
Abstract
Nonequilibrium ecology and resilience theory have transformed rangeland ecology and management by challenging the traditional assumptions of ecological stability and linear successional dynamics. These alternative interpretations indicate that ecosystem dynamics are strongly influenced by disturbance, heterogeneity, and existence of multiple stable states. The nonequilibrium persistent model indicates that plant production and livestock numbers are seldom in equilibrium in pastoral systems because reoccurring drought maintains livestock number below the ecological carrying capacity. However, it has recently been demonstrated that livestock are often in equilibrium with key dry-season resources, even though they may only be loosely coupled to abundant wet-season resources. Similarly, state-and-transition models were initially influenced by nonequilibrium ecology, but they have subsequently been organized around resilience theory to represent both equilibrial dynamics within states and existence of multiple states. Resilience theory was introduced to describe how ecosystems can be dynamic, but still persist as self-organized systems. It envisions that community structure is maintained by ecological processes representing feedback mechanisms and controlling variables to moderate community fluctuation in response to disturbance. Appropriate qualification of equilibrium ecology within resilience theory, rather than its complete replacement by nonequilibrium models, provides more realistic interpretations for both plant–herbivore interactions and vegetation dynamics than does complete reliance on disturbance-driven events. Resilience thinking represents a “humans-in-nature” perspective that emphasizes human values and goals and it seeks to guide change in social-ecological systems by creating opportunities for multiple stakeholders to adaptively design management strategies and policies.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others

Keywords
- Alternative stable states
- Multiple equilibria
- Rangeland ecology
- Resilience-based management
- Resilience thinking
- Social resilience
- Thresholds
1 Introduction
Humans interact with nature through the use of simplified and incomplete perceptions of its structure, interrelationships, and dynamics. These perceptions are based on experience, specific to place, and subject to change (Jones et al. 2011). They influence which problems are considered, how they are envisioned, and the potential solutions to address them (Lynam and Stafford Smith 2004). Consider the following questions regarding ecosystem dynamics. How stable are ecosystems? Do limits exist to ecosystem recovery following disturbance? What management actions are most likely to sustain desired ecosystems? A major shift in our perception of nature would greatly alter our responses to these questions, and the manner in which we interact with nature to promote sustainable ecosystem management and human well-being.
Nonequilibrium ecology and resilience theory represent such a change in the human perception of nature. Nonequilibrium ecology challenged the prevailing perception of ecosystem stability and rapid, linear recovery following natural or human disturbances. Equilibrium ecology is reflected in the “balance-of-nature ” metaphor and is exemplified by the controversial Gaia hypothesis which suggests that the Earth system is in part self-regulated to maintain conditions for life. Equilibrial ecology was initially challenged by theoretical evidence of nonlinear system dynamics in the mid-twentieth century and, thereafter, by inconsistencies in natural resource management outcomes (Holling 1973; Folke 2006).
Nonequilibrium ecology represents a more dynamic and less predictable perception of ecosystem dynamics that recognizes the contributions of disturbance, spatial heterogeneity, and multiple stable states, in addition to internal biotic regulation (Wu and Loucks 1995). It further challenges the prevailing model of natural resource management —the steady-state management model—that was founded upon equilibrium ecology. This management model emphasizes the maximum sustainable yield of one or a few resources through implementation of management actions to minimize variability and redundancy that may interfere with maximum sustainable production (Holling and Meffe 1996). Practices that optimize harvest efficiency and reduce diversity and heterogeneity—fire prevention, plant control measures, and planting of monocultures—are representative of this management approach that often relies on technological solutions to increase production and reduce uncertainty. It is now recognized that this management model along with the command-and-control management strategy—top-down regulation by a centralized authority—can destabilize the very ecosystems that they were intended to sustain (Holling and Meffe 1996). The adverse outcomes originating from these management approaches have been termed the “pathology of natural resource management .”
Resilience theory emerged in response to recognition that the prevailing concept of ecological stability was not a realistic interpretation of observed ecosystem dynamics. For example, ecosystems can exhibit wide fluctuations in species composition, but still be very resilient (Curtain and Parker 2014). This inconsistency was resolved by dividing stability into two components—resistance and resilience. Resistance describes the capacity of systems to remain unchanged by disturbance, while resilience is the capacity to return to a former configuration following disturbance (Holling 1973). Resilience also recognizes the existence of threshold conditions that contribute to the formation of alternative stable states. Grassland conversion to woodland and perennial shrub steppe conversion to annual grasslands are widely recognized examples of nonreversible dynamics that result in the formation of alternative ecosystems on the same site. Resilience-based management further provides an alternative to steady-state management that encourages managers to anticipate and guide the direction of change, rather to prevent change, so that ecosystems can sustainably provide ecosystem services to society (Holling 1973; Chapin et al. 2010).
Resilience theory has recently been extended to social systems to provide a “humans-in-nature” perspective to ecosystem management and policy. Adaptive management —learning by doing—and social learning—the capacity of groups of people to achieve goals—have emerged as essential components of resilient human-dominated systems. These resilience-based approaches are collectively termed “resilience thinking ” and they are intended to provide a path toward greater sustainability by embracing uncertainty, variability, and recognition of incomplete knowledge (Walker and Salt 2012; Curtain and Parker 2014).
The goals of this chapter are to provide a synopsis of the origins and development of nonequilibrium ecology and resilience theory and to describe how these concepts have influenced the ecology, management, and governance of rangeland systems. Specific objectives are to:
-
(1)
Summarize equilibrium and nonequilibrium ecology and resilience theory
-
(2)
Assess the consequences of these concepts to rangeland ecology and management
-
(3)
Explore the application and utility of resilience in social-ecological systems
-
(4)
Describe future perspectives regarding further integration of resilience in rangeland systems
2 Conceptual Advances
2.1 Equilibrium and Nonequilibrium Ecology
Equilibrium ecology and its associated metaphor, “the balance of nature ,” is an ancient human concept, but the modern foundation was derived from systems theory in the 1960s. It is founded on the assumption that ecosystems are highly self-regulated by internal biotic processes, including intra- and interspecific competition and plant–animal interactions that restrict their dynamics to a single stable state (Wu and Loucks 1995). It is further assumed that this state will return to its original pre-disturbance condition after a disturbance has ceased. The predictable and directional response of plant succession that passes through anticipated, sequential stages toward a single equilibrium point or stable state provides a well-known example (Pickett and Ostfeld 1995). Equilibrium ecology experienced growing criticism in the mid-twentieth century for several reasons, including (1) limited supporting evidence of equilibrium conditions in ecosystems, (2) an inability to account for the occurrence of alternative stable states in some ecosystems, and (3) slow or nonexistent recovery of alternative states when they had formed (Wu and Loucks 1995; Briske et al. 2003).
Nonequilibrium theory emerged from investigation of theoretical competition models in the mid-1950s (Petraitis 2013) and the potential existence of multiple ecological states was first described some 15 years later (Lewontin 1969). However, this theory did not enter the ecological mainstream until the following decade when several non-equilibrial systems , include rangelands, were described (May 1977). Nonequilibrium ecology and its associated metaphor, “the flux of nature ” (Pickett and Ostfeld 1995), are founded on the assumption that ecosystems possess a finite capacity for internal regulation such that they may be strongly influenced by disturbances (Wiens 1984; Wu and Loucks 1995). This implies that nonequilibrium systems possess greater potential for change than do equilibrium systems, including the potential to exhibit multiple stable states (Table 6.1).
2.2 Engineering Versus Ecological Resilience
Holling (1973) initially envisioned resilience theory by recognizing the potential occurrence of multiple stable states associated with the nonlinear dynamics in theoretical predator–prey models . Resilience was initially defined as the “persistence of relationships within a system and is a measure of the ability of these systems to absorb changes of state variables, driving variables and parameters, and still persist” (Holling 1973, p. 17). Two expressions of resilience later emerged to describe unique categories of ecosystem dynamics (Gunderson 2000). Engineering and ecological resilience broadly correspond to, but do not originate from, equilibrium and nonequilibrium ecological models, respectively. Engineering resilience describes system behavior near an individual equilibrium point and, therefore, system dynamics are assumed to be more consistent and predictable. Engineering resilience represents the time required for a system to return to its original equilibrium state after it has been modified by a disturbance (Holling 1973; Folke 2006). In contrast, ecological resilience describes system dynamics far from an equilibrium point and it recognizes the possibility that ecosystems may not return to their original equilibrium point and that they may reorganize around alternative equilibrium points (Gunderson 2000). Ecological resilience is currently defined as the capacity of systems to absorb disturbances and reorganize while undergoing change so as to still retain essentially the same function, structure, identity, and feedbacks (Walker et al. 2004).
