
Abstract
Numerous studies in airways, ileum, and urinary bladder have demonstrated that relaxation by β-adrenoceptor agonists has lower potency and/or efficacy when contraction was elicited by muscarinic receptor agonists as compared to other G-protein-coupled receptors, KCl, or basal tone, but the molecular mechanisms behind this relative resistance remain unclear. A paper by Huang et al. in this issue demonstrates that NAV2729, an inhibitor of ADP ribosylation factor 6, inhibits contraction of isolated blood vessels elicited by muscarinic receptor agonists, but not by α1-adrenoceptor agonists or KCl. Against this background, we discuss the role of ADP ribosylation factor 6 in cellular responses to G-protein-coupled receptor stimulation. While ADP ribosylation factor 6 apparently is the only promising molecular explanation for the relative resistance of smooth muscle contraction elicited by muscarinic agonists, the existing data are insufficient for a robust conclusion.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Muscarinic receptors are important mediators of smooth muscle contraction in various tissues including airways, gut, and urinary bladder; this largely involves the M3 subtype but M2 receptors can also affect smooth muscle tone (Hegde and Eglen 1999). M3 receptors typically couple to G-proteins of the Gq/11 type leading to activation of a phospholipase C (PLC) (Caulfield and Birdsall 1998). Surprisingly, M3-mediated smooth muscle contraction is not explained by PLC activation, for instance in the bladder (Frazier et al. 2008), but alternative molecular mechanisms to elicit smooth muscle contraction have not been well-defined.
The potency and/or efficacy of β-adrenoceptor agonists to relax smooth muscle is lower when tested against muscarinic agonists such as carbachol than against agonists at other receptors, against passive tension or against receptor-independent contraction elicited by KCl (Dale et al. 2014). Such observations have been made in airways (Russel 1984; Raffestin et al. 1985; Ostrom and Ehlert 1999; Sarria et al. 2002; Naline et al. 2007), ileum (Ostrom and Ehlert 1999), and bladder (Longhurst and Levendusky 1999; Witte et al. 2011; Kanie et al. 2012) of multiple species including rat (Longhurst and Levendusky 1999; Michel and Sand 2009; Witte et al. 2011; Cernecka et al. 2014), guinea pig (Ostrom and Ehlert 1999), dog (Russel 1984), and human (Raffestin et al. 1985; Sarria et al. 2002; Naline et al. 2007; Kanie et al. 2012) and with agonists acting at histamine (Russel 1984; Raffestin et al. 1985; Ostrom and Ehlert 1999; Naline et al. 2007), 5-hydroxytryptamine (5-HT) (Michel and Sand 2009; Cernecka et al. 2014), bradykinin (Michel and Sand 2009; Cernecka et al. 2014), prostanoid receptors (Sarria et al. 2002), passive tone (Naline et al. 2007; Michel and Sand 2009; Cernecka et al. 2014), or KCl (Longhurst and Levendusky 1999; Michel and Sand 2009; Kanie et al. 2012). While most of the above studies have used general β-adrenoceptor agonists such as isoprenaline, similar findings have also been obtained with agonists selective for β2-adrenoceptors such as formoterol, indacaterol, salbutamol, and salmeterol in human bronchi (Naline et al. 2007) or fenoterol in rat bladder (Erdogan et al. 2021) or selective for β3-adrenoceptors such as TRK 380 in human bladder (Kanie et al. 2012) or KUC 7322 in rat bladder (Cernecka et al. 2014). However, it has remained elusive why smooth muscle contraction elicited by muscarinic receptor agonists is more resistant to relaxation by β-adrenoceptor agonists than that elicited by other stimuli. It appears logical that such selective resistance should be related to a signaling pathway activated preferentially by muscarinic receptors as compared to other receptors, but we are not aware of such signaling pathways.
