde might be a key stop-over along a potentially longer movement corridor.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Manta rays are a group of two planktivorous batoid ray species in the genus Mobula (White et al. 2018), with a third putative species having been identified in the western central Atlantic ocean (Hinojosa-Alvarez et al. 2016; Hosegood et al. 2020). The reef manta ray (Mobula alfredi) is considered vulnerable to extinction by the IUCN Red List (Marshall et al. 2022b) and the oceanic manta ray (Mobula birostris) is considered endangered on a global scale (Marshall et al. 2022a), since their populations have been significantly declining (Ward-Paige et al. 2013) due to direct and accidental fishing, among other factors (Croll et al. 2016; Marshall et al. 2022a, b).
The oceanic manta ray is the largest of the manta rays, and is circumglobally distributed in tropical and temperate waters (Marshall et al. 2022a). Despite a tendency for being more solitary and with a preference for oceanic environments (Armstrong et al. 2020; Marshall et al. 2022a) compared to reef manta rays, oceanic manta rays are also known to frequent coastal habitats (e.g.,Medeiros et al. 2015; Andrzejaczek et al. 2021), and in some cases display philopatry to certain aggregation sites (Dewar et al. 2008; Stewart et al. 2016). While many important seasonal or stable aggregation sites have been identified for both M. alfredi and M. birostris (Dewar et al. 2008; Luiz et al. 2009; Kitchen-Wheeler 2010; Deakos et al. 2011; Marshall et al. 2011; Graham et al. 2012; Germanov and Marshall 2014; McCauley et al. 2014; Jaine et al. 2014; Braun et al. 2015), little information is available for much of their range. In particular, virtually no information exists on the oceanic manta ray populations of the Eastern Central Atlantic Ocean (ECA). The presence of oceanic manta rays has been listed in published species indexes of Cabo Verde (Freitas et al. 2018) and the Azores (Sobral 2013), and they are sometimes mentioned in bycatch reports of tuna fisheries in the region (Zeeberg et al. 2006; Amandé et al. 2011; Torres-Irineo et al. 2014; Clavareau et al. 2018). No thorough assessment of manta ray presence exists for any country or region of the eastern Atlantic, and no key aggregation sites like the ones described for other parts of their range have been identified and formally described.
The ECA is a highly productive region for marine resources and multiple ecosystems, which sustain high biodiversity (Roberts et al. 2002) as well as productive fisheries (FAO 2018). It is therefore plausible that substantial manta ray populations inhabit this region. Simultaneously, the existence of numerous, highly active industrial and artisanal fishing operations (FAO 2018) is a potential threat to these populations. Manta rays are susceptible to capture in multiple fishery types and fishing gears, but especially in purse seine operations (Croll et al. 2016). In the Atlantic, reports exist on manta rays being accidentally caught in tuna fisheries (Zeeberg et al. 2006; Amandé et al. 2011; Torres-Irineo et al. 2014), though the ECA is one of the regions with the highest proportion of illegal, unregulated, and unreported (IUU) catches (Agnew et al. 2009; Carneiro 2012), suggesting that bycatch frequency might be much higher than official data suggests. Manta rays have extremely conservative life histories with very low fecundity (Dulvy et al. 2014), and are therefore sensitive to even modest increases in mortality (Croll et al. 2016).
In Cabo Verde, sightings of the oceanic manta ray (hereafter referred to only as manta ray. M. alfredi will be referred to as reef manta ray) are common among traditional fisher folks and tourism guides, notably in the Eastern islands of Sal, Boavista, and Maio, though no information exists on this population in the scientific literature. Fishermen on the island of Boavista report that manta rays are present around the island year-round, though with potential peaks in abundance in March–April and August–October. Manta rays are also advertised as potential encounters during SCUBA trips by dive shops on the island of Sal, and occasional sightings of the species have also been reported in other islands, including Maio, Santiago, and Sao Vicente (FG, personal observations). Although the presence of manta rays in the archipelago is therefore known, it remains unclear whether they are long-term residents or if Cabo Verde represents a stop in seasonal movements throughout the Eastern Atlantic Ocean.
A better understanding of the manta ray population in the western Atlantic and its distribution in this region, as well as its interactions with fishing activities, is required to inform regional conservation planning and evaluate the status of manta rays in this part of their range. Here, we present the first description of manta ray presence in the ECA, employ satellite tracking to describe movements of individuals within the country’s territorial waters, and use this information to infer, through ensemble ecological niche modeling, the likely wider distribution of manta rays in Cabo Verde and the adjacent region.