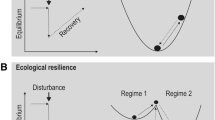
Resilience theory is often presented graphically in an attempt to clarify this abstract concept. The “basin-of-attraction ” or “ball-and-cup” graphic is among the most commonly used. In this highly simplified presentation the ball represents the current state of the system (state variables; structural system characteristics) with respect to the slow or controlling variables (parameters; ecological processes) of the system, the limits of which are represented by the size and shape of the basin (Beisner et al. 2003; Walker et al. 2012). Engineering resilience reflects the shape of the basin—its depth and degree of inclination—that determines the rate of recovery following disturbance (i.e., rate at which the ball returns to the bottom of the same basin). Ecological resilience is signified by the width of the basin of attraction , rather than its depth and inclination, as in the case for engineering resilience. If a disturbance forces the ball (structural system or community) beyond the rim of the basin (threshold) or if the width of the basin is narrowed by the modification of a controlling variable, resilience is exceeded and an alternative state may be formed as the ball moves into an adjacent basin (Gunderson 2000; Beisner et al. 2003). Multiple basins of attraction are representative of ecological resilience indicating that an ecosystem may possess more than one equilibrium state. The total number and shape of the basins in which an ecosystem may reside are collectively termed the resilience landscape (Walker et al. 2004) (Fig. 6.1).

The basin of attraction model illustrates ecological resilience as a ball (the community) that can reside in one or more basins of attraction (alternative states). Drivers may sufficiently modify controlling variables to force a community out of the original basin, beyond the ridge representing the threshold, and into an alternative basin forming a new stable state (see Table 6.1 for concept definitions) (from Beisner et al. 2003)
2.3 Drivers, Controlling Variables, and Feedback Mechanisms
As indicated in the previous section, resilience is influenced by interactions among several variables and these interactions can be modified by events both internal and external to the system (Walker et al. 2004, 2012). Drivers, controlling variables, and feedback mechanisms, in addition to the state variables previously introduced, are among the most important components of resilience (Text Box 6.1). Drivers are considered to be external to the system and they are not coupled to the system by feedback mechanisms (e.g., climatic regimes and extreme weather events). Controlling variables have a major influence on resilience and most systems are assumed to be regulated by a rather small number (3–5) of these variables (Chapin et al. 2010; Walker et al. 2012). They are often relatively stable through time, because they are buffered by feedback mechanisms (see below). Important controlling variables are major ecological processes—primary production and nutrient cycling; biodiversity—plant functional groups and woody plant encroachment; and historical disturbances—fire and grazing regimes. The modification of controlling variables directly by drivers or indirectly by feedback mechanisms represents the major way that resilience is altered (Beisner et al. 2003; Walker et al. 2012). These modifications are depicted as changes in the resilience landscape of the basin-of-attraction model (Fig. 6.1). Fast variables —annual plant and animal production—are more obvious than controlling (slow) variables because they fluctuate widely throughout an annual cycle. Critical interactions among these components occur when a driver—drought—modifies important controlling variables—grazing and fire regimes to influence numerous fast variables—grass growth and livestock gains.
Text Box 6.1: Concept Definition and Application to Resilience Theory
System or stable state—collection of multiple state variables and the feedback mechanisms that exist among them. State variables are broadly categorized as fast and slow (controlling).
Examples: Grassland, savanna, or shrubland communities.
Fast variables—variables characterized by dynamic and rapid responses to controlling variables and external drivers.
Examples: seasonal plant and animal production, compositional shifts in annual and transient species, soil water availability, pathogen, and insect dynamics.
Controlling (slow) variables—variables that often operate at slow rates and have a controlling influence on fast variables, feedbacks, and collectively system resilience; they are the central focus of resilience management.
Examples: dominant plant species, including plant functional groups and woody plant encroachment; grazing and fire regimes, soil characteristics, invasive species.
Drivers—events that are external to the system and do not possess feedbacks within the system; drivers may be of natural or human origin. They influence both fast and slow variables and their interactions within systems.
Examples: climatic regimes, extreme weather events, globalized markets, and human population growth.
Interpretation—drivers directly impact both fast and slow variables and the feedback mechanisms that exist between them. When a driver of sufficient magnitude modifies one or more slow (controlling) variables, threshold conditions may be established and existing stable states may transition to alternative stable states.
Application of resilience concepts to woody plant encroachment
System—grassland or savanna characterized by contiguous grass production that provides fine fuel to support regular fire regimes.
Fast variables—soil water availability, grass production, and fine fuel accumulation.
Controlling (slow) variables—dominant grass species, sustained intensive grazing regimes that reduce fine fuel accumulation, negative human perceptions and regulations limiting use of prescribed burning.
Drivers—severe drought that contributes to mortality of dominant grasses, human-induced land cover change, and increasing atmospheric carbon dioxide that increases woody plant growth.
Interpretation—interaction of natural and human drivers suppresses the controlling variable of fire frequency to enable threshold conditions to develop and grasslands or savannas to transition to alternative woodland states .
Feedback mechanisms are ecological processes that influence the rate of change among system variables. More specifically, they are secondary effects of one variable interacting with another to either enhance or dampen the rate of change of the initial variable. Stabilizing (negative) feedbacks reduce the rate of change of the initial variable (Gunderson 2000; Walker et al. 2012). For example, a high density of dominant grass species provides abundant, continuous fine fuel loads capable of supporting frequent fires to prevent woody plant encroachment and maintain grasslands. Amplifying (positive) feedbacks have the opposite effect and accelerate change of the initial variable. For example, increasing abundance of the invasive annual grass cheatgrass (Bromus tectorum) in the western USA increases fire frequency that contributes to the mortality of native vegetation to further increase cheatgrass dominance. In both examples, feedbacks interacted with a controlling variable—dominance of native grasses and invasion of an exotic plant species, respectively.
In the basin-of-attraction graphic, stabilizing feedbacks are greatest when communities reside near the bottom of the basin, while the relative strengths of stabilizing and amplifying feedbacks are assumed to be equivalent near the rim of the basin, which represents threshold conditions (Scheffer and Carpenter 2003; Walker et al. 2012). When the relative strength of amplifying feedbacks exceeds that of stabilizing feedbacks, one or more controlling variables may be sufficiently modified to create threshold conditions and initiate formation of an alternative state within a different basin of attraction (Fig. 6.2). Once an alternative stable state has been formed, resilience is established through a unique set of controlling variables and feedback mechanisms. The strengthening of stabilizing, relative to amplifying, feedbacks will support controlling variables and increase resilience of the alternative state. The potential for multiple interactions among external drivers, controlling variables , and feedback mechanisms over various time periods contributes to the difficulty of anticipating and describing thresholds and identifying ecological indicators of their occurrence (Briske et al. 2006; Bestelmeyer et al. 2011) (Text Box 6.1).

The feedback switch model depicts thresholds as the point where feedbacks switch from a dominance of negative (stabilizing) feedbacks (NFB) that maintain resilience (solid ball) to a dominance of positive (amplifying) feedbacks (PFB) that decrease resilience. The dominance of positive feedbacks contributes to formation of an alternative state (cross-hatched ball) in a different basin of attraction. Resilience of the alternative state requires that NFBs exceed PFBs (from Briske et al. 2006)
2.4 Threshold Indicators
The difficultly associated with threshold identification has focused attention on the search for early warning indicators. Indicators are assumed to signify modifications to state variables (structural characteristics), controlling variables, and to a lesser extent feedback mechanisms that determine the ecological resilience of a state. From a management perspective, indicators can be used to identify the trajectory of systems toward pending thresholds so that management strategies can be implemented or modified to prevent thresholds from being crossed (Briske et al. 2008; Standish et al. 2014). Alternatively, restoration ecologists may use threshold indicators to promote restoration of previous states that were considered more desirable (Suding and Hobbs 2009; Limb et al. 2014). Indicator effectiveness is a function of (1) the rate at which a system will respond to management actions to modify its resilience, (2) the amount and type of variability (noise) within systems that may mask indicator detection, and (3) the number of feedback mechanisms and controlling variables that contribute to system resilience (Contamin and Ellison 2009). In addition, threshold indicators are most relevant to systems where resilience is associated with gradual modification of controlling variables, rather than abrupt events that are difficult or even impossible to detect in advance (Hastings and Wysham 2010).
Two categories of theoretical early warning signals have been developed for threshold identification. The first emphasizes an increase in time required for recovery of system variables following disturbances that is termed “critical slowing down” (Scheffer et al. 2012; Dakos et al. 2012). The second category focuses on increasing variance and autocorrelation among system variables as thresholds are approached. It is assumed that both categories of indicators reflect a decrease in stabilizing feedback mechanisms as thresholds are approached (Walker et al. 2012). However, the limited scope of these early warning signals suggests that specific knowledge of systems dynamics, especially the major controlling variables, is still of greatest value (Dakos et al. 2012). Consequently, threshold identification on rangelands currently relies on ecological indicators that have been previously developed for evaluation of rangeland health and implementation of the range model last century and they are primarily implemented within the STM framework (Bestelmeyer et al. 2013) (Chapter 9, this volume).
2.5 Rethinking Rangeland Ecology
The concepts of nonequilibrium and resilience profoundly altered rangeland ecology by supporting development of the nonequilibrium persistent (NEP) model and state-and-transition model (STM) , respectively, in the late 1980s. However, these two models functioned independently because they focused on unique aspects of rangeland systems. The NEP model is based on the occurrence of nonequilibrium dynamics among vegetation and livestock, but it does not reference the existence of multiple stable states as does the STM. The following three subsections contrast the traditional equilibrium range model with the nonequilibrium persistent and multi-equilibrium state-and-transition model.
