
Abstract
Costa Rica hosts two renowned olive ridley (Lepidochelys olivacea) arribada beaches: Nancite and Ostional. In addition, the country hosts a third nascent arribada beach in Corozalito. We documented the frequency of arribada events in Corozalito (9°50′55.40″ N; 85°22′ 47.67″ W) from 2008 to 2021 and the abundance of egg-laying females during 12 arribadas. Moreover, we report on tag recaptures at Corozalito that were initially deployed at other nesting beaches in Costa Rica. Finally, we estimated incubation temperature, and performed quadrat and nest excavations to estimate hatching rates. We registered 29 arribadas at Corozalito since 2008, all between August and January. We estimated more than 150,000 olive ridleys nested during 12 arribadas from 2019 to 2021. Mean hatching success was 65% in quadrats and 59% for in situ marked nests. The mean incubation temperature was 32.3 °C. We found turtles tagged at other locations on the Pacific coast of Costa Rica nesting at Corozalito, providing evidence of philopatric plasticity. We suggest nascent arribada rookeries form rapidly via recruitment from regional turtle aggregations. Our findings show that hatching success is far higher at Corozalito than Ostional or Nancite. Continued monitoring of nesting dynamics at this site may shed light upon the mechanisms that rule how arribada beaches are born and evolve. Corozalito offers a unique opportunity to study and understand the arribada phenomenon and warrants official protection under official National Wildlife Refuge status.
Similar content being viewed by others

Introduction
The olive ridley (Lepidochelys olivacea, Eschscholtz, 1829) is the most abundant sea turtle species in the world (Pritchard 1997). This species is currently listed as ‘vulnerable’, with populations globally declining according to the International Union for the Conservation of Nature’s Red List of Threatened Species (Abreu-Grobois and Plotkin 2008). Besides nesting in solitary fashion, the olive ridley is also known to nest synchronously en masse, a phenomenon termed “arribada”, for the Spanish word meaning arrival (Hughes and Richard 1974). Arribadas are exclusive to the genus Lepidochelys (Pritchard 1997). Solitary nesting behavior is shared with the other six sea turtle species in which the turtle emerges, nests individually, and returns to the sea (Bernardo and Plotkin 2007). However, the arribada is an unusual nesting behavior in which hundreds to thousands of female ridley turtles emerge synchronously at a specific beach to nest over a few days (Hughes and Richard 1974; Cornelius et al. 1991, 2007; Valverde et al. 1998; Bernardo and Plotkin 2007). To distinguish arribada nesting from a night of heavy solitary nesting at major arribada beaches, a threshold number of 100 turtles or more present on the beach simultaneously has been suggested to designate the onset of an arribada (Valverde and Gates 1999). However, the number of nesting turtles typically increases significantly over the night the arribada starts. In general, mass synchronous nesting events occur once a month or intermittently throughout the year at established arribada rookeries (Valverde et al. 1998; Coria-Monter and Durán-Campos 2017; Bézy et al. 2020). The timing and duration of these events is highly variable and unpredictable, and generally depend on the number of nesting turtles.
Worldwide, arribadas occur on only a few nesting beaches in the eastern Pacific, the western Atlantic, and the northern Indian Ocean (Bernardo and Plotkin 2007). Mass synchronous nesting of over 100,000 olive ridleys has been reported in the Pacific of Mexico (Ocana et al. 2012) and Costa Rica (Valverde et al. 1998, 2012; Fonseca et al. 2009), as well as in Orissa, on the east coast of India (Shanker et al. 2004). Two of the most important arribada beaches, in terms of the number of nesting females and relevance as focal points of research, are located in Costa Rica: Ostional and Nancite, both of which were unknown to science until the 1970s (Hughes and Richard 1974; Cornelius et al. 1991; Valverde et al. 1998; Fonseca et al. 2009, 2022). Ostional hosts the largest arribadas in Costa Rica with up to 476,550 egg-laying females estimated during a single mass synchronous nesting event (Valverde et al. 2012). Nancite formerly hosted large arribadas as well, with as many as 115,000 egg-laying females in a single mass synchronous nesting event (Valverde et al. 1998; Clusella Trullas and Paladino 2007; Fonseca et al. 2009). However, after experiencing a steep and continuous population decline over nearly 4 decades, arribadas currently are increasing at Nancite to consist of ~ 57,000 individuals per arribada (Fonseca et al. 2022).
Mass synchronous nesting events confer arribada nesting populations several advantages. First, the large number of hatchlings emerging synchronously acts as a predator satiation tactic (Eckrich and Owens 1995). The brief occurrence of prey at high densities reduces the probability of an individual being eaten, regardless of the usually high number of predators present at arribada beaches. In addition, arribada nesters can find mates more easily, as males also gather near arribada beaches searching for mates. In fact, the Ostional olive ridley population exhibits the highest level of multiple paternity determined for any sea turtle population (Jensen et al. 2006). Finally, arribada ridleys can delay nesting until weather conditions are suitable and the nesting process has a low energetic cost during the inter-nesting period, as the turtles remain near nesting sites (Bernardo and Plotkin 2007). Mass synchronous nesting, however, also has its disadvantages. Nest destruction for instance, is common at arribada beaches due to the high density of clutches and egg-laying females (Bézy et al. 2014; Cornelius et al. 1991). Nesting turtles dig up each other’s nests, resulting in a high microbial load, which may reduce hatching success (Fonseca et al. 2009; Valverde et al. 2012). Furthermore, the competition for space at the nesting site, attacks by a higher number of predators, and a relatively long nesting season are other disadvantages presented at arribadas nesting beaches (Bernardo and Plotkin 2007).
Corozalito is the third arribada beach to be documented in Costa Rica. Small-scale arribadas have been witnessed in this small Pacific coast beach over the last two decades (Viejobueno et al. 2011). Although the initial occurrence of arribadas at the site was not properly recorded, according to the neighboring community´s traditional knowledge they seem to have begun in the early 2000s (Mejías-Balsalobre et al. 2019). Unfortunately, until recently monitoring of nesting activity at this beach has focused on solitary nesting activity and not the arribada phenomenon, due in part to the site’s isolation and difficult access, as well as the unpredictability of arribada events and the additional resources required to monitor the event. However, we predict that the number and frequency of arribadas will increase in the next years, and that the hatching success will decrease as in other established arribada nesting beaches. Hence, the objectives of our study were to (1) report and assess the numbers of arribadas occurring at Corozalito since 2008, (2) estimate the number of egg-laying females using the strip transect in time method for the 2019, 2020, and 2021 arribadas, (3) assess hatching success and emergence rates via nest quadrat excavations, (4) estimate the incubation temperature in marked nests, and (5) record sand temperature and rainfall during the dry and rainy seasons.