A paper in this issue of the journal demonstrates that NAV2729, an inhibitor of ADP ribosylation factor 6 (ARF6), inhibits coronary vascular smooth muscle contraction elicited by the muscarinic agonists carbachol and methacholine but not that elicited by agonists at other receptors including α1-adrenoceptors, 5-HT, endothelin-1, or prostanoid TP receptors or those elicited by KCl (Huang et al. 2022). The same group had previously shown the existence of ARF6 expression human prostate smooth muscle tissue (Hennenberg et al. 2013) and that NAV2729 inhibited contraction in isolated human prostate strips by α1-adrenoceptor agonists, whereas that elicited by a prostanoid TP receptor agonist, endothelin-1 or by KCl, was not inhibited (Yu et al. 2019). Furthermore, NAV2729 inhibited contraction in human prostate smooth muscle cells, and this was also observed in ARF6 knockout cells (Wang et al. 2021). Inhibition of ARF6 by NAV2729 inhibited contraction elicited by α1-adrenoceptor agonists in human prostate (Yu et al. 2019) but not porcine blood vessels (Huang et al. 2022). Interestingly, the inhibition of contraction in human prostate was accompanied by an inhibition of ARF6 in pull-down assays, while ARF6 was not activated by noradrenaline, phenylephrine, or methoxamine (Yu et al. 2019). The selective inhibition of muscarinic receptor responses in the blood vessels (Huang et al. 2022) raises the possibility that coupling to ARF6 may be a mechanism that distinguishes inhibition by β-adrenoceptor agonists of responses to a muscarinic agonist as compared to those elicited by other means.
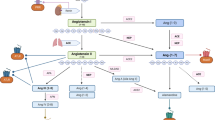
ARF6 is a small GTP-binding protein that contributes to several cellular processes including G-protein-coupled receptor (GPCR) trafficking, actin organization, and contractile response through diverse downstream component interaction (Fig. 1, Table 1). ARF6 function is modulated by two distinct components. Guanine nucleotide exchange factors (GEFs) mediate the activation of ARF6, whereas GTPase-activating proteins (GAPs) mediate inhibition. NAV2729 interferes in the formation of ARF6-GEF complex to inhibit the ARF6 activation (Yamauchi et al. 2017).

Schematic drawing of ARF6-mediated cellular effects in GPCRs agonist stimulation. Dashed red arrow, negative regulatory effect; black arrow, positive regulatory effect; GAP, GTPase-activating proteins; GDP, guanosine diphosphate; GEF, guanine nucleotide exchange factors; GTP, guanosine triphosphate; ↑, increase; ↓, decrease; ↔ , no effect
ARF6 was shown to be a prerequisite component further promoting either clathrin- or caveolin-mediated pathway in agonist-induced endocytosis of several GPCRs such as β-adrenoceptor in adipocytes (Liu et al. 2010) and β2-adrenoceptor in HEK293 cells (Claing et al. 2001; Lawrence et al. 2005; Macia et al. 2012); M2 muscarinic receptors in HeLa cells (Delaney et al. 2002), in JEG-3 human choriocarcinoma cells (Reiner and Nathanson 2008), and in HEK293 cells (Houndolo et al. 2005); the luteinizing hormone chorionic gonadotropin receptor in HEK 293 cells (Kanamarlapudi et al. 2012); angiotensin type 1 receptor (Houndolo et al. 2005; Cotton et al. 2007); μ-opioid receptor (Rankovic et al. 2009); the vasopressin V2 receptor; and endothelin type B receptor (Houndolo et al. 2005) in HEK293 cells. Upon agonist stimulation, GPCR endocytosis was found mostly activated via ARF6-dependent pathway with some exceptions which show inhibitor regulatory effect of ARF6 in M2 muscarinic receptor (Delaney et al. 2002; Reiner and Nathanson 2008) and in β2-adrenoceptor (Macia et al. 2012) internalization. Furthermore, VIP receptor internalization was not affected by ARF6 depletion (Houndolo et al. 2005). ARF6 involvement in trafficking did not exist for some receptors such as M4 muscarinic receptor (Reiner and Nathanson 2008) and vasopressin V2 receptor (Madziva and Birnbaumer 2006). ARF6 requirement in endocytosis was mostly demonstrated in agonist-induced settings, which may not reflect the ARF6 function for basal condition for the same receptor (Cotton et al. 2007). Moreover, ARF6 involvement of muscarinic receptor internalization has mostly been studied with the M2 subtype because of the well-defined, clathrin-dependent pathway-mediated internalization of M1, M3, and M4 receptors (Reiner and Nathanson 2008).