2.5.1 Range Model
The assessment of rangeland vegetation in response to grazing was initially linked to successional theory by Arthur Sampson, a former student of Fredric Clements, shortly following the publication of Clements’ influential work on succession in 1916 (Briske et al. 2005). However, a quantitative assessment of this procedure was not developed for another 30 years when Dyksterhuis (1949) published his classic paper outlining rangeland condition and trend analysis (here termed the range model). This procedure was adopted and applied to rangelands throughout the world during the last half of the twentieth century even though it encountered considerable criticism (Joyce 1993). The range model envisioned vegetation dynamics to occur along a single axis in which grazing intensity linearly counteracted secondary succession. The species composition of plant communities along a succession–grazing axis was compared to that of a single historic plant community to define a range condition rating. The more closely the species composition of a plant community approached that of the reference community, the higher the condition rating. These ratings were used to draw inferences for both production goals and ecological assessments (Joyce 1993). Range trend described the relative change in range condition ratings on specific sites through time. The adoption of Clementsian succession as the basis for vegetation assessment deeply embedded equilibrium ecology within the rangeland profession from its very beginnings (Fig. 6.3).

The range model assumes that the species composition of plant communities is a result of the opposing forces of plant succession and grazing intensity. Grazing can slow, stop, or reverse secondary succession to produce communities that differ in species composition from the historical climax plant community that represents the single reference (equilibrium) point (from Briske et al. 2005)
The expansion of woody plants and the persistence of these plant communities following the reduction or removal of grazing resulted in strong criticism of the range model in the 1970s and 1980s (Laycock 1991; Briske et al. 2005). However, in retrospect, the decision to use the grassland-savanna fire climax community (e.g., pre-European, Native American), as opposed to the climatic climax community (e.g., shrubland or woodland in wetter regions), as the primary reference state in the range model was a major contributor to these inconsistent outcomes. The selection of this reference state likely resulted from the recognized value of grasslands and savannas to livestock production , but climatic climax communities began to be expressed as historical fire regimes were minimized by grazing induced fuel reductions and direct fire suppression (Smith 1988; Westoby et al. 1989). However, the range model is still considered an appropriate interpretation of vegetation dynamics in more productive grasslands similar to those in which Dyksterhuis devised the range model (Fort Worth Prairie in north central Texas) (Dyksterhuis 1949). The occurrence of relatively linear vegetation dynamics in these grasslands is a result of intense plant competition and stronger plant–livestock interactions that are characteristic of equilibrium ecology (Díaz et al. 2001) (Table 6.1).
2.5.2 Nonequilibrium Persistent Model
The nonequilibrium persistent model was introduced by Ellis and Swift (1988) while conducting research in pastoral systems of the Turkana region of East Africa. This region is characterized by low annual and high interannual rainfall variability. This variability, especially when expressed as multiyear drought, frequently contributes to substantial livestock mortality in spite of attempts by nomadic pastoralists to track this variation. Livestock numbers recover less rapidly than plant production in the intervening favorable rainfall years such that they lag behind the availability of forage resources. This weak relationship between plant production and animal numbers contributed to the interpretation that these were nonequilibrium ecosystems. This interpretation gave rise to an alternative set of management and policy recommendations for pastoral systems in the early 1990s that was termed the “New Range Ecology ” (Behnke et al. 1993). These recommendations rejected the equilibrial concepts of carrying capacity, stocking rate, and the potential for livestock to degrade rangeland resources that were inherent to traditional grazing management (Cowling 2000) (Fig. 6.4).

The persistent nonequilibrium model indicates that multiyear droughts occurring on approximately decadal intervals minimize equilibrium between plant production and livestock numbers because vegetation recovers much more rapidly than livestock numbers following drought (redrawn from Ellis and Swift 1988)
Interannual variability in rainfall is negatively correlated with mean annual rainfall , and so the limitation on animal numbers imposed by low primary production in arid and semiarid rangelands is compounded by intra-annual rainfall variation . This makes it difficult to separate the effects of low annual rainfall from those of seasonal variability. A critical level of rainfall variability—an interannual coefficient of variation (CV, annual variability relative to the long-term mean) of ≥33 %—was established as the value at which animal numbers are no longer in equilibrium with plant production (Ellis and Swift 1988) (Fig. 6.4).
Reassessment of NEP. The NEP model was critically evaluated by Illius and O’Connor (1999, 2000) from the perspective of spatial and temporal forage availability to livestock. They concluded that the NEP model did not sufficiently consider livestock use of distinct vegetation resources between wet and dry seasons within an annual cycle. They reasoned that livestock numbers in environments characterized by wet and dry seasons are closely couple to a subset of “key” resources that are accessible in the dry season, while being largely uncoupled from more abundant forage in the wet season. The critical parameter establishing livestock herd size is often animal survival over the dry season, which is a function of forage availability during this period. Therefore, the ultimate determinant of animal numbers and their potential impacts on vegetation is the relative proportion of the grazed ecosystem that provides key resources during the dry season. It is this partitioning of forage resources, and not rainfall variability per se, that is the true determinant of livestock persistence and the potential for grazing to impact vegetation (llius and O’Connor 2000).
A reduction in access to key resource areas would cause livestock numbers to decline over the course of several years, especially during drought periods. In contrast, an abundance of non-key resources is likely to occur during the wet season, because animal numbers have been reduced by a scarcity of, and competition for, high-quality forage during the dry season . This interpretation establishes that livestock are closely coupled to forage resources, for at least part of the year, to meet their energy and nutrient requirements for survival, growth, and reproduction. This interpretation has been experimentally corroborated in a pastoral system located in an arid and highly stochastic environment (Hempson et al. 2015). In this investigation, livestock body condition followed density-dependent depletion of the limited dry-season riverine vegetation (key resources), and annual demographic parameters of animal populations tracked dry-season conditions. Dry-season length and previous population size were the main determinants of the animal population trajectory, with no clear evidence for an effect of growing season conditions over the vast area accessible to them. Therefore, wet-season rangeland can be categorized as nonequilibrium, because animal populations are only loosely coupled with it, but livestock do exist in equilibrium with dry-season resources.
Implications to Grazing Management. Reinterpretation of the NEP model has several important implications for management of livestock grazing. It indicates that the potential for grazing to modify vegetation and potentially degrade rangeland resources during the wet season increases as the proportion of key dry-season resource areas increases because it is these resources that establish the maximum number of livestock supported over the long term. Consequently, a high ratio of key dry:wet-season resources could support livestock numbers which are sufficient to produce high grazing intensities on wet-season resources, even though they may not be in equilibrium with them. Key resource areas themselves are obviously of considerable importance, and since they represent an equilibrial part of the grazed ecosystem they generally respond to increasing grazing intensity through reduced productivity and altered species composition (von Wehrden et al. 2012; Muthoni et al. 2014). These negative vegetation impacts will likely have direct, negative feedbacks on animal populations as described by traditional grazing management .
Commercial ranching represents a situation where livestock are often provided with supplemental feed during the dry or winter season to minimize animal mortality and weight loss. In these cases, livestock numbers would become uncoupled from both wet- and dry-season resources because grazing intensity is determined by animal numbers maintained by supplemental feeding. The maintenance of high livestock numbers during these periods increases the potential to adversely impact vegetation during the growing season and it reduces the opportunity for vegetation recovery following drought compared with less intensively managed systems. In principle, this interpretation would also apply to wild herbivores that migrate from wet-season (summer) resources to dry-season resources during the winter and then return to wet-season resources. Vegetation on summer range would be impacted to the extent that resource availability in winter range can support total animal numbers.
This reassessment strongly qualifies the NEP model by indicating that livestock will always maintain an equilibrial relationship with forage in key resource areas, even though this is not necessarily the case for abundant forage during the wet season . It also minimizes legitimacy of the “new” rangeland ecology by reaffirming that stocking rate and carrying capacity are valid concepts for grazing management, albeit in the context of larger landscapes and longer time periods (Cowling 2000).
2.5.3 The State-and-Transition Model
State-and-transition models were introduced as a “management language ,” rather than an ecological theory, to organize and interpret rangeland vegetation dynamics (Westoby et al. 1989). They provided an alternative to the range model that had been severely criticized in the 1970s and 1980s for being overly reliant on linear, directional vegetation dynamics that were unable to account for nonreversible vegetation change , especially woody plant encroachment and invasion by exotic invasive species (Briske et al. 2005). An influential report by the US National Research Council (NRC 1994) endorsed development and adoption of an alternative management model based on the STM framework and the Society for Range Management quickly supported this endorsement (SRM Task Group 1995). The US Department of Agriculture—Natural Resource Conservation Service (USDA-NRCS) formally adopted STMs for rangeland assessment in the late 1990s and established programs to develop and organize these models for all 50 states in the USA. A National Ecological Site Manual was developed and adopted in 2010 to standardize the use of ecological site descriptions and STMs among the NRCS, Bureau of Land Management, and US Forest Service (BLM 2010).
