
Abstract
The purpose of the study was to investigate the influence of contraction history on force steadiness and the associated EMG activity during submaximal isometric contractions performed with the dorsiflexor muscles. The key feature of the protocol was a triangular ramp contraction performed in the middle of a steady contraction at a lower target force. The target force during the ramp contraction was 20% MVC greater than that during the steady contraction. Thirty-seven healthy individuals (21 men and 16 women) performed the submaximal tasks with the ankle dorsiflexors. Electromyography (EMG) signals were recorded from tibialis anterior with a pair of surface electrodes. The coefficient of variation for force was significantly greater during the second steady contraction compared with the first one at each of the seven target forces (p < 0.015; d = 0.38–0.92). Although the average applied force during the steady contractions before and after the triangular contraction was the same (p = 0.563), the mean EMG amplitude for the steady contractions performed after the triangular contraction was significantly greater at each of the seven target forces (p < 0.0001; d = 0.44–0.68). Also, there were significant differences in mean EMG frequency between the steady contractions performed before and after the triangular contraction (p < 0.01; d = 0.13–0.82), except at 10 and 20% MVC force. The greater force fluctuations during a steady submaximal contraction after an intervening triangular contraction indicate a change in the discharge characteristics of the involved motor units.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
When performing a steady isometric contraction, the applied force fluctuates about an average value (Galganski et al. 1993; Laidlaw et al. 2000; Negro et al. 2016). The coefficient of variation for force—the normalized amplitude of these force fluctuations—provides a measure of force steadiness. Two features of the underlying motor unit activity can contribute to the amplitude of the force fluctuations: variability in the interspike intervals of individual motor units and the common low-frequency oscillations in discharge rate of the involved motor units (Moritz et al. 2005; Negro et al. 2009). Experimental measurements and computer simulations suggest that the fluctuations in force at low target forces is primarily attributable to the variability in interspike intervals, whereas the common low-frequency oscillations in discharge rate are responsible for the force fluctuations over most of the operating range of a muscle (Dideriksen et al. 2012; Farina and Negro 2015). However, experimental measures indicate marked overlap in the target forces at which the two measures of motor unit activity are significantly correlated with the fluctuations in force (Tsatsaki et al. 2022).
In addition to these discharge characteristics, force fluctuations can be modulated by the contraction history of the involved motor units. For example, evidence indicates that the discharge of action potentials by motor units can be maintained by persistent inward currents even after the termination of depolarizing synaptic input (Afsharipour et al. 2020; Gorassini et al. 1998; Heckman et al. 2008; Hounsgaard et al. 1984; Lee and Heckman 1998). The influence of this intrinsic motor neuron property on motor unit activity was demonstrated by Beauchamp et al. (2023) when they compared the discharge of motor units in the tibialis anterior and medial gastrocnemius muscles during steady contractions at 10% MVC force that were performed before and after an intervening triangular contraction to a greater target force. Many of the motor units recruited during the triangular contraction sustained discharge when performing the second steady contraction, which reduced the average discharge rate of the entire pool. These adjustments are expected to influence the coefficient of variation for force during the steady contractions, which is what we examined in our study.
The purpose of our study was to investigate the influence of contraction history on force steadiness and the associated EMG activity during submaximal isometric contractions performed with the dorsiflexor muscles. We used the protocol developed by Beauchamp et al. (2023) to compare the coefficient of variation for force and a global measure of muscle activation (mean amplitude and frequency of the surface EMG) during the steady contractions performed before and after a triangular contraction. As an extension of this protocol, we compared the effects for seven target forces performed with the dorsiflexor muscles. We hypothesized that the coefficient of variation for force (force steadiness) during the submaximal isometric contractions would increase after an intervening triangular ramp contraction.