Phospholipase D (PLD) is known to be involved in smooth muscle contraction through PKC activation but its contribution in urinary bladder contraction was proposed to be minor (Frazier et al. 2008). ARF6-mediated PLD activation was reported by several researchers in in vivo animal (Le Stunff et al. 2000) and in vitro cultured cell line studies (Mitchell et al. 2003; Johnson et al. 2006; Rankovic et al. 2009; Chakraborti et al. 2017; Charles et al. 2018). In human pulmonary artery smooth muscle cells (HPASMCs), stimulation of prostanoid TP receptor stimulates cytohesin-1 coupling to ARF6 which further leads to PLD2 isoform and subsequent NADPH oxidase activation (Chakraborti et al. 2017). In HPASMCs, involvement of ARF6 in endothelin-1-induced PLD and NADPH oxidase activation was shown by same study group (Chakraborti et al. 2018). ARF6 was shown to be involved in PLD activation in N376D mutant 5-HT2A-stimulated PLD activation but not in mediated via WT 5-HT2A stimulation (Johnson et al. 2006). This study also showed the sensitivity of other class A GPCRs which contain DPxxY motif such as purinergic P2u, thrombin PAR, and gonadotropin-releasing hormone receptor to ARF6 for further PLD activation in COS7 cells (Johnson et al. 2006). In the same cell line, agonist-stimulated M3 activation induced both PLD1/2 activation through ARF6-mediated pathway, whereas PLD2 activation was found linked to PKC and ARF6 in purinergic P2U receptor and to only PKC in N376D mutant 5-HT2a receptor (Mitchell et al. 2003). ARF6 is involved in the promotion of prostate smooth muscle contraction. Inhibition of ARF6 activation by cytohesin (a GEF) inhibitor resulted in reduced noradrenaline, phenylephrine-, thromboxane A2-, and endothelin-1- and endothelin-3-induced contraction (Herlemann et al. 2018). In vascular smooth muscle cell, both ARF1 and ARF6 are involved in actin polymerization which subsequently migrate and proliferate but only ARF1 affected contractile responses (Charles et al. 2018). However, in a latter study, ARF6 was found to promote contraction and proliferation in human prostate stromal cells (WPMY-1) (Wang et al. 2021).
ARF6 activation interrupts the recycling of β2-adrenoceptors and lead desensitization of receptor in HEK293 cells (Macia et al. 2012); similarly, it is postulated that ARF6 may involve in α1-adrenoceptor desensitization in human prostate tissue (Hennenberg et al. 2013). In contrast, ARF6-mediated endocytosis was suggested beneficial in development of opioid tolerance through preventing receptor desensitization HEK293 cells (Rankovic et al. 2009).
Other effects of ARF6 include the calcium-sensing receptor-mediated plasma membrane ruffling which is required for chemotaxis in HEK cells (Bouschet et al. 2007). In endothelial cells, endothelin ETB receptor stimulation by endothelin 1 results in ARF6 activation which facilitates cell migration via actin reorganization. Moreover, endothelin 1 stimulation did not promote capillary tube formation in ARF6 depleted cells which indicates ARF6 involvement in angiogenesis (Daher et al. 2008). ARF6 also has regulatory effects on metabolic pathway. Depletion of ARF6 resulted in inhibition of isoproterenol-induced lipolysis in 3T3-L1 adipocytes. ARF6 mRNA and protein level was found higher in WAT tissue of ob/ob mice compared to obesity resistance mice (Liu et al. 2010). In 3T3-L1 adipocytes, endothelin ETA receptor-mediated lipolysis was found to be dependent downstream ARF6-ERK1/2 signaling (Davies et al. 2014). Additionally, endothelin 1 stimulated GLUT4 translocation through heterotrimetric Gq protein signaling pathway activated by ARF6 in 3T3-L1 adipocytes (Bose et al. 2001; Lawrence and Birnbaum 2001) and specifically Gα11 isoform of Gαq family.