Methods
Visual surveys
Over the course of 4 years (2016–2019), 48 surveys aimed at encountering and identifying megaplanktivores were conducted around the island of Boavista, Cabo Verde (Fig. 1a). Surveys were mostly (80%) conducted during the summer and autumn months (July–October; when sea conditions are most favorable to fieldwork) and were focused on the south-western quadrant of the island, where some of the most productive fishing sites of the island are located and fishermen often reported sightings of manta rays. Several surveys (n = 12, 27%) were also conducted at other times of the year and/or in other parts of the island, when conditions at sea allowed. Two surveys were also opportunistically conducted in the island of Sal, north of Boavista. Surveys were conducted aboard traditional fishing vessels (6 m-long, open wooden boats), with two to three observers continually scanning the water for sightings. Occasional sightings were also collected opportunistically whenever manta rays were encountered during the course of surveys aimed at other species. Upon sighting of a manta ray, two trained observers would enter the water and obtain pictures of the ventral side of the animals for later photo-identification (Marshall et al. 2011), as well as determine the sex of individuals and estimate their size (size estimation was carried out visually by comparing the animals to an observer of known length or to a 2 m pole spear). Whenever size could be estimated, sexual maturity was then inferred by comparison with published size-at-maturity estimates for the species (Stewart et al. 2018).

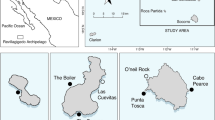
a Location of Cabo Verde within the Eastern Central Atlantic; b Example photo of oceanic manta ray (M. birostris) encountered in Cabo Verde during surveys; c Oceanic manta ray (M. birostris) sightings (yellow dots) and space use (red squares) in Cabo Verde, measured as the total length of track segments intersecting each cell, multiplied by the number of individuals that have been tracked within the cell. Darker shading indicates higher intensity of use of the cell. Only cells used by more than one individual are shown
Finally, feeding behavior was noted whenever it could be observed. Feeding was indicated in this study by manta ray swimming with open mouth and unfurled cephalic fins. Comparison of photo-identification samples was conducted visually by two trained researchers.
Satellite tracking
A total of 11 manta rays were fitted with towed satellite transmitting position-only tags (Wildlife Computers SPOT5 and SPOT6; www.wildlifecomputers.com), as per methods described in (Graham et al. 2012). Tags were attached while swimming behind and above the animal, using a small percutaneous nylon umbrella dart attached to a 1 m long 1/160 (1.59 mm) nylon cable, inserted into the lower left or lower right quadrant shoulder musculature using a 2 m pole spear. The tags were covered with dark blue antifouling paint to minimize bio-fouling. The tags were programmed to transmit continuously at the sea surface and to record the time each animal spent at pre-established temperature bins within 6-h periods. The tag’s position was determined by the Argos System (www.argos-system.org). Detachment of each tag from the manta ray was estimated based on clear changes in traveling speed, and the number, quality, and time of daily transmissions ((Graham et al. 2012; Hearn et al. 2013); Supplementary Information Appendix 1). All locations received after the estimated detachment time were removed. The remaining Argos data were filtered to only include location classes (LC) 1, 2, 3, A, and B, for which location accuracy is known (Witt et al. 2010). Locations estimated to be on land, or to have required speeds greater than 20 km/h between locations (Graham et al. 2012) were removed.
A hierarchical state-space model (SSM) was applied to the filtered Argos data to regularize positions and improve data retention. The SSM was computed through the R package bsam (Jonsen et al. 2005; Jonsen 2016), which estimated locations at 12-h intervals, as the majority (78%) of locations were 12 h apart or less (Lea et al. 2015). A total of 15,000 iterations were run, of which 5,000 assigned to adaptation and burn-in respectively, and were thinned by a factor of 5. The Argos data had some large temporal gaps in transmissions, so periods with no “real” transmissions for 1 week were eliminated, to avoid spurious interpolation.
Since most tagged manta rays undertook long movements between islands, description of a home range through kernel density estimation was deemed inappropriate. Instead, a grid covering the study area with resolution 0.1° was created within the EEZ of Cabo Verde; for each cell, the number of individual manta ray tracks that crossed it was counted, and the length of each segment was measured. Space use of manta rays was then defined as all the cells containing manta ray tracks, while the relative importance of different cells was inferred by the sum of all track segments length within the cell, weighted by the number of individuals present in the cell. The length of tracks was used, instead of the number of locations received within any cells, because it was deemed better suited at capturing manta ray space use, given that most of the time manta rays are found underwater and hence most of their positions are not reflected in satellite transmissions.
Distribution modeling
Species distribution modeling was used to investigate the broader habitat distribution for manta rays in Cabo Verde, as well as potential seasonal shifts in their distribution. The spatial domain for modeling was fixed to be within latitudes 10° and 22° and from the coast of continental Africa to longitude − 33°. An ensemble approach was used (Garzon et al. 2020), which included general linear models (GLM), general addictive models (GAM), boosted regression models (GBM), and Maximum entropy models (MaxEnt) (Araújo and New 2007). In the absence of real absence records (i.e., areas where manta rays have never been recorded despite continuous monitoring), models were trained using data derived from the state-space models as ‘presence’, and an equal number of randomly placed pseudo-absences (Wisz and Guisan 2009; Barbet-Massin et al. 2012; Garzon et al. 2020). To minimize stochastic effects, five pseudo-absence sets were created and used in turn for modeling with the same presence set (See Supplementary Information Appendix 2 for details of the modeling process). Pseudo-absence sets contained an equal number of points as the presence set, each of which was placed randomly within the modeling spatial domain. In order to avoid overfitting of the models, both presence and pseudo-absence points were clustered using K-means clustering with a radius of 4 km to match the resolution of environmental data.
