
Abstract
The most recent Ebola outbreak in West Africa highlighted critical weaknesses in the medical infrastructure of the affected countries, including effective diagnostics tools, sufficient isolation wards, and enough medical personnel. Here, we develop and analyze a mathematical model to assess the impact of early diagnosis of pre-symptomatic individuals on the transmission dynamics of Ebola virus disease in West Africa. Our findings highlight the importance of implementing integrated control measures of early diagnosis and isolation. The mathematical analysis shows a threshold where early diagnosis of pre-symptomatic individuals, combined with a sufficient level of effective isolation, can lead to an epidemic control of Ebola virus disease.
Diego Chowell* and Muntaser Safan* both the authors contributed equally to this chapter
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others

Keywords
1 Introduction
The Ebola viral strains are re-emerging zoonotic pathogens and members of the Filoviridae family consisting of five distinct species: Bundibugyo, Cotes d’Ivoire, Reston, Sudan, and Zaire with a high case-fatality rate in humans [1]. Filoviruses are long filamentous enveloped, non-segmented, single-stranded viruses, consisting of a negative-sense RNA genome [2]. Each Ebola species genome encodes seven linearly arranged genes: nucleoprotein (NP), polymerase cofactor (VP35), matrix protein (VP40), glycoprotein (GP), replication-transcription protein (VP30), matrix protein (VP24), and RNA-dependent RNA prolymerase (L) [2]. While there are no proven effective vaccines or effective antiviral drugs for Ebola, containing an outbreak relies on contact tracing and on early detection of infected individuals for isolation and care in treatment centers [2]. The most recent Ebola outbreak in West Africa, which began in December 2013, due to the Zaire strain, demonstrated several weaknesses in the medical infrastructure of the affected countries, including the urgent need of effective diagnostics, which have a fundamental role in both disease control and case management.
The Ebola virus is transmitted as a result of direct contact with bodily fluids containing the virus [3]. The virus enters via small skin lesions and mucus membranes where it is able to infect macrophages and other phagocytic innate immune cells leading to the production of a large number of viral particles [2]. The macrophages, monocytes, and dendritic cells infected in the early stage of the disease serve to spread the virus throughout the organs, particularly in the spleen, liver, and lymph nodes [2]. Consequently, critically ill patients display intensive viremia [4]. Recognizing signs of Ebola viral disease is challenging because it causes common non-specific symptoms such as fever, weakness, diarrhea, and vomiting, and the incubation period typically lasts 5–7 days [3]. Therefore, functioning laboratories and effective point-of-care diagnostic tests are critically needed in order to minimize transmission, allow better allocation of scarce healthcare resources, and increase the likelihood of success of antiviral treatments as they are developed [5].
There is an ongoing effort in place to improve Ebola diagnostics, primarily to detect the disease early. Currently, the cost and difficulty of testing limit diagnostic facilities to small mobile laboratories or centralized facilities with turnaround times measured in days rather than in a few hours, meaning that diagnosis is largely used to confirm disease. Ebola diagnosis can be achieved in two different ways: measuring the host-specific immune response to infection (e.g. IgM and IgG antibodies) and detection of viral particles (e.g. ReEBOV Antigen Rapid Test Kit for VP40), or particle components in infected individuals (e.g. RT-PCR or PCR). The most general assay used for IgM and IgG antibody detection are direct ELISA assays. Considering the physiological kinetics of the humoral immune system as well as impaired antigen-presenting cell function as a result of viral hemorrhagic fever, antibody titers are low in the early stages and often undetectable in severe patients prior to death [6]. This leaves polymerase chain reaction (PCR) for antigen detection as a viable option for early diagnostic assays. PCR is a chemical reaction that amplifies pieces of a virus’s genes floating in the blood by more than a millionfold, which makes detection of pre-symptomatic individuals likely identifiable. Indeed, a research article published in 2000, illustrates the power of this technology to detect Ebola virus in humans in the pre-symptomatic stage [7]. In this study, 24 asymptomatic individuals who had been exposed to symptomatic Ebola patients were tested using PCR. Eleven of the exposed patients eventually developed the infection. Seven of the 11 tested positive for the PCR assay. And none of the other 13 did.
In this chapter, we extend the work presented in [8]. Here, we have developed and analyzed a mathematical model to evaluate the impact of early diagnosis of pre-symptomatic individuals on the transmission dynamics of Ebola virus disease in West Africa, under the assumption that the disease is maintained possibly at very low levels due to the deficiencies in health systems and our incomplete understanding of Ebola infection as illustrated by the case of Pauline Cafferkey. Therefore, eliminating Ebola may require a more sustained and long-term control effort.