Methods
Participants
This study was approved by the Institutional ethics committee (reference no. 2023-06/02) and conducted in accordance with the Declaration of Helsinki. Thirty-seven healthy individuals (21 men and 16 women; 22.8 ± 5.5 years, height 1.7 ± 0.1 m, body mass 70 ± 16 kg) were recruited for the study. Participants self-reported no history of neurological or musculoskeletal disorders. Possible benefits and risks of the study were explained prior to providing written informed consent. Participants were required to abstain from alcohol, caffeine, and high-intensity exercise for 24-h before their visit.
Experimental procedures
Participants were informed about the experimental protocol upon arrival at the laboratory. After being secured in an isokinetic dynamometer (IsoMed 2000, D&R, Ferstl, Hemau, Germany), participants practiced the experimental task. This involved a triangular ramp up-and-down contraction performed in the middle of several submaximal isometric contractions with the ankle dorsiflexors. Maximal voluntary contractions (MVC) were performed before and after the submaximal tasks (Fig. 1). EMG electrodes were attached to the skin over the tibialis anterior muscle.

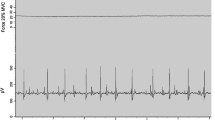
Experimental protocol. (a) The order of the measurements during the single visit. Maximal voluntary contractions (MVC) with the dorsiflexors were performed before and after the submaximal tasks. The primary task was a sombrero contraction, which involved a triangular ramp contraction performed in the middle of a steady contraction. The steady contractions were performed in a counterbalanced order at 7 target forces: 10, 20, 30, 40, 50, 60, and 70%. The target force during the ramp contraction was 20% greater than that for the steady contraction. The steady contractions each lasted ~10 s. (b) Participant position for ankle dorsiflexion tasks. (c) An example of the low-pass filtered force (black), smoothed EMG (blue), and interference EMG data collected for one participant. The steady contraction was performed at 40% MVC and the peak triangular force was 60% MVC. (d) As in c but for a different trial in which the two target forces were 10 and 30% MVC. Gray-shaded regions along the horizontal axis denote the ~10 s windows used for data analysis (Colour figure online)
The submaximal isometric contraction comprised a 5 s ramp up to the target force, a 10 s steady contraction, a 5 s ramp up to a target force that was 20% MVC force greater than the preceding steady contraction, a 5 s ramp back down to the lower target force, another 10 s steady contraction, and 5 s ramp down to baseline. The force trajectory during this task is known as a sombrero contraction (Beauchamp et al. 2023). The primary outcome variable was the coefficient of variation for force (force steadiness) during the steady contractions before and after the triangular contraction. When performing the tasks, visual feedback was provided on a computer screen placed in front of the participant. The vertical axis of the screen was proportional to the force registered by the isokinetic dynamometer. Participants were instructed to reach the target force displayed on the screen and maintain a steady contraction.
The isometric contractions were performed with the dorsiflexors at an ankle and knee angles of 90° (Fig. 1b). The seven target forces (Fig. 1a) for the steady contractions were set relative to the MVC force: 10, 20, 30, 40, 50, 60, and 70% MVC force. The steady contractions at the different target forces were performed in a counterbalanced order across participants, with 120 s of rest between each trial.
Two sets of MVCs (three trials each) for the ankle dorsiflexors were performed before and after the steady contractions. Each MVC trial lasted ~6 s and involved a 3 s increase from rest up to maximum that was held for 3 s. There was 2 min of rest between trials. Strong verbal encouragement was provided during each MVC trial.
EMG recordings
A wireless surface EMG system (Ultium EMG, Noraxon USA Inc., Scottsdale, AZ) was used to record muscle activity of tibialis anterior. The signals were sampled at 2000 Hz and band-pass filtered (10–500 Hz). The skin over tibialis anterior was shaved and cleaned with abrasive gel (Nuprep; Weaver and Company, Aurora, CO) and water. A pair of disposable Ag/AgCl self-adhesive gel electrodes (diameter of 1.3 cm, Noraxon Product No. 272; Noraxon USA Inc.) were attached to the skin with 2 cm between the centers of the two electrodes. The EMG and force signals were synchronized using an analog input system (16 channel BNC, Noraxon USA Inc. Scottsdale, AZ) and transferred to the same computer.