Environmental data to inform models included static and dynamic variables that have been found to relate to presence or increased abundance of manta rays and other mega-planktivores (Panigada et al. 2008; Embling et al. 2010; Freedman and Sen Roy 2012; Jaine et al. 2012; McKinney et al. 2012; Sequeira et al. 2012; Hacohen-Domené et al. 2017; Garzon et al. 2020; Farmer et al. 2022), including bathymetric depth (General Bathymetric Chart of The Ocean; http://www.gebco.net), seafloor slope (created from bathymetry data in QGIS), night-time sea surface temperature (NSST, https://www.oceancolor.gsfc.nasa.gov), and chlorophyll-a concentration (Chl-a, used as a proxy for plankton abundance; https://www.oceancolor.gsfc.nasa.gov). Dynamic variables were sourced as monthly averages (mean) for all years in which manta rays were tracked (2016–2019). All environmental variables were downloaded as spatially referenced gridded layers (from now on referred to as rasters) and re-sampled to match the spatial resolution of the coarsest dataset (Chl-a and NSST, 4 × 4km2) using linear interpolation. Environmental conditions at each presence and pseudo-absence point were extracted from temporally matching rasters (according to the month of the location fix). Correlation between environmental variables was tested using Spearman rank correlation, and only non-correlated variables (r < 0.5%) were kept for modeling.
Ecological niche modeling was carried out using the biomod2 package for the software R (R Core Team 2022); For each pseudo-absence dataset, 10 runs were computed for each model (GLM, GAM, GBM, and MaxEnt) using a 75–25% random split of the data for training and testing, respectively. True Skill Statistic (TSS) scores were computed to evaluate the predictive ability of the models (Allouche et al. 2006). Only models with a TSS ≥ 0.7 were used to compute the ensemble model, which was created as their weighted average (weights assigned according to TSS scores).
Predictions of habitat suitability were created for each month of the year in the form of rasters that have the same spatial resolution of the environmental variables used for training, where each cell was given a value ranging from 0 to 1 indicating its relative suitability (RS) for manta rays (0 indicates lowest suitability, 0.5 corresponds to average suitability, and 1 indicates highest suitability). Monthly rasters were turned into binary layers where areas having RS ≥ 0.5 (i.e., areas that contained suitable habitat) were assigned the value of 1. These layers were then summed, so that the resulting raster cells contained values ranging from 0 to 12, corresponding to the number of months of the year in which they contained suitable habitat for manta rays.
Results
Surveys produced 51 manta rays sightings (n = 7 in 2015, n = 29 in 2016, n = 3 in 2017, n = 6 in 2018, and n = 6 in 2019). Mantas were mostly female (n = 29, though sex could not be determined for 14 individuals) and had an average disk width of 3.6 ± 0.6 m (range 2.8–5.0 m). Of all sightings for which disk width could be measured (n = 39), 49% were deemed to be of likely reproductive size. Of all mantas encountered, a ventral photo could be obtained for 24 individuals. Visual inspection of these photographs did not reveal any re-encounter instances. All but two sightings were recorded in the southwest of the island of Boavista (where most tagging events took place, Fig. 1), and the remaining were sighted between the islands of Sal and Boavista. Manta rays were observed feeding on only 5 occasions throughout the study.
All manta rays fitted with satellite tags (Table 1) were encountered and tagged around the island of Boavista. Tagging took place between August and October each year. Satellite tags transmitted positions for an average of 57 days (range 13–99, Table 1) before detaching from the animal or interrupting transmissions, with high variability in the number of transmissions received among individuals (range 27–199, after filtering and regularization). All data were received between August and January, while no data was received between February and July of any year.
Most manta rays spent all or some time around the island of Boavista following tagging (Fig. 1), with the longest remaining around Boavista for 99 days (PTT 172,735, Fig. 1). Manta rays only moved either slightly further offshore (PTT 163,667, 163,668, and 172,735) before returning to Boavista, or to the southern islands of Maio and Santiago (PTT 163,669, 172,734, 172,736, and 172,745). Two individuals (PTT 163,667 and 172,736) briefly visited the island of Sal, but no locations were received from the northern islands of the archipelago, despite manta ray sightings having been reported there in the past. All tagged manta rays remained within the Cabo Verdean Exclusive Economic Zone (EEZ) for the duration of the tracking (Fig. 1), with the exception of manta ray 163,667, whose last transmitted positions placed it just outside the northwest boundary of the EEZ (Fig. 1). Space use of manta rays was concentrated around the island of Boavista, especially in the southeast quadrant of the island (Fig. 2). The shallow corridor linking the islands of Boavista and Maio was also used by 5 individuals, while no other clear movement corridors could be identified.