State-and-transition models are organized as a collection of all recognized or anticipated stable states that individual ecological sites may support (Bestelmeyer et al. 2003; Stringham et al. 2003). Individual stable states (e.g., grassland or shrubland) include transient and reversible shifts in species composition that occur in response to disturbances or self-regulating processes. These internal state dynamics are referred to as community phases and represent variation in species composition associated with wet and dry years, periodic intensive grazing, and fire frequency. In contrast, individual states are assumed to be separated by thresholds that are considered to be irreversible without management intervention . Ecological indicators of state variables, controlling variables, and to lesser extent feedback mechanisms that underpin state resilience are used to determine if a state is trending toward or away from pending thresholds (Briske et al. 2008). This information can inform managers of the need to implement actions to modify state resilience to achieve desired outcomes (Watson et al. 1996; Bagchi et al. 2013) (Fig. 6.5).

State-and-transition models are a representation of all known or anticipated stable states that may occupy an individual ecological site. States are assumed to be separated by thresholds that are considered to be nonreversible without management intervention. Community phases represent recognizable variations of a state that are readily reversible (from Stringham et al. 2003)
State-and-transition models have subsequently been organized around ecological resilience to link them to an accepted ecological theory and to accommodate scientific, in addition to management knowledge (Briske et al. 2008). In relation to resilience, individual states exist within a single basin of attraction that is consistent with engineering resilience and thresholds represent boundaries between multiple equilibrium states. Consequently, STMs are best interpreted as multiple equilibria rather than disequilibrium models (Petraitis 2013). Rangeland ecologists adopted the “nonequilibrium” terminology utilized by Westoby et al. (1989) because STMs were introduced as an alternative to the range model that was severely criticized for overemphasis of equilibrium dynamics associated with Clementsian succession (Joyce 1993; Briske et al. 2005). In addition, the focus of STMs has moved beyond threshold identification to the management of controlling variables and feedback mechanisms supporting resilience of stable states, but the nonequilibrium terminology has remained. Chapter 9 by Bestelmeyer and coauthors comprehensively describe the development, implementation, and interpretation of STMs.
2.6 What Has Been Learned?
Nonequilibrium ecology as described by the NEP model has been reinterpreted to indicate that while livestock may not necessarily be in equilibrium with forage during the wet season, they will always be in equilibrium with key forage resources during the dry season (Illius and O’Connor 1999). The recognition of two categories of forage resources with grazed systems indicates that those with low and highly variable rainfall do not function exclusively as “nonequilibrium systems” , because an equilibrial relationship with forage availability and livestock exists during the dry or winter season. Similarly, systems considered to be equilibrial are likely to experience intervals of nonequilibrium between livestock and forage during wet or dry seasons or years when forage production is high or when animal numbers are low. The current status of this rigorous debate is that appropriate qualification of equilibrium ecology, rather than its complete replacement by nonequilibrium models , offers more realistic interpretations for both plant–herbivore interactions and vegetation dynamics than does complete reliance on disturbance-driven events.
Resilience provides a framework to accommodate the occurrence of dynamic equilibria within ecosystems (i.e., engineering resilience) and the potential for ecosystems to transition to alternative stable states (i.e., ecological resilience). Recognition of nonlinear and nonreversible shifts between stable states initially focused attention on identification and characterization of thresholds separating these states. However, thresholds have proven very difficult to identify prior to their occurrence which minimizes their management value. An alternative approach that has greater management value focuses on the trajectory of state resilience relative to the proximity of potential thresholds, with the use of ecological indicators , rather than on thresholds themselves (Watson et al. 1996; Briske et al. 2008). State-and-transition models developed and maintained by the USDA-NRCS and its partners represent the major framework for application of ecological resilience to rangelands in the USA and elsewhere. Greater insight into feedback mechanisms and controlling variables establishing resilience, and ecological indicators to assess the trajectory of state resilience, are needed to promote implementation of resilience-based management (Bestelmeyer and Briske 2012).
3 Resilience of Social-Ecological Systems
Resilience theory is currently being explored in coupled social-ecological systems (SESs), in addition to ecological systems previously described. The objective is to provide a “humans-in-nature” perspective that serves to guide natural resource management , policy, and governance . As the name suggests, SESs are integrated systems of ecosystems, humans, institutions, and social organizations that contain feedbacks and interdependences among system components (Folke et al. 2010). This approach has provided common ground for social and natural sciences to interact and evaluate multiple knowledge sources addressing human–environment relationships. SESs are founded on the recognition that ecological information represents necessary but insufficient knowledge to manage ecosystems because they are strongly influenced by human needs, values, and goals (Chapter 8, this volume). Put more simply, “natural resource problems are human problems” (Ludwig 2001).
The concept of SESs emerged from interaction between social and ecological scientists in response to what was seen as failed natural resource management policy from the 1970s to the early 1990s. These policies often resulted in unintended negative consequences because they neglected the complex and often unrecognized interactions that exist between social and ecological components of these systems. These failures primarily originated from the ineffectiveness of the steady-state management model that emphasized maximum sustainable production of one or a few ecosystem services such as livestock production from rangelands and timber from forests, with little concern for other components of these complex systems. The deficiencies of this management model reside in the narrow interpretation of sustainability, inevitable trade-offs between sustainability and maximum resource yield, and tendency to overestimate resource availability and the consistency of resource yield (Holling and Meffe 1996; Ascher 2001). In contrast, resilience and resilience-based management focus on the variability and uncertainty of SESs and encourage managers to guide the direction of change, rather than to prevent change, to provide diverse ecosystem services to society (Chapin et al. 2010). However, in spite of the recognized deficiencies of the steady-state management model, it still remains the most widely used natural resource management strategy today (Anderies et al. 2006). Its persistence is likely a consequence of the central role that optimal control procedures have played in natural resource economics and the absence of a viable alternative management strategy.
At the same time that C.S. Holling and coworkers were studying the problems caused by the “command-and-control pathology of natural resource management ” discussed above, Elinor Ostrom was also questioning the rationality of top-down, command-and-control governance structures from a policy perspective (Ostrom 1990). Her work demonstrated that small groups of people can effectively manage complex natural resource challenges without top-down governance. She further suggested that top-down interventions could have “pathological” effects on social systems by reducing their capacity to solve problems similar to the way in which Holling envisioned the negative impact of top-down control on ecosystems. These two independent assessments of natural resource management failures —one ecological and other sociological—eventually converged to giving rise to the concept of SESs in the 1900s (Berkes et al. 2003) (Chapter 8, this volume).
3.1 Resilience Thinking
Resilience thinking provides a general framework for organizing and analyzing information regarding SESs to guide sustainable development and natural resource use (Folke 2006; Cote and Nightingale 2012). In this context, resilience is more appropriately interpreted as an approach and set of assumptions to analyze and guide SESs, rather than a system property as described in the previous sections (Biggs et al. 2015). The application of resilience to SESs implies “general” resilience that considers the potential existence of multiple drivers, disturbances, and thresholds, as opposed to “specified” resilience that emphasizes the impact of a smaller number of impacts on a specific threshold (Anderies et al. 2006). Resilience thinking is widely viewed as being comprised of three broad components (Folke et al. 2010). First, as previously defined for ecological systems, resilience describes the capacity of SESs to continually change and adapt, and yet remain within their current basin of attraction . Second, adaptive capacity describes the ability of humans to guide and direct change by enabling SESs to respond and adapt to internal and external events so that they can maintain their integrity and function. Third, transformation describes the capacity to create an alternative SES when resilience of the previous system can no longer be maintained by incremental adaptation (Folke et al. 2010). Each of these components will be discussed further in the following sections.
3.1.1 Social Resilience
Social resilience refers to the ability of human communities to withstand external shocks to their social infrastructure , such as environmental variability or social, economic, and political disruption (Adger 2000). However, attempts to transfer resilience theory from ecological systems to social systems have encountered several major concerns. Central among them is the validity of the assumption of persistent relationships—feedback mechanisms as described for ecosystems—which determine the ability of SESs to absorb natural and social disruption, change, and continue to persist (Cote and Nightingale 2012; Brown 2014). Social resilience emphasizes societal values and human behaviors, including power relations, equity and justice, and function of social organizations, which are central to human–environment relationships and social change, but are absent from ecological systems (Cote and Nightingale 2012; Olsson et al. 2015). Therefore, thresholds may represent changes in institutional procedures, including power sharing in decision making, wealth distribution, and land tenure, rather than irreversible divisions between two stable states (Christensen and Krogman 2012). In addition, neither social nor ecological resilience are normative so they do not provide a basis to distinguish between desirable and undesirable expressions of resilience (Brown 2014). Consequently, the value of resilience as an analytical tool for SESs has proven even more difficult to define and implement than for ecological systems (Benson and Garmestani 2011). The broad integration of knowledge and perspectives that conveys value to resilience as an organizing framework likely represents the same attributes that restrict its application to standardized management protocols and policies (Cote and Nightingale 2012).