Data analysis
All data were processed in myoMuscle software (Noraxon USA Inc. Scottsdale, AZ) and MATLAB (2021b, MathWorks, Natick, MA). Mean EMG amplitude and frequency along with the coefficient of variation for force were calculated for the steady contractions performed before and after each triangular ramp contraction. The force signal was low-pass filtered (20 Hz, fourth-order, Butterworth). Subsequently, the coefficient of variation for force was calculated (standard deviation/mean × 100) for each 10 s steady contraction.
The root mean square (RMS, with 250-ms moving windows) value of the high-pass filtered (20 Hz, fourth-order, Butterworth) EMG signal was calculated. Mean EMG amplitude was quantified as the average of RMS value during each 10 s steady contraction and normalized to the maximal EMG amplitude during the initial MVC. The mean EMG frequency for the submaximal steady contractions was calculated across 1-s non-overlapping windows. Mean frequency was quantified as the frequency that corresponded to the peak value in the power density spectrum derived from the interference EMG signal.
Statistics
All statistical analyses were performed in R (version 4.3.1; R core team, Vienna, Austria) using rstatix library (version 0.7.2). The normality of the distributions and the equality of variances for the outcome variables were confirmed using the Shapiro–Wilk test and the Levene’s test, respectively. A two-way repeated-measures ANOVA was used to evaluate the differences in the coefficient of variation for force and the EMG measures across the target forces (10–70% MVC force) and the 10 s steady contractions (before and after the triangular contraction). Holm’s correction was applied to adjust p values for pairwise comparisons. MVCs performed before and after the submaximal tasks were compared using paired t tests.
Effect sizes (Cohen’s d) were calculated to indicate the magnitude of the difference in outcome variables between the steady contractions performed before and after the triangular contraction at each target force. The effect size thresholds were as follows: |d| < 0.2 negligible, |d| < 0.5 small, |d| < 0.8 medium and |d| ≥ 0.8 large (Cohen 1992).
Results
It seems unlikely that the protocol reduced the force capacity of the dorsiflexor muscles as each trial lasted approximately 40 s and was followed by a rest period of 120 s. Consistent with this expectation, MVC torque for the dorsiflexors after all the submaximal contractions (26.6 ± 8.0 Nm) was not statistically different (p = 0.46) from that at the start of the protocol (26.5 ± 8.0 Nm).
Examples of the force and EMG signals recorded during this experiment are plotted in Fig. 1. One notable feature of our observations was that many participants found it challenging to reduce the applied force back to the lower target force after the triangular contraction, especially at lower target forces. For example, the applied force shown in Fig. 1d fluctuated above the target force for approximately 3 s before settling on the target. We analyzed only the steady portions of the contractions at each target force (Fig. 1d, gray-shaded regions).
Decrease in force steadiness
The target force for the steady contractions performed before and after the triangular contraction was not statistically different (p = 0.563). However, the coefficient of variation for force was significantly greater during the second steady contraction compared with the preceding one at each of the 7 target forces (Fig. 2). The effect size for the difference in the coefficient of variation for force between the two steady contractions was greatest at the lowest (10%: d = 0.62) and three highest (50%: d = 0.90; 60%: d = 0.86; and 70%: d = 0.92) target forces (Table 1).

Amplitude of force fluctuations quantified as (a) coefficient of variation (CV) for force and (b) standard deviation (SD) for force during the 10 s submaximal steady contractions at the 7 target forces before (gray) and after (blue) the triangular ramp contraction. Data are mean ± standard error. *p < 0.05; **p < 0.01; ***p < 0.001 compared with the Before values (Colour figure online)
The main effect of contraction intensity on the coefficient of variation for force was also statistically significant. The coefficient of variation for force during the initial steady contraction (before) was significantly different for two comparisons: (1) 10% MVC vs. the other six target forces (p < 0.001), and (2) between 30 and 70% MVC (p = 0.025). Similarly, the coefficient of variation for force during the second steady contraction (after) was significantly greater at 10% MVC than at the other six target forces (p < 0.01). The other statistically significant differences in the coefficient of variation for force for the steady contractions performed after the triangular contraction included: (1) 70 vs. 50% (p = 0.019), 40% (p < 0.0001), and 30% MVC (p = 0.003); (2) 60 vs. 40% (p = 0.02) and 30% MVC (p = 0.032); and (3) 20 vs. 30% MVC (p = 0.03).