Methods
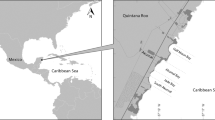
Corozalito (9°50′55.40′′ N; 85°22′ 47.67′′ W) is a small beach approximately 768 m long, located on the southern Nicoya Peninsula, Pacific coast of Costa Rica (Viejobueno et al. 2011) (Fig. 1). Rocky outcrops border both ends of the beach, with an estuary on the southern end that floods during the rainy season. The typical rainy and dry season patterns (July to December and December to June, respectively) of Costa Rica’s north Pacific are present in Corozalito, (Bernardo and Plotkin 2007; Dornfeld et al. 2015), with the largest number of olive ridley nesting events taking place between June and December (Binhammer et al. 2019). The site is uninhabited, with the 162-resident village of Corozalito located approximately 2.5 km inland (Picón-Cruz et al. 2014).

a Geographic location of Costa Rica in Central America. b Corozalito in the southern Nicoya Peninsula. Circles show the number olive ridleys tagged at other locations in Costa Rica that we detected at Corozalito
Arribada estimation
We conducted this study using olive ridley nesting data compiled during arribadas at Corozalito by the Sea Turtle Restoration Program (PRETOMA for its Spanish acronym) from 2008 to 2013 and by the Rescue Center for Endangered Marine Species (CREMA for its Spanish acronym) from 2014 to 2021. We recorded the total number of arribadas that occurred during this period as well as the duration (d) of each arribada event. Unfortunately, we were unable to provide an estimate of the size of arribadas up to 2018 because we had not yet implemented standardized protocols to census arribadas. In addition, during the 2020 nesting season, it was not possible to monitor the first night of the September, October and December arribadas due to lockdowns and restrictions resulting from the COVID-19 global health crisis. Thus, the number of egg-laying females during these arribadas is underestimated.
We used the strip transect in time method (Gates et al. 1996; Valverde and Gates 1999) to estimate the number of egg-laying females during arribada nesting events at Corozalito from June 2019 to December 2021. We modified the original arribada protocol by adjusting the length of the beach to 768 m and changing the definition of the formal initiation of the arribada from 100 nesting turtles simultaneously to 50, after which transect censuses begun. To conduct our study, we divided the beach into 30 sectors, each 25 m in width, using pole markers placed along the vegetation line and numbered in ascending order from North to South. We setup 2-m wide transects extending from the vegetation line to the high tide line at each sector post and left them for the duration of each arribada (Valverde and Gates 1999). We counted only egg-laying females that were inside the transects every 2 h throughout each arribada. We used the Arribada Portal website (Valverde 2013) to analyze the number of egg-laying female counts in each arribada and calculated the 95% confidence interval (CI95%) (Valverde et al. 2012).
In addition, we conducted nightly patrols from July to January to document the presence of turtles with tags (tag recaptures). For each recapture, we recorded the date, tag number, and nesting turtle’s behavior (arribada/solitary) (Tripathy and Pandav 2008). Subsequently, we tallied the number of recaptured turtles per year and location, and the distance from their tagging sites to Corozalito, if our teams did not originally tag them.
Nest quadrat excavations
We conducted quadrat nest excavations to estimate hatching success and emergence rates of nests laid during the October, November, and December arribadas of 2019, and for the arribada of December 2020. Our objective was to report on hatchling production of this new rookery. We excavated the nests 50–60 days after each arribada event. Nests were selected for excavations by placing 50 random 1 m2 PVC quadrats along the same transects that had been used to count nesting turtles, and each quadrat was excavated to a depth of 60 cm (Bézy et al. 2016). Since nest density was high at times, clutches tended to overlap. For this reason, we counted all eggs from all clutches of the corresponding arribada found in the quadrats as if they belonged to a single clutch. Clutches from other arribadas were ignored by distinguishing the condition of egg shells, as described by Bézy et al. (2016). Throughout 2020, we only excavated nests laid during the December arribada due to the COVID-19 logistical issues mentioned above.
Data from each quadrat excavation included: number of empty shells, hatchlings found inside each quadrat (either dead or alive), and unhatched eggs (with or without embryonic development) (Valverde et al. 2012). We then classified unhatched eggs into six different stages: stage 0 (no embryonic development), stage 1 (embryo size covered 1–25% of the egg), stage 2 (26–50%), stage 3 (51–75%), stage 4 (76–100%) and stage 5 (broken egg shells) (Valverde et al. 2010). The hatching success for each quadrat was defined as the number of broken eggshells/total number of eggs in the quadrat × 100 (Valverde et al. 2010; Bézy et al. 2016), whereas the emergence rate was defined as the number of turtles that emerged from the quadrat/total number of eggs in the quadrat × 100. We conceived the Index of Development (IoD) as a way to assess and characterize the quality of the incubation microenvironment. The objective was to have an idea of the developmental stage at which the embryos perished, if at all. We calculated the IoD using the nest excavation values for each quadrat as follows: \(\mathrm{IoD}= {\sum }_{s=1-6}\left(\frac{S*{n}_{i}}{n.}\right)\), where S = is the stage category (1–6); ni = number of embryos per stage, and n. = total number of eggs. The IoD value ranges from 1 to 6, 1 being the lowest value and thus represents the poorest incubation conditions, and 6 is the maximum possible value, conducive to proper embryonic development and hatching.
We evaluated monthly differences in hatching success, emergence rate and IoD by means of an analysis of variance (one-way ANOVA) and a post-hoc test to determine which month or months were significantly different for each variable (α = 0.05).
Nest, sand temperature and rainfall
We marked 18 nests with Onset HOBO pendant temperature loggers (Onset Computer, UA-001-08, accuracy: ± 0.53 °C, resolution: 0.1 °C) during the arribadas of September 2021 and November 2021. We selected nests randomly during the arribadas by choosing females that were laying eggs in the arribada nesting area (Valverde et al. 2010). We counted the eggs directly using a red light as they dropped in the nest, and we inserted the data logger into the center of each nest after approximately 50 eggs were laid without disturbing the nesting process (Bézy et al. 2014). To protect the nest from other nesters, predators, or egg extraction, we covered each nest with a 40 × 40 × 25 cm wire mesh cage (Valverde et al. 2010, 2012) and we buried it entirely below the surface of the sand. On day 40, we removed the wire mesh cages, and we monitored each nest daily for signs of hatchling emergence (Bézy et al. 2014). Finally, nests were excavated 2 days after the last hatchling emergence (Valverde et al. 2012). Upon excavations, we retrieved data loggers and recorded the developmental stage for all nests (Valverde et al. 2010).