Filtered and regularized tracks of oceanic manta rays (M. birostris) tagged in Cabo Verde between 2016 and 2019. Numbers in the top left of each panel indicate the identification number of each tracker. Color of locations indicates time elapsed from tag deployment, with darker shades corresponding to most recent locations. Bottom-right panel shows estimated tracks for all individuals in relation to Cabo Verde’s Exclusive Economic Zone
All 200 ecological niche models (10 runs for 4 algorithms, repeated for 5 pseudo-absence datasets) ran without errors. Since bathymetry was found to be significantly and strongly correlated (r > 0.5) with chlorophyll concentration, it was excluded from modeling. Models had generally high TSS scores (mean 0.81 ± 0.06 s.d.). The ensemble model was built using 189 models that had TSS ≥ 0.7. The Ensemble Ecological Niche Model (EENM) indicated that chlorophyll-a concentration was the most important variable in predicting habitat suitability for manta rays (relative importance, RI = 0.44), with NSST receiving only slightly less importance (RI = 0.42); bathymetric slope was deemed to have the lowest importance in determining habitat suitability (RI = 0.14).
The monthly predictions of the model suggest that manta ray presence in the archipelago undergoes considerable seasonal fluctuations, although some suitable habitat for manta rays is present in the EEZ year-round (Fig. 3). The peak of predicted suitable habitat extent and relative suitability was obtained in November, with a significant reduction of habitat extent and relative suitability between February and June. The monthly shift in suitable habitat somewhat follows a northward movement from southern regions of west Africa, passing through Cabo Verde from July to November/December, and then moving offshore—perhaps toward the Western Atlantic. Cabo Verde nonetheless retains suitable habitat for manta rays for at least six months of the year around all of the islands and the two prominent seamounts to the east of the archipelago (in waters shallower than 2000 m, Fig. 4). The only other region that hosts suitable manta ray habitat for over half the year is found in the southeast corner of the study area, in Guinea-Bissau’s EEZ and nearby international waters (Fig. 4).

Monthly predictions of habitat suitability for oceanic manta rays (M. birostris) in the study area (months are indicated in the top-left corner of each panel). Shading indicates relative suitability from 0 (unsuitable, dark blue) to 1 (extremely suitable, yellow). White cells represent areas where environmental data was absent and suitability could not be calculated. Dotted white lines delimit exclusive economic zones (EEZ) of the countries in the study area

Gray shading indicates the number of months of the year in which areas host suitable manta ray habitat. The orange line defines key areas where suitable habitat is encountered for at least half of the year. Black lines delimit exclusive economic zones (EEZ) of the countries in the study area
Discussion
The data presented here suggest a reliable presence of manta rays between July and January in the archipelago of Cabo Verde across at least six years. This is the only documented region in the eastern Central Atlantic where manta rays appear to be regularly encountered, although it is likely that further research in this region will uncover other important locations for the species. Despite the repeated occurrence of manta rays in Cabo Verde, this site is very different to other “aggregation” sites, since manta rays are never sighted in groups bigger than two individuals, their movements are distributed over a relatively large area, and there are no confirmed instances of re-sighting of individuals which would suggest seasonal fidelity to the site. The lack of re-encounters suggests that Cabo Verde could represent a stop along a larger movement route. While the low number of individuals encountered in any year makes it impossible to assess the long-term residency of manta rays from sightings alone, satellite tracking data also suggest that most individuals left the island of Boavista to move to other islands or, in some cases, toward the central Atlantic Ocean, hence supporting the possibility of relatively short residency time within national waters. However, given the relatively low survey effort in relation to manta ray density at this site, it is possible that likelihood of re-sighting was underestimated.
While satellite tracking could elucidate the movements of individuals within Cabo Verdean national waters, it failed to provide long-term insights into the movement of these animals—the longest tag retention time was 99 days. Only one manta was observed leaving Cabo Verde’s EEZ in this time, but it appears unlikely that the manta rays tracked in the present study are resident to Cabo Verde year-round. The comparison of identifying features of manta rays in Cabo Verde with those held in other repositories could also further elucidate the long-distance movements of these animals within the Atlantic Ocean.
While it seems that the island of Boavista is a particularly important site for manta rays in Cabo Verde, with all tagged individuals spending time around the island before moving to other islands, it wasn’t possible to determine the reason for their stay in this study. The majority of manta rays sighted during the present study had reached a size where maturity was plausible (Table 1, Stewart et al. 2018); yet, no courtship behavior was observed during surveys and individuals were rarely observed feeding, instead usually cruising with rolled-up cephalic fins. The ensemble distribution model suggests a seasonal cycle in manta ray occurrence, and little suitable habitat with overall lower predicted relative suitability scores present from February to June in the country. In the pattern, the model describes concords with reports by fishermen that place the annual peak in manta ray abundance between July and October. Sightings of the species are also reported in the winter and spring months (ZS, ZLM, and FG personal observations), when the model largely predicts absence of manta rays, although these records occur irregularly and less frequently than in summer months. It is possible that the lack of data points in the first half of the year in the presence database used to train the model, has led the model to underestimate the breadth of the manta rays ecological niche in Cabo Verde, suggesting that environmental conditions are unsuitable for manta rays when they might in fact be able to tolerate a wider spectrum of temperatures and chlorophyll concentrations. Additional data, obtained either from a more rigorous monthly monitoring system through aerial or in-water surveys, or better telemetry attachment systems that secure longer tag retention times, are required to confirm or refute the seasonality of manta ray presence in Cabo Verde.