Increase in EMG amplitude
Mean EMG amplitude during the 10-s steady contractions performed after the triangular contraction was significantly greater than that during the initial steady contraction at each of the 7 target forces (p < 0.0001) (Fig. 3). The increase in EMG amplitude averaged 3.24 ± 1.2% MVC [95% CI: (2.16 4.33)] across the seven target forces. The elevated EMG amplitude, however, did not influence the applied force as this did not differ between the pairs of steady contractions (p = 0.563). The effect sizes for the differences in mean EMG amplitude between the pairs of steady contractions was moderate at 10% (d = 0.56), 20% (d = 0.6), 40% (d = 0.68), and 50% MVC (d = 0.63), whereas it was less at 30% (d = 0.48), 60% (d = 0.49), and 70% MVC (d = 0.44).

Mean EMG amplitude (%MVC) for tibialis anterior during the 10 s steady contractions performed before and after the triangular ramp contraction. Each dot indicates the EMG value for one participant with each target force (%MVC) indicated in a different color. Black lines show the average EMG amplitude at each target force. The difference in EMG amplitude before and after the ramp was significantly different at each target force (p < 0.0001)
Decrease in mean EMG frequency
Mean EMG frequency during the first steady contraction increased progressively with target force up to the second highest target force (60%). After the triangular contraction, however, the peak mean EMG frequency occurred at the 40% target force and the value for all subsequent target forces was less than that for the first steady contraction (Fig. 4). The difference in mean EMG frequency between the pairs of steady contractions was statistically significant at the target forces of 30% (p < 0.01; d = 0.13), 40, 50, 60, and 70% MVC (p < 0.001; d = 0.29–0.82). Also, the magnitude of the difference in mean EMG frequency between the two steady contractions increased with force level; for example, the difference was ~7 Hz at 40% MVC force (d = 0.29), whereas it was ~20 Hz at 70% MVC force (d = 0.82).

Mean EMG frequency (Hz) during the 10 s steady contractions at the 7 target forces before (gray) and after (blue) the triangular ramp contraction. Data are mean ± standard error. **p < 0.01; ***p < 0.001 compared with the Before values (Colour figure online)
Discussion
The main finding of the study was that the coefficient of variation for force, EMG amplitude for tibialis anterior, and the mean frequency of the interference EMG signal during the steady submaximal contraction were all altered by an intervening triangular contraction. Despite the mean applied force not differing during the first and second steady contractions, both the coefficient of variation for force and mean EMG amplitude were greater during the second steady contraction. Also, the mean EMG frequency did not increase progressively across target forces during the second steady contraction, as it did during the steady contractions performed before the triangular contraction.
Force steadiness
Although contraction history can influence muscle force and that of its constituent motor units (Burke et al. 1976; Gorassini et al. 1998; Proske et al. 2014; Rassier and Herzog 2004; Shi et al. 2023; Van Cutsem and Duchateau 2005), its impact on the normalized force fluctuations (force steadiness) largely reflects changes in the modulation of discharge rate (Dideriksen et al. 2012; Martinez-Valdes et al. 2022; Mazzo et al. 2022; Nagamori et al. 2021; Thompson et al. 2018). Our finding of an increase in the coefficient of variation for force during the second steady contraction replicates the result reported by Beauchamp et al. (2023). In their study, the sombrero contraction went from a steady contraction of 10% MVC torque to a peak triangular torque of 30% MVC, and the task was performed with both the plantar flexor and dorsiflexor muscles. In both studies, the average force was not statistically different between the first and second steady contractions. Our results extend those of Beauchamp et al. by demonstrating the decline in steadiness (coefficient of variation for force) was evident at all the target forces we examined, with the greatest effect at the lowest (10%) and four highest (40–70%) target forces.