3.1.2 Adaptive Capacity and Social Learning
Adaptive capacity describes the ability of humans to create, and shape variability and change in SESs, and it represents a central component of social resilience. Adaptive management, social learning, and adaptive governance are all components of adaptive capacity that are necessary for putting resilience into practice. Adaptive management is often described as learning by doing and it operates in an iterative manner acknowledging that our understanding of complex systems is incomplete and constantly changing. Adaptive management was introduced shortly following development of resilience theory to incorporate the inevitable constraints of uncertainty and insufficient knowledge into natural resource management (Holling 1978). Social learning describes the process by which groups of individuals assess social-ecological conditions and respond in ways that meet desired objectives. This represents a central component of the ability of small coordinated groups of natural resource users to develop more effective local governance than top-down policies (Ostrom 1990) (Chapter 11, this volume).
3.1.3 Anticipating System Transformation
Transformation becomes necessary when adaptive capacity and available adaptation strategies are no longer sufficient to maintain resilience of SESs. Transformation describes the capacity to create fundamentally new SESs when ecological, economic, or social conditions make the existing system unsustainable (Walker et al. 2004). Ecological change in the form of increased climatic variability or social disruption regarding the availability and allocation of land, labor, and capital can initiate the need for transformation. The intended goal of transformation is to reorganize SESs around alternative and likely novel basins of attraction that can provide ecosystem services to sustainably support human livelihoods when previous SESs have failed (Walker and Salt 2012).
The capacity of SESs to successfully manage transformation has only recently been considered and is not yet well developed. However, successful transformation may be dependent on five key considerations: awareness, incentives, networks, experimentation, and assets. Awareness of the need to implement transformative strategies is dependent upon the ability of members of the SES to recognize and broadly communicate the occurrence of unsustainable conditions and the need for transformation (Carpenter and Folke 2006; Marshall et al. 2011). Incentives may be required to encourage voluntary change because indecision or denial is often an immediate response to the loss of resilience in SESs (Walker and Salt 2012). The strength of social networks , ability of participants to experiment , preferably at local to regional scales until cost-effective strategies have been established, and leadership are important components for implementing transformation (Nelson et al. 2007; Folke et al. 2010). Finally, effective transformation requires flexibility in the assets or resources necessary to implement change. Recognizing and communicating the need for transformation and developing policies, programs, and actions to support determination of when and how to initiate transformational change represent important challenges for the rangeland profession (Joyce et al. 2013) (Chapter 15, this volume).
3.2 Resilience-Based Governance and Policy
Institutions and policies governing the behavior of SESs influence resilience by defining the rights and capacities of managers to make decisions regarding social-ecological trade-offs (Horan et al. 2011). A framework capable of supporting resilience thinking in SESs must be able to address the problems of governance, including management and policy decisions in the face of uncertainty, disputed values, and potential shifts to alternative states (Carpenter and Brock 2008). The failure to recognize and manage feedbacks in SESs, including limited monitoring of policy outcomes and insufficient adaptive management , may be among the primary reasons previous institutions have contributed to natural resource management failures (Holling and Meffe 1996; Ascher 2001). Therefore, a key component of the resilience of SESs is the reorganization of institutions to monitor policy outcomes and implement information feedbacks to learn from previous actions. This may be best achieved by focusing on experimentation, adaptation, and social learning within local communities, rather than on implementation of generalized policies originating from static institutions that are assumed to be efficient or “right” (Benson and Garmestani 2011; Anderies and Janssen 2013).
Designing governance for resilient SESs requires understanding of how biophysical conditions, social structure, and institutional policies interact and affect each other and the entire system. Resilient governance regimes are those that achieve a good fit between these different system components so that they continue to function under conditions of uncertainty and disturbance to the system (Anderies and Janssen 2013). In this way, “resilience thinking” departs considerably from traditional policy and resource management which typically takes a narrower view of imposing top-down policies to drive the system toward specific outcomes. Understanding how different components in SESs respond and interact to management strategies and policies and what outcomes will be produced has proven extremely difficult. Consequently, only general principles of governance have been identified to promote resilience in SESs (Anderies et al. 2006). Three of these major principles follow:
-
(1)
Collaboration to build trust and promote dialogue toward a shared understanding of the system among stakeholders is necessary to mobilize action and self-organize SESs (Ostrom 1990).
-
(2)
Multilayered institutions located at various scales within the system improve the fit between knowledge, action, and social-ecological interactions in ways that promote adaptive societal responses at appropriate times and locations (Ostrom 1990; Anderies and Janssen 2013).
-
(3)
Accountable authorities that pursue the just distribution of benefits and involuntary risks among stakeholders to enhance the adaptive capacity of the most vulnerable groups (Lebel et al. 2006).
3.3 Resilience Analysis and Management
Resilience management in SESs emphasizes two major, interrelated goals. First is to prevent the system from transitioning to an undesirable stable state, and second is to retain and promote the ability of SESs to reorganize following major change (Walker et al. 2002). The intent is not to direct the trajectory of SESs toward a predetermined endpoint, but rather to strengthen the internal feedbacks to enhance general resilience to both anticipated and unanticipated future change. A planning approach consisting of four broad, interrelated steps has been developed as a means to retain and promote general resilience of SESs (Walker et al. 2002). The planning process is to be conducted by multiple stakeholders possessing diverse interests and knowledge of the SES under consideration (Fig. 6.6).
-
Step 1. Develop a Conceptual System Model . The initial step is to establish boundaries for the SESs, and to identify the major management issues, critical stakeholders groups, and primary drivers of change. Identification of major ecosystem services, primary controlling variables, and institutional and governance arrangements is also important. Investigation of the historical profile of SESs emphasizing the impact and adaptive responses to previous disturbances will provide valuable baseline information.
-
Step 2. Create Scenarios of Future System Trajectories . The second step is the development of a limited set (3–5) of future scenarios that address a range of potential SES trajectories in response to the major drivers of change identified in step 1. The scenarios may include a business-as-usual scenario, a more confined or conservative scenario, and one or two more exploratory scenarios. These scenarios are not intended to be predictions, but rather broad plausible visions of potential outcomes that are consistent with existing evidence.
-
Step 3. Resilience Analysis . The goal is to assess how the system may change within each of the scenarios identified in the previous step. This assessment should emphasize anticipated responses of SESs to the drivers and processes that influence stakeholder interests. The identification of thresholds, alternative states , and other potential surprises are of primary importance. This step is highly context dependent and therefore difficult to define in specific detail.
-
Step 4. Resilience Management . The final step involves stakeholder evaluation of the knowledge created in the previous steps for management and policy considerations. The implications of this knowledge for assessment and management of critical feedback mechanisms and controlling variables that determine general resilience of SESs are especially relevant. A specific trajectory for SESs is not selected because it is assumed that insufficient information and occurrence of unanticipated change will limit the value of predictions.

A resilience analysis and management procedure that consists of four broad, interrelated steps that are to be conducted by multiple stakeholder groups as a means to retain and promote resilience of social-ecological systems (Walker et al. 2002)
3.4 What Has Been Learned?
The concepts of social resilience and SESs emphasize a “humans-in-nature” perspective that recognizes that ecological knowledge alone is insufficient to sustainably manage human-dominated systems. These concepts recognize the importance of human values and goals to sustainable natural resource management and they create opportunities for multiple stakeholders to adaptively design management strategies and policies. In the context of social resilience, sustainability is pursued by acknowledging the existence of uncertainty, incomplete knowledge, and the potential for abrupt shifts to alternative states , as opposed to steady state management that attempts to minimize variability and target specific outcomes. The application of resilience to SESs emphasizes “general” resilience to various human and natural disturbances, rather than “specified” resilience that emphasizes the impact of a small number of known impacts on specific thresholds. Adaptive capacity has emerged as a key feature of general resilience in SESs that includes adaptive management , social learning, and adaptive governance. Adaptive capacity may be best achieved by focusing on experimentation, adaptation, and collaboration within local communities, rather than on implementation of generalized policies that are assumed to be efficient or “right.” Rangeland SESs and the human livelihoods they support are especially vulnerable to a loss of resilience given that they are often characterized by resource scarcity and variability . In cases where the resilience of SESs has been exceeded, transformational change will be required to reorganize these systems within other basins of attraction to support human livelihoods through the production of alternative ecosystem services with different management strategies .
4 Future Perspectives
In this section we summarize several perspectives regarding the future development and implementation of resilience that emerged during the writing of this chapter.
4.1 Heterogeneity and Livestock-Vegetation Dynamics
Reinterpretation of the NEP model has indicated that grazed systems often contain two unique resource categories: one in which livestock may not necessarily be in equilibrium during the wet season and the other is key resource areas with which livestock are always in equilibrium (Illius and O’Connor 1999). These resource categories are created by heterogeneity in the relative proportion and spatial arrangements of wet- and dry-season forage resources within the landscape (Hempson et al. 2015). The manner in which these spatial attributes influence coupling of vegetation and livestock in relation to length of the wet and dry seasons would provide greater insight into this component of grazed systems. The importance of functional heterogeneity needs to be more effectively incorporated into management recommendations and policy decisions (Chapter 5, this volume).