We counted all broken shells and undeveloped eggs in the marked nests and classified them as explained above. We calculated hatching rate for each nest as the following: (number of hatchlings in nest)/(total number of eggs) × 100. In addition, we made general observations of the nests (Valverde et al. 2012). Hatching success differences between September and November arribadas in 2021 were evaluated with a t test; the significance level was set at α = 0.05. We used STATISTICA®v7 for all statistical analysis.
We recorded sand temperature (°C) hourly from July 2019 to January 2020, and from September to December in 2021. We measured sand temperature in 2019 using a single data logger, whereas in 2021 an additional data logger was used to measure air temperature. We buried sand temperature loggers in the middle of the beach and close to the vegetation line, at 40 cm of depth. We placed air data logger in the shade on a palm tree at a height of approximately 1.50 m. All data loggers were programmed to collect temperature data every 2 h. Data logger recording consistency and accuracy was ensured by placing sensors in a bag simultaneously and keeping them in a shaded room to record air temperature for a period of 38 h (recordings included 2-night periods) (Valverde et al. 2010). We downloaded temperature data and converted it to spreadsheets using HOBOware Pro 3.2.0 (Valverde et al. 2010). Rainfall (mm) was recorded with a rain gauge twice a day in 2019 and 2021, at 0700 h and 1900 h. We then calculated monthly and seasonal (rainy and dry seasons) average temperature and precipitation (Valverde et al. 2010).
Results
Arribada estimates
Twenty-nine arribada events were recorded at Corozalito during the study period (2008 to 2021), all of which occurred from August to January. No mass synchronous nesting events were recorded in 2008, 2009, 2010 and 2015, although we did record the occurrence of nesting activity during those years with over 50 nesting turtles but not simultaneously. A single arribada per year occurred in 2011, 2012 and 2013, with durations from 1 to 3 nights. Three arribadas occurred in a single year for the first time in 2014, one with a duration of 4 nights. One of the four arribadas that occurred in 2017 had a duration of five nights. Five arribadas occurred in 2018, and for the first time two arribadas occurred in a single month, twice (in September and December). In 2019, one arribada occurred per month from August to December and in 2020, four arribadas occurred and for the third time two arribadas occurred in a single month (October). Finally, in 2021, we recorded three arribadas and for second time one of the arribadas lasted 5 days (Fig. 2).

Duration of arribadas per month and year of occurrence at Corozalito from 2011 to 2021. No arribada occurred in 2008, 2009, 2010, and 2015
We estimated that 150,150 egg-laying females participated during the 12 arribadas that occurred at Corozalito during the 2019, 2020, and 2021 nesting seasons, respectively. The largest mass synchronous nesting event we recorded thus far occurred in October 2021, with an estimated 21,653 ± 3327 (estimate ± CI95%) egg-laying females. In contrast, the smallest arribada occurred in December of 2019, with an estimated 2,139 ± 807 egg-laying females (Fig. 3). The average interval between arribada events from 2011 to 2021 was 28.1 ± 1.0 days (range 16–70 days; median 25.0), and the mean time span or duration of the arribadas was 2.82 ± 1.1 nights.

Estimated number of olive ridley egg-laying females and associated 95% confidence intervals (CI 95%) for each mass synchronous nesting event in 2019, 2020 and 2021 nesting seasons at Corozalito, Costa Rica. Non-overlapping CI95% indicate significantly different estimates
We recaptured (tag recovery) 460 turtles that had been tagged previously at Corozalito and 87 turtles that had been tagged at 8 different locations along the Pacific coast of Costa Rica, ranging in distance from 8 km (Camaronal) to 222 km (Osa Peninsula) (Fig. 1b). A slight majority of the recaptured turtles were found nesting solitarily, whereas the remaining ones were nesting during arribada nesting events (51.1%, n = 235 and 48.9%, n = 225 respectively). Among the turtles tagged at other locations, Montezuma and Camaronal contributed the greatest number of recaptures (40.2%, n = 35 and 39.1%, n = 34, respectively). Twenty-six of these turtles (29.9%) were recorded during solitary nesting activity and 61 (70.1%) during arribada nesting activity in Corozalito. Most of the recaptures occurred in 2018 (24.1%, n = 21) and 2016 (22.9%, n = 20), whereas no recaptures occurred in 2008, 2010, nor 2012 (Table 1). We did not detect tags from other countries at Corozalito. In addition, 47 turtles tagged by various CREMA projects around the Nicoya Peninsula have been found nesting at Ostional beach since 2012. Tag searching effort was not uniform throughout the study period.
Quadrat hatching success, emergence rates, and IoD
Mean hatching success of quadrat nests for the October, November, and December arribadas of 2019 was 66.2% ± 34.6, n = 150 (range 0–100%), and 65.3% ± 33.3, n = 50 (range 0–100%) for the arribada event of December 2020. Hatching success of all arribadas was significantly different among them (ANOVA, F (1,3) = 525.7430, p < 0.00). The mean emergence rate recorded for the three 2019 arribadas was 65.8% ± 34.9, n = 150 (range 0–100%), and 65.2% ± 33.2, n = 50 (range 0–100%) for the December 2020 arribada. The highest values for both hatching success (70.4% ± 32.2, n = 50) and emergence rate (69.7% ± 32.6, n = 50) were recorded in October 2019 and the lowest (56.3% ± 36.8, n = 50) in November 2019 (Fig. 4).

a Hatching (%) and b emergence rates (%) estimated for the October, November and December arribadas of 2019 and the December arribada of 2020 in Corozalito. Triangles indicate that hatching rate and emergence success for November of 2019 were significantly lower than the other months
Average IoD for the October, November, and December arribadas of 2019 was 4.53 ± 1.59, n = 150 (range 1.00–6.00), and 4.50 ± 1.48, n = 50 (range 1.00–6.00) for the arribada of December 2020. IoD differed significantly among arribadas (ANOVA, F (1,3) = 1120.596, p < 0.00), with the November 2019 arribada displaying the lowest IoD (Tukey, p = 0.0498) (Fig. 5).