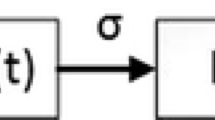
Compartmental model showing the transition between model states
2 Model Formulation
The total population is assumed to be classified into six mutually independent subgroups: susceptible S(t), non-detectable latent \(E_1(t)\), detectable latent \(E_2(t)\), infectious I(t), isolated J(t), and recovered R(t) individuals. Table 1 shows the state variables and their physical meaning. The transition between all these states is shown in Fig. 1. And model parameters and their description are presented in Table 2. Parameter values have been obtained from previous studies [9, 10].
It is assumed that individuals are recruited (either through birth or migration) into the susceptible class at a rate \(\varLambda \) and die naturally with rate \(\mu \). Susceptible individuals get infected due to successful contacts with infectious or not perfectly isolated infected individuals at rate \(\lambda \). As a consequence, they become latent undetectable, who develop their state of infection to become latent detectable at rate \(\kappa _1\). We assume that the latent detectable class represent individuals whose viral load is above the detection limit of the PCR-based diagnostic test [7, 11]. Latent detectable individuals either are diagnosed and get isolated with probability \(f_T\) or develop symptoms to become infectious, who sequentially either get isolated at rate \(\alpha \), or are removed from the system by recovery or Ebola-induced death at rate \(\gamma \). It is assumed here that Ebola-induced deaths occur for the infectious individuals with probability \(q_1\). Similarly, isolated individuals leave their class at rate \(\gamma _r\), by either dying due to Ebola with probability \(q_2\), or they get recovered and become immune. It is assumed that isolation is partially effective so that successful contacts with susceptible individuals may lead to infection with probability r; this parameter is a measure of isolation effectiveness of infectious individuals. Thus, the force of infection is given by
The assumptions mentioned above lead to the following model of equations
where
is the total population size at time t. On adding all equations of system (2) together, we get
3 Model Analysis
3.1 Basic Properties
Since model (2) imitates the dynamics of human populations, all variables and parameters should be non-negative. Thus, following the approach shown in appendix A of [12], we show the following result.
Theorem 1
The variables of model (2) are non-negative for all time.
Lemma 1
The closed set
is positively invariant for model (2) and is absorbing.
Proof: Equation (3) implies that
It follows from (4) that
and from (5) that
If we assume \(N(0) > \varLambda /\mu \), then \(dN/dt < 0\) and therefore (based on inequality (6)), N(t) decreases steadily until reaching \(\varLambda /\mu \) when t tends to \(\infty \). Similarly, if we assume \(N(0) < \varLambda /(\mu + q_1\gamma + q_2\gamma _r)\), then \(dN/dt > 0\) and therefore (based on inequality (7)), N(t) increases steadily until reaching a maximum at \(\varLambda /(\mu + q_1\gamma + q_2\gamma _r)\) when t tends to \(\infty \). It remains to check the case if N(0) lies in the phase between \(\varLambda /(\mu + q_1\gamma + q_2\gamma _r)\) and \(\varLambda /\mu \). To this end, both inequalities (6) and (7) are combined together to get
On taking the limit when t tends to \(\infty \), we find that N(t) remains within the same phase. Thus, the set \(\varOmega \) is positively invariant and absorbing.
3.2 Equilibrium Analysis
3.2.1 Ebola-Free Equilibrium and the Control Reproduction Number \(\mathscr {R}_c\)
It is easy to check that model (2) has the Ebola-free equilibrium
where the prime “ \({}^{\prime }\) ” means vector transpose.
The basic reproduction number, \(\mathscr {R}_0\), is a measure of the average number of secondary cases produced by a typical infectious individual during the entire course of infection in a completely susceptible population and in the absence of control interventions [13, 14]. On the other hand, the control reproduction number, \(\mathscr {R}_c\), quantifies the potential for infectious disease transmission in the context of a partially susceptible population due to the implementation of control interventions. When \(\mathscr {R}_c > 1\), the infection may spread in the population, and the rate of spread is higher with increasingly high values of \(\mathscr {R}_c\). If \(\mathscr {R}_c < 1\), infection cannot be sustained and is unable to generate an epidemic. For our model, \(\mathscr {R}_c\) is computed using the next generation matrix approach shown in [15]. Accordingly, we compute the matrices \(\mathbf {F}\) (for the new infection terms) and \(\mathbf {V}\) (for the transition terms) as
Thus, the control reproduction number is given by
where \(\rho \) is the spectral radius (dominant eigenvalue in magnitude) of the matrix \(\mathbf {F}\mathbf {V}^{-1}\) and
is the basic reproduction number for the model.
The local stability of the Ebola-free equilibrium, \(E_0\), for values of \(\mathscr {R}_c < 1\) is established based on a direct use of Theorem 2 in [15]. We summarize our result in the following lemma.
Lemma 2
The Ebola-free equilibrium \(E_0\) of model (2) is locally asymptotically stable if and only if \(\mathscr {R}_c < 1\).
3.2.2 Ebola-Endemic Equilibrium
On putting the derivatives in the left hand side of (2) equal zero and solving the resulting algebraic system with respect to the variables \(\bar{S}, \bar{E}_1, \bar{E}_2, \bar{I}, \bar{J}\), and \(\bar{R}\), we obtain
where
is the equilibrium force of infection. On substituting from (11) into (12) and simplifying (with the assumption that \(\lambda \ne 0\)), we get
where
Hence, the Ebola-endemic equilibrium is unique and we show the following lemma.
Lemma 3
Model (2) has a unique endemic equilibrium that exists if and only if \(\mathscr {R}_c > 1\).
3.3 Normalized Sensitivity Analysis on \(\mathscr {R}_c\)
In considering the dynamics of the Ebola system (2), we conduct normalized sensitivity analysis on \(\mathscr {R}_c\) to determine the impact of parameter perturbations on the transmission dynamics of the system. By computing the normalized sensitivity indices, we consider the percent change in the output with respect to a percent change in the parameter input. Those parameters with the largest magnitude of change impact the compartment model the most; the sign indicates whether the change produces an increase or a decrease on \(\mathscr {R}_c\). The normalized sensitivity indices for \(\mathscr {R}_c\) are calculated by taking the partial derivative of \(\mathscr {R}_c\) with respect to each parameter and multiply the derivative with the ratio of the parameter to \(\mathscr {R}_c\). This value represents the percent change in \(\mathscr {R}_c\) with respect to a 1 % change in the parameter value [16].
We use the parameters values from Table 2 to study the sensitivity of \(\mathscr {R}_c\) to each parameter. We compute normalized sensitivity analysis on all parameters, but we just consider the impact of parameters that are the most sensitive: \(\beta , r, \ell , \gamma _r, \gamma , \alpha \), and \(f_T\). The other parameters (\(\mu , \kappa _1\), and \(\kappa _2\)) have a very low impact, namely less than \(0.001\,\%\). The numerical simulations to the sensitivity of \(\mathscr {R}_c\) with respect to each of the most sensitive parameters are given in Table 3, for two different levels of isolation effectiveness (\(r = 0.35\) and \(r = 0.95\)) and two values of \(f_T\) (\(f_T = 0.25\) and \(f_T = 0.75\)), which is the fraction of pre-symptomatic individuals diagnosed and isolated. The other parameter values are kept as shown in Table 2.
In the case of high isolation effectiveness (\(r = 0.95\)), simulations show that both the removal rate, \(\gamma _r\), of isolated individuals and the relative transmissibility parameter \(\ell \) of isolated individuals with respect to infectious individuals are the least sensitive parameters (with \(0.053\,\%\) change of \(\mathscr {R}_c\)), while the parameter of isolation effectiveness, r, is the most sensitive one, where a \(1\,\%\) increase in r causes a \(1.014\,\%\) reduction in the value of \(\mathscr {R}_c\). Also, the rate at which infectious individuals get isolated, \(\alpha \), and the fraction of pre-symptomatic individuals detected and isolated, \(f_T\), impact negatively on the level of \(\mathscr {R}_c\), where a \(1\,\%\) percent increase in the value of \(f_T\) causes approximately a \(0.31\,\%\) decline in the value of the reproduction number \(\mathscr {R}_c\). Thus, as pre-symptomatic individuals are diagnosed and as isolation is highly effective, the number of available infectious individuals who are capable of transmitting Ebola decreases and therefore, the reproduction number decreases. Also, the removal (by recovery or Ebola-induced death) rate \(\gamma \) of infectious individuals affects negatively on \(\mathscr {R}_c\). Hence, for the case of highly effective isolation, the parameters concerning early diagnosis and isolation have a significant impact on the reproduction number.
This percent impact of the parameters on \(\mathscr {R}_c\) remains so as long as isolation is highly effective. However, if the effectiveness of isolation is low, in the sense that all parameter values are kept the same except the value of the parameter r, which is reduced to 0.35, then we get the results presented in Table 3. In this case, both the relative transmissibility \(\ell \) and the removal rate of isolated individuals, \(\gamma _r\), are the second most sensitive parameters, after \(\beta \) which is the most impactful one. Also, \(\ell \) became more sensitive than r. The implication is that, when isolation is less effective, there exists the possibility for isolated people to make successful contacts with susceptible individuals and therefore the possibility of causing new infections increases. This causes an increase in the reproduction number. Also, it is noted that the effect of \(f_T\) and \(\alpha \) is reduced, which means that diagnosing and isolating infected individuals becomes a weak strategy if the effectiveness of isolation is low.
On repeating the previous analyses, but this time for a higher value of \(f_T\) (\(f_T = 0.75\)), we obtain the results shown in Table 3. In comparison to the scenario when \(f_T = 0.25\), the simulations show that increasing the fraction of pre-symptomatic individuals who are diagnosed and isolated, \(f_T\), increases the percent impact of the parameters \(r, \ell , \gamma _r,\) and \(f_T\), and decreases the percent impact of the parameters \(\gamma \) and \(\alpha \), on the value of the control reproduction number \(\mathscr {R}_c\).

Impact of early detection of pre-symptomatic individuals on the value of \(\mathscr {R}_c\)
3.4 Impact of Early Detection and Isolation on the Value of \(\mathscr {R}_c\)
To study the impact of early detection of pre-symptomatic individuals and isolation on the reproduction number, we first depict \(\mathscr {R}_c\) as a function of \(f_T\), for different levels of isolation effectiveness r. Figure 2 shows that the control reproduction number declines as the proportion, \(f_T\), of pre-symptomatic individuals, who get diagnosed and isolated, increases. Simulations are done using parameter values from Table 2, but for three different values of r. It further shows that the curve corresponding to a low and an intermidate value of isolation effectivenes r (e.g. \(r = 0.35\) for the solid curve and \(r = 0.65\) for the dashed curve) hits \(\mathscr {R}_c = 1\) at some critical value of \(f_T\) (say \(f_T^{\star }\)), while for the high value of r (\(r = 0.95\)), it never hits the critical threshold \(\mathscr {R}_c = 1\), as the curve is totally below the critical threshold. This indicates that for a high effectiveness of isolation, the control reproduction number is less than one and therefore the infection dies out. Analytically, the exact form of \(f_T^{\star }\) is
The critical proportion \(f_T^{\star }\) represents the minimum proportion of pre-symptomatic individuals who are detected and get isolated to ensure an effective control of Ebola. This critical value remains feasible as long as the following inequality holds
If we keep all parameters fixed except r, then condition (15) could be rewritten in a more convenient form
This gives the minimum level of effectiveness of isolation required to obtain an isolation and early diagnosis-based control strategy for Ebola tranmission.

Impact of isolating infectious individuals on the value of \(\mathscr {R}_c\)
Now, we could also ask a similar question on the role of isolating infectious individuals to contain Ebola transmission. Figure 3 shows the impact of changing the rate at which infectious individuals get isolated, \(\alpha \), on \(\mathscr {R}_c\), for the same three different levels of isolation effectivenes, as used above. The analysis shows that it is possible to control the epidemic if and only if \(\alpha > \alpha ^\star \), where
and with the implementation of condition (15).
4 Discussion and Conclusion
The Ebola epidemic has shown us major weaknesses not only in health systems in West Africa, but also in our global capacity to respond early to an outbreak with effective diagnostic capacities. After multiple outbreaks of infectious diseases, from severe acute respiratory syndrome (SARS) to Middle East respiratory syndrome coronavirus (MERS-CoV), we still do not have effective diagnostic tools to rapidly respond to a number of potential epidemics. The main reason why we lack of such diagnostic preparedness against infectious diseases is because of the lack of a financed global strategy that can be implemented ahead, rather than during an epidemic. This strategy must primarily focus on two critical aspects: First, a continuous interaction between the field to detect small outbreaks and collect samples, and reference laboratories with advanced sequencing tools to identify the pathogen. Second, the need of assay development for early diagnosis, their regulatory approval, and a plan of implementation in anticipation of an outbreak.
Here, motivated by some studies showing that PCR assay can detect Ebola virus in both humans and non-human primates during the pre-symptomatic stage [7, 11], we have developed and analyzed a mathematical model calibrated to the transmission dynamics of Ebola virus disease in West Africa to evaluate the impact of early diagnosis of pre-symptomatic infections . In the absence of effective treatments and vaccines, our results show the importance of implementing integrated control measures of early diagnosis and isolation. Importantly, our analysis identifies a threshold where early diagnosis of pre-symptomatic individuals, combined with a sufficient level of effective isolation, can lead to an epidemic control of Ebola virus disease. Furthermore, the need to incorporate vital dynamics is justified by our still limited understanding of Ebola infection including whether or not Ebola virus may persist among recovered individuals. The use of \(\mathscr {R}_c\) in this context reflect our view that control measures should be sustainable and not just in response to an outbreak.
References
Kugelman, J.R., Sanchez-Lockhart, M., Andersen, K.G., Gire, S., Park, D.J., Sealfon, R., Lin, A.E., Wohl, S., Sabeti, P.C., Kuhn, J.H., Palacios, G.F.: Evaluation of the potential impact of Ebola virus genomic drift on the efficacy of sequence-based candidate therapeutics (2015)
Beeching, N.J., Fenech, M., Houlihan, C.F.: Ebola virus disease. BMJ 349, g7348 (2014)
Fauci, A.S.: Ebola–underscoring the global disparities in health care resources. N. Engl. J. Med. 371(12), 1084–1086 (2014)
McElroy, A.K., Erickson, B.R., Flietstra, T.D., Rollin, P.E., Nichol, S.T., Towner, J.S., Spiropoulou, C.F.: Ebola hemorrhagic fever: novel biomarker correlates of clinical outcome. J. Infect. Dis. jiu088 (2014)
Fink, S.: Ebola drug aids some in a study in West Africa. The New York Times, 4 Feb 2015
Ippolito, G., Feldmann, H., Lanini, S., Vairo, F., Di Caro, A., Capobianchi, M.R., Nicastri, E.: Viral hemorrhagic fevers: advancing the level of treatment. BMC Med. 10(1), 31 (2012)
Leroy, E.M., Baize, S., Volchkov, V.E., Fisher-Hoch, S.P., Georges-Courbot, M.C., Lansoud-Soukate, J., McCormick, J. B., : Human asymptomatic Ebola infection and strong inflammatory response. Lancet 355(9222), 2210–2215 (2000)
Chowell, D., Castillo-Chavez, C., Krishna, S., Qiu, X., Anderson, K.S.: Modelling the effect of early detection of Ebola. Lancet Infect. Dis. 15(2), 148–149 (2005)
Chowell, G., Nishiura, H.: Transmission dynamics and control of Ebola virus disease (EVD): a review. BMC Med. 12, 196 (2014)
Fasina, F., Shittu, A., Lazarus, D., et al.: Transmission dynamics and control of Ebola virus disease outbreak in Nigeria, July to September 2014. Euro Surveill. 19, 20920 (2014)
Qiu, X., Wong, G., Audet, J., et al.: Reversion of advanced Ebola virus disease in nonhuman primates with ZMapp. Nature 514, 4753 (2014)
Thieme, H.R.: Mathematics in Population Biology. Princeton University Press, Princeton (2003)
Brauer, F., Castillo-Chavez, C.: Mathematical Models in Population Biologyand Epidemiology. Springer, Berlin (2011)
Anderson, R.M., May, R.M.: Infectious Diseases of Humans. Oxford University Press, Oxford (1991)
van den Driessche, P., Watmough, J.: Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission. Math. Biosci. 180, 29–48 (2002)
Caswell, H.: Matrix Population Models, 2nd edn. Sinauer Associates, Sunderland (2001)
Acknowledgments
We thank Benjamin Katchman for the helpful discussions about the different types of molecular diagnostics of Ebola.
Author information
Authors and Affiliations
Corresponding authors
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2016 Springer International Publishing Switzerland
About this chapter
Cite this chapter
Chowell, D., Safan, M., Castillo-Chavez, C. (2016). Modeling the Case of Early Detection of Ebola Virus Disease. In: Chowell, G., Hyman, J. (eds) Mathematical and Statistical Modeling for Emerging and Re-emerging Infectious Diseases. Springer, Cham. https://doi.org/10.1007/978-3-319-40413-4_5
Download citation
DOI: https://doi.org/10.1007/978-3-319-40413-4_5
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-40411-0
Online ISBN: 978-3-319-40413-4
eBook Packages: Mathematics and StatisticsMathematics and Statistics (R0)