Previous studies suggest that the greater fluctuations in force during the second steady contraction were attributable to either an increase in the coefficient of variation for interspike interval or an increase in the amplitude of the common low-frequency oscillations in discharge rate (Dideriksen et al. 2012; Farina and Negro 2015; Moritz et al. 2005; Negro et al. 2009). The significant adjustments in motor unit activity during the steady contraction performed after the triangular contraction are presumably responsible for the increase in the coefficient of variation for force (Beauchamp et al. 2023). This association needs to be examined explicitly. Moreover, it would be of interest to determine if the adjustments in discharge activity are accompanied by changes in the distribution of common synaptic input to the motor neuron pool as assessed by the identification of motor unit modes (Del Vecchio et al. 2023; Levine et al. 2023; Ricotta et al. 2023).
EMG amplitude
The second main finding was the greater EMG amplitude during the second steady contraction relative to the one performed before the triangular ramp contraction. The elevated EMG amplitude can be explained by the results of Beauchamp et al. (2023) who decomposed high-density EMG recordings into the discharge times of action potentials by many concurrently active motor units. They identified three groups of motor units: one group was activated during the first steady contraction, a second group was only active during the triangular ramp contraction, and a third group was recruited during the triangular contraction and remained active during the second steady contraction. This activity pattern was observed for motor units in tibialis anterior during dorsiflexion and in medial gastrocnemius during plantar flexion. The sustained discharge of the third group of motor units was attributed to the activation of persistent inward currents and was presumably responsible for the increase in EMG amplitude in our study.
Although statistically significant, the increase in EMG only averaged 3.24 ± 1.2% MVC across the seven target forces. This adjustment did not influence the average force during the second steady contraction, which did not differ from that during the first steady contraction. Nonetheless, the sustained discharge of the third group of motor units observed by Beauchamp et al. must have contributed to the increase in the coefficient of variation for force. Our results did not provide insight on this issue, but previous studies suggest that the greater fluctuations in force during the second steady contraction were attributable to either an increase in the coefficient of variation for interspike interval or an increase in the amplitude of the common low-frequency oscillations in discharge rate (Dideriksen et al. 2012; Farina and Negro 2015; Moritz et al. 2005; Negro et al. 2009).
As the final common pathway for the transformation of an activation signal into contractile force, motor unit activity establishes the quantity and quality of the resulting muscle force. Beauchamp et al. (2023) found that the target force during the second steady contraction was achieved with a reduction in the average discharge rate in combination with motor units that were recruited during the triangular contraction. Average discharge rate for tibialis anterior motor units was approximately 10 pps during the first steady contraction and this declined by 1.75 pps during the second plateau, both of which are rather low on the sigmoidal relation between discharge rate and force (Macefield et al. 1996). This decline in discharge rate was accompanied by the recruitment of an average of five motor units during the second steady contraction. Nonetheless, these adjustments were sufficient to achieve the target torque. What remains unknown, however, is those discharge characteristics that were responsible for the increase in the coefficient of variation for force.
Mean EMG frequency
The third main finding was a shift in the mean frequency for the interference EMG signal during the second steady contraction. Interpretation of changes in the power density spectrum is challenging (Farina et al. 2004). Although an interference EMG signal comprises the sum of muscle fiber action potentials (Dideriksen et al. 2011), its mean amplitude is relatively insensitive to modest changes in motor unit activity (Christie et al. 2009; Farina et al. 2014). Moreover, the spectral content of the interference EMG signal cannot detect either motor unit recruitment or average discharge rate (Del Vecchio et al. 2017; Dideriksen and Farina 2019; Farina et al. 2004), and the mean frequency is not associated with the amount of motor unit activity (Farina et al. 2014). Rather, the mean frequency is most influenced by the shape of the intracellular action potential and the impact of the volume conductor on the shape of motor unit action potentials (Boonstra and Breakspear 2012; Dideriksen et al. 2011; Farina et al. 2014).