Taken together, the above data demonstrate that ARF6 is involved in cellular processes and is activated by cellular effects stimulated by various GPCR, including smooth muscle contraction. Within a given tissue, the involvement of ARF6 in pathways leading to smooth muscle contraction, e.g., in arteries or prostate, appears to be preferential for some GPCR (muscarinic receptors and α1-adrenoceptors, respectively) over others apparently coupling to the same G-proteins. Some of these data are in line with the hypothesis that coupling to ARF6 may explain the relative resistance of smooth muscle contraction elicited by muscarinic receptors as compared to other GPCR or receptor-independent contraction such as receptor desensitization to relaxation by β-adrenoceptor agonists. However, not all data support this hypothesis. Thus, the role of ARF6 in this phenomenon cannot be considered proven, but it remains as a reasonable molecular candidate to explain the resistance of muscarinic receptor-mediated smooth muscle contraction against relaxation. Further studies at the molecular level are required to further explore this, specifically studies in which the role of agonists at various GPCR is compared quantitatively.
References
Bose A, Cherniack AD, Langille SE, Nicoloro SM, Buxton JM, Park JG, Chawla A, Czech MP (2001) Gα11 signaling through ARF6 regulates F-actin mobilization and GLUT4 glucose transporter translocation to the plasma membrane. Mol Cell Biol 21:5262–5275
Bouschet T, Martin S, Kanamarlapudi V, Mundell S, Henley JM (2007) The calcium-sensing receptor changes cell shape via a β-arrestin-1–ARNO–ARF6–ELMO protein network. J Cell Sci 120:2489–2497
Caulfield MP, Birdsall NJM (1998) International Union of Pharmacology. XVII. Classification of muscarinic acetylcholine receptors. Pharmacol Rev 50:279–290
Cernecka H, Sand C, Michel MC (2014) The odd sibling: features of ß3-adrenoceptor pharmacology. Mol Pharmacol 86:479–484
Chakraborti S, Sarkar J, Chowdhury A, Chakraborti T (2017) Role of ADP ribosylation factor6− cytohesin1− phospholipase D signaling axis in U46619 induced activation of NADPH oxidase in pulmonary artery smooth muscle cell membrane. Arch Biochem Biophys 633:1–14
Chakraborti S, Sarkar J, Bhuyan R, Chakraborti T (2018) Role of catechins on ET-1-induced stimulation of PLD and NADPH oxidase activities in pulmonary smooth muscle cells: determination of the probable mechanism by molecular docking studies. Biochem Cell Biol 96:417–432
Charles R, Bourmoum M, Claing A (2018) ARF GTPases control phenotypic switching of vascular smooth muscle cells through the regulation of actin function and actin dependent gene expression. Cell Signal 46:64–75
Claing A, Chen W, Miller WE, Vitale N, Moss J, Premont RT, Lefkowitz RJ (2001) β-Arrestin-mediated ADP-ribosylation factor 6 activation and β2-adrenergic receptor endocytosis. J Biol Chem 276:42509–42513
Cotton M, Boulay P-L, Houndolo T, Vitale N, Pitcher JA, Claing A (2007) Endogenous ARF6 interacts with Rac1 upon angiotensin II stimulation to regulate membrane ruffling and cell migration. Mol Biol Cell 18:501–511
Daher Z, Noël J, Claing A (2008) Endothelin-1 promotes migration of endothelial cells through the activation of ARF6 and the regulation of FAK activity. Cell Signal 20:2256–2265
Dale PR, Cernecka H, Schmidt M, Dowing M, Charlton SJ, Michel MC (2014) The pharmacological rationale for combining muscarinic receptor antagonists and ß-adrenoceptor agonists in the treatment of airway and bladder disease. Curr Opin Pharmacol 16C:31–42
Davies JC, Bain SC, Kanamarlapudi V (2014) ADP-ribosylation factor 6 regulates endothelin-1-induced lipolysis in adipocytes. Biochem Pharmacol 90:406–413