4.2 Procedures to Implement Resilience-Based Management
Resilience has gained wide acceptance as a framework to guide natural resource management , but the procedures necessary to implement it require additional development. Thresholds and alternative states have received the greatest attention, but they do not necessarily provide the best information to guide resilience-based management because they are often only recognized after their occurrence. Greater emphasis needs to be focused on the identification of ecological indicators of controlling variables and feedback mechanisms to assist managers in assessing the trajectory of ecosystem resilience and to identify appropriate management strategies to modify these trajectories when desired.
The search for procedures to implement resilience has encountered some friction between traditional and contemporary scientific approaches regarding the trade-off between precision and vagueness. It has been argued that vagueness, which is normally viewed as being detrimental to scientific progress, may actually promote creativity and problem solving within the context of resilience-based management (Strunz 2012). This creates considerable uncertainty regarding the extent to which resilience procedures should be standardized and formalized for application. State-and-transition models as a component of Ecological Site Descriptions are currently the primary procedure for implementation of resilience-based management on rangelands. It remains uncertain whether procedures addressing the resilience of SESs should be incorporated into this framework or if a separate framework specifically focusing on SESs is required. Emphasis on SESs will require a reorientation from specified resilience emphasizing specific stable states and thresholds, as widely applied today, to general resilience of entire SESs that exist at larger spatial scales.
4.3 Recognizing and Guiding Transformation
Globalized markets, climate change, loss of biodiversity, and species invasion collectively contribute to conditions in which SESs and the human livelihoods they provide may become unsustainable. Rangeland SESs may be especially vulnerable because they are frequently characterized by resource scarcity and variability, limited infrastructure and financial capital, and few viable alternatives to sustain human livelihoods (Sayre et al. 2013). Development of policies and programs to increase awareness and communication of the need for transformational change represents an important challenge. The ability to determine when an SES is becoming unsustainable and how to effectively guide transformation of SESs toward a more sustainable alternative requires greater consideration. This will require an assessment of resilience over multiple timescales (Anderies et al. 2013). Short-term decisions primarily focus on specified resilience to maintain stabilizing feedbacks of a desirable state that will minimize development of an alternative state . Mid- and long-term decisions require emphasis on general resilience by adapting SESs to increasing uncertainty and new conditions, and on transformational change when appropriate. Strategies focused on general resilience will require a greater understanding of the organization and function of SESs than is currently available.
4.4 Institutional Reorganization to Promote Resilience
A centralized organizational structure controlling both power and financial resources often supports a command-and-control management strategy which is recognized as an impediment to resilience-based management (Holling and Meffe 1996). Consequently, the traditional management, policy, and institutions responsible for natural resource management present major challenges to the implementation of resilience thinking (Benson and Garmestani 2011). Formal institutional arrangements within existing laws and regulations often ignore ecological complexity and variability, and emphasize a “preservation paradigm” that is focused on minimizing or mitigating human damage to ecosystems. Current natural resource management policies and incentives are often designed to prevent change in the desired “optimal” state and they often represent perverse incentives that may eventually reduce resilience of the system (Anderies et al. 2006). In addition, management agencies often focus exclusively on ecological components of natural resource challenges, but largely overlook the associated social components. This is largely a consequence of legal and policy frameworks that separate decision making regarding these two systems. However, as previously indicated, ecological and social systems are tightly linked through reciprocal feedbacks that require simultaneous consideration for development of effective management and policy.
Barriers that must be overcome to modify governance to enhance resilience of SESs include (1) the tendency for political expediency to modify, rather than change, existing institutions; (2) reliance on traditional procedures and knowledge to address novel, complex problems; and (3) fragmented governance among land ownership patterns and institutional jurisdictions (Brunckhorst 2002). Potential implementation of the Endangered Species Act to address conservation and management of greater sage-grouse (Centrocercus urophasianus) and its habitat represents an excellent example of an institutional mismatch with complex natural resource management challenges (Boyd et al. 2014). The regulatory approach is incapable of addressing the ecological and social complexity of these challenges because it is not designed to empower collaborative adaptive management among diverse stakeholders (Chapter 11, this volume).
5 Summary
Rangeland ecology and management have undergone a major transformation in the past quarter century as nonequilibrium ecology and resilience theory were adopted to increase consistency with observed ecological dynamics and management outcomes. Equilibrium ecology had long been a guiding ecological principle that emphasized linear and predictable ecosystem dynamics and it supported the steady-state management model that prevailed throughout the twentieth century. Equilibrium ecology was challenged on the basis of both theoretical inconsistencies and its inability to account for observed ecosystem dynamics. Nonequilibrium ecology recognizes that disturbance, spatial heterogeneity, and multiple stable states, in addition to internal biotic regulation, have a major influence on ecosystem dynamics (Wu and Loucks 1995).In addition to these broad implications of nonequilibrium ecology, rangeland ecology was explicitly challenged by the persistent nonequilibrium model that originated in pastoral systems in East Africa. This model indicated that forage availability and livestock numbers were seldom in equilibrium because vegetation recovered more rapidly than livestock numbers following multiyear drought (Ellis and Swift 1988). It was further assumed that this weak coupling between vegetation and livestock minimized the potential for grazing to degrade rangeland resources. However, the persistent nonequilibrium model has been qualified by recognizing that livestock are always in equilibrium with the key resource areas of a grazed system, even though they may only be loosely coupled to abundant wet-season resources. Reinterpretation of the nonequilibrium persistent model challenges the legitimacy of the “new” rangeland ecology by reaffirming that stocking rate and carrying capacity are valid concepts for grazing management, albeit in the context of larger landscapes and longer time periods.
Two categories of resilience exist to describe unique patterns of ecosystem dynamics. Engineering resilience assumes that systems are confined to a single basin of attraction and it is represented by the time required for a system to return to its original equilibrium point following disturbance (Holling 1973). In contrast, ecological resilience recognizes that ecosystems may not return to their original equilibrium point, but that they may reorganize around alternative equilibrium points (Gunderson 2000). Resilience is often expressed graphically with “basin-of-attraction” or “ball-and-cup” diagrams to further clarify this abstract concept. In this graphic representation, the ball represents the structural variables of the system in relation to the controlling variables and feedback mechanisms that are represented by the shape and number of the basins (Beisner et al. 2003; Walker et al. 2012). Engineering resilience emphasizes the depth of the basin which determines the rate of recovery following disturbance. Ecological resilience emphasizes the width of the basin to represent the amount of disturbance that a system can withstand without the ball rolling beyond the rim of the basin (threshold) into an alternative basin. Ecological resilience is most commonly applied to natural resource management where thresholds and the formation of alternative stable states are of major concern.
State-and-transition models represent a conceptual advance from the traditional range model that was founded on Clementsian succession by recognizing the occurrence of nonlinear vegetation dynamics and the potential existence of alternative stable states on individual ecological sites. These models were originally introduced as a “management language,” rather than an ecological theory, to organize and interpret rangeland vegetation dynamics. However, the STM framework has become a major tool for interpreting and integrating both management and ecological information. Subsequently, these models are broadly viewed as being supported by ecological resilience and are most appropriately considered equilibrium models because individual states exist with a single basin of attraction and thresholds represent boundaries between multiple equilibrium states. Ecological indicators of state variables, controlling variables, and to lesser extent feedback mechanisms that underpin resilience are used to determine if a state is trending toward or away from pending thresholds (Briske et al. 2008). This information can inform managers of the need to implement actions to modify state resilience to achieve desired outcomes.
The adoption of resilience theory has had broad consequences for natural resource management by providing an alternative to command-and-control management. Command and control employs a top-down, regulatory strategy that often ignores variation and the complexity of interactions among ecological and social system components (Anderies et al. 2006). The typical response to uncertainty and surprise is to increase control which often further reduces resilience and moves the system toward pending thresholds (Holling 1973; Holling and Meffe 1996). Management strategies to minimize variability and optimize production efficiency in one portion of the system frequently increase vulnerability in another portion of the system. These top-down management strategies have also been founded on unrealistic assumptions regarding the ability of managers to anticipate and implement actions, often as technological solutions, in time to prevent natural resource degradation or ecosystem shifts to alternative states.
In contrast to command and control, resilience-based management recognizes both the inevitability of change and the need for change, such that it seeks to guide change, rather than to control it to maintain a single optimal state. A family of concepts—alternative stable states, thresholds, controlling variables, and feedbacks—has evolved around resilience theory that has both increased its potential conceptual value and introduced considerable ambiguity (Strunz 2012). The vagueness inherent to resilience theory is counter to the clarity and precision normally associated with science, but this trade-off may be necessary to promote creativity, trans-disciplinarity, and cooperation among diverse stakeholder groups that is needed to contend with the complexity of natural resource problems currently confronting society. The broad and often ambiguous elements of resilience that have contributed to its intuitive value and appeal are likely the same attributes that make it challenging to interpret and implement (Cote and Nightingale 2012).