At a national level, threats to the survival of manta rays are likely low within Cabo Verde. No directed fishery exists for this species in the country, and given the concentration of suitable habitat nearer the islands, manta rays are most likely to interact with small-scale or semi-industrial fishing vessels, which may be less likely to by-catch, although occasional catches of mobulid species have been observed in the country (FG personal observation). However, large industrial vessels, mainly coming from Europe, Japan, and China, are active within and outside Cabo Verde’s EEZ and could also represent a threat to the species. Bycatch of mobulid rays has been reported in the ECA (Zeeberg et al. 2006; Amandé et al. 2011; Torres-Irineo et al. 2014), and though the frequency and overall magnitude of the phenomenon are not clear, mobulid mortality following release from purse seine nets can exceed 50% (Francis and Jones 2017).
This study elucidates the role of coastal and pelagic waters up to 2000 m deep in Cabo Verde in providing favorable conditions and habitat for manta ray habitat for at least half of the year. Similarly, pockets of long-lasting suitable habitat seem to be present further south, off the coast of Guinea-Bissau, suggesting that perhaps a connection could exist between Cabo Verde and the plateau and complex bathymetric region fronting Guinea and Guinea-Bissau. The existence and nature of this connection should be further elucidated, as it could have important consequences for conservation planning for this species in the region. Moving forward, further research to better elucidate the spatial ecology of manta rays in this region of the world is needed. Within Cabo Verde, the cooperation of the fishing community could be enlisted to record sightings of manta rays across the country to obtain a better picture of manta ray space use and residency. Indeed, successful cases of cooperation between conservation organizations and fishermen communities already exists in many islands of the archipelago, highlighting the opportunity for expansion of the project to other islands and to include new species. The diving community, albeit comparatively smaller in Cabo Verde than other known manta ray aggregations sites, may also offer a source of sighting and photo identification data.
Identifying links between Cabo Verde and other countries, as well as the role that the country plays in the life cycle of manta rays in the region also remains a priority. Strengthening the collection and sharing of individual identifying photos could provide a mean to establish links between regions, while improved satellite telemetry systems could not only reveal movement corridors but also further elucidate the importance of different areas through identification of behaviors.
Data availability
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
Agnew DJ, Pearce J, Pramod G, Peatman T, Watson R, Beddington JR, Pitcher TJ (2009) Estimating the worldwide extent of illegal fishing. PLoS ONE 4:e4570. https://doi.org/10.1371/journal.pone.0004570
Allouche O, Tsoar A, Kadmon R (2006) Assessing the accuracy of species distribution models: prevalence, kappa and the true skill statistic (TSS). J Appl Ecol 43:1223–1232. https://doi.org/10.1111/j.1365-2664.2006.01214.x
Amandé M, Ariz J, Chassot E, De MAD, Gaertner D, Murua H, Pianet R, Ruiz J, Chavance P, Amandé M, Ariz J, Chassot E, De MAD, Gaertner D, Amandè MJ, Ariz J, Chassot E, De MAD, Gaertner D (2011) Bycatch of the European purse seine tuna fishery in the Atlantic Ocean for the 2003–2007 period. Aquat Living Resour. https://doi.org/10.1051/alr/2011003
Andrzejaczek S, Schallert RJ, Forsberg K, Arnoldi NS, Cabanillas-Torpoco M, Purizaca W, Block BA (2021) Reverse diel vertical movements of oceanic manta rays off the northern coast of Peru and implications for conservation. Ecol Solut Evid 2:e12051. https://doi.org/10.1002/2688-8319.12051
Araújo MB, New M (2007) Ensemble forecasting of species distributions. Trends Ecol Evol 22:42–47. https://doi.org/10.1016/j.tree.2006.09.010
Armstrong AJ, Armstrong AO, Bennett MB, McGregor F, Abrantes KG, Barnett A, Richardson AJ, Townsend KA, Dudgeon CL (2020) The geographic distribution of reef and oceanic manta rays (Mobula alfredi and Mobula birostris) in Australian coastal waters. J Fish Biol 96:835–840. https://doi.org/10.1111/jfb.14256