Index of Development (IoD) for the three mass synchronous nesting events monitored in 2019 and the single arribada monitored in 2020 at Corozalito. Asterisk indicates that the arribada of November 2019 presented a significantly lower IoD
Sand, air temperature and rainfall
Average sand temperature during the 2019 nesting season was 31.17 °C ± 2.21, n = 204 (range 24.86–35.16 °C) and 29.69 °C ± 1.11, n = 174 (range 24.06–34.05 °C) for the 2021 nesting season (Fig. 6). In 2019 and 2021, we recorded the highest mean values in December (33.68 °C ± 0.82; 31.85 °C ± 1.57, respectively). We recorded the lowest mean temperatures in October of 2019 (28.83 °C ± 1.28) and in August of 2021 (28.49 °C ± 0.82). We recorded a mean air temperature in 2021 of 26.72 °C ± 2.84, n = 174.

Monthly mean (± SE) sand temperature (dots) and rainfall (bars) recorded at Corozalito during the a 2019 and b the 2021 nesting seasons
Average rainfall during 2019 and 2021 was 11.0 ± 3.5 mm, n = 202 (range 0.00–254.00 mm) and 4.01 ± 3.1 mm, n = 174 (range 0.00–112.00 mm), respectively. Greatest precipitation occurred during October 2019 (34.1 mm ± 9.4) and September 2021 (7.9 mm ± 14.88) (Fig. 6).
Nest incubation temperature and hatching success of marked nests
The mean incubation temperature of marked nests from September to December 2021 was 32.3 °C ± 2.41, n = 15. Nest incubation temperatures were monitored for nests laid in September (31.62 °C ± 1.38, n = 8; range 26.45–37.54 °C) and November (33.16 °C ± 1.57, n = 7; range 27.01–37.66 °C) (Fig. 7). The lowest mean incubation temperature for these nests occurred during the first days of incubation (30.88 °C ± 2.2 and 31.87 °C ± 2.2, respectively). Nest temperatures increased during the incubation period (32.06 °C ± 2.16 and 34.50 °C ± 2.04, respectively). Among the data loggers (n = 18), one of them was vandalized along with the nest and two of them did not record the temperature; however, hatching success was documented in the remaining nests.

Daily nest incubation temperatures (45 days) for a September (n = 8) and b November nests (n = 7) (open squares), sand temperature (closed circles), and air temperature (closed triangles) at Corozalito. The red line indicates field lethal temperatures for nests (35.0 °C) (Valverde et al. 2010), and the blue line indicates olive ridley sex determination mean pivotal temperature (30.5 °C) (Wibbels et al. 1998)
All of the nests marked during the September and November 2021 arribadas produced hatchlings. Mean hatching success for September marked nests was 57.1% ± 26.5, n = 9 (range 18.7–93.7%), and 58.2% ± 25.0, n = 7 (range 35.1–97.4%) for November marked nests. Hatching success differed significantly between the nests marked during these 2 months (t test = 7.05749, p < 0.00). Embryonic development was prone to stop in nests with incubation temperatures reaching above 31.5 °C (n = 12) (Fig. 8).

Correlation between hatching success and mean incubation temperature (°C) recorded during the incubation period (45 days) of nests marked in September and November arribadas of 2021
Discussion
Corozalito arribadas
We recorded 29 arribadas at Corozalito from 2008 to 2021, which displayed a clear growing trend in annual frequency and days of duration. At present, there is a lack of information available regarding the initial stages of these mass nesting events at Corozalito. In 1970, survey flights were conducted along the coast of the Nicoya Peninsula, during which less than ten tracks were recorded near Corozalito (Richard and Hughes 1972). The first official report of an arribada occurring in Corozalito was made in 2007 by the local community (R. Arauz personal communication). Arribadas in Corozalito nonetheless, may have begun earlier (Mejías-Balsalobre et al. 2019).
Little is known regarding the mechanisms and the cues that lead to the establishment of arribadas at specific beaches, likely because so far, they do not follow any pattern, are highly ephemeral, occur suddenly or provide no clues to facilitate our understanding, and because they are discovered in remote locations (Bernardo and Plotkin 2007; Pritchard 2007; Fonseca et al. 2009). Corozalito seems to be in its early stages of development as an arribada beach, given the large fluctuations in size and frequency observed relative to more established arribada rookeries like Ostional (Pritchard 2007). We documented the largest arribadas in September 2019 (17,982 ± 3220 egg-laying turtles) and October 2021 (21,653 ± 3327 egg-laying turtles), coinciding with the rainiest months of the year. This is consistent with the occurrence of the largest arribada events in the Eastern Tropical Pacific (ETP) region, which have been associated with rainy season months and hence, lower sand temperatures (Hughes and Richard 1974; Plotkin et al. 1997; Bernardo and Plotkin 2007; Corina-Monter and Durán-Campos 2017). Overall, arribadas at Corozalito are small when compared to those of larger well-known arribada beaches, such as Ostional with as many as 476,550 egg-laying turtles (Valverde et al. 2012), Nancite with 57,907 (Fonseca et al. 2022), La Flor, Nicaragua with 60,816 (Honarvar et al. 2016), La Escobilla, México with 317,219 (Corina-Monter and Durán-Campos 2017) and Orissa, India with 180,000 (Shanker et al. 2004). It appears that the abundance of nesting olive ridley turtles at Corozalito is insufficient to yield monthly arribadas across an entire year, as does occur at nearby Ostional.
While arribadas rarely occur simultaneously at Ostional and Nancite, with a time lag of days or even weeks apart (Hughes and Richard 1974; Bernardo and Plotkin 2007; Bézy et al. 2020), synchronicity has been observed during the occurrence of nine arribadas at Corozalito and Ostional for the last 3 years, initiating on either the same day or a couple days apart. Interestingly, the current interval between arribadas at Corozalito (28.1 ± 1.0 days) seems different to Ostional’s (34 ± 1.4 days) (Bézy et al. 2020). Synchronicity between arribada sites may be due to independent turtles responding to the same cues that trigger arribada events (Plotkin et al. 1995), a possibility that increases with proximity. It is possible that arribadas at Corozalito may continue to increase and evolve for a few decades, only to wane and collapse when the arribada becomes self-limiting, as occurred at Nancite (Fonseca et al. 2009, 2022; Pritchard 2007; Valverde et al. 2012), which is similar in length to Corozalito (1 km). Despite the lack of estimates regarding the size of the arribadas at Corozalito from 2008 to 2018, our observations are relevant to document their historic occurrence, particularly considering the lack of published information on the development of this unique behavior in its early stages. Further monitoring is needed at this site to assess the fate and trend of the nesting population and to formulate adequate management plans to ensure its protection, as currently this arribada rookery is located on public unprotected land.