Some of the critical challenges confronting the application of resilience to natural resource management are represented by the following questions. How much disturbance can ecosystems absorb before they cross thresholds and reorganize as alternative stable state (Standish et al. 2014)? Is it possible to identify indicators of resilience within existing stable states so that management actions can be implemented to reduce the probability of a threshold being crossed? How can restoration programs best prioritize efforts to reestablish stable states after they have crossed a threshold and reorganized as an alternative stable state? How can resilience thinking be most effectively incorporated into management recommendations and policy decisions without impairing generality and flexibility inherent to the theory (Strunz 2012)? Is the resilience framework relevant to SESs (Anderies et al. 2006)? These questions are especially challenging given that experimental evidence is very limited and that experiments to address these questions are extremely difficult to conduct in large complex systems. Historical data has been suggested as a means to investigate resilience by retrospective analysis of events in both ecological (Standish et al. 2014) and SESs (Stafford Smith et al. 2007). All indications are that resilience will continue to be a work in progress given both the complexity of the concepts involved and the enormity of natural resource challenges to be addressed.
Resilience has been envisioned as a framework to guide society on a path toward sustainability (Folke 2006). It has even been suggested that resilience should replace sustainability as an organizing framework to support environmental management and ecological governance because it is often impossible to know what should and can be sustained in a world of increasing complexity, uncertainty, and rapid change (Benson and Craig 2014). Resilience-based management emphasizes adaptive management and the development of adaptive capacity to guide change, rather than management actions to reduce variability in an attempt to stabilize desired systems. In this context, resilience may represent a more valuable framework than sustainability for natural resource management because it anticipates uncertainty and emphasizes learning to contend with future challenges (Strunz 2012). However, for resilience to meaningfully contribute to this enormous challenge it must be put it into practice at multiple scales of application or it may encounter the same ambiguous outcome as sustainability.
References
Adger, W.N. 2000. Social and ecological resilience: Are They Related? Progress in Human Geography 24: 347–364.
Anderies, J.M., and M.A. Janssen. 2013. Robustness of social–ecological systems: Implications for public policy. Policy Studies Journal 41(3): 513–536.
Anderies, J.M., B.H. Walker, and A.P. Kinzig. 2006. Fifteen weddings and a funeral: Case studies and resilience-base management. Ecology & Society 11(1): 21.
Anderies, J.M., C. Folke, B. Walker, and E. Ostrom. 2013. Aligning key concepts for global change policy: Robustness, resilience and sustainability. Ecology & Society 18(2): 8.
Ascher, W. 2001. Coping with complexity and organizational interests in natural resource management. Ecosystems 4: 742–757.
Bagchi, S., D.D. Briske, B.T. Bestelmeyer, and X.B. Wu. 2013. Assessing resilience and state-transition models with historical records of cheatgrass Bromus tectorum invasion in North American sagebrush-steppe. Journal of Applied Ecology 50: 1131–1141.
Behnke, R.H., I. Scoones, and C. Kerven. 1993. Range ecology at disequilibrium: New models of natural variability and pastoral adaptation in African savannas. London: Overseas Development Institute.
Beisner, B.E., D.T. Haydon, and K. Cuddington. 2003. Alternative stable states in ecology. Frontiers in Ecology and the Environment 1: 376–382.
Benson, M.H., and R.K. Craig. 2014. The end of sustainability. Society and Natural Resources 27: 777–782.
Benson, M.H., and A.S. Garmestani. 2011. Can we manage for resilience? The integration of resilience thinking into natural resource management in the United States. Environmental Management 48: 392–399.
Berkes, F., J. Colding, and C. Folke. 2003. Navigating social-ecological systems: Building resilience for complexity and change. Cambridge: Cambridge University Press.
Bestelmeyer, B.T., and D.D. Briske. 2012. Grand challenges for resilience-based management of rangelands. Rangeland Ecology & Management 65: 654–663.
Bestelmeyer, B.T., J.R. Brown, K.M. Havstad, R. Alexander, G. Chavez, and J.E. Herrick. 2003. Development and use of state-and-transition models for rangelands. Journal of Range Management 56: 114–126.
Bestelmeyer, B.T., A.M. Ellison, W.R. Fraser, K.B. Gorman, S.J. Holbrook, C.M. Laney, M.D. Ohman, D.P.C. Peters, F.C. Pillsbury, A. Rassweiler, R.J. Schmitt, and S. Sharma. 2011. Analysis of abrupt transitions in ecological systems. Ecosphere 2: 129. doi:10.1890/ES11-00216.1.
Bestelmeyer, B.T., M.C. Duniway, D.K. James, L.M. Burkett, and K.M. Havstad. 2013. A test of critical thresholds and their indicators in a desertification-prone ecosystem: More resilience than we thought. Ecology Letters 16: 339–345.
Biggs, R., M. Schluter, and M.L. Schoon. 2015. An introduction to the resilience approach and principles to sustain ecosystem services in social–ecological systems. In Principles for building resilience: Sustaining ecosystem services in social–ecological systems, ed. R. Biggs, M. Schluter, and M.L. Schoon, 1–31. Cambridge, UK: Cambridge University Press.
Boyd, C.S., D.D. Johnson, J.D. Kerby, T.J. Svejcar, and K.W. Davies. 2014. Of grouse and golden eggs: Can ecosystems be managed within a species-based regulatory framework? Rangeland Ecology & Management 67: 358–368.
Briske, D.D., S.D. Fuhlendorf, and F.E. Smeins. 2003. Vegetation dynamics on rangelands: A critique of the current paradigms. Journal of Applied Ecology 40: 601–614.
———. 2005. State-and-transition models, thresholds, and rangeland health: A synthesis of ecological concepts and perspectives. Rangeland Ecology & Management 58: 1–10.
———. 2006. A unified framework for assessment and application of ecological thresholds. Rangeland Ecology & Management 59: 225–236.
Briske, D.D., B.T. Bestelmeyer, T.K. Stringham, and P.L. Shaver. 2008. Recommendations for development of resilience-based state-and-transition models. Rangeland Ecology & Management 61: 359–367.
Brown, K. 2014. Global environmental change I: A social turn for resilience? Progress in Human Geography 38: 107–117.
Brunckhorst, D.J. 2002. Institutions to sustain ecological and social systems. Ecological Management & Restoration 3: 108–116.
Bureau of Land Management (BLM). 2010. Rangeland interagency ecological site manual. Washington, DC: BLM Manual 1734-1.
Carpenter, S.R., and W.A. Brock. 2008. Adaptive capacity traps. Ecology & Society 13(2): 40.
Carpenter, S.R., and C. Folke. 2006. Ecology for transformation. Trends in Ecology and Evolution 21: 309–315.
Chapin III, F.S., S.R. Carpenter, G.P. Kofinas, C. Folke, N. Abel, W.C. Clark, P. Olsson, D.M. Stafford Smith, B. Walker, O.R. Young, F. Berkes, R. Biggs, J.M. Grove, R.L. Naylor, E. Pinkerton, W. Stephen, and F.J. Swanson. 2010. Ecosystem stewardship: Sustainability strategies for a rapidly changing planet. Trends in Ecology and Evolution 25: 241–249.
Christensen, L., and N. Krogman. 2012. Social thresholds and their translation into social–ecological management practices. Ecology & Society 17(1): 5.
Contamin, R., and A.M. Ellison. 2009. Indicators of regime shifts in ecological systems: What do we need to know and when do we need to know it? Ecological Applications 19(3): 799–816.
Cote, M., and A.J. Nightingale. 2012. Resilience thinking meets social theory: Situating social change in socio-ecological systems. Progress in Human Geography 36: 475–489.
Cowling, R.M. 2000. Challenges to the ‘new’ rangeland science. Trends in Ecology and Evolution 15: 303–304.
Curtain, C.G., and J.P. Parker. 2014. Foundations of resilience thinking. Conservation Biology 28(4): 912–923.
Dakos, V., S.R. Carpenter, W.A. Brock, A.M. Ellison, V. Guttal, et al. 2012. Methods for detecting early warnings of critical transitions in time series illustrate using simulated ecological data. PLoS ONE 7(7), e41010.
Díaz, S., I. Noy-Meir, and M. Cabido. 2001. Can grazing response of herbaceous plants be predicted from simple vegetative traits? Journal of Applied Ecology 38: 497–508.
Dyksterhuis, E.J. 1949. Condition and management of rangeland based on quantitative ecology. Journal of Range Management 2: 104–105.
Ellis, J.E., and D.M. Swift. 1988. Stability of African pastoral ecosystems: Alternate paradigms and implications for development. Journal of Range Management 41: 450–459.
Folke, C. 2006. Resilience: The emergence of a perspective for social–ecological systems analyses. Global Environmental Change 16: 253–267.
Folke, C., S.R. Carpenter, B. Walker, M. Scheffer, T. Chapin, and J. Rockstrom. 2010. Resilience thinking: Integrating resilience, adaptability and transformability. Ecology & Society 15(4): 20.
Gunderson, L.H. 2000. Ecological resilience—In theory and application. Annual Review of Ecology and Systematics 31: 425–439.
Hastings, A., and D.B. Wysham. 2010. Regime shifts in ecological systems can occur with no warning. Ecology Letters 13: 464–472.
Hempson, G.P., A.W. Illius, H.H. Hendricks, W.J. Bond, and S. Vetter. 2015. Herbivore population regulation and resource heterogeneity in a stochastic environment. Ecology 96: 2170–2180.