Barbet-Massin M, Jiguet F, Albert CH, Thuiller W (2012) Selecting pseudo-absences for species distribution models: how, where and how many? Methods Ecol Evol 3:327–338. https://doi.org/10.1111/j.2041-210X.2011.00172.x
Braun CD, Skomal GB, Thorrold SR, Berumen ML (2015) Movements of the reef manta ray (Manta alfredi) in the Red Sea using satellite and acoustic telemetry. Mar Biol 162:2351–2362. https://doi.org/10.1007/s00227-015-2760-3
Carneiro G (2012) “They Come, They Fish, and They Go”: EC Fisheries Agreements with Cape Verde and São Tomé e Príncipe. Mar Fish Rev 73:1–25
Clavareau L, Sabarros PS, Escalle L, Bach P (2018) Elasmobranches bycatch in the french tropical purse-seine fishery of the eastern Atlantic Ocean: spatio-temporal distributions, life stages, sex-ratio and mortality rates. ICCAT Collective Vol Sci Pap 74:3740–3753
Croll DA, Dewar H, Dulvy NK, Fernando D, Francis MP, Galván-Magaña F, Hall M, Heinrichs S, Marshall A, Mccauley D, Newton KM, Notarbartolo-Di-Sciara G, O’Malley M, O’Sullivan J, Poortvliet M, Roman M, Stevens G, Tershy BR, White WT (2016) Vulnerabilities and fisheries impacts: the uncertain future of manta and devil rays. Aquat Conserv Mar Freshw Ecosyst 26:562–575. https://doi.org/10.1002/aqc.2591
Deakos MH, Baker JD, Bejder L (2011) Characteristics of a manta ray Manta alfredi population off Maui, Hawaii, and implications for management. Mar Ecol Prog Ser 429:245–260. https://doi.org/10.3354/meps09085
Dewar H, Mous P, Domeier M, Muljadi A, Pet J, Whitty J (2008) Movements and site fidelity of the giant manta ray, Manta birostris, in the Komodo Marine Park, Indonesia. Mar Biol 155:121–133. https://doi.org/10.1007/s00227-008-0988-x
Dulvy NK, Pardo SA, Simpfendorfer CA, Carlson JK (2014) Diagnosing the dangerous demography of manta rays using life history theory. PeerJ 2014:1–19. https://doi.org/10.7717/peerj.400
Embling CB, Gillibrand PA, Gordon J, Shrimpton J, Stevick PT, Hammond PS (2010) Using habitat models to identify suitable sites for marine protected areas for harbour porpoises (Phocoena phocoena). Biol Cons 143:267–279. https://doi.org/10.1016/j.biocon.2009.09.005
FAO (2018) The state of the world fisheries and aquaculture 2018—Meeting the sustainable development goals. Rome
Farmer NA, Garrison LP, Horn C, Miller M, Gowan T, Kenney RD, Vukovich M, Willmott JR, Pate J, Harry Webb D, Mullican TJ, Stewart JD, Bassos-Hull K, Jones C, Adams D, Pelletier NA, Waldron J, Kajiura S (2022) The distribution of manta rays in the western North Atlantic Ocean off the eastern United States. Sci Rep 12:6544. https://doi.org/10.1038/s41598-022-10482-8
Francis MP, Jones EG (2017) Movement, depth distribution and survival of spinetail devilrays (Mobula japanica) tagged and released from purse-seine catches in New Zealand. Aquat Conserv Mar Freshw Ecosyst 27:219–236. https://doi.org/10.1002/aqc.2641
Freedman R, Sen Roy S (2012) Spatial patterning of Manta birostris in United States east coast offshore habitat. Appl Geogr 32:652–659. https://doi.org/10.1016/j.apgeog.2011.07.015
Freitas R, Burnett KA, Dureuil M, Caruso JH (2018) New and confirmed records of fishes from the Cabo Verde archipelago based on photographic and genetic data. Life Mar Sci 35:67–83
Garzon F, Graham RT, Witt MJ, Hawkes LA (2020) Ecological niche modeling reveals manta ray distribution and conservation priority areas in the Western Central Atlantic. Anim Conserv. https://doi.org/10.1111/acv.12663
Germanov ES, Marshall AD (2014) Running the Gauntlet: Regional movement patterns of Manta alfredi through a complex of parks and fisheries. PLoS ONE 9:1–9. https://doi.org/10.1371/journal.pone.0110071
Graham RT, Witt MJ, Castellanos DW, Remolina F, Maxwell S, Godley BJ, Hawkes LA (2012) Satellite tracking of manta rays highlights challenges to their conservation. PLoS ONE 7:3–8. https://doi.org/10.1371/journal.pone.0036834
Hacohen-Domené A, Martínez-Rincón RO, Galván-Magaña F, Cárdenas-Palomo N, Herrera-Silveira J (2017) Environmental factors influencing aggregation of manta rays (Manta birostris) off the northeastern coast of the Yucatan Peninsula. Mar Ecol 38:1–9. https://doi.org/10.1111/maec.12432
Hearn AR, Green JR, Espinoza E, Peñaherrera C, Acuña D, Klimley AP (2013) Simple criteria to determine detachment point of towed satellite tags provide first evidence of return migrations of whale sharks (Rhincodon typus) at the Galapagos Islands, Ecuador. Anim Biotelemetry 1:1–10. https://doi.org/10.1186/2050-3385-1-11