Olive ridleys tag recapture in Corozalito
We observed an exchange of olive ridleys between Corozalito and other nesting sites through Inconel flipper tag returns, 51.1% of which belonged to solitary nesters and 48.9% to arribada nesters. The majority of turtles recaptured at Corozalito were tagged at Montezuma (40 km away) and the nearby beach of Camaronal (8 km away), but also included turtles that had been tagged at Ostional and Nancite, or at other nesting sites along the Pacific coast of Costa Rica. An occasional exchange of either solitary or arribada nesting olive ridley sea turtles has been reported between Ostional and Nancite (Cornelius and Robinson 1981; Cornelius and Robinson-Clark 1986). Likewise, a similar phenomenon of nesting turtles at sites that had been tagged nesting elsewhere has been observed in Chacocente and La Flor arribada rookeries in Nicaragua (Heidy Salazar, personal communication). Our tag recovery data shows that nesting turtles tagged at different beaches along the Pacific coast of Costa Rica also nest in Corozalito during solitary or arribada nesting events. It is known that olive ridleys tagged in Ostional and Nancite move to other regions of the ETP, and that they potentially nest at beaches as far as Oaxaca, Mexico (Cornelius and Robinson-Clark 1986; Plotkin 2010). Taking together our recapture observations plus those at other arribada nesting sites, it seems that large arribadas do not build gradually over time at a beach spurred by high hatching rates, as currently thought (Valverde et al. 1998; Fonseca et al. 2009). Rather, it seems that these events first establish relative quickly as large groups of olive ridleys that spin off aggregations from regional waters. It is also possible that both hypotheses work in concert to some extent to build over time an arribada aggregation.
It is known that arribada nesters display some degree of site fidelity in Costa Rica’s arribada rookeries (Plotkin et al. 1995; Plotkin 2002), although our findings clearly show that this fidelity is not absolute. Olive ridleys exhibit philopatric plasticity, where animals return to nest to a given region, but not necessarily to the same beach over their lifetime. This behavior is not rare and has been documented for several sea turtle species using satellite tags, metal tags and even genetics (López-Castro and Rocha-Olivares 2005; Georges et al. 2007; Nordmoe et al. 2004; Tucker 2010).
Mark recapture studies have suggested that each arribada is in part regulated by cues present in the local environment (Bernardo and Plotkin 2007). Thus, each arribada rookery may exhibit its own idiosyncrasy. Unfortunately, the low number of tag recovery rates characteristic of arribada rookeries is a factor that limits our understanding of olive ridley beach selection and overall nesting behavior. Tag recapture may vary due to the relatively small number of turtles tagged and significant tag loss; moreover, it is difficult to detect tagged turtles during mass nesting events given the difficulty of finding those few animals among the thousands that participate in the events (Cornelius and Robinson-Clark 1986). Finally, few monitoring programs deploy teams dedicated to finding previously tagged turtles given the multiple tasks conducted during the events and the limitations on numbers of workers. Altogether, it is possible that the exchange of nesting beaches by particular nesters is part of a complex behavior to colonize new areas or beaches (Tripathy and Pandav 2008). If so, this may explain the seeming lack of genetic population structure in olive ridleys across most of the ETP (López-Castro and Rocha-Olivares 2005). Unfortunately, we do not know why olive ridleys are currently choosing to nest at Corozalito, or the mechanism by which nesting animals are recruited to this nesting beach.
Hatching success and development index at Corozalito
Mass nesting events may lead to olive ridley arribada beaches reaching their carrying capacity, resulting in a decrease in hatchling production and low recruitment to the adult population over time, as is suspected to have occurred at Nancite (Bernardo and Plotkin 2007; Cornelius et al. 1991; Fonseca et al. 2009, 2022; Pritchard 2007; Valverde et al. 1998). However, this is not the case for Corozalito at this point in time, since current hatching success in arribada quadrats (65%) and marked nests (59%) were higher than those of other arribada beaches such as Nancite (33.4%) (Fonseca et al. 2022), Ostional (18.4%) (Valverde et al. 2012) and La Flor (19.0%) (Honarvar et al. 2016). The high hatching success relative to other arribada beaches is indicative that Corozalito is a nascent and thus, young arribada beach, and that nest destruction and microbial load are not yet critical. In this context, current hatching success may be high enough to support the relatively quick establishment of a large arribada population at the site. In contrast, the hatching success that we observed at Corozalito, when compared with the between 80 and 90% hatching rates of solitary olive ridley nesting beaches (Dornfeld et al. 2015; Binhammer et al. 2019), appear to be low, suggesting that Corozalito is on a trend to reaching its carrying capacity. However, if arribada populations are largely supported by regional turtles (Plotkin et al. 1995), it is possible that the establishment of Corozalito as an arribada rookery is independent of its hatchling production.
According to our findings, the IoD at Corozalito was 4.5. This indicates that in general, significant mortality took place in later stages of development. It is possible that this mortality was due to factors that characterize well-established arribada beaches such as Ostional and Nancite, owing to their suboptimal incubation conditions, which drive high embryo mortality rates (Clusella Trullas and Paladino 2007; Honarvar et al. 2008; Valverde et al. 2010, 2012; Bézy et al. 2014, 2016). It has been suggested that optimal embryo development is regulated by a range of environmental variables that define a suitable microclimate for development, where the physical characteristics of the beach, local climate, and the number of eggs laid in the clutch are key factors that guarantee embryo development (Ackerman 1997). We feel that the IoD reflects those variables and that it will serve as a tool to determine optimal incubation conditions. Finally, we are confident the continued estimation of an IoD at solitary and arribada rookeries subject to various environmental conditions will eventually enable us to properly interpret the meaning of these values. For now, it seems that a value of 4.5 is reasonably good for Corozalito given the hatching success obtained.