Given the minimal demands of our protocol, the change we observed in mean frequency across the seven target forces likely reflect changes in the average shape of motor unit action potentials. Accordingly, the increase in mean frequency across target forces during the first steady contraction suggest a compression in the time domain of the average shape of the motor unit action potentials, presumably due to the recruitment of high-threshold motor units (Del Vecchio et al. 2017; Keenan et al. 2005; Milner-Brown and Stein 1975). In contrast, the profile differed during the second steady contraction with the mean frequency declining at the two highest target forces (60 and 70% MVC) despite a progressive increase in EMG amplitude. Potential explanations for this finding include the recruitment of motor units further away from the recording electrode or a change in the proportion of motor units that were recruited during the triangular contraction and sustained discharge into the second steady contraction. This issue needs to be resolved by recording the activity of motor units with grid electrodes during strong sombrero contractions.
Future directions
Our study has only explored one part of the contraction-history landscape and our results are not intended to generalize across conditions. For example, subsequent studies should examine the influence of joint angle, manipulate the features of the intervening triangular contraction, identify the motor unit discharge characteristics that are responsible for the reduction in force steadiness, and evaluate the functional consequences of the increase in force fluctuations during submaximal isometric contractions. This protocol, which was developed by Beauchamp et al. (2023), provides an opportunity to increase our knowledge of motor unit physiology.
In conclusion, control of force applied by the dorsiflexor muscles during a steady submaximal contraction was compromised by a prior ramp-up and ramp-down (triangular) isometric contraction. Although participants were able to match the seven target forces before and after the triangular contractions, the coefficient of variation for force and EMG amplitude for tibialis anterior were greater during the second steady contraction. Moreover, the mean frequency for the interference EMG signal suggested that there was a change in the involved motor units during the second steady contraction. This work, however, underscores the need for more studies on the influence of contraction history on motor unit physiology.
Data availability
Data will be made available by the authors upon reasonable request.
References
Afsharipour B, Manzur N, Duchcherer J, Fenrich KF, Thompson CK, Negro F, Quinlan KA, Bennett DJ, Gorassini MA (2020) Estimation of self-sustained activity produced by persistent inward currents using firing rate profiles of multiple motor units in humans. J Neurophysiol 124(1):63–85. https://doi.org/10.1152/jn.00194.2020
Beauchamp JA, Pearcey GEP, Khurram OU, Negro F, Dewald JPA, Heckman CJ (2023) Intrinsic properties of spinal motoneurons degrade ankle torque control in humans. bioRxiv 2023.10.23.563670. https://doi.org/10.1101/2023.10.23.563670
Boonstra TW, Breakspear M (2012) Neural mechanisms of intermuscular coherence: implications for the rectification of surface electromyography. J Neurophysiol 107:796–807. https://doi.org/10.1152/jn.00066.2011
Burke RE, Rudomin P, Zajac FE III (1976) The effect of activation history on tension production by individual muscle units. Brain Res 109:515–529
Christie A, Inglis JG, Kamen G, Gabriel DA (2009) Relationships between surface EMG variables and motor unit firing rates. Eur J Appl Physiol 107:177–185. https://doi.org/10.1007/s00421-009-1113-7
Cohen J (1992) A power primer. Psychol Bull 112:155–159