Hinojosa-Alvarez S, Walter RP, Diaz-Jaimes P, Galván-Magaña F, Paig-Tran EM (2016) A potential third Manta Ray species near the Yucatán Peninsula? Evidence for a recently diverged and novel genetic Manta group from the Gulf of Mexico. PeerJ 4:e2586. https://doi.org/10.7717/peerj.2586
Hosegood J, Humble E, Ogden R, de Bruyn M, Creer S, Stevens GMW, Abudaya M, Bassos-Hull K, Bonfil R, Fernando D, Foote AD, Hipperson H, Jabado RW, Kaden J, Moazzam M, Peel LR, Pollett S, Ponzo A, Poortvliet M, Salah J, Senn H, Stewart JD, Wintner S, Carvalho G (2020) Phylogenomics and species delimitation for effective conservation of manta and devil rays. Mol Ecol 29:4783–4796. https://doi.org/10.1111/mec.15683
Jaine FRA, Couturier LIE, Weeks SJ, Townsend KA, Bennett MB, Fiora K, Richardson AJ (2012) When giants turn up: sighting trends, environmental influences and habitat use of the manta ray Manta alfredi at a coral reef. PLoS ONE 7:e46170. https://doi.org/10.1371/journal.pone.0046170
Jaine FRA, Rohner C, Weeks S, Couturier L, Bennett M, Townsend K, Richardson A (2014) Movements and habitat use of reef manta rays off eastern Australia: offshore excursions, deep diving and eddy affinity revealed by satellite telemetry. Mar Ecol Prog Ser 510:73–86. https://doi.org/10.3354/meps10910
Jonsen ID (2016) Joint estimation over multiple individuals improves behavioural state inference from animal movement data. Sci Rep 6:1–9. https://doi.org/10.1038/srep20625
Jonsen ID, Flemming JM, Myers RA (2005) Robust state-space modeling of animal movement data. Ecology 86:2874–2880. https://doi.org/10.1890/04-1852
Kitchen-Wheeler AM (2010) Visual identification of individual manta ray (Manta alfredi) in the Maldives Islands, Western Indian Ocean. Mar Biol Res 6:351–363. https://doi.org/10.1080/17451000903233763
Lea JSE, Wetherbee BM, Queiroz N, Burnie N, Aming C, Sousa LL, Mucientes GR, Humphries NE, Harvey GM, Sims DW, Shivji MS (2015) Repeated, long-distance migrations by a philopatric predator targeting highly contrasting ecosystems. Sci Rep 5:1–11. https://doi.org/10.1038/srep11202
Luiz OJ, Balboni AP, Kodja G, Andrade M, Marum H (2009) Seasonal occurrences of Manta birostris (Chondrichthyes: Mobulidae) in southeastern Brazil. Ichthyol Res 56:96–99. https://doi.org/10.1007/s10228-008-0060-3
Marshall AD, Dudgeon CL, Bennett MB (2011) Size and structure of a photographically identified population of manta rays Manta alfredi in southern Mozambique. Mar Biol 158:1111–1124. https://doi.org/10.1007/s00227-011-1634-6
Marshall A, Barreto R, Carlson J, Fernando D, Fordham S, Francis MP, Herman K, Jabado RW, Liu KM, Rigby CL, Romanov E (2022a) Mobula birostris (amended version of 2020 assessment). The IUCN Red List of Threatened Species 2022a:e.T198921A214397182. doi: https://doi.org/10.2305/IUCN.UK.2022-1.RLTS.T198921A214397182.en
Marshall AD, Barreto R, Carlson J, Fernando D, Fordham S, Francis M, Herman K, Jabado RW, Liu KM, Pacoureau N, Rigby CL, Romanov E, Sherley RB (2022b) Mobula alfredi (amended version of 2019 assessment). IUCN Red List of Threatened Species 2022b:e.T195459A214395983. doi: https://doi.org/10.2305/IUCN.UK.2022-1.RLTS.T195459A214395983.en
McCauley DJ, DeSalles PA, Young HS, Papastamatiou YP, Caselle JE, Deakos MH, Gardner JPA, Garton DW, Collen JD, Micheli F (2014) Reliance of mobile species on sensitive habitats: A case study of manta rays (Manta alfredi) and lagoons. Mar Biol 161:1987–1998. https://doi.org/10.1007/s00227-014-2478-7
McKinney JA, Hoffmayer ER, Wu W, Fulford R, Hendon JM (2012) Feeding habitat of the whale shark Rhincodon typus in the northern Gulf of Mexico determined using species distribution modelling. Mar Ecol Prog Ser 458:199–211. https://doi.org/10.3354/meps09777
Medeiros AM, Luiz OJ, Domit C (2015) Occurrence and use of an estuarine habitat by giant manta ray Manta birostris. J Fish Biol 86:1830–1838. https://doi.org/10.1111/jfb.12667
Panigada S, Zanardelli M, MacKenzie M, Donovan C, Mélin F, Hammond PS (2008) Modelling habitat preferences for fin whales and striped dolphins in the Pelagos Sanctuary (Western Mediterranean Sea) with physiographic and remote sensing variables. Remote Sens Environ 112:3400–3412. https://doi.org/10.1016/j.rse.2007.11.017
R Core Team (2022) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Roberts CM, McClean CJ, Veron JEN, Hawkins JP, Allen GR, McAllister DE, Mittermeier CG, Schueler FW, Spalding M, Wells F, Vynne C, Werner TB (2002) Marine biodiversity hotspots and conservation priorities for tropical reefs. Science 295:1280–1284. https://doi.org/10.1126/science.1067728