Nest incubation temperature
Sea turtles exhibit temperature-dependent sex determination, which is characterized by the exclusive production of a single sex below (male) or above (female) a transitional range of temperatures that can vary among sea turtle species (Ackerman 1997; Porter et al. 2021; Wibbels 2003; Wibbels et al. 1998), as well as a pivotal temperature that produces a sex ratio of 1:1 (Mrosovsky and Yntema 1980). Mean nest incubation temperature at Corozalito (32.3 °C) was higher than the estimated sex determination pivotal temperature for olive ridleys (30.5 °C) in the ETP (Wibbels et al. 1998; Wibbels 2007; Morales Mérida et al. 2021) and below the lethal temperature (35 °C) documented at Ostional (Valverde et al. 2010). The absence of rainfall during 2021 likely led to the high incubation temperatures recorded during the study period, as rainfall causes sand and nest temperatures to drop for several days (Laloë et al. 2021; Porter et al. 2021). The high temperatures recorded at Corozalito led to embryonic mortality in several nests, as occurred at Ostional (Valverde et al. 2010). According to our findings, most of Corozalito’s incubation temperatures are below the lethal threshold (Valverde et al. 2010), and hatching success and the recruitment rates are high, with the beach currently producing a hatchling sex ratio skewed towards females.
The preponderant theoretical production of females at Corozalito may contribute to an increase in the number of nesting olive ridleys in the region, which may eventually contribute to an increase in the size of arribadas at this and other nesting sites. However, it is important to consider the effect the variation of temperatures during the incubation period could have on hatchling rates (Morales Mérida et al. 2021) and recruitment to mass nesting events (Wibbels 2007). Further research is needed at Corozalito to assess incubation temperature in olive ridley turtles during several seasons and compare them with larger nesting beaches such Ostional, Nancite, or La Flor, to understand the development of this nesting beach over time.
Nest predation and egg extraction
Predation and egg extraction by humans increase dramatically during arribadas (Eckrich and Owens 1995). Egg extraction (poaching) has been reported as a major threat to sea turtle along the beaches of the southern Nicoya Peninsula (Viejobueno et al. 2011). We did not assess predation or poaching at Corozalito, although one of the nests that we marked during the November arribada with a data logger was stolen a couple of days later. In addition, Pheasey et al. (2020) tracked a poached turtle nest from Corozalito to a supermarket in the Central Valley of Costa Rica, 137 km away. In this regard, it is important to mention the extraction of sea turtle eggs for domestic commercialization is legal in Costa Rica only at Ostional (Campbell 1998; Valverde et al. 2012). Furthermore, it is essential to assess all threats present at Corozalito, including the illegal extraction of eggs and predation, particularly at this early developmental stage for the proper study of the evolution of this nascent arribada beach.
Towards conservation in Corozalito
This study provides supporting evidence that the number and frequency of arribadas in Corozalito will increase over time. We also, predict that hatching success will decrease over time but will be higher than more established arribada nesting beaches. As such, only the continued monitoring of nesting dynamics at this site will shed light on the fate of Corozalito as an arribada rookery, and perhaps on the mechanisms by which arribada beaches are born, and evolve. Our research provides insight into the contribution of nesting turtles from independent nesting beaches to the creation and consolidation of arribada beaches, and opens new questions about this phenomenon. Finally, it is extremely important to provide Corozalito with National Wildlife Refuge status for the official implementation of a management plan to ensure its long-term protection, as its current location on public unprotected land is to be considered a threat.
Data availability
The authors declare the availability of the data.
Code availability
Not applicable.
References
Abreu-Grobois A, Plotkin PT (2008) Lepidochelys olivacea, olive ridley. The IUCN Red List Threat Species. e.T11534A3292503. https://doi.org/10.2305/IUCN.UK.2008.RLTS.T11534A3292503.en%5CnCopyright
Ackerman RA (1997) The nest environment and the embryonic development of sea turtles. In: Lutz PL, Musick JA (eds) The biology of sea turtles, vol I. CRC Press, Boca Raton, pp 83–106
Bernardo J, Plotkin PT (2007) An evolutionary perspective on the arribada phenomenon and reproductive behavioral polymorphism of olive ridley sea turtles (Lepidochelys olivacea). In: Plotkin PT (ed) Biology and conservation of ridley sea turtles. Johns Hopkins University Press, Baltimore, pp 59–87
Bézy VS, Valverde RA, Plante CJ (2014) Olive ridley sea turtle hatching success as a function of the microbial abundance and the microenvironment of in situ nest sand at Ostional, Costa Rica. J Mar Biol. https://doi.org/10.1371/journal.pone.0118579
Bézy VS, Girondot M, Valverde RA (2016) Estimation of the net nesting effort of olive ridley arribada sea turtles based on nest densities at Ostional beach, Costa Rica. J Herpetol 50:409–415. https://doi.org/10.1670/14-152
Bézy VS, Putman NF, Umbanhowar JA, Orrego CM, Fonseca LG, Quirós-Pereira WM, Valverde RA, Lohmann KJ (2020) Mass-nesting events in olive ridley sea turtles: environmental predictors of timing and size. Anim Behav 163:85–94. https://doi.org/10.1016/j.anbehav.2020.03.002
Binhammer MR, Beange M, Arauz R (2019) Sand temperature, sex ratios, and nest success in olive ridley sea turtles. Mar Turt Newsl 159:5–9
Campbell LM (1998) Use them or lose them? Conservation and the consumptive use of marine turtle eggs at Ostional, Costa Rica. Environ Conserv 25:305–319. https://doi.org/10.1017/S0376892998000393
Clusella Trullas S, Paladino FV (2007) Micro-environment of olive ridley turtle nests deposited during an aggregated nesting event. J Zool 272:367–376. https://doi.org/10.1111/j.1469-7998.2006.00277.x