Delaney KA, Murph MM, Brown LM, Radhakrishna H (2002) Transfer of M2 muscarinic acetylcholine receptors to clathrin-derived early endosomes following clathrin-independent endocytosis. J Biol Chem 277:33439–33446
Erdogan BR, Yesilyurt ZE, Arioglu-Inan E, Michel MC (2021) Validation of fenoterol to study β2-adrenoceptor function in the rat urinary bladder. Pharmacology 107:116–121
Frazier EP, Peters SLM, Braverman AS, Ruggieri MR Sr, Michel MC (2008) Signal transduction underlying control of urinary bladder smooth muscle tone by muscarinic receptors and ß-adrenoceptors. Naunyn-Schmiedebergs Archives of Pharmacology 377:449–462
Hegde SS, Eglen RM (1999) Muscarinic receptor subtypes modulating smooth muscle contractility in the urinary bladder. Life Sci 64:419–428
Hennenberg M, Miersch J, Rutz B, Strittmatter F, Waidelich R, Stief CG, Gratzke C (2013) Noradrenaline induces binding of Clathrin light chain A to α1-adrenoceptors in the human prostate. Prostate 73:715–723
Herlemann A, Keller P, Schott M, Tamalunas A, Ciotkowska A, Rutz B, Wang Y, Yu Q, Waidelich R, Strittmatter F (2018) Inhibition of smooth muscle contraction and ARF6 activity by the inhibitor for cytohesin GEFs, secinH3, in the human prostate. American Journal of Physiology-Renal Physiology 314:F47–F57
Houndolo T, Boulay P-L, Claing A (2005) G protein-coupled receptor endocytosis in ADP-ribosylation factor 6-depleted cells. J Biol Chem 280:5598–5604
Huang R, Li B, Tamalunas A, Waidelich R, Stief CG, Hennenberg M (2022) Inhibition of neurogenic contractions in renal arteries and of cholinergic contractions in coronary arteries by the presumed inhibitor of ADP-ribsolytion factor 6, NAV2729. Naunyn Schmiedeberg’s Arch Pharmacol. https://doi.org/10.1007/s00210-022-02218-2
Johnson MS, Robertson DN, Holland PJ, Lutz EM, Mitchell R (2006) Role of the conserved NPxxY motif of the 5-HT2A receptor in determining selective interaction with isoforms of ADP-ribosylation factor (ARF). Cell Signal 18:1793–1800
Kanamarlapudi V, Thompson A, Kelly E, Bernal AL (2012) ARF6 activated by the LHCG receptor through the cytohesin family of guanine nucleotide exchange factors mediates the receptor internalization and signaling. J Biol Chem 287:20443–20455
Kanie S, Otsuka A, Yoshikawa S, Morimoto T, Hareyama N, Okazaki S, Kobayashi R, Hasebe K, Nakao K, Hayashi R, Mochizuki H, Matsumoto R, Ozono S (2012) Pharmacological effect of TRK-380, a novel selective human ß3-adrenoceptor agonist, on mammalian detrusor strips. Urology 79:744.e741-744.e747
Lawrence JTR, Birnbaum MJ (2001) ADP-ribosylation factor 6 delineates separate pathways used by endothelin 1 and insulin for stimulating glucose uptake in 3T3-L1 adipocytes. Mol Cell Biol 21:5276–5285
Lawrence J, Mundell SJ, Yun H, Kelly E, Venkateswarlu K (2005) Centaurin-α1, an ADP-ribosylation factor 6 GTPase activating protein, inhibits β2-adrenoceptor internalization. Mol Pharmacol 67:1822–1828
Le Stunff H, Dokhac L, Bourgoin S, Bader M-F, Harbon S (2000) Phospholipase D in rat myometrium: occurrence of a membrane-bound ARF6 (ADP-ribosylation factor 6)-regulated activity controlled by βγ subunits of heterotrimeric G-proteins. Biochemical Journal 352:491–499
Liu Y, Zhou D, Abumrad NA, Su X (2010) ADP-ribosylation factor 6 modulates adrenergic stimulated lipolysis in adipocytes. Am J Physiol Cell Physiol 298:C921–C928
Longhurst PA, Levendusky M (1999) Pharmacological characterization of ß-adrenoceptors mediating relaxation of the rat urinary bladder in vitro. Br J Pharmacol 127:1744–1750
Macia E, Partisani M, Paleotti O, Luton F, Franco M (2012) Arf6 negatively controls the rapid recycling of the β2 adrenergic receptor. J Cell Sci 125:4026–4035