Sequeira A, Mellin C, Rowat D, Meekan MG, Bradshaw CJA (2012) Ocean-scale prediction of whale shark distribution. Divers Distrib 18:504–518. https://doi.org/10.1111/j.1472-4642.2011.00853.x
Sobral AFL (2013) Biology, ecology and conservation of Mobulid rays in the Azores. Univeristy of the Azores
Stewart JD, Beale CS, Fernando D, Sianipar AB, Burton RS, Semmens BX, Aburto-Oropeza O (2016) Spatial ecology and conservation of Manta birostris in the Indo-Pacific. Biol Cons 200:178–183. https://doi.org/10.1016/j.biocon.2016.05.016
Stewart JD, Jaine FRA, Armstrong AJ, Armstrong AO, Bennett MB, Burgess KB, Couturier LIE, Croll DA, Cronin MR, Deakos MH, Dudgeon CL, Fernando D, Froman N, Germanov ES, Hall MA, Hinojosa-Alvarez S, Hosegood JE, Kashiwagi T, Laglbauer BJL, Lezama-Ochoa N, Marshall AD, McGregor F, Notarbartolo di Sciara G, Palacios MD, Peel LR, Richardson AJ, Rubin RD, Townsend KA, Venables SK, Stevens GMW (2018) Research priorities to support effective manta and devil ray conservation. Front Mar Sci 5:314. https://doi.org/10.3389/fmars.2018.00314
Torres-Irineo E, Amandè MJ, Gaertner D, de Molina AD, Murua H, Chavance P, Ariz J, Ruiz J, Lezama-Ochoa N (2014) Bycatch species composition over time by tuna purse-seine fishery in the eastern tropical Atlantic Ocean. Biodivers Conserv 23:1157–1173. https://doi.org/10.1007/s10531-014-0655-0
Ward-Paige CA, Davis B, Worm B (2013) Global population trends and human use patterns of manta and mobula rays. PLoS ONE 8:e74835. https://doi.org/10.1371/journal.pone.0074835
White WT, Corrigan S, Yang L, Henderson AC, Bazinet AL, Swofford DL, Naylor GJP (2018) Phylogeny of the manta and devilrays (Chondrichthyes: Mobulidae), with an updated taxonomic arrangement for the family. Zool J Linn Soc 182:50–75. https://doi.org/10.1093/zoolinnean/zlx018
Wisz MS, Guisan A (2009) Do pseudo-absence selection strategies influence species distribution models and their predictions? An information-theoretic approach based on simulated data. BMC Ecol 9:1–13. https://doi.org/10.1186/1472-6785-9-8
Witt MJ, Åkesson S, Broderick AC, Coyne MS, Ellick J, Formia A, Hays GC, Luschi P, Stroud S, Godley BJ (2010) Assessing accuracy and utility of satellite-tracking data using Argos-linked Fastloc-GPS. Anim Behav 80:571–581. https://doi.org/10.1016/j.anbehav.2010.05.022
Zeeberg JJ, Corten A, de Graaf E (2006) Bycatch and release of pelagic megafauna in industrial trawler fisheries off Northwest Africa. Fish Res 78:186–195. https://doi.org/10.1016/j.fishres.2006.01.012
Acknowledgements
The authors would like to thank João Lima, Angelito and Carlos Monteiro, and Tchoca Moreno, as well as the fishing community of Sal Rei, for their invaluable contribution to the success of the study and continued efforts in promoting shark conservation in the country. The authors would also like to thank the Norvig Family, Wildlife Conservation Network, Foundation Ensemble, and National Geographic Society for providing financial support to the study.
Funding
This work was supported by funding from the Norvig Family, Wildlife Conservation Network, Foundation Ensemble, and National Geographic Society (grant number EC-50499C-18).
Author information
Authors and Affiliations
Contributions
RG, ZS, and FG contributed to the study conception and design. All authors participated in data collection. Data analysis was performed by FG. The first draft of the manuscript was written by FG and RG. All authors approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no relevant financial or non-financial interests to disclose.
Ethical approval
The study was conducted under research permits granted by the Cabo Verdean government. No specific ethical approval was necessary for the study, but field operations were conducted according to best-practice methodology in the sector.
Additional information
Responsible Editor: J. Carlson.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Deceased: Z.T.A. Seymour, Z.L. Monteiro.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Garzon, F., Seymour, Z.T.A., Monteiro, Z.L. et al. Spatial ecology of a newly described oceanic manta ray population in the Atlantic Ocean. Mar Biol 170, 68 (2023). https://doi.org/10.1007/s00227-023-04219-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00227-023-04219-y