Coria-Monter E, Durán-Campos E (2017) The relationship between the massive nesting of the olive ridley sea turtle (Lepidochelys olivacea) and the local physical environment at la Escobilla, Oaxaca, Mexico, during 2005. Hidrobiologica 27:201–209. https://doi.org/10.24275/uam/izt/dcbs/hidro/2017v27n2/coria
Cornelius SE, Robinson DC (1981) Abundance, distribution and movements of olive ridley sea turtles in Costa Rica. Endangered Species Report. U.S. Fish and Wildlife Service, New Mexico, Albuquerque, NM
Cornelius SE, Robinson-Clark DC (1986) Post-nesting movements of female olive ridley turtles tagged in Costa Rica. Vida Silvestre Neotropical 1(1):12–23
Cornelius SE, Ulloa MA, Castro JC, Mata Del Valle M, Robinson DC (1991) Management of olive ridley sea turtles (Lepidochelys olivacea) nesting at Playas Nancite and Ostional, Costa Rica. In: Robinson JGRK (ed) Neotropical widlife use and conservation. Neotropica University of Chicago Press, Chicago, pp 111–135
Cornelius SE, Arauz R, Fretey J, Godfrey MH, Márquez-M R, Shanker K (2007) Effect of land-based harvest of Lepidochelys. In: Plotkin PT (ed) The biology and conservation of ridley sea turtles. Johns Hopkins University Press, Baltimore, pp 231–251
Dornfeld TC, Robinson NJ, Tomillo PS, Paladino FV (2015) Ecology of solitary nesting olive ridley sea turtles at Playa Grande, Costa Rica. Mar Biol 162:123–139. https://doi.org/10.1007/s00227-014-2583-7
Eckrich CE, Owens DW (1995) Solitary versus arribada nesting in the olive ridley sea turtles (Lepidochelys olivacea): a test of the predator-satiation hypothesis. Herpetologica 51:349–354
Fonseca LG, Murillo GA, Guadamúz L, Spínola RM, Valverde RA (2009) Downward but stable trend in the abundance of arribada olive ridley sea turtles (Lepidochelys olivacea) at Nancite beach, Costa Rica (1971–2007). Chelonian Conserv Biol 8:19–27. https://doi.org/10.2744/CCB-0739.1
Fonseca LG, Villachica WN, Rangel E, Palola E, Gilbert M, Valverde RA (2022) Reassessment of the olive ridley sea turtle Lepidochelys olivacea (Testudines: Cheloniidae) nesting population at Nancite beach, Costa Rica. Marine Biology, submitted
Gates CE, Valverde RA, Mo CL, Chaves AC, Ballesteros J, Peskin J (1996) Estimating arribada size using a modified instantaneous count procedure. J Agric Biol Environ Stat 1:275–287
Georges JY, Fossette S, Billes A, Ferraroli S, Fretey J, Grémillet D, Le Maho Y, Myers AE, Tanaka H, Hays GC (2007) Meta-analysis of movements in Atlantic leatherback turtles during the nesting season: conservation implications. Mar Ecol Prog Ser 338:225–232. https://doi.org/10.3354/meps338225
Honarvar S, O’Connor MP, Spotila JR (2008) Density-dependent effects on hatching success of the olive ridley turtle, Lepidochelys olivacea. Oecologia 157:221–230. https://doi.org/10.1007/s00442-008-1065-3
Honarvar S, Brodsky MC, Van Den Berghe EP, O’Connor MP, Spotila JR (2016) Ecology of olive ridley sea turtles at arribadas at playa la Flor, Nicaragua. Herpetologica 72:303–308. https://doi.org/10.1655/Herpetologica-D-16-00014.1
Hughes DA, Richard JD (1974) The nesting of the Pacific ridley turtle Lepidochelys olivacea on Playa Nancite, Costa Rica. Mar Biol 24:97–107
Jensen MP, Abreu-Grobois FA, Frydenberg J, Loeschcke V (2006) Microsatellites provide insight into contrasting mating patterns in arribada vs. non-arribada olive ridley sea turtle rookeries. Mol Ecol 15:2567–2575. https://doi.org/10.1111/j.1365-294X.2006.02951.x
Laloë JO, Tedeschi JN, Booth DT, Bell I, Dunstan A, Reina RD, Hays GC (2021) Extreme rainfall events and cooling of sea turtle clutches: implications in the face of climate warming. Ecol Evol 11:560–565. https://doi.org/10.1002/ece3.7076
López-Castro MC, Rocha-Olivares A (2005) The panmixia paradigm of eastern Pacific olive ridley turtles revised: consequences for their conservation and evolutionary biology. Mol Ecol 14:3325–3334. https://doi.org/10.1111/j.1365-294X.2005.02652.x
Mejías-Balsalobre C, Rojas-Cañizales D, Fusté-Mach R, Webster EG, Arauz D, Barrios-Garrido H, Arauz R (2019) Using traditional ecological knowledge to complement the gaps of infromation: the shifting baseline of arribadas as case study. In: Proceedings of the thirty-nine annual symposium on sea turtle Biology and Conservation. Charleston, USA., p 2019
Morales Mérida A, Helier A, Cortés-gómez AA, Girondot M (2021) Hatching success rather than temperature-dependent sex determination as the main driver of olive ridley (Lepidochelys olivacea) nesting activity in the pacific coast of central America. Animals 11:3168. https://doi.org/10.3390/ani11113168
Mrosovsky N, Yntema CL (1980) Temperature dependence of sexual differentiation in sea turtles: implications for conservation practices. Biol Conserv 18:271–280
Nordmoe ED, Sieg AE, Sotherland PR, Spotila JR, Paladino FV, Reina RD (2004) Nest site fidelity of leatherback turtles at Playa Grande, Costa Rica. Anim Behav 68:387–394. https://doi.org/10.1016/j.anbehav.2003.07.015
Ocana M, Harfush-Melendez M, Heppell S (2012) Mass nesting of olive ridley sea turtles Lepidochelys olivacea at La Escobilla, Mexico: linking nest density and rates of destruction. Endanger Species Res 16:45–54. https://doi.org/10.3354/esr00388
Pheasey H, Roberts DL, Rojas-Cañizales D, Mejías-Balsalobre C, Griffiths RA, Williams-Guillen K (2020) Using GPS-enabled decoy turtle eggs to track illegal trade. Curr Biol 30:R1066–R1068. https://doi.org/10.1016/j.cub.2020.08.065
Picón-Cruz JC, Rodríguez Quirños R, Bravo Chacón J (2014) Experiencias en investigación y manejo de humedales interiores y marino-costeros en Costa Rica. Experiencias en investigación y manejo de humedales interiores y marino-costeros en Costa Rica
Plotkin P (2002) Adult migrations and habitat use. In: Lutz PL, Musick JA (eds) The biology of sea turtles, vol II. CRC Press, Boca Raton, pp 225–241
Plotkin PT (2010) Nomadic behaviour of the highly migratory olive ridley sea turtle Lepidochelys olivacea in the eastern tropical Pacific Ocean. Endanger Species Res 13:33–40. https://doi.org/10.3354/esr00314
Plotkin PT, Byles RA, Rostal DC, Owens DW (1995) Independent versus socially facilitated oceanic migrations of the olive ridley, Lepidochelys olivacea. Mar Biol 122:137–143. https://doi.org/10.1007/BF00349287