Del Vecchio A, Negro F, Felici F, Farina D (2017) Associations between motor unit action potential parameters and surface EMG features. J Appl Physiol 123(4):835–843. https://doi.org/10.1152/japplphysiol.00482.2017
Del Vecchio A, Germer CM, Kinfe TM, Nuccio S, Hug G, Eskofier B, Farina D, Enoka RM (2023) The forces generated by agonist muscles during isometric contractions arise from motor unit synergies. J Neurosci 43:2860–2873
Dideriksen JL, Farina D (2019) Amplitude cancellation influences the association between frequency components in the neural drive to muscle and the rectified EMG signal. PLoS Comput Biol 15(5):e1006985. https://doi.org/10.1371/journal.pcbi.1006985
Dideriksen JL, Enoka RM, Farina D (2011) Neuromuscular adjustments that constrain submaximal EMG amplitude at task failure of sustained isometric contractions. J Appl Physiol 111(2):485–494. https://doi.org/10.1152/japplphysiol.00186.2011
Dideriksen JL, Negro F, Enoka RM, Farina D (2012) Motor unit recruitment strategies and muscle properties determine the influence of synaptic noise on force steadiness. J Neurophysiol 107(12):3357–3369. https://doi.org/10.1152/jn.00938.2011
Farina D, Negro F (2015) Common synaptic input to motor neurons, motor unit synchronization, and force control. Exerc Sport Sci Rev 43(1):23–33. https://doi.org/10.1249/JES.0000000000000032
Farina D, Merletti R, Enoka RM (2004) The extraction of neural strategies from the surface EMG. J Appl Physiol 96(4):1486–1495. https://doi.org/10.1152/japplphysiol.01070.2003
Farina D, Merletti R, Enoka RM (2014) The extraction of neural strategies from the surface EMG: an update. J Appl Physiol 117(11):1215–1230. https://doi.org/10.1152/japplphysiol.00162.2014
Galganski ME, Fuglevand AJ, Enoka RM (1993) Reduced control of motor output in a human hand muscle of elderly subjects during submaximal contractions. J Neurophysiol 69(6):2108–2115. https://doi.org/10.1152/jn.1993.69.6.2108
Gorassini MA, Bennett DJ, Yang JF (1998) Self-sustained firing of human motor units. Neurosci Lett 247(1):13–16. https://doi.org/10.1016/s0304-3940(98)00277-8
Heckman CJ, Johnson M, Mottram C, Schuster J (2008) Persistent inward currents in spinal motoneurons and their influence on human motoneuron firing patterns. Neuroscientist 14(3):264–275. https://doi.org/10.1177/1073858408314986
Hounsgaard J, Hultborn H, Jespersen H, Kiehn O (1984) Intrinsic membrane properties causing a bistable behaviour of α-motoneurons. Exp Brain Res 55:391–394. https://doi.org/10.1007/BF00237290
Keenan KG, Farina D, Maluf KS, Merletti R, Enoka RM (2005) Influence of amplitude cancellation on the simulated surface electromyogram. J Appl Physiol 98:120–131. https://doi.org/10.1152/japplphysiol.00894.2004
Laidlaw DH, Bilodeau M, Enoka RM (2000) Steadiness is reduced and motor unit discharge is more variable in old adults. Muscle Nerve 23:600–612. https://doi.org/10.1002/(SICI)1097-4598(200004)23:4%3c600:AID-MUS20%3e3.0.CO;2-D
Lee RH, Heckman CJ (1998) Bistability in spinal motoneurons in vivo: systematic variations in persistent inward currents. J Neurophysiol 80:583–593. https://doi.org/10.1152/jn.1998.80.2.583
Levine J, Avrillon S, Farina D, Hug F, Pons JL (2023) Two motor neuron synergies, invariant across ankle joint angles, activate the triceps surae during plantarflexion. J Physiol 601:4337–4354
Macefield VG, Fuglevand AJ, Bigland-Ritchie B (1996) Contractile properties of single motor units in human toe extensors assessed by intraneural motor axon stimulation. J Neurophysiol 75:2509–2519. https://doi.org/10.1152/jn.1996.75.6.2509
Martinez-Valdes E, Negro F, Botter A, Pincheira PA, Cerone GL, Falla D, Lichtwark GA, Cresswell AG (2022) Modulations in motor unit discharge are related to changes in fascicle length during isometric contractions. J Appl Physiol 133:1136–1148