Madziva MT, Birnbaumer M (2006) A role for ADP-ribosylation factor 6 in the processing of G-protein-coupled receptors. J Biol Chem 281:12178–12186
Michel MC, Sand C (2009) Effect of pre-contraction on ß-adrenoceptor-mediated relaxation of rat urinary bladder. World J Urol 27:711–715
Mitchell R, Robertson DN, Holland PJ, Collins D, Lutz EM, Johnson MS (2003) ADP-ribosylation factor-dependent phospholipase D activation by the M3 muscarinic receptor. J Biol Chem 278:33818–33830
Naline E, Trifilieff A, Fairhurst RA, Advenier C, Molimard M (2007) Effect of indacaterol, a novel long-acting ß2-agonist, on isolated human bronchi. Eur Respir J 29:575–581
Ostrom RS, Ehlert FJ (1999) Cross-functional antagonism between isoproterenol and M 2 muscarinic receptors in guinea pig ileum and trachea. J Pharmacol Exp Ther 288:969–976
Raffestin B, Cerrina J, Boullet C, Labat C, Benveniste J, Brink C (1985) Response and sensitivity of isolated human pulmonary muscle preparations to pharmacological agents. J Pharmacol Exp Ther 233:186–194
Rankovic M, Jacob L, Rankovic V, Brandenburg L-O, Schröder H, Höllt V, Koch T (2009) ADP-ribosylation factor 6 regulates mu-opioid receptor trafficking and signaling via activation of phospholipase D2. Cell Signal 21:1784–1793
Reiner C, Nathanson NM (2008) The internalization of the M2 and M4 muscarinic acetylcholine receptors involves distinct subsets of small G-proteins. Life Sci 82:718–727
Russel JA (1984) Differential inhibitory effect of isoproterenol on contractions of canine airways. J Appl Physiol 57:801–807
Sarria B, Naline E, Zhang Y, Cortijo J, Molimard M, Moreau J, Therond P, Advenier C, Morcillo EJ (2002) Muscarinic M2 receptors in acetylcholine-isoproterenol functional antagonism in human isolated bronchus. Am J Physiol 283:L1125–L1132
Wang R, Schneider S, Keppler OT, Li B, Rutz B, Ciotkowska A, Stief CG, Hennenberg M (2021) ADP ribosylation factor 6 promotes contraction and proliferation, suppresses apoptosis and Is specifically Inhibited by NAV2729 in prostate stromal cells. Mol Pharmacol 100:356–371
Witte LPW, de Haas N, Mammen M, Stangeland EL, Steinfeld T, Aiyar J, Michel MC (2011) Muscarinic receptor subtypes and signalling involved in the attenuation of isoprenaline-induced rat urinary bladder relaxation. Naunyn Schmiedebergs Arch Pharmacol 384:555–563
Yamauchi Y, Miura Y, Kanaho Y (2017) Machineries regulating the activity of the small GTPase Arf6 in cancer cells are potential targets for developing innovative anti-cancer drugs. Advances in Biological Regulation 63:115–121
Yu Q, Gratzke C, Wang R, Li B, Kuppermann P, Herlemann A, Tamalunas A, Wang Y, Rutz B, Ciotkowska A, Wang X, Strittmatter F, Waidelich R, Stief CG, Hennenberg M (2019) A NAV2729-sensitive mechanism promotes adrenergic smooth muscle contraction and growth of stromal cells in the human prostate. J Biol Chem 294:12231–12249
Acknowledgements
The figure was generated by using Servier Medical Art by Servier licensed under a Creative Commons Attribution 3.0 unported license. http://smart.servier.com.
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Contributions
MCM conceptualized the idea. BRE performed the literature search. Both BRE and MCM drafted and revised the manuscript and approved the final version.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Erdogan, B.R., Michel, M.C. Does coupling to ADP ribosylation factor 6 explain differences between muscarinic and other receptors in interaction with β-adrenoceptor-mediated smooth muscle relaxation?. Naunyn-Schmiedeberg's Arch Pharmacol 395, 381–386 (2022). https://doi.org/10.1007/s00210-022-02221-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00210-022-02221-7