Plotkin PT, Rostal DC, Byles RA, Owens DW (1997) Reproductive and developmental synchrony in female Lepidochelys olivacea. J Herpetol 31:17–22. https://doi.org/10.2307/1565323
Porter E, Booth DT, Limpus CJ, Staines MN, Smith CE (2021) Influence of short-term temperature drops on sex-determination in sea turtles. J Exp Zool Part A Ecol Integr Physiol 335:649–658. https://doi.org/10.1002/jez.2509
Pritchard PCH (1997) Evolution, phylogeny and current status. In: Lutz PL, Musick JA (eds) The biology of sea turtles. CRC Press, Boca Raton, pp 1–28
Pritchard PCH (2007) Arribadas I have known. In: Plotkin PT (ed) Biology and conservation of ridley sea turtles. Johns Hopkins University Press, Baltimore, pp 7–21
Richard JD, Hughes DA (1972) Some observations of sea turtle nesting activity in Costa Rica. Mar Biol 16:297–309. https://doi.org/10.1007/BF00347753
Shanker K, Pandav B, Choudhury BC (2004) An assessment of the olive ridley turtle (Lepidochelys olivacea) nesting population in Orissa, India. Biol Conserv 115:149–160. https://doi.org/10.1016/S0006-3207(03)00104-6
Tripathy B, Pandav B (2008) Beach fidelity and internesting movements of olive ridley turtles (Lepidochelys olivacea) at Rushikulya, India. Herpetol Conserv Biol 3:40–45
Tucker AD (2010) Nest site fidelity and clutch frequency of loggerhead turtles are better elucidated by satellite telemetry than by nocturnal tagging efforts: implications for stock estimation. J Exp Mar Bio Ecol 383:48–55. https://doi.org/10.1016/j.jembe.2009.11.009
Valverde RA (2013) Arribada portal: the globalization of arribada estimates. Mar Turt Newsl 137:16–17
Valverde R, Gates C (1999) Population surveys on mass nesting beaches. In: Eckert KL, Bjorndal KA, Abreu-Grobois FA (eds) Research and management techniques for the conservation of sea turtles, vol 2. IUCN/SSC Marine Turtle Specialist Publication, pp 56–60
Valverde RA, Cornelius SE, Mo CL (1998) Decline of the olive ridley des turtle (Lepidochelys olivacea) nesting assemblage at Nancite Beach, Santa Rosa National Park, Costa Rica. Chelonian Conserv Biol 3:58–63
Valverde RA, Wingard S, Gómez F, Tordoir MT, Orrego CM (2010) Field lethal incubation temperature of olive ridley sea turtle Lepidochelys olivacea embryos at a mass nesting rookery. Endanger Species Res 12:77–86. https://doi.org/10.3354/esr00296
Valverde RA, Orrego CM, Tordoir MT, Gómez FM, Solís DS, Hernández RA, Gómez GB, Brenes LS, Baltodano JP, Fonseca LG, Spotila JR (2012) Olive ridley mass nesting ecology and egg harvest at Ostional beach, Costa Rica. Chelonian Conserv Biol 11:1–11. https://doi.org/10.2744/CCB-0959.1
Viejobueno S, Adams C, Arauz R (2011) Oportunidades para el desarrollo sostenible de las comunidades costeras de Nandayure (Nicoya sur, Guanacaste). Rev Ciencias Ambient 41:37. https://doi.org/10.15359/rca.41-1.5
Wibbels T (2003) Critical Approaches to Sex Determination in Sea Turtles. In: Lutz PL, Musick JA, Jeanette W (eds) The biology of sea turtles, vol II. CRC press, Boca Raton, pp 103–134
Wibbels T (2007) Sex determination and sex ratios in ridley turtles. In: Plotkin PT (ed) Biology and conservation of ridley sea turtles. Johns Hopkins University Press, Baltimore, pp 167–211
Wibbels T, Rostal D, Byles R (1998) High pivotal temperature in the sex determination of the olive ridley sea turtle, Lepidochelys olivacea, from Playa Nancite. Costa Rica Copeia. https://doi.org/10.2307/1447364
Acknowledgements
We would like to thank to research assistants, locals and volunteers who over the past years have helped us collecting data and monitoring the beach. We thank Turtle Trax S.A for all the logistics support during this research. We also would like to thank Madelaine Beange for her collaboration and encouragement during the early stages of the monitoring project at Corozalito. We thank to Whitley Fund for Nature and Sandler Foundation for their support. We are grateful with Gary Kaasa, Mike Hopkins and John Lindsay for all their support since the Corozalito project started. This project was funded by RIESTER Foundation. This study was conducted under MINAE, SINAC, and ACT research permits [ACT-OR-DR-055-2020; ACT-OR-DR-063-2021]. We would also like to thank the contribution of two valuable reviewers, who greatly helped to improve the quality of this paper.
Funding
This project and study was funded by Riester Foundation.
Author information
Authors and Affiliations
Contributions
DR-C: conceptualization, methodology, formal analysis, investigation, resources, data curation, project administration, funding acquisition, writing—original draft, and visualization. CM-B: methodology, investigation, resources, data curation, writing—review and editing, and visualization. NE-R: methodology, investigation, resources, data curation, and writing—review and editing. VB: methodology, resources, and writing—review and editing. IN: methodology, writing—review and editing, resources, and project administration. RA: methodology, resources, funding acquisition, visualization, and writing—review and editing. RAV: conceptualization, methodology, resources, data curation, visualization, writing—review and editing, and supervision.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no conflicts of interest to declare that are relevant to the content of this article.
Financial interests
Daniela Rojas-Cañizales and Isabel Naranjo received research support from Riester Foundation. Carmen Mejías-Balsalobre, Nínive Espinoza-Rodriguez, Vanessa Bézy, Randall Arauz and Roldán A Valverde declare they have no financial interests.
Ethical approval
This study has been approved by MINAE, SINAC, and ACT of Costa Rica, research permits [ACT-OR-DR-055-2020; ACT-OR-DR-063-2021].
Consent to participate
The authors declare that they participated in this research.
Consent for publication
The authors agree and give their consent for the publication of this study.
Additional information
Responsible Editor: P. Casale.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Rojas-Cañizales, D., Mejías-Balsalobre, C., Espinoza-Rodríguez, N. et al. Corozalito: a nascent arribada nesting beach in Costa Rica. Mar Biol 169, 59 (2022). https://doi.org/10.1007/s00227-022-04039-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00227-022-04039-6