Mazzo MR, Holobar A, Enoka RM (2022) Association between effective neural drive to the triceps surae and fluctuations in plantar-flexor torque during submaximal isometric contractions. Exp Physiol 107:489–507
Milner-Brown HS, Stein RB (1975) The relation between the surface electromyogram and muscular force. J Physiol 246:549–569. https://doi.org/10.1113/jphysiol.1975.sp010904
Moritz CT, Barry BK, Pascoe MA, Enoka RM (2005) Discharge rate variability influences the variation in force fluctuations across the working range of a hand muscle. J Neurophysiol 93(5):2449–2459. https://doi.org/10.1152/jn.01122.2004
Nagamori A, Laine CM, Loeb GE, Valero-Cuevas FJ (2021) Force variability is mostly not motor noise: theoretical implications for motor control. PLoS Comput Biol 17:e1008787
Negro F, Holobar A, Farina D (2009) Fluctuations in isometric muscle force can be described by one linear projection of low-frequency components of motor unit discharge rates. J Physiol 587:5925–5938. https://doi.org/10.1113/jphysiol.2009.178509
Negro F, Yavuz UŞ, Farina D (2016) The human motor neuron pools receive a dominant slow-varying common synaptic input. J Physiol 594(19):5491–5505. https://doi.org/10.1113/JP271748
Proske U, Tsay A, Allen T (2014) Muscle thixotropy as a tool in the study of proprioception. Exp Brain Res 232:3397–3412
Rassier DE, Herzog W (2004) Consideration on the history dependence of muscle contraction. J Appl Physiol 96:419–427
Ricotta JM, Nardon M, De SD, Jiang J, Graziani W, Latash ML (2023) Motor unit-based synergies in a non-compartmentalized muscle. Exp Br Res 241:1367–1379
Shi F, Rymer WZ, Son J (2023) Ankle joint angle influences relative torque fluctuation during isometric plantar flexion. Bioengineering 10:373
Thompson CK, Negro F, Johnson MD, Holmes MR, McPherson LM, Powers RK, Farain D, Heckman CJ (2018) Robust and accurate decoding of motoneuron behaviour and prediction of the resulting force output. J Physiol 596:2643–2659
Tsatsaki E, Amiridis IG, Holobar A, Trypidakis G, Arabatzi F, Kellis E, Enoka RM (2022) The length of tibialis anterior does not influence force steadiness during submaximal isometric contractions with the dorsiflexors. Eur J Sport Sci 22(4):539–548. https://doi.org/10.1080/17461391.2021.1922506
Van Cutsem M, Duchateau J (2005) Preceding muscle activity influences motor unit discharge and rate of torque development during ballistic contractions in humans. J Physiol 562:635–644. https://doi.org/10.1113/jphysiol.2004.074567
Acknowledgements
We thank the volunteers who participated in our study and the members of our laboratory for helpful discussions on this project. R.M.E. was supported by an award from the National Multiple Sclerosis Society in the USA (project RG-2206-39688).
Funding
Open access funding provided by Northeastern University Library. National Multiple Sclerosis Society, RG-2206-39688, Roger Enoka.
Author information
Authors and Affiliations
Contributions
AD and RME contributed to the conception and design of research, interpretation of the results, and the preparation of the manuscript. AD performed experiments and analyzed data. AD prepared figures. Both authors have read and approved the final version of the manuscript and agree to be held accountable for all aspects of the work in ensuring that questions related to the accuracy or integrity of any part of the work are appropriately investigated and resolved. All persons designated as authors qualify for authorship, and all those who qualify for authorship are listed.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Communicated by Winston D Byblow.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Darendeli, A., Enoka, R.M. Control of motor output during steady submaximal contractions is modulated by contraction history. Exp Brain Res 242, 675–683 (2024). https://doi.org/10.1007/s00221-023-06774-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-023-06774-8