Holling, C.S. 1973. Resilience and stability of ecological systems. Annual Review of Ecology and Systematics 4: 1–23.
———. 1978. Adaptive environmental assessment and management. Chinchester: John Wiley and Sons.
Holling, C.S., and G.K. Meffe. 1996. Command and control and the pathology of natural resource management. Conservation Biology 10: 328–337.
Horan, R.D., E.P. Fenichel, K.L.S. Drury, and D.M. Lodge. 2011. Managing ecological thresholds in coupled environmental–human systems. Proceedings of the National Academy of Science, USA 108: 7333–7338.
Illius, A.W., and T.G. O’Connor. 1999. On the relevance of non-equilibrium concepts to arid and semi-arid grazing systems. Ecological Applications 9: 798–813.
Illius, A.W., and T.G. O’Connor. 2000. Resource heterogeneity and ungulate population dynamics. Oikos 89: 283–294.
Jones, N.A., H. Ross, T. Lynam, P. Perez, and A. Leitch. 2011. Mental models: An interdisciplinary synthesis of theory and methods. Ecology & Society 16:46 [online]. Retrieved from http://www.ecologyandsociety.org/vol16/iss1/art46/.
Joyce, L.A. 1993. The life cycle of the range condition concept. Journal of Range Management 46: 132–138.
Joyce, L.A., D.D. Briske, J.R. Brown, H.W. Polley, B.A. McCarl, and D.W. Bailey. 2013. Climate change and North American rangelands: Assessment of mitigation and adaptation strategies. Rangeland Ecology & Management 66: 512–528.
Laycock, W.A. 1991. Stable states and thresholds of range condition on North American rangelands: A viewpoint. Journal of Range Management 44: 427–433.
Lebel, L., J.M. Anderies, B. Campbell, C. Folke, S. Hatfield-Dodds, T.P. Hughes, and J. Wilson. 2006. Governance and the capacity to manage resilience in regional social–ecological systems. Ecology & Society 11(1): 19.
Lewontin, R.C. 1969. The meaning of stability. In Diversity and stability of ecological systems. Brookhaven Symposia in Biology No. 22. Brookhaven, New York.
Limb, R.F., D.M. Engle, A.L. Alford, and E.C. Hellgren. 2014. Plant community response following removal of Juniperus virginiana from tallgrass prairie: Testing for restoration limitations. Rangeland Ecology & Management 67: 397–405.
llius, A.W., and T.G. O’Connor. 2000. Resource heterogeneity and ungulate population dynamics. Oikos 89: 283–294.
Ludwig, D. 2001. The era of management is over. Ecosystems 4: 758–764.
Lynam, T.J.P., and M. Stafford Smith. 2004. Monitoring in a complex world—Seeking slow variables, a scaled focus, and speedier learning. African Journal of Range & Forage Science 21: 69–78.
Marshall, N.A., I.J. Gordon, and A.J. Ash. 2011. The reluctance of resource-users to adopt seasonal climate forecast to enhance resilience to climate variability on the rangelands. Climatic Change 107: 511–529.
May, R.M. 1977. Thresholds and breakpoints in ecosystems with a multiplicity of stable states. Nature 269: 471–477.
Muthoni, F.K., T.A. Groen, A.K. Skidmore, and P. van Oel. 2014. Ungulate herbivory overrides rainfall impacts on herbaceous regrowth of residual biomass in a key resource area. Journal of Arid Environments 100–101: 9–17.
Nelson, D.R., W.N. Adger, and K. Brown. 2007. Adaptation to environmental change: Contributions of a resilience framework. Annual Review of Environment and Resources 32: 395–419.
NRC (National Research Council). 1994. Rangeland health: New methods to classify, inventory, and monitor rangelands. Washington, DC: National Academy Press. 180 p.
Olsson, L., A. Jerneck, H. Thoren, J. Persson, and D. O’Byrne. 2015. Why resilience is unappealing to social science: Theoretical and empirical investigations of the scientific use of resilience. Science Advances 1(4), e1400217.
Ostrom, E. 1990. Governing the commons: The evolution of institutions for collective action. New York: Cambridge University Press.
Petraitis, P. 2013. Multiple stable states in natural ecosystems, 188. Oxford, UK: Oxford University Press.
Pickett, S.T.A., and R.S. Ostfeld. 1995. The shifting paradigm in ecology. In A new century for natural resource management, ed. R.L. Knight and S.F. Bates, 261–278. Washington, DC: Island Press.
Sayre, N.F., R.R.J. McAllister, B.T. Bestelmeyer, M. Moritz, and M.D. Turner. 2013. Earth stewardship of rangelands: Coping with ecological, economic and political marginality. Frontiers in Ecology and the Environment 11: 348–354.
Scheffer, M., and S. Carpenter. 2003. Catastrophic regime shifts in ecosystems: Linking theory to observation. Trends in Ecology and Evolution 18: 648–656.
Scheffer, M., S.R. Carpenter, T.M. Lenton, J. Bascompte, W. Brock, et al. 2012. Anticipating critical transitions. Science 338: 344–348.
Smith, E.L. 1988. Successional concepts in relation to range condition assessment, 113–133. Netherlands: Springer.
SRM Task Group (Society for Range Management Task Group on Unity in Concepts and Terminology Committee, Society for Range Management). 1995. New concepts for assessment of rangeland condition. Journal of Range Management 48: 271–282.
Stafford Smith, D.M., G.M. McKeon, I.W. Watson, B.K. Henry, G.S. Stone, W.B. Hall, and S.M. Howden. 2007. Learning from episodes of degradation and recovery in variable Australian rangelands. Proceedings of the National Academy of Sciences USA 104: 20690–20695.
Standish, R.J., R.J. Hobbs, M.M. Mayfield, B.T. Bestelmeyer, et al. 2014. Resilience in ecology: Abstraction, distraction, or where the action is? Biological Conservation 177: 43–51.
Stringham, T.K., W.C. Krueger, and P.L. Shaver. 2003. State and transition modeling: An ecological process approach. Journal of Range Management 56: 106–113.
Strunz, S. 2012. Is conceptual vagueness an asset? Arguments from philosophy of science applied to the concept of resilience. Ecological Economics 76: 112–118.
Suding, K.N., and R.J. Hobbs. 2009. Threshold models in restoration and conservation. Trends in Ecology and Evolution 24: 271–279.
von Wehrden, H., J. Hanspach, P. Kaczensky, J. Fischer, and K. Wesche. 2012. Global assessment of the non-equilibrium concept in rangelands. Ecological Applications 22: 393–399.
Walker, B., and D. Salt. 2012. Resilience practice: Building capacity to absorb disturbance and maintain function, 226. Washington, DC: Island Press.
Walker, B., S. Carpenter, J. Anderies, N. Abel, et al. 2002. Resilience management in social-ecological systems: A working hypothesis for a participatory approach. Conservation Ecology 6(1):14 [online]. Retrieved from www.consecol.org/vol6/iss1/art14.
Walker, B., C.S. Holling, S.R. Carpenter, and A. Kinzig. 2004. Resilience, adaptability and transformability in social–ecological systems. Ecology & Society 9(2):5. Retrieved from http://www.ecologyandsociety.org/vol9/iss2/art5.
Walker, B.H., S.R. Carpenter, J. Rockstrom, A.-S. Crepin, and G.D. Peterson. 2012. Drivers, “slow” variables, “fast” variables, shocks, and resilience. Ecology & Society 17(3): 30.
Watson, I.W., D.G. Burnside, and A.M. Holm. 1996. Event-driven or continuous; which is the better model for managers? Australian Rangeland Journal 18: 351–369.
Westoby, M., B.H. Walker, and I. Noy-Meir. 1989. Opportunistic management for rangelands not at equilibrium. Journal of Range Management 42: 266–274.
Wiens, J.A. 1984. On understanding a nonequilibrium world: Myth and reality in community patterns and processes. In Ecological communities: Conceptual issues and the evidence, ed. D.R. Strong, D. Simberloff, L. Abele, and A.B. Thistle, 439–458. New Jersey: Princeton University Press.
Wu, J., and O.L. Loucks. 1995. From balance of nature to hierarchical patch dynamics: A paradigm shift in ecology. Quarterly Review of Biology 70: 439–466.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
This chapter is distributed under the terms of the Creative Commons Attribution-Noncommercial 2.5 License (http://creativecommons.org/licenses/by-nc/2.5/) which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
The images or other third party material in this chapter are included in the work’s Creative Commons license, unless indicated otherwise in the credit line; if such material is not included in the work’s Creative Commons license and the respective action is not permitted by statutory regulation, users will need to obtain permission from the license holder to duplicate, adapt or reproduce the material.
Copyright information
© 2017 The Author(s)
About this chapter
Cite this chapter
Briske, D.D., Illius, A.W., Anderies, J.M. (2017). Nonequilibrium Ecology and Resilience Theory. In: Briske, D. (eds) Rangeland Systems. Springer Series on Environmental Management. Springer, Cham. https://doi.org/10.1007/978-3-319-46709-2_6
Download citation
DOI: https://doi.org/10.1007/978-3-319-46709-2_6
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-46707-8
Online ISBN: 978-3-319-46709-2
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)