
Abstract
The North Sea region contains a vast number of lakes; from shallow, highly eutrophic water bodies in agricultural areas to deep, oligotrophic systems in pristine high-latitude or high-altitude areas. These freshwaters and the biota they contain are highly vulnerable to climate change. As largely closed systems, lakes are ideally suited to studying climate-induced effects via changes in ice cover, hydrology and temperature, as well as via biological communities (phenology, species and size distribution, food-web dynamics, life-history traits, growth and respiration, nutrient dynamics and ecosystem metabolism). This chapter focuses on change in natural lakes and on parameters for which their climate-driven responses have major impacts on ecosystem properties such as productivity, community composition, metabolism and biodiversity. It also points to the importance of addressing different temporal scales and variability in driving and response variables along with threshold-driven responses to environmental forces. Exceedance of critical thresholds may result in abrupt changes in particular elements of an ecosystem. Modelling climate-driven physical responses like ice-cover duration, stratification periods and thermal profiles in lakes have shown major advances, and the chapter provide recent achievements in this field for northern lakes. Finally, there is a tentative summary of the level of certainty for key climatic impacts on freshwater ecosystems. Wherever possible, data and examples are drawn from the North Sea region.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others

Keywords
- North Atlantic Oscillation
- Dissolve Organic Matter
- North Atlantic Oscillation Index
- Lake Ecosystem
- Boreal Lake
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1 Introduction
Freshwaters and freshwater biota are highly vulnerable to climate change (IPCC 2013). The various impacts and threats differ substantially between biomes and among geographical regions, and even within geographical regions hydrological factors and catchment properties will be major determinants of the responses in freshwater ecosystems to climate change. Effects on freshwater resources in agricultural or densely populated catchments for example will differ from those in pristine boreal or alpine catchments. In the North Sea region, water scarcity per se is not seen as the most immediate threat of climate change, with the exception of during summer in some areas. Strong flooding events may pose greater challenges. The North Sea region contains a vast number of aquatic systems, including shallow, highly eutrophic water bodies in agricultural areas; systems highly influenced by the input of terrestrially derived, coloured dissolved organic matter in coniferous areas; and large, deep and oligotrophic systems in pristine, sub-alpine areas. Clearly, the review presented in this chapter cannot cover every aspect related to climate and surface waters in the North Sea region; hence a focus on natural lakes and parameters where there is empirical support for climate change effects, as well as on systems for which long-term data sets exist and for which their responses have major impacts on ecosystem properties such as productivity, community composition and biodiversity. Hydrological effects, wetland effects and past climate effects inferred from paleolimnological surveys are covered elsewhere in this report, although there is no doubt that hydrology will also have strong bearings on water chemistry (e.g. by dilution through increased precipitation) and biota (changed fluxes in key elements like nitrogen, phosphorus, carbon, silicon, iron and calcium), dissolved organic matter and pollutants.
Monitoring the impacts of climate change poses challenges because of the many responses within an ecosystem and the spatial variation within the landscape. A substantial body of research demonstrates the sensitivity of freshwater ecosystems to climate forcing and shows that physical, chemical, and biological lake properties respond rapidly to changes in this forcing (Rosenzweig 2007; Adrian et al. 2009; MacKay et al. 2009; Tranvik et al. 2009; IPCC 2013). Fast turnover times from the scale of organisms to entire lake ecosystems are the prerequisite for these rapid changes. Studies of lake ecosystems have provided some of the earliest indications of the impact of current climate change on ecosystem structure and function (Adrian et al. 1995; Magnuson et al. 2000; Verburg et al. 2003) and the consequences for ecosystem services (O’Reilly et al. 2003). Some climate-related signals are highly visible and easily measured in lakes. For instance climate-driven fluctuations in lake level have been observed on a regional-scale across North America (Williamson et al. 2009), and shifts in the timing of ice formation and melt reflect climate warming at a global scale (Magnuson et al. 2000). Other signals may be more complex and difficult to detect in lakes, but may be equally sensitive to climate or more informative regarding impacts on ecosystem services. Long-term historical records and reconstructions from sediment cores have yielded insight into less visible climate-related changes, thus increasing understanding of the mechanisms driving these changes. In particular, paleolimnological records have been critical for reconstructing the climate record over recent geological periods, making it possible to interpret current climate change and predict its future impacts.
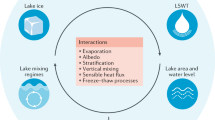
Lake ecosystems are excellent sentinels for current climate change. In this context a sentinel is a lake ecosystem that provides indicators of climate change either directly or indirectly through the influence of climate on the catchment (Fig. 10.1; Carpenter et al. 2007; Adrian et al. 2009; Williamson et al. 2009). The indicators are measureable response variables, such as water temperature, dissolved organic carbon, or phytoplankton composition. Lakes are particularly good sentinels for current climate change for several reasons: they are well-defined ecosystems and studied in a sustained fashion; they respond directly to climate change and incorporate the effects of climate change within the catchment; they integrate responses over time, which can filter out random noise; and they are distributed worldwide and so cover many different geographic locations and climatic regions. However, the large range in lake morphology, geographic location, and catchment characteristics means that broad statements about the ability of lakes to capture the impacts of the current, rapidly changing climate must be made with caution. On the other hand, this also means that there are many different types of sensors in the landscape ranging from small shallow turbid lakes to large, deep, clear lakes that may capture, or provide sentinel information on different aspects of climate change, including temperature and precipitation-related components.

Water and its context: lakes respond to climatic forcing, atmospheric deposition and the properties of their catchments. 1 Atmospheric forcing via temperature, precipitation and deposition of key constituents like nitrogen. 2 The catchment responds to climatic drivers and atmospheric inputs via vegetation and soil processes. 3 This determines the inputs of organic matter, nutrients and key elements determining parameters like retention time, transparency, pH and temperature. 4 The biota respond in terms of phenology, productivity, metabolism, community composition, diversity and food web interactions, to the direct forcing (1), catchment properties (2) and water properties (3)
This chapter focuses on both the direct and indirect (e.g. via the catchment) effects of climate change, as well as on internal physical, chemical and biological processes and on the role of temporal scale. Wherever possible, data and examples are drawn from the North Sea region, but for important phenomena where North Sea studies are not available, information is ‘borrowed’ from other parts of the world.
2 Climate Warming and Impacts on Lake Physics
Data-based studies on the impact of climate change on lake temperature began in the early 1990s with the seminal studies of Magnuson et al. (1990) and Schindler et al. (1990). The former demonstrated that near-surface lake temperatures in Wisconsin fluctuate coherently in response to regional climatic forcing, while the latter demonstrated that lake temperatures in the Canadian Experimental Lakes Area were undergoing a long-term increase. Since the 1990s, an increasing number of studies have demonstrated that near-surface lake temperatures can fluctuate coherently over several hundred kilometres (e.g. Benson et al. 2000; Livingstone and Padisák 2007; Livingstone et al. 2010a), and that individual lakes in many parts of the world have been undergoing long-term warming at all depths. Lake warming has been demonstrated in Europe (e.g. Livingstone 2003; Salmaso 2005), North America (e.g. Arhonditsis et al. 2004; Coats et al. 2006; Austin and Colman 2008), East Africa (e.g. O’Reilly et al. 2003; Verburg et al. 2003), Siberia (Hampton et al. 2008) and Antarctica (Quayle et al. 2002). A recent study based on satellite thermal infrared images from 1985 onwards (Schneider and Hook 2010) confirmed that lake surface temperatures have been undergoing a long-term increase over large areas of the northern hemisphere. In an extensive world wide survey of lake's summer surface temperatures O’Reilly et al. (2015) found an average warming trend of 0.34 °C per decade for the period 1985 until 2009.
Modelling studies on strictly dimictic lakes, which are ice-covered in winter and mix twice per year, suggest that surface temperatures will increase faster than deep-water temperatures as a result of climate change (Robertson and Ragotzkie 1990; Hondzo and Stefan 1993). In monomictic lakes, which mix once per year, the divergence between surface and deep-water temperatures is likely to be less strong as a result of heat carry-over in winter (Peeters et al. 2002). However, even in such lakes historical data show that near-surface water temperatures are increasing faster than deep-water temperatures, which implies an increase in thermal stability leading to an increase in the duration of stratification in summer and a corresponding decrease in the duration of homothermy in winter and spring (Livingstone 2003).
At high latitudes or high altitudes, where lakes are generally ice-covered for many months of the year, the more frequent occurrence of mild winters associated with climate warming will imply a general decrease in the duration of ice cover and a corresponding increase in the duration of summer stratification. Since the mid-19th century, there has been a general long-term decrease in the duration of ice cover in northern hemisphere lakes at a mean rate of about 1.2 days per decade (Magnuson et al. 2000), with the rate for individual lakes ranging from 0.9 to 1.7 days per decade (Benson et al. 2012). This decline seems to be accelerating: over the last 30 years, the equivalent is 1.6–4.3 days per decade (Benson et al. 2012). In regions with relatively brief or mild winters, where lakes are ice-covered for a comparatively short period—for instance in southern Sweden, Denmark and northern Germany—increasingly milder winters are likely to result in ice cover becoming intermittent or even disappearing (Livingstone and Adrian 2009; Weyhenmeyer et al. 2011). The disappearance of ice cover from a deep lake, implying a shift in mixing regime from dimixis to monomixis (Boehrer and Schultze 2008; Livingstone 2008), is likely to cause a change in its physical response to further climate warming, as mixing will no longer necessarily occur at the temperature of maximum density (4 °C) before ice-on and after ice-off. However, mixing can still occur at temperatures higher than 4 °C. Deep lakes that are already monomictic will experience individual years in which some form of stratification persists throughout the year, reducing the intensity of mixing and inhibiting deep-water renewal. Thus some deep monomictic lakes may show a tendency towards becoming oligomictic; i.e. will not mix fully every year. Shallow polymictic lakes, which lose their winter ice cover, are likely to undergo permanent mixing during winter.
2.1 Lake Water Temperature
Research on lakes within the North Sea region has demonstrated the occurrence of many of the phenomena mentioned in the previous section. In the UK—at the western boundary of the region, where the influence of the North Atlantic is at its greatest—research has focused on the English Lake District, where several decades of data are available from some of the larger lakes. Here, water temperatures both near the surface and in the deep water have been shown to respond coherently to climatic forcing throughout the year (George et al. 2000), with a clear long-term increase recorded in both near-surface temperatures (George et al. 2007a) and deep-water temperatures (Dokulil et al. 2006). Long-term increases in surface water temperatures have also been recorded in lakes in Sweden, at the region’s eastern boundary (Adrian et al. 2009), and in northern Germany, at its southern boundary (Adrian et al. 2006, 2009; Wilhelm et al. 2006). It is thus likely that long-term increases in lake temperature are occurring throughout the North Sea region.
It has become evident that the climatic forcing acting on lakes in the North Sea region is extremely large-scale in nature, with the climate prevailing over the North Atlantic playing a major role in determining the physical behaviour of the lakes. Interannual fluctuations in thermal stratification in the lakes of the English Lake District, for instance, are related to north-south displacements of the Gulf Stream, with the early summer thermocline tending to be shallower and more well-defined when the Gulf Stream has its most northern direction (George and Taylor 1995). This effect appears to be related to the effect of the Gulf Stream on wind speed (George et al. 2007b). However, an even more potent determinant of physical lake behaviour is the climate mode known as the North Atlantic Oscillation (NAO), which governs winter weather in western, northern and central Europe to a very large degree (Hurrell 1995; Hurrell et al. 2003) and which is known to play an important role in determining the behaviour of lakes in this region (Straile et al. 2003). This climate mode, which can be considered a regional manifestation of the Arctic Oscillation (AO) (Thompson and Wallace 1998), is associated with interannual fluctuations in the meridional surface air pressure gradient in the north-east Atlantic (between about 35°N and 65°N). A positive NAO index implies a large meridional air pressure gradient, which results in the strong zonal transport of warm, moist maritime air from the North Atlantic towards north-west Europe. In winter, this implies predominantly mild, wet weather in the North Sea region. When the NAO index is negative, however, the eastward transport of warm, moist air from the North Atlantic is much weaker, implying predominantly cold, dry weather in the North Sea region. In lakes throughout northern and central Europe, surface water temperatures, near-bottom temperatures and the duration of ice cover (Fig. 10.2) are correlated to some extent with the winter NAO index (Blenckner et al. 2007).

Dependence of the number of days of total ice cover on Lake Erken (southern Sweden) and Müggelsee (northern Germany) on the winter NAO index (NAOw) of Hurrell (1995), based on data from winter 1976/77 to winter 2005/06. Interannual variability in NAOw explains 35 % of the interannual variability of the duration of ice cover for Lake Erken and 47 % for Müggelsee. The linear regressions (regression lines illustrated) are significant at p < 0.01 (from Livingstone et al. 2010b)
In the English Lake District, lake surface temperatures in winter are tightly correlated with the winter NAO index (George et al. 2000, 2004b, 2007a) as, but to a lesser extent, are deep-water temperatures (George et al. 2004b), with the highest correlations being observed in the shallower lakes (George et al. 2004b). Mean annual water temperature of Lake Veluwe, a shallow lake in the Netherlands, is correlated with the winter NAO index (Scheffer et al. 2001), as are the surface water temperatures of Vänern, Vättern, and Mälaren, the three largest lakes in Sweden, in spring (Weyhenmeyer 2004). However, in Mälaren, a morphometrically complex lake with many sub-basins, the significance of the correlation varies substantially among the different sub-basins, suggesting that local lake characteristics can modify the effect of large-scale climatic forcing even on surface water temperature, which apart from the timing of ice-off is probably the lake variable least affected by internal lake processes. In Müggelsee, a well-studied, shallow, polymictic lake in northern Germany, several studies have demonstrated the effect of the winter NAO on lake temperatures (Gerten and Adrian 2000, 2001; Straile and Adrian 2000).
In a comparative study of Müggelsee and two neighbouring lakes, Gerten and Adrian (2001) showed that the winter NAO leaves a signal at all depths in lake temperature, but that the temporal persistence of this signal can differ substantially from lake to lake. Near the surface, the NAO signal is in general confined to late winter and early spring. In the deeper water, however, the persistence of the NAO signal depends on the morphometry and mixing characteristics of the lake. The NAO signal persists only through spring in shallow, polymictic Müggelsee, but throughout much of the following summer in the shallow, dimictic Heiligensee, and throughout the whole of summer and autumn in the much deeper, dimictic Stechlinsee. Thus, although an NAO signal is likely to be present to some extent in the temperature of all lakes within the North Sea region, individual lake characteristics are certain to result in a large degree of variability in the strength and persistence of this signal.
In the context of the NAO, one other phenomenon should be mentioned: the late 1980s climate regime shift. In the late 1980s, an abrupt regime shift occurred in the atmospheric, oceanic, terrestrial, limnological and cryospheric systems in many regions of the world. Evidence suggests that this large-scale regime shift involved abrupt changes in the AO and NAO (Alheit et al. 2005; Rodionov and Overland 2005; Lo and Hsu 2010), had a substantial impact on air temperature in northern Europe (Lo and Hsu 2010), and affected fish populations in the North Sea (Reid et al. 2001; Alheit et al. 2005). It is not surprising therefore that a regime shift in lake temperature in the late 1980s was also detected in Müggelsee (Gerten and Adrian 2000, 2001), and it can be fairly confidently hypothesised that a similar regime shift, at least in lake surface temperatures, is likely to have occurred in many lakes within the North Sea region.
In summer, a more regional approach to determining the effects of climatic forcing on lakes is necessary owing to the smaller spatial scales of the weather systems involved. In the case of the UK, the Lamb synoptic weather classification system (Lamb 1950) has proven useful. For both Windermere, the largest lake in the English Lake District, and Lough Feeagh, a lake located near the west coast of Ireland, the highest lake surface temperatures were recorded during a westerly circulation type in winter (corresponding to a positive NAO index), but a southerly circulation type in summer (George et al. 2007b).
On a multi-annual time scale, large-scale regional coherence is greater in winter than in summer. However, on shorter time scales the opposite appears to be the case. Short-term, high-resolution surface temperature measurements in Scottish Highland lochs (Livingstone and Kernan 2009) show a high degree of regional coherence in daily means from late spring to autumn, but much lower coherence in winter and early spring. Short-term regional coherence is high in summer because the surface mixed layer is thin and surface temperatures respond sensitively to climate forcing, and is high in autumn because surface temperatures are dominated by convective cooling, which is governed by regionally coherent air temperature. Short-term coherence is comparatively low in winter because fluctuations in lake surface temperature are small and may be buffered by partial ice cover, and is low in early spring because the lochs warm up and stratify at different times during the season depending on their altitude and distance from the maritime influence of the Atlantic. This latter effect may be important on the western boundary of the North Sea region, because the ameliorating influence of the Atlantic Ocean acts to increase winter surface temperatures, and hence to reduce the duration of inverse stratification or circulation.
Based on observed air temperature and physical lake characteristics, George et al. (2007b) were able to model well the surface temperatures of the lakes of the English Lake District. Using a regional climate model (RCM) driven by the SRES A2 scenario, they also projected lake surface temperatures in the 2050s. This showed increases of up to 1.1 °C in winter and up to 2.2 °C in summer, with the greatest increase in winter occurring in the shallowest lake, and the greatest increase in summer occurring in the lake with the shallowest thermocline.
2.2 Lake Ice Phenology
It is likely that the phenology of ice cover on the lakes of the North Sea region will follow the global trend; with ice-on occurring later, ice-off occurring earlier, and a general reduction apparent in the duration of ice cover (Magnuson et al. 2000; Benson et al. 2012). However, several factors will modify this general trend. Because of the approximately sinusoidal form of the air temperature curve, the dates on which the air temperature falls below and rises above 0 °C, which are crucial for the timing of ice-on and ice-off, respectively, are not linear functions of air temperature. Instead, they are arc cosine functions of air temperature, which implies that the sensitivity of the timing of ice-on, the timing of ice-off, and the duration of ice cover are greater in warmer regions than in colder regions, and so will increase as the climate warms (Weyhenmeyer et al. 2004a, 2011; Jensen et al. 2007; Livingstone and Adrian 2009). Thus, in the North Sea region, the impact of climate warming on lake ice phenology will be disproportionately large in those areas where winters are mild or variable and the duration of ice cover on lakes is already short (i.e. the UK, northern France, Belgium, Netherland, Luxemburg, northern Germany and southern Scandinavia) compared to those areas where winters are consistently cold and the duration of ice cover is much longer (i.e. northern Scandinavia). This is despite the IPCC (Intergovernmental Panel on Climate Change) projections implying that climate warming in winter in northern Scandinavia will be stronger than in the rest of the region (Christensen et al. 2007). In Sweden, several decades of historical data confirm that the timing of ice-off on lakes in the south of the country, where winters are relatively mild, has been responding significantly more sensitively to interannual fluctuations in mean winter air temperature than the timing of ice-off on lakes in the north of the country, where winters are longer and more severe (Fig. 10.3; Weyhenmeyer et al. 2004a).

a Dependence of the median date of ice-off on annual mean air temperature (1961–1990) for 70 lakes in Sweden. b Temporal variation in the relationship between the median date of ice-off and annual mean air temperature for 14 lakes in a northern region of Sweden (61–67°N) and 14 lakes in a southern region of Sweden (56–60°N) for the periods 1961–1990 (white) and 1991–2002 (black), showing that the form of the relationship is strongly dependent on latitude but not on the time period chosen. Annual mean air temperatures are calculated from July to June, and the curves illustrated are based on the arc cosine model of Weyhenmeyer et al. (2004a) (Livingstone et al. 2010b; after Weyhenmeyer et al. 2004a)
A further study of ice phenology on 54 Swedish lakes during the 30-year IPCC reference period 1961–1990 showed a statistically significant (p < 0.05) trend towards earlier ice-off in 47 lakes, with the shift towards earlier ice-off varying between 1 and 29 days (Weyhenmeyer et al. 2005). Again, the shift towards earlier ice-off was stronger in the milder, southern part of Sweden than in the colder, northern part. During the IPCC reference period, the mean air temperature of the northern hemisphere increased by 0.4 °C. This resulted in a shift in the timing of ice-off by ~70 days in southern Sweden, but only ~10 days in northern Sweden. Applying the arc cosine model suggests that interannual variability in the duration of lake ice cover will be far greater in southern Scandinavia, Scotland and northern Germany than in northern Scandinavia (Weyhenmeyer et al. 2011). A more sophisticated probability model applied to Müggelsee, which now shows extremely variable, intermittent ice cover, predicts that the percentage of ice-free winters for this lake will increase from ~2 % now to over 60 % by the end of the 21st century (Livingstone and Adrian 2009).
As for winter lake surface temperatures, the duration of ice cover on lakes in the North Sea region also appears to be strongly related to the NAO. In the case of Müggelsee, 47 % of the interannual variability of the duration of ice cover can be explained statistically (p < 0.01) in terms of the interannual variability of the winter NAO index, while for Lake Erken, in east-central Sweden, the equivalent value is 35 % (Fig. 10.2). Even in the UK, where total lake ice cover does not occur frequently, there is evidence that the number of days of partial ice cover is strongly linked to the winter NAO (George et al. 2004a; George 2007).
As well as the duration of ice cover, the timing of both ice-on and ice-off also appear to be determined to some degree by the NAO. For lakes in Sweden, the timing of ice-off is strongly related to the winter NAO while the timing of ice-on shows a weaker relationship to the autumn NAO (Blenckner et al. 2004). These results agree with those of similar studies showing that the winter NAO is an important determinant of ice phenology in the neighbouring Baltic region (Livingstone 2000; Yoo and d’Odorico 2002; Blenckner et al. 2004; George et al. 2004a). As in the case of lake water temperatures, there is evidence to suggest that the late 1980s climate regime shift may have resulted in an abrupt shift in lake ice phenology in the North Sea region: a study of Swedish lakes showed an abrupt shift in 1988/1989 that was substantial in southern Sweden but not in northern Sweden, again emphasising the relative sensitivity of ice phenology in warmer regions to external climatic forcing (Temnerud and Weyhenmeyer 2008).
3 Catchment–Lake Interactions
While lakes are commonly seen as closed entities, which is partly true in terms of populations with low or no immigration or emigration, lakes are strongly influenced by catchment properties such as the proportions of forest, bogs, and arable land that serve as major determinants of element fluxes (and water) to lakes. This is however modified by anthropogenic impacts. For example, acidifying elements such as nitrogen (N) and sulphur (S) will modify the catchment export of ions, dissolved organic matter (DOM) and nutrients, and deforestation or afforestation will also have a major impact on the flux of elements to lakes. This is especially striking for nitrogen, where alpine or otherwise sparsely vegetated catchments may have very low N-retention, while forested catchments may retain almost all nitrate (NO3) and ammonium (NH4) inputs during the growing season (Hessen 1999).
Different catchment properties have vastly different effects on lakes, even within the North Sea region, and these properties and effects will also be differently modified by climate drivers (Fig. 10.1). Although large, deep and oligotrophic lakes are common in alpine areas of the North Sea drainage basin, the North Sea region is dominated by two major types of lake system and it is these that will be most affected by climate change. These are the boreal lakes generally found in forested, less impacted, catchments with limited input of bioavailable nutrients but high loads of humic substances of terrestrial origin. The other lake type occurs in agricultural areas with higher nutrient loads. Both categories of lake/catchment encompass a wide range of size, volume, renewal rates, productivity and community composition.
While catchment responses to climate affect downstream rivers, lakes and ecosystems, hydrology (runoff) also plays a major role both in mobilising and diluting dissolved matter and key elements (such as nitrogen, phosphorus, iron, silicon and calcium). Hydrology also affects the water renewal rate, which has major impacts on the physical, chemical and biological properties of waters. These aspects are less well studied and there is a need for ‘bridging the gap’ between hydrology and especially aquatic biology.
3.1 Boreal Lakes
Rising temperatures and changes in precipitation patterns and amounts will have direct effects on lake ecosystems as well as indirect effects via impacts within the terrestrial catchments (Schindler 2001; Kernan et al. 2010). Atmospheric deposition of nitrogen and sulphur will add to these climate-related impacts on ecosystems (Wright et al. 2010). The well-documented increase in dissolved organic matter in boreal lake ecosystems is one of the most obvious indirect effects (Monteith et al. 2007). This ‘browning’ is mainly due to coloured dissolved organic carbon (DOC) which absorbs light and so affects lake production at all trophic levels (Karlsson et al. 2009; Xenopoulos et al. 2009) as well as species composition (Watkins et al. 2001). The export of DOC from wetlands, bogs and forest via rivers (Fig. 10.4) to lakes and coastal areas is projected to increase, causing reduced primary and secondary production due to light limitation.

Export of coloured dissolved organic carbon (DOC) from wetlands, bogs and forest via rivers (Photo D.O. Hessen)
Changes in temperature and precipitation drive seasonal and interannual variations in the export of DOC and nutrients by affecting soil organic matter mineralisation and the production of DOC and mineral nitrogen species (Kalbitz et al. 2000; Hobbie et al. 2002a, b), as well as hydrology within the catchment. On a longer timescale, climate change can allow forests to expand into areas at present under heathland and alpine vegetation (Hofgaard 1997), thereby affecting nutrient retention, nutrient export and soil organic matter pools and quality (Kammer et al. 2009). A major climatic response expected in boreal, coniferous areas is an increase in the concentrations of terrestrially derived DOM and DOC. For a large dataset of boreal lakes (1041 Swedish lakes along a 13° latitudinal gradient), Weyhenmeyer and Karlsson (2009) demonstrated a nonlinear response of DOC to increasing temperature, with the number of days above 0 °C as the major predictor. Analysis of a correspondingly large dataset of boreal Norwegian watersheds within the North Sea region, indicates that even a moderate (2 °C average, downscaled Hadley scenario) increase in temperature with associated increase in precipitation and vegetation density has the potential to increase DOC export substantially (Fig. 10.5; Larsen et al. 2011a, b). For other North Sea regions, especially the UK, reduced deposition of acidifying compounds (notably sulphur) has also been shown to affect DOC concentrations (Evans et al. 2006), although the Nordic studies strongly suggest that climatic drivers like temperature and/or precipitation are the key drivers of rising DOC concentrations in lakes.

Projected change in total organic carbon concentrations and catchment fluxes at a new steady state based on a downscaled 2 °C increase (Hadley model) (Larsen et al. 2011b)
The ecosystem responses to elevated DOC export may be profound, and may also act in concert with elevated N-deposition and N-export to lakes, which also may increase as a result of elevated N-deposition due to increased precipitation (de Wit et al. 2008; de Wit and Wright 2008; Hessen et al. 2009). This in turn is likely to affect productivity and autotroph community composition; a large survey of US and Scandinavian lakes suggested a transition from N-limitation to P-limitation in many freshwater systems as a result of chronic, elevated N-deposition (Elser et al. 2009).
An increase in allochthonous carbon may fuel microbial production and serve as an alternative food resource for zooplankton (Hessen 1998; Jansson et al. 2007). Changes in water colour and loading of organic carbon may also affect the relative roles of the benthic and pelagic parts of the lake ecosystem, in favour of the pelagic (Ask et al. 2009; Karlsson et al. 2009). However, increased DOC loads are expected to have negative impacts on overall lake productivity. Recent studies from boreal catchments suggest that P-loads especially will decrease in most areas due to increased terrestrial uptake, intensifying P-limitation and in concert with increased light limitation will reduce the overall productivity of boreal freshwaters (Weyhenmeyer et al. 2007; Jones et al. 2012; Thrane et al. 2014). Since DOM may also be a source of phosphorus in pristine catchments, a unimodal response in fish yield over DOC was found for a large number of boreal, Norwegian lakes (Finstad et al. 2014), where an initial stimulus of DOC-associated phosphorus was superseded by light limitation at higher DOC-concentrations.
Hansson et al. (2012) studied how a combination of warming and increased lake colour affects spring plankton phenology and trophic interactions in a mesocosm experiment. Elevated temperature was crossed with increased water colour. Overall, they found temperature to have a stronger effect on phytoplankton and zooplankton abundance than humic substances, but importantly also found synergistic effects between the two stressors. Thermal properties will also be affected by changes in ice cover and basin morphometry (MyLake-model, Saloranta and Andersen 2007), and temperature and reduced nutrients may both induce smaller algal cell size (Daufresne et al. 2009; Hessen et al. 2013) and community changes.
3.2 Lakes in Agricultural Areas
While changes in DOC may be the main climatic response in boreal lakes, they will also be affected by changes in nitrogen, phosphorus and silicon loads. The nutrient impact is far more severe in urban or agricultural lakes, however, and such lakes are also more susceptible to catchment erosion promoted by extreme rainfall as well as reduced periods of snow cover or frozen ground in winter. Land use and agricultural practices such as harvesting and fertiliser applications largely determine loads of nutrients and particulate matter to these lakes, but climatic factors, not least precipitation patterns, will add to these effects. Typically the nitrogen load from agricultural areas is expected to increase, but with a seasonal shift to increased N-export in winter, reflecting both land-use practices and climatic change (Jeppesen et al. 2011).
Phosphorus loads will also increase due to higher winter precipitation and erosion, but again the effects of land-use practices will be superimposed on the effects of climate change. The net impact on lake productivity is unclear, not least because increased turbidity has such a profound impact on lake productivity and stability (Mooij et al. 2005; Jeppesen et al. 2011). Specific ecosystem responses in lakes in agricultural areas are addressed in the following sections.
4 Ecosystem Dynamics
Climate change is expected to alter community structure and ecosystem functioning within lakes worldwide as well as within the North Sea region. Changes may occur in phenology, species and size distribution, food-web dynamics, life-history traits, growth and respiration, nutrient dynamics and ecosystem metabolism. Temperature-induced changes of this type are expected to interact with the increased nutrient flows resulting from enhanced precipitation and runoff (Blenckner et al. 2007; Jeppesen et al. 2010a; Moss et al. 2011).
4.1 Trophic Structure and Function
4.1.1 Fish
Several studies indicate that fish community assemblages, size structure and dynamics will change with global warming. A long-term study of 24 European lakes revealed a decline in the abundance of cold-stenothermal fish species, particularly in shallow lakes, and an increase in the abundance of eurythermal fish species, even in deep, stratified lakes (Jeppesen et al. 2012). This occurred despite a reduction in nutrient loading in most of the case studies, supposedly favouring fish in cold-water and low-nutrient lakes. The cold-stenothermic Arctic charr Salvelinus alpinus has been particularly affected, showing a clear decline in Lake Elliðavatn in Iceland, Lake Windermere in the UK (Winfield et al. 2010), Lake Vättern in Sweden (Jeppesen et al. 2012), and Scandinavian hydroelectric reservoirs (Milbrink et al. 2011).
Other cold-water-adapted species such as coregonids and smelt Osmerus eperlanus are affected at the southern border of their distribution. The harvest of whitefish Coregonus spp. has declined substantially in Lake Vättern in Sweden and Lake Peipsi in Estonia (Kangur et al. 2007; Jeppesen et al. 2012). In the UK and Ireland, a decline in the coregonid pollan Coregonus autumnalis in recent decades has been attributed to changes in temperature (Harrod et al. 2002). A drastic reduction in the population of smelt has occurred in shallow Lake Peipsi as shown from commercial fishing, with the decline particularly strong in years with heat waves (Kangur et al. 2007; Jeppesen et al. 2012). In contrast, the abundance of eurythermal species, including the thermo-tolerant carp Cyprinus carpio (Lehtonen 1996; Jeppesen et al. 2012) is rising in several lakes in the North Sea region (Jeppesen et al. 2012).
It is well-established that high-latitude fish species are not only often larger but also often grow more slowly, mature later, have longer life spans and allocate more energy to reproduction than populations at lower latitudes (Blanck and Lamouroux 2007). Even within species such differences can be seen along a latitudinal gradient (Blanck and Lamouroux 2007) and within North Temperate lakes (Jeppesen et al. 2010b). Thus, changes in life history and size can be expected with warming, and may in fact already have occurred (Daufresne et al. 2009; Jeppesen et al. 2010b, 2012).
4.1.2 Plankton
Changes in fish community structure are likely to have cascading effects in lakes, most implying increased predation on larger zooplankton which in turn means less grazing on phytoplankton and so higher algal biomass per unit of available phosphorus (Lehtonen and Lappalainen 1995; Gyllström et al. 2005; Balayla et al. 2010; Ruuhijärvi et al. 2010; Jeppesen et al. 2010a, b; Meerhoff et al. 2012). Decreasing body size has been suggested as a universal biological response to global warming (Gardner et al. 2011; Hessen et al. 2013). However, there is no consensus about the underlying causality.
The predatory effect due to fish that prefer large zooplankton prey could be reversed or partially reversed if the prevailing or additional predators are invertebrates that prefer small prey. In this case, stronger predation at higher temperature would lead to a stronger removal of small species. A shift towards smaller species can also result from stronger resource competition under higher temperatures and competitive advantage for smaller species. Stronger invertebrate predation at higher temperatures has been suggested, particularly for primary producers, because heterotrophic metabolic rates increase faster with rising temperature than photosynthesis (Yvon-Durocher et al. 2011).
Further evidence of warming-induced changes in plankton size structure comes from mesocosm studies that mimic British shallow lakes. In these mesocosms, warming increased the steepness of the plankton community size spectrum by increasing the prevalence of small organisms, primarily within the phytoplankton assemblage. Mean and maximum size of phytoplankton was reduced by about an order of magnitude. The observed shifts in phytoplankton size structure were reflected in changes in phytoplankton community composition, while zooplankton taxonomic composition remained unaffected by warming (Yvon-Durocher et al. 2011). See Sect. 10.6 for more information on responses of lake plankton communities in the context of global warming. Weak changes in the species composition of benthic macroinvertebrates following shifts towards warmer water temperatures were found in Swedish lakes (Burgmer et al. 2007).
4.1.3 Cyanobacteria Biomass
Higher phytoplankton biomass, particularly higher biomass of cyanobacteria during summer may be expected as a direct response to enhanced water temperatures and as an indirect response to prolonged stratification. Prolonged stratification causes an increase in internal P-loading (Jensen and Andersen 1992; Søndergaard et al. 2003; Wilhelm and Adrian 2008; Wagner and Adrian 2009b), boosting the decomposition of organic matter and thus oxygen depletion at the water-sediment interface, which further exacerbates the P-release from the sediment (Søndergaard et al. 2003). In polymictic lakes, for example, climate warming extended the periods of stratification, and this lengthening of stratified periods led to more frequent and/or stronger internal nutrient pulses between stratified and mixed periods which again promoted cyanobacteria proliferation during summer (Wilhelm and Adrian 2008; Wagner and Adrian 2009b). In dimictic lakes, on the other hand, longer periods of summer stratification may cause longer periods of nutrient limitation in the epilimnion along with higher water temperature and stronger nutrient pulses during the autumn overturn (Adrian et al. 1995; Huisman et al. 2004; Mooij et al. 2005; Elliott et al. 2006; Jöhnk et al. 2008). Immediate access to the hypolimnetic nutrient pools will be limited to migrating species such as buoyant cyanobacteria species, which are often capable of N-fixation (Reynolds 1984; Paerl 1988). Thus, in addition to causing an increase in algal biomass (particularly for cyanobacteria), climate warming may also lead to a change in ecosystem functionality such as a predominance of species capable of N-fixation (Wagner and Adrian 2009b; Huber et al. 2012).
4.1.4 Microbial Loop
How the microbial community and microbial processes are affected by global warming has only been studied in a few large-scale experiments. In mesocosm studies in Denmark the abundance of picoalgae, bacteria and heterotrophic nanoflagellates showed no direct response to experimental warming (Christoffersen et al. 2006). However, experimental warming modified the effects of nutrient addition (Christoffersen et al. 2006; Özen et al. 2013), indicating that interactive effects may be significant in the future given the expected increase in nutrient loading to shallow lakes worldwide (Jeppesen et al. 2009, 2010b; Moss et al. 2011). Increased DOM levels will alter the balance between phytoplankton and heterotrophic bacteria, and thus shift systems (further) towards net heterotrophy (see also Sect. 10.4.4).
4.1.5 Macrophytes
Owing to the climate-induced increase in eutrophication there is an increased likelihood of losing submerged macrophytes and thereby shifting shallow lakes from benthic- to pelagic-dominated systems with a consequent reduction in biodiversity. Indications of such developments are based on long-term data of Danish (Jeppesen et al. 2003) and Dutch shallow lakes (van Donk et al. 2003) as well as modelling studies (Mooij et al. 2007, 2009). Moreover, a dominance of filamentous green algae rather than phytoplankton seems possible under elevated temperatures (Trochine et al. 2011). Space-for-time approaches indicate that macrophyte cover will decrease in lakes with fewer days of ice cover, unless nutrient levels also decline (Kosten et al. 2009). Netten et al. (2011) predicted that milder winters may cause submerged macrophytes with an evergreen overwintering strategy as well as free-floating macrophytes, to outcompete submerged macrophytes that die back in winter. Neophytes such as the free-floating species Salvinia natans and the submerged species Vallisneria spiralis have been shown to be successful under elevated temperatures at the expense of native submerged macrophytes (Netten et al. 2010; Hussner et al. 2014). Mormul et al. (2012) tested the effects of elevated temperature (3 °C) on native and non-native aquatic plant production in mesocosms in combination with ‘browning’ (increased DOC), a potentially important change in the northern hemisphere and found browning to be more important for species invasion than warming. Climate change is likely to have a direct effect if non-native species respond positively to climate change and an indirect effect through species interactions, for example, because browning impairs the growth of native macrophytes and reduces biotic resistance to invasion.
Native competitors of the invasive Elodea canadensis were less successful in browner waters indicating a reduced resistance to invasion. Warming of mesocosms in the UK by 3 °C also significantly altered the proportions of three macrophyte species due to a higher growth rate and higher relative abundance of the neophyte Lagarosiphon major (McKee et al. 2002). A subsequent trial with temperatures 4 °C higher and higher nutrient concentrations resulted in a dominance of floating duckweed Lemna spp. which severely reduced oxygen availability and resulted in a fish kill (Moss 2010). In general, however, there are few long-term studies on the effect of climate change on macrophyte species distribution and invasions, coverage and subsequent effects on other trophic levels in aquatic ecosystems.
For large areas of northern Europe, mass occurrences (‘nuisance growth’) of the macrophyte Juncus bulbosus have been recorded (Moe et al. 2013). The reasons for this phenomenon are still unclear, but nuisance growth has been linked with hydrology, carbon dioxide and elevated N-deposition (Moe et al. 2013).
4.2 Phenology
The most prominent examples of climate-induced changes in lakes are changes in phenology. Coherent changes in ice phenology (see Sect. 10.2.2), and changes in spring and early summer plankton phenology in the North Sea region in recent years have been attributed to climate change (Adrian et al. 1999; Weyhenmeyer et al. 1999; Gerten and Adrian 2000; Straile 2002) as synchronised by large-scale climatic signals such as the NAO (for a review see Blenckner et al. 2007; Gerten and Adrian 2002a; Straile et al. 2003). While indirect temperature effects such as early ice-off, which improves underwater light conditions have brought forward the start of algal bloom development in spring, direct temperature effects caused changes in the timing of rotifer and daphnid spring maxima (Gerten and Adrian 2000; Adrian et al. 2006; Straile et al. 2012) cascading into an earlier clear water phase (Straile 2002). For zooplankton, phenological shifts were most immediate for fast-growing species such as cladocerans and rotifers (but see Seebens et al. 2007), whereas longer-lived plankton such as copepods showed a lag in response. Copepods responded to altered day-length-specific water temperature affecting the timing of the emergence of resting stages in spring (Gerten and Adrian 2002b; Adrian et al. 2006). In addition, warming-induced accelerated ontogenetic development may enable the development of additional generations within a year as has been shown for copepod species (Gerten and Adrian 2002b; Schindler et al. 2005; Adrian et al. 2006; Winder et al. 2009).
4.3 Metabolism
Recent research suggests that global warming tends to shift the metabolic regime of entire lakes toward a dominance of respiration (Allen et al. 2005). This fundamental difference in temperature response between autotrophic and heterotrophic processes may have major implications for biological communities and for ecosystems in general. However, different members of a food web react differently to temperature, for example while cell-division rates may increase with temperature, as may grazing rates and metabolic demands of zooplankton. Thus, the net effect of warming on metabolism is not straightforward also because metabolic rates differ at the species level. As a result, how a changing climate interacts with increased nutrient supply to alter ecosystem metabolism is more uncertain than the change in trophic structure. Although evidence suggests that processes such as deoxygenation, decomposition and denitrification are influenced both by nutrients and by warming, interactions are complex and variable and there are discrepancies in study results about the end result for system components as well as for systems as a whole.
Mesocosm studies in the UK indicated that gross primary production and respiration increase with warming, while results for net production and carbon storage differ. Two experiments (Moss 2010; Yvon-Durocher et al. 2010) showed a marked increase (18–35 %) in the ratio of diurnal community respiration rates and gross photosynthesis for a warming of up to 4 °C. If extrapolated to the large number of shallow northern lakes, this could have immense implications for positive feedbacks in the Earth’s future carbon cycle (Moss 2010). However, these UK experiments were all of short duration (less than one year) and may only have described the transient state after warming, which may lead to overestimation of the net release of carbon. Long-term mesocosm experiments will provide more reliable indications about the net effect of warming on ecosystem metabolism (Jeppesen et al. 2010a; Liboriussen et al. 2011), complemented by long-term research and modelling of whole lake ecosystems (Trolle et al. 2012).
4.4 Greenhouse Gases and Heterotrophy
Ecosystems not only respond as recipients of climatic change, but also provide feedbacks, not least via greenhouse gases such as carbon dioxide (CO2), methane (CH4) and nitrous oxide (N2O). As previously mentioned, the input of organic carbon to lakes through run-off from the catchment has increased in the North Sea region, inducing changes in water colour (Hongve et al. 2004; Erlandsson et al. 2008) (see also Sect. 10.3). Recovery of soils from acidification and changed hydrological conditions are believed to be important factors determining this development (Monteith et al. 2007). On a somewhat longer perspective, climate effects on hydrology and vegetation density may also promote a substantial browning of boreal surface waters (Larsen et al. 2011a, b).
Autotrophs and heterotrophic bacteria compete for the same essential elements, but utilise different energy sources. While DOC is a major energy source for heterotrophic bacteria, it is also an important absorbent in the photosynthetically active part of the spectrum. DOC thus has negative impacts on primary producers both by competing for photons and by stimulating their major nutrient competitors. Increased levels of terrestrially-derived DOC in concert with reduced availability of inorganic phosphorus in lakes may shift systems further towards net heterotrophy and thus a net CO2 release (Sobek et al. 2003; Larsen et al. 2011c).
Increased loads of DOM in boreal lakes would also promote anoxia in the deeper water layers, which promotes CH4 production and its net flux to the atmosphere (Juutinen et al. 2009). Increased N-deposition in concert with elevated export of DOM may promote export of N2O (Hong et al. 2015). For lakes in agricultural systems, increased productivity due to increased nutrient inputs from the catchment may promote the net efflux of CH4 and N2O (Juutinen et al. 2009; Hong et al. 2015), while the net impact of the CO2-balance will also depend on lake morphometry and stratification and so is harder to predict.
5 Biodiversity
Climate conditions are as important for freshwater biodiversity as for terrestrial and marine biodiversity, and consistently explain a major proportion of the geographic variation in species richness of different freshwater taxa such as amphibians, fish, mammals, crayfish and waterbirds (Tisseuil et al. 2013). Even so, other geographic patterns such as the latitudinal gradient in biodiversity is not as strong in freshwater as it is in the marine or terrestrial realms (Hillebrand 2004). However, these geographic trends (often measured at global to regional scales) cannot easily be transferred into predictions on temporal shifts in biodiversity at regional to local scales under climate change. Freshwater systems are particularly vulnerable to climate change for several reasons: owing to the isolated nature of freshwater habitats embedded in a terrestrial matrix; because climate change has direct influences on local temperatures and temperature-associated factors (such as oxygen saturation); and because many freshwater systems already absorb other anthropogenic stressors such as nutrient loading or an altered hydrological regime (Woodward et al. 2010). This section addresses these issues by reviewing existing information on biodiversity shifts in the North Sea region (Sect. 10.5.1) and deriving more general predictions from theoretical and experimental literature identifying research needs in the North Sea region (Sect. 10.5.2).
5.1 Shifts in Biodiversity
Predicting shifts in freshwater biodiversity in the North Sea region is difficult, because few studies have been explicitly conducted in lentic or lotic water bodies in this region. Predictions concerning future biodiversity are often derived from bioclimatic envelope models, which project future range shifts based on the current distribution of species (Parmesan and Yohe 2003). These models typically predict a northward (and often eastward) shift in ranges, such that warm-adapted species expand their ranges, and the ranges of cold-adapted species narrow.
Although changes in marine biodiversity in the North Sea region have frequently been predicted using this approach (Beaugrand et al. 2002), studies for freshwater systems in this region are rare. The species richness of macroinvertebrates across running waters in Europe has been predicted to decline in the southern North Sea countries (UK, Germany, Netherlands, Denmark), but to increase in the North (Norway) (Domisch et al. 2013). Changes in distribution and diversity have already been observed in some freshwater groups, such as odonates in the UK (Hickling et al. 2005). Other diversity shifts caused by local, climate-related range retractions have been reported for fishes in Iceland (Jeppesen et al. 2010b) and crustaceans in Norway (Lindholm et al. 2012). Correspondingly, analyses of long-term monitoring data on community composition revealed shifts in macroinvertebrate assemblages associated with ambient temperatures in Greenland, Iceland, Norway, Denmark and Sweden (Burgmer et al. 2007; Friberg et al. 2013). However, few long-term monitoring data sets still exist for freshwater systems within the North Sea region.
Thus it seems clear that specialised communities in colder regions around the North Sea have a high potential for reduced biodiversity. Macroinvertebrates in glacier-fed river systems will be characterised by lower local species richness and lower beta-diversity if warming leads to glacier retreat (Jacobsen et al. 2012). In boreal regions of northern Europe, the riparian zones of running waters are predicted to be affected by additive or interactive combinations of higher temperature, increased annual discharge but less seasonal variation in runoff, changes in groundwater supply, and altered ice regimes (Nilsson et al. 2013). Potential consequences for biodiversity can be negative or positive. Negative consequences are likely if the riparian zone narrows, such as by hydrologic changes, and thus species richness locally declines. In contrast, higher temperatures might allow invasion of exotic species leading to higher local species richness. Other types of change include altered disturbance regimes (e.g. altered freezing and thawing regimes during winter), which could foster a more dynamic and species-rich riparian vegetation, but also a more specific and species-poor assemblage of stress-tolerant species.
Floodplain systems around the North Sea coast have been massively altered by human regulation of flow regimes and inundation (Tockner et al. 2010). At the same time, they harbour a diverse fauna and flora shaped by the interaction of different climatic, hydrological and biological drivers as well as by the interaction between aquatic and terrestrial ecosystems with respect to the exchange of water, nutrients and organisms. Climate-induced shifts in flow regime are thus of primary importance for biodiversity. European-scale modelling scenarios predict that North Sea region floodplains will experience moderately higher flow levels (contrasting with predictions for other regions, such as the Mediterranean floodplains) (Schneider et al. 2013). Consequences for biodiversity remain largely unknown.
Climate-driven changes in biodiversity are very likely to interact with changes associated with other anthropogenic pressures, such as eutrophication (Moss et al. 2009). Some synergistic effects have been found in UK mesocosm studies (Feuchtmayr et al. 2009). For example, warming promoted increased phosphorus concentrations and the frequency of severe benthic anoxia in the mesocosms, with the potential to exacerbate existing eutrophication problems (McKee et al. 2003).
5.2 Predictions, Theory and Experimental Studies
Although functional aspects of ecosystem impacts (biomass, productivity, element cycling) under climate change can be predicted with some accuracy (see previous sections), information on shifts in freshwater biodiversity in the North Sea region remains vague (Moss et al. 2009). This is due not only to the scarcity of studies themselves, but also to the focus on larger spatial scales (i.e. regional climate-envelope models on range shifts), which makes predictions for local ecosystems difficult. Information from ecological theory and experiments can help to fill this gap by identifying potential changes in freshwater biodiversity and associated pathways (Fig. 10.6). Local attributes may also be superimposed on large-scale patterns. In an analysis of global lake zooplankton data, Shurin et al. (2010) found increased biodiversity in lakes showing greater temperature variation on different time scales (intra- and interannual). A recent review argued strongly for including temperature variability in climate-change experiments (Thompson et al. 2013).

Schematic representation of pathways leading to altered local biodiversity and thus ecosystem function. Main pathways include changes in species pools (species turnover) and on the fundamental as well as realised niches of species
The survival of a species can be directly impaired if temperatures are shifted beyond the fundamental thermal niche, leading to local extinction (Fig. 10.6). Temperature also affects the fundamental niche of a species with respect to other conditions, which are often directly related to temperature (e.g. oxygen saturation, hydrological regimes, solute concentrations). Indirect consequences of temperature change can be seen in freshwaters from around the North Sea, for example with respect to stratification regimes (Wagner and Adrian 2011) or oxygenation (Wilhelm and Adrian 2008). These changes alter the environmental filtering of colonising species, such that local diversity can increase or decrease depending on the number of species that are precluded or enabled to establish viable populations.
The meta-community context (Leibold et al. 2004) reflects the interplay of dispersal and local processes and will become an important tool to predict future changes in aquatic biodiversity. This is even more likely given that much of the regional biodiversity in aquatic systems is contributed by ponds and shallow lakes as these have high beta-diversity due to their often isolated nature (Scheffer et al. 2006) and provide important ecosystem services such as carbon storage due to their high number (Giller et al. 2004; Downing et al. 2006; Tranvik et al. 2009).
At the same time, there is high potential for small isolated systems to lose species with global climate change (Burgmer and Hillebrand 2011) and spatial dynamics are important for maintaining biodiversity in these systems. Meta-community dynamics can also provide spatial insurance for freshwater ecosystems (Loreau et al. 2003). In a meta-community mesocosm study, colonisation from a regional species pool and higher biodiversity positively affected the recovery of the pond ecosystems from heat stress (Thompson and Shurin 2012).
Changes in species turnover with time reflect changes in biodiversity dynamics, i.e. local immigration and extinction. Most experiments suggest a more rapid turnover of species at higher temperatures reflecting an acceleration of colonisation–extinction dynamics. In the thermal effluent of a nuclear power plant, higher temperatures resulted in a faster turnover of species composition but without affecting species richness (Hillebrand et al. 2010). Laboratory experiments also indicated faster change in species composition with increasing temperature (Hillebrand et al. 2012). Analysing temporal turnover in community composition could thus be a better means of identifying climate-induced changes in biodiversity than simple univariate measures of biodiversity such as richness or evenness (Angeler and Johnson 2012). Especially because even major changes such as a shift in dominance between functional groups (e.g. from diatoms to cyanobacteria in phytoplankton) might occur without a change in species richness (Wagner and Adrian 2011).
Temperature also modifies species interactions and their consequences for biodiversity (Fig. 10.6). Increased temperatures are often associated with higher rates of consumption (Hillebrand et al. 2009), which can lead to either lower biodiversity (higher mortality) or higher biodiversity (more consumer-mediated coexistence). Competitive interactions are also strengthened by higher temperatures, leading to more rapid exclusion of inferior species—an effect which was shown to depend on consumer presence (Burgmer and Hillebrand 2011). Not only can the strength of interactions be altered by temperature, but also the temporal match of the interacting species through changes in phenology (Berger et al. 2010). Information on temperature-dependent changes in mutualistic interactions in freshwaters is currently missing.
Both experiments and models indicate that warming-induced shifts in biodiversity have functional consequences for ecosystems (Fig. 10.6), among others with respect to primary production, resource use efficiency and temporal stability of ecosystem functions (Hillebrand et al. 2012; Schabhuettl et al. 2013). In a long-term freshwater phytoplankton experiment, temperature-induced reductions in species richness were associated with lower biomass production, and higher extinction rates were associated with higher variability in biomass production (Burgmer and Hillebrand 2011). Similar strong relationships between diversity and resource use efficiency are found in freshwater field data from the UK and Scandinavia (Ptacnik et al. 2008).
6 Importance of Temporal Scale
Responses to climate change in lake ecosystems operate on various temporal scales. Physical forces such as variation in temperature and mixing regimes span sub-daily to monthly time scales. Organisms differ in their generation time from daily to yearly time scales (Adrian et al. 2009). Thus, to understand the impacts of single climatic forcing events in the context of longer term dynamics it is necessary to consider not only sufficiently long periods (several decades) but also to consider appropriate small temporal scales within the yearly cycle. For example, ecological variables may respond to meteorological forcing only during short critical time windows, or to short-lived exceedance of ecologically-relevant critical thresholds. Thus, annual, seasonal or monthly climate data may not be enough to capture the thermal dynamics to which organisms actually respond (Fig. 10.7; Adrian et al. 2012).

Timing of ecosystem responses to meteorological forcing. Ecological responses are often triggered by changes in critical time windows. The triggering mechanism frequently involves the crossing of critical thresholds in forcing variables (dashed-dotted line), and responses tend to occur with a time lag. In this conceptual sketch, analysis at the monthly timescale (dashed line) would not be sufficient to detect threshold exceedance, in contrast to analysis at the daily timescale (solid line) (Adrian et al. 2012)
Members of the grassroots organisation GLEON (Global Lake Observatory Network; www.gleon.org) or the European project NETLAKE (Networking Lake Observatories in Europe; www.cost.eu/domains_ctions/essem/Actions/ES1201) established an international network of automatic stations in lakes to address dynamics of ecosystem properties at sub-hourly scales.
The following sections focus on the role of temporal scale in climate impact research and provide examples of responses at small (sub-daily) to large (decadal) temporal scales, the role of critical time windows, and the significance of exceeding critical thresholds for lakes within the North Sea region (which also applies for lakes throughout the North Temperate Zone).
6.1 Critical Temporal Scales
The importance of addressing critical temporal scales in climate impact research has been documented for various lakes in the North Sea region (see Adrian et al. 2012; Sect. 10.2). A closer look at sub-hourly measurements showed that the rate of increase in the daily minima (night-time water temperature) exceeded that of the daily maxima (daytime water temperature) (Wilhelm et al. 2006). The consequences of this day-night asymmetry for the biota are unclear, but may contribute to some of the unexplained changes observed in ecosystem dynamics over time. Day-to-day variation in respiration seems to be common in lakes worldwide, including those in the North Sea region (Solomon et al. 2013). Daily variation in gross primary production explained 5–85 % of the daily variation in respiration. Solomon et al. (2013) found respiration to be closely coupled to gross primary production at a diurnal-scale in oligotrophic and dystrophic lakes, but more weakly coupled in mesotrophic and eutrophic lakes.
Known changes in the thermal regime of lakes in the North Sea region (Sect. 10.2) operate over a broad range of temporal scales, and are closely related to lake morphometry: on sub-daily (Wilhelm and Adrian 2008) to weekly scales in polymictic lakes (Wagner and Adrian 2009b), and on weekly to monthly scales in monomictic or dimictic lakes (Gerten and Adrian 2000; Livingstone 2003). While variation in the timing of spring overturn affects underwater light conditions and thus the start of algal growth (Weyhenmeyer et al. 1999; Gerten and Adrian 2000; Peeters et al. 2007), variation in the timing of summer stratification affects water temperature and internal nutrient loading and subsequent plankton development and species composition in productive lakes (Wilhelm and Adrian 2008; Wagner and Adrian 2011). Differences in water temperature between mixed and stratified periods can be up to 5 °C within days or a just few weeks in summer, favouring thermophilic copepod species for example (Wagner and Adrian 2011).
Changes in phenology in abiotic and biotic variables (see Sect. 10.4.2) operate on time scales of weeks (Weyhenmeyer et al. 1999; Gerten and Adrian 2000; Straile et al. 2003). Thus, in terms of their duration, seasons should be defined by cardinal events within the lake itself, rather than by fixed calendar dates. Important markers successfully used to define phenology-adjusted seasons in lakes include temperature thresholds, ice-off dates, the timing of the clear-water phase, and periods of stable thermal stratification (Rolinski et al. 2007; Wagner and Adrian 2009a; Huber et al. 2010).
Wagner et al. (2012) proposed a seasonal classification scheme tuned to specific hydrographic-sensitive phases for dimictic lakes across a latitudinal gradient: inverse stratification (winter), spring overturn, early stratification and the summer stagnation period. They estimated a mean latitudinal shift of 2.2 days per degree of latitude for the start of these sensitive phases. After accounting for latitudinal time shifts, mean water temperatures during the defined hydrographic cycles were similar in lakes spanning the gradient between 47° and 54°N. Adjusting seasons in this way thus enhances the probability of identifying the major driving forces in climate impacts on lake ecosystems.
6.2 Critical Time Windows
Responses to warming trends are often expressed in terms of average changes in temperature on seasonal or annual scales, however species exhibiting short generation times such as planktonic organisms respond only during specific time windows within a season. Shatwell et al. (2008) showed how short time windows can open for cyanobacteria in warm springs in an otherwise diatom-dominated season. They argued that if cyanobacteria attain a critical biomass during that critical time window they can dominate the phytoplankton during summer. In terms of zooplankton, the abundance of cyclopoid (Gerten and Adrian 2002b; Seebens et al. 2009) and calanoid copepods (Seebens et al. 2007) in summer and autumn are determined by conditions in spring—probably related to temperature-induced changes in the emergence of resting stages (Adrian et al. 2006) or short time windows of high food availability which increases offspring survival (Seebens et al. 2009).
Temperature-driven changes in the timing of food availability and of predation by young-of-the-year fish during critical time windows in spring/early summer determined the mid-summer decline in daphnids (Benndorf et al. 2001). More specifically, water temperatures in narrow time windows either before (2.2 weeks) or after the typical clear-water phase (3.2 weeks) affected the start-up populations of summer crustacean zooplankton and explained some of their contrasting success during three hot summers characterised by more or less the same average summer water temperature (Huber et al. 2010).
6.3 Critical Thresholds
Threshold-driven responses to environmental forces have gained attention in ecology because of their seeming unpredictability and their potentially large effects at all levels of ecosystems. The crossing of critical thresholds may result in abrupt changes in particular elements of an ecosystem (Andersen et al. 2009; Scharfenberger et al. 2013) or entire ecosystems—the famous example being the alternative stable states of clear versus turbid lakes (Scheffer and Carpenter 2003). Abrupt changes within ecosystems are already known under warming trends experienced in the recent past for a number of variables spanning abiotic and biotic components, such as nutrients or algal blooms (Wagner and Adrian 2009a; for review see Adrian et al. 2009, 2012). The underlying forces are often unclear, but may involve competition for common resources and the crossing of critical thresholds in the abundance of conspecifics (Scharfenberger et al. 2013) or multiple overlapping environmental forces (Huber et al. 2008).
Critical thresholds are known to have been exceeded within lake ecosystems (Hargeby et al. 2004). For example, Peeters et al. (2007) quantified the exceedance of critical thresholds in spring for several meteorological variables to determine early or late onset of phytoplankton growth in Lake Constance (Germany). In a recent model, Straile et al. (2012) used water temperature phenology as a predictor for Daphnia seasonal dynamics in North Temperate lakes. The day of the year when surface water temperatures reached a threshold of 13 °C explained 49 % of the variability of the timing of the spring Daphnia maximum in two German lakes (Lake Constance, Müggelsee) and in Lake Washington (USA). The Daphnia phenology model also performed well for predicting the timing of the Daphnia maxima in 49 lakes within the northern hemisphere—many located in the North Sea region (Straile et al. 2012). Early spawning of zebra mussel Dreissena polymorpha in Müggelsee was related to early attainment of the same critical water temperature threshold of 13 °C, known to initiate the first spawning event of the year (Wilhelm and Adrian 2007).
Exceeding direct and indirect temperature thresholds (length of thermal stratification) has been shown to trigger processes such as the onset and magnitude of cyanobacteria blooms (Wagner and Adrian 2009a; Huber et al. 2012). Stratification periods of more than three weeks caused a switch from a dominance of non-N-fixing cyanobacteria to a dominance of N-fixing cyanobacteria species, thus affecting not only biomass of cyanobacteria but also ecosystem functioning (Wagner and Adrian 2009a). In addition to warming-related changes in species composition (Adrian et al. 2009), habitat shifts northward toward temperate-zone lakes over the last few decades have been observed for Cylindrospermopsis raciborskii, an invasive freshwater cyanobacterium, originating in the tropics (Padisak 1997). Observations of Cylindrospermopsis raciborskii in pelagic populations were found to be temperature-mediated. Filaments emerged in the pelagic habitat when water temperature rose above 15–17 °C in two north German lakes (Wiedner et al. 2007).
6.4 Extreme Events
Extreme events, which are expected to become more frequent in the future, are often ecologically more relevant than fluctuations in mean climate. For lakes, extreme events principally refer to exceptionally mild winters, summer heat waves, or extreme storms or heavy rainfall events. For example, an extreme event for a lake that typically freezes in winter would be if the lake did not freeze at all. Based on regional climate model forecasts, Livingstone and Adrian (2009) predicted that the percentage of ice-free winters for a lake in northern Germany would increase from about 2 % at present to over 60 % by the end of the century; see Sect. 10.2.2 for more detail on the frequency of extremely late freeze-up, early break-up and short ice duration. These extremes affect thermal stratification patterns and underwater light conditions, with implications for oxygen conditions and phytoplankton development (see Sect. 10.4.2).
Central Europe has recently experienced extreme heat waves, most notably that of summer 2003. Mean air temperature in summer that year exceeded the long-term average by around 3 °C over much of Europe (Schär et al. 2004). Although heat waves are likely to promote cyanobacteria blooms (Jöhnk et al. 2008), water temperatures above average to the same extent in Müggelsee (Germany) in two recent summers (2003 and 2006) resulted in very different situations: a cyanobacteria bloom in 2006 but a record low cyanobacteria biomass in 2003. This difference was due to the thermal stratification pattern being critically intense only in 2006 (Huber et al. 2012).
Summer fish kills and a change in fish community structure have been attributed to summer temperature extremes in combination with eutrophication in shallow Lake Peipsi (Estonia/Russia) related to a decline in near-bottom oxygen conditions and a decrease in water transparency (Kangur et al. 2013). An extreme rainy period in 2000 caused a strong increase in chemical loading, particularly for organic carbon, in Lake Mälaren (the third largest lake in Sweden) followed by an increase in water colour by a factor of 3.4 and a doubling of spring cryptophyte biomass. This increase in algal mass required changes in the treatment of raw water from Lake Mälaren for the drinking water supply of Stockholm city (Weyhenmeyer et al. 2004b). Extreme summer rain events have also altered CO2 and CH4 fluxes in southern Finish lakes, with the systems switching from being a net sink to a net source of CO2 to the atmosphere (Ojala et al. 2011).
Episodic events of extreme wind speed or rain events exceeding two standard deviations of the seasonal means have strong but complex impacts on thermal structure and stability, DOC loading and underwater PAR (photosynthetically active radiation) levels in northern European lakes—the magnitude and direction of change depending on the location of the lake and catchment characteristics (Jennings et al. 2012). A comprehensive summary as to how extreme weather events affect freshwater ecosystems is provided by the British Ecological Society (BES 2013).
6.5 Regime Shifts
While ecosystems may be buffered against single short-lived critical threshold exceedance events, gradual changes over longer periods may cause lake ecosystems to switch abruptly from one state to another. The most likely and widespread climate warming-induced shift in lakes will be shifts in thermal regime (see Sect. 10.2). Climate change alters heat redistribution over time (within the annual cycle) and space (vertically within the water column), and eventually leads to transitions in the seasonal mixing regime of a lake much in the sense of scenarios described by regime shift theory (Scheffer and Carpenter 2003).
On the basis of existing climate scenarios, Kirillin (2010) predicted a shift from a dimictic to a monomictic regime in the majority of European dimictic lakes by the end of the 21st century, with the loss of ice cover in the cold season meaning that winter stratification in these lakes would completely disappear. In summer, climate warming has an opposite, stabilising effect that may eventually lead to the mixing regime shifting to a dimictic regime in hitherto polymictic lakes. The ecological consequences of this type of regime shift may be even more far-reaching than for di-/monomictic transitions, because the abrupt detachment of the nutrient-rich hypolimnion from the euphotic layer is likely to trigger stronger competition between autotrophic species resulting in changes in phytoplankton species composition and ecosystem functionality (Wilhelm and Adrian 2008; Wagner and Adrian 2009a, b, 2011).
7 Modelling
Predicting the fate of freshwater ecosystem processes under climate change is non-trivial, because many physical, chemical and biological processes interact, and may be affected on different spatial and temporal scales by climate forcing. In attempting to account for these complex interactions, mechanistic numerical models continue to play a greater role in hypothesis testing (system understanding) and for predicting the future state of ecosystems given the projections of future climatic forcing according to climate models (Trolle et al. 2012). Thus, the ability to link—and equally importantly to quantify—complex interactions between physical, chemical and biological processes makes models one of the most important tools of modern science, and for the past decade, models have been used extensively, aiming to establish the potential effects of future climate on freshwater ecosystems (Mooij et al. 2007; Trolle et al. 2011; Elliott 2012).
7.1 Linking Physical and Ecological Dynamics
A widely accepted effect of increased climate warming on lakes is increased stability of the water column, which is readily quantified by hydrodynamic models (see Fig. 10.8). Increased stability can cause prolonged periods of stratification, with subsequent effects on biogeochemical cycling, for example by increasing the duration of anoxia in bottom waters and thereby the potential for release of iron-bound phosphorus.

Simulated water temperature for the deep Lake Ravn, Denmark, for current climate (represented by 1989–1993) (a) and future climate (represented by 2094–2098) (b), and simulated oxygen concentration for current climate (c) and future climate (d). Simulations were performed using DYRESM-CAEDYM. Future climate forcing was derived by the Danish Meteorological Institute using the regional HIRHAM model and the SRES A2 scenario for Denmark. Y-axis represent water level (m) (D. Trolle, original)
Climate warming is also expected to result in decreased duration and thickness of ice cover on lakes, and models such as MyLake have recently been applied to provide quantitative estimates of such decreases.
Models with a strong focus on food-web dynamics, such as PCLake (Janse 1997) have also been used to quantify the effects of warming in facilitating lake ecosystems to shift from a clear, macrophyte-dominated state to a turbid, phytoplankton-dominated state, under different combinations of warming scenarios and external nutrient load scenarios (Fig. 10.9). Modelling exercises of this type are readily undertaken using a standard desktop computer, in contrast to testing such scenarios in experimental mesocosm studies, for example, which is extremely time consuming and expensive.

Three scenarios with elevated temperatures (a) simulated using PCLake adapted for shallow Dutch lakes, and evaluated for average summer chlorophyll-a concentration (b) and the percentage of cyanobacteria relative to total phytoplankton biomass (c). The simulations include four scenarios: a control (closed circles), an all year round temperature increase of 3 °C (open circles), an increase in summer maximum temperature of 3 °C but no change in the winter minimum (open diamonds), and an increase in winter minimum temperature of 3 °C, but no change in the summer maximum (open triangles) (adapted from Mooij et al. 2007)
One of the key messages from the model study by Mooij et al. (2007) shown in Fig. 10.9 was that the critical loading of phosphorus to a lake, at which a shift from a clear to a turbid state occurs, is likely to decrease as warming continues. This effectively means, that according to the model, external loading will need to be reduced in the future, if lakes are to retain the ecological quality of present day.
7.2 Predictions Versus Observations
Modelling studies of the long-term effects of climate change on northern hemisphere temperate lakes (e.g. Elliott et al. 2005; Mooij et al. 2007; Trolle et al. 2011) generally imply that overall phytoplankton biomass is likely to increase, and that cyanobacteria will become a more dominant feature of the phytoplankton species composition. This is in line with empirical observations from time series (Jöhnk et al. 2008; Wagner and Adrian 2009a, b; Posch et al. 2012) and with cross-system analyses (Jeppesen et al. 2009; Kosten et al. 2012). However, the effects may be even more severe than suggested by models, as current models do not fully account for structural changes in the lake ecosystems that could occur due to warming. As stated in Sect. 10.4.1 the composition of fish stocks could change towards smaller and faster reproducing fish, implying more predation on zooplankton and so less grazing on phytoplankton (Jeppesen et al. 2010a; Meerhoff et al. 2012). Such changes have yet to be included in dynamic models.
7.3 Linking Landscape Activities and Nutrient Losses
A recent trend is to assemble multidisciplinary modelling teams with expertise both in hydrological and ecological modelling. Coupling hydrological and ecological models (Norton et al. 2012; Nielsen et al. 2013) enables a direct and quantitative link between land use activities and surface water quality. For example, Norton et al. (2012) used a farm-scale nutrient budget model (PLANET; Planning Land Applications of Nutrients for Efficiency) combined with a generic nutrient runoff model (GWLF; Generalized Watershed Loading Function) to estimate nutrient losses to Loweswater, a small lake in northwest England. The simulated nutrient losses were subsequently used as input to an ecological model for the lake, PROTECH (Phytoplankton RespOnses To Environmental CHange). This model-chain has been used to forecast the abundance of different phytoplankton types within the lake in response to a range of catchment management measures. Ongoing research projects are currently expanding such model chains to include the effects of future climate on both hydrology and aquatic ecosystems.
8 Conclusion
Freshwater systems, and notably lakes, are well suited to trace climate-induced effects both directly via changes in ice cover, hydrology and temperature, but also indirectly via biotic communities since they represent closed bodies (aquatic islands in the landscape). Moreover, they also offer strong tools as ‘sentinels’ by integrating changes at the catchment scale that elsewhere would be hard to detect (Adrian et al. 2009). The North Sea region has a wide range of freshwaters that reflect changes related directly and indirectly to climate forcing. This includes a number of long-term monitoring sites, and experimental studies offer insights into climate change in freshwaters—and subsequent impacts on downstream coastal recipients. The major impacts on lakes, both those known to date and those predicted to occur, can be addressed with different levels of certainty (tentatively summarised in Table 10.1). The level of certainty falls substantially, once higher trophic levels and the complexity of trophic interactions are included. Nevertheless, this summary may be used to identify systems and parameters that are already well suited for addressing climate responses, those that need further attention, and those that are likely to need addressing at local scales and where general statements and predictions are hard to achieve.
References
Adrian R, Deneke R, Mischke U, Stellmacher R, Lederer P (1995) A long-term study of Heiligensee (1975-1992). Evidence for effects of climate change on the dynamics of eutrophied lake ecosystems. Arch Hydrobiol 133:315–337
Adrian R, Hintze T, Walz N, Hoeg S, Rusche R (1999) Effects of winter conditions on the plankton succession during spring in a shallow polymictic lake. Freshwater Biol 41:621–632
Adrian R, Wilhelm S, Gerten D (2006) Life-history traits of lake plankton species may govern their phenological response to climate warming. Glob Change Biol 12:652–661
Adrian R, O’Reilly CM, Zagarese H, Baines SB, Hessen DO, Keller W, Livingstone DM, Sommaruga R, Straile D, van Donk E, Weyhenmeyer GA, Winder M (2009) Lakes as sentinels of climate change. Limnol Oceanogr 54:2283–2297
Adrian R, Gerten D, Huber V, Wagner C, Schmidt SR (2012) Windows of change: temporal scale of analysis is decisive to detect ecosystem responses to climate change. Mar Biol 159:2533–2542
Alheit J, Möllmann C, Dutz J, Kornilovs G, Loewe P, Mohrholz V, Wasmund N (2005) Synchronous ecological regime shifts in the central Baltic and the North Sea in the late 1980 s. ICES J Mar Sci 62:1205–1215
Allen AP, Gillooly JF, Brown JH (2005) Linking the global carbon cycle to individual metabolism. Funct Ecol 19:202–213
Andersen T, Carstensen J, Hernándes-García E, Duarte CM (2009) Ecological thresholds and regime shifts: Approaches to identification. Trends Ecol Evol 24:49–57
Angeler DG, Johnson RK (2012) Patterns of temporal community turnover are spatially synchronous across boreal lakes. Freshwater Biol 57:1782–1793
Arhonditsis GB, Brett MT, DeGasperi CL, Schindler DE (2004) Effects of climatic variability on the thermal properties of Lake Washington. Limnol Oceanogr 49:256–270
Ask J, Karlsson J, Persson L, Ask P, Bystrom P, Jansson M (2009) Terrestrial organic matter and light penetration: effects on bacterial and primary production in lakes. Limnol Oceanogr 54:2034–2040
Austin J, Colman S (2008) A century of temperature variability in Lake Superior. Limnol Oceanogr 53:2724–2730
Balayla DJ, Lauridsen TL, Søndergaard M., Jeppesen E (2010) Larger zooplankton in Danish lakes after cold winters: are fish kills of importance? Hydrobiologia 646:159–172
Beaugrand G, Reid PC, Ibanez F, Lindley JA, Edwards M (2002) Reorganization of North Atlantic marine copepod biodiversity and climate. Science 296:1692–1694
Benndorf J, Kranich J, Mehner T, Wagner AK (2001) Temperature impact on the midsummer decline of Daphnia galeata: An analysis of long-term data from the biomanipulated Bautzen Reservoir (Germany). Freshwater Biol 46:199–211
Benson BJ, Lenters JD, Magnuson JJ, Stubbs M, Kratz TK, Dillon PJ, Hecky RE, Lathrop RC (2000) Regional coherence of climatic and lake thermal variables of four lake districts in the Upper Great Lakes Region of North America. Freshwater Biol 43:517–527
Benson BJ, Magnuson JJ, Jensen OP, Card VM, Hodgkins G, Korhonen J, Livingstone DM, Stewart KM, Weyhenmeyer GA, Granin NG (2012) Extreme events, trends, and variability in Northern Hemisphere lake-ice phenology (1855-2005). Climatic Change 112:299–323
Berger SA, Diehl S, Stibor H, Trommer G, Ruhenstroth M (2010) Water temperature and stratification depth independently shift cardinal events during plankton spring succession. Glob Change Biol 16:1954–1965
BES, 2013. The Impact of Extreme Events on Freshwater Ecosystems. British Ecological Society, www.britishecologicalsociety.org/public-policy/our-position/ecological-issues
Blanck A, Lamouroux N (2007) Large-scale intraspecific variation in life-history traits of 44 European freshwater fish. J Biogeogr 34:862–875
Blenckner T, Järvinen M, Weyhenmeyer GA (2004) Atmospheric circulation and its impact on ice phenology in Scandinavia. Boreal Environ Res 9:371–380
Blenckner T, Adrian R, Livingstone DM, Jennings E, Weyhenmeyer GA, George DG, Jankowski T, Jarvinen M, Aonghusa CN, Nõges T, Straile D, Teubner K (2007) Large-scale climatic signatures in lakes across Europe: a meta-analysis. Glob Change Biol 13:1314–1326
Boehrer B, Schultze M (2008) Stratification of lakes. Rev Geophys 46:RG2005. doi:10.1029/2006RG000210
Burgmer T, Hillebrand H (2011) Temperature mean and variance alter phytoplankton biomass and biodiversity in a long-term microcosm experiment. Oikos 120:922–933
Burgmer T, Hillebrand H, Pfenninger M (2007) Effects of climate-driven temperature changes on the diversity of freshwater macroinvertebrates. Oecologia 151:93–103
Carpenter SR, Benson BJ, Biggs R et al (2007) Understanding regional change: A comparison of two lake districts. Bioscience 57:323–335
Christensen JH, Hewitson B, Busuioc A, Chen A, Gao X, Held I, Jones R, Kolli RK, Kwon W-T, Laprise R, Magaña Rueda V, Mearns L, Menéndez CG, Räisänen J, Rinke A, Sarr A, Whetton P (2007) Regional climate projections. In: Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press
Christoffersen K, Andersen N, Søndergaard M, Liboriussen, L, Jeppesen E (2006) Implications of climate-enforced temperature increases on freshwater pico- and nanoplankton populations studied in artificial ponds during 16 months. Hydrobiologia 560:259–266
Coats R, Perez-Losada J, Schladow G, Richards R, Goldman C (2006) The warming of Lake Tahoe. Climatic Change 76:121–148
Daufresne M, Lengfellner K, Sommer U (2009) Global warming benefits the small in aquatic ecosystems. Proc Nat Acad Sci USA 106:12788–12793
de Wit HA, Wright RF (2008) Projected stream water fluxes of NO3 and total organic carbon from the Storgama headwater catchment, Norway, under climate change and reduced acid deposition. Ambio 37:56–63
de Wit HA, Hindar A, Hole L (2008) Winter climate affects long-term trends in stream water nitrate in acid-sensitive catchments in southern Norway. Hydrol Earth Sys Sci 12:393–403
Dokulil MT, Jagsch A, George GD, Anneville O, Jankowski T, Wahl B, Lenhart B, Blenckner T, Teubner K (2006) Twenty years of spatially coherent deepwater warming in lakes across Europe related to the North Atlantic Oscillation. Limnol Oceanogr 51:2787–2793
Domisch S, Araujo MB, Bonada N, Pauls SU, Jahnig SC, Haase P (2013) Modelling distribution in European stream macroinvertebrates under future climates. Glob Change Biol 19:752–762
Downing JA, Prairie YT, Cole JJ, Duarte CM, Tranvik LJ, Striegl RG, McDowell WH, Kortelainen P, Caraco NF, Melack JM, Middelburg JJ (2006) The global abundance and size distribution of lakes, ponds, and impoundments. Limnol Oceanogr 51:2388–2397
Elliott JA (2012) Is the future blue-green? A review of the current model predictions of how climate change could affect pelagic freshwater cyanobacteria. Water Res 46:1364–1371
Elliott JA, Thackeray SJ, Huntingford C, Jones RG (2005) Combining a Regional Climate Model with a phytoplankton community model to predict future changes in phytoplankton in lakes. Freshwater Biol 50:1404–1411
Elliott JA, Jones ID, Thackeray, SJ (2006) Testing the sensitivity of phytoplankton communities to changes in water temperature and nutrient load, in a temperate lake. Hydrobiologia 559:401–411
Elser JJ, Andersen T, Baron JS, Bergström A-K, Kyle M, Nydick KR, Steger L, Hessen DO (2009) Atmospheric nitrogen deposition distorts lake N:P stoichiometry and alters phytoplankton nutrient limitation. Science 326:835–837
Erlandsson M, Buffam I, Folster J, Laudon H, Temnerud J, Weyhenmeyer GA, Bishop K (2008) Thirty-five years of synchrony in the organic matter concentrations of Swedish rivers explained by variation in flow and sulphate. Glob Change Biol 14:1191–1198
Evans CD, Chapman PJ, Clark JM, Monteith DT, Cresser MS (2006) Alternative explanations for rising dissolved organic carbon export from organic soils. Glob Change Biol 12:2044–2053
Feuchtmayr H, Moran R, Hatton K, Connor L, Heyes T, Moss B, Harvey I, Atkinson D (2009) Global warming and eutrophication: effects on water chemistry and autotrophic communities in experimental hypertrophic shallow lake mesocosms. J Appl Ecol 46:713–723
Finstad AG, Helland IP, Ugedal O, Hesthagen T, Hessen DO (2014) Unimodal response of fish yield to dissolved organic carbon. Ecol Lett 17:36–43
Friberg N, Bergfur J, Rasmussen J, Sandin L (2013) Changing Northern catchments: Is altered hydrology, temperature or both going to shape future stream communities and ecosystem processes? Hydrol Proc 27:734–740
Gardner JL, Peters A, Kearney MR, Joseph L, Heinsohn R (2011) Declining body size: a third universal response to warming? Trends Ecol Evol 26:285–291
George DG (2007) The impact of the North Atlantic Oscillation on the development of ice on Lake Windermere. Climatic Change 81:455–468
George DG, Taylor AH (1995) UK lake plankton and the Gulf Stream. Nature 378:139
George DG, Talling JF, Rigg E (2000) Factors influencing the temporal coherence of five lakes in the English Lake District. Freshwater Biol 43:449–461
George DG, Järvinen M, Arvola L (2004a) The influence of the North Atlantic Oscillation on the winter characteristics of Windermere (UK) and Pääjärvi (Finland). Boreal Environ Res 9:389–399
George DG, Maberly SC, Hewitt DP (2004b) The influence of the North Atlantic Oscillation on the physical, chemical and biological characteristics of four lakes in the English Lake District. Freshwater Biol 49:760–774
George DG, Hewitt DP, Jennings E, Allott N, McGinnity P (2007a) The impact of changes in the weather on the surface temperatures of Windermere (UK) and Lough Feeagh (Ireland). In: Water in Celtic Countries: Quantity, Quality and Climatic Variability. Proceedings of the Fourth Inter-Celtic Colloquium on Hydrology and Management of Water Resources, Guimarães, Portugal, July 2005, IAHS Publications 310
George G, Hurley M, Hewitt D (2007b) The impact of climate change on the physical characteristics of the larger lakes in the English Lake District. Freshwater Biol 52:1647–1666
Gerten D, Adrian R (2000) Climate-driven changes in spring plankton dynamics and the sensitivity of shallow polymictic lakes to the North Atlantic Oscillation. Limnol Oceanogr 45:1058–1066
Gerten D, Adrian R (2001) Differences in the persistency of the North Atlantic Oscillation signal among lakes. Limnol Oceanogr 46:448–455
Gerten D, Adrian R (2002a) Effects of climate warming, North Atlantic Oscillation and El Niño on thermal conditions and plankton dynamics in European and North American lakes. The Scientific World Journal 2: 586–606
Gerten D, Adrian R (2002b) Species specific response of freshwater copepods to recent summer warming. Freshwater Biol 47:2163–2173
Giller PS, Hillebrand H, Berninger UG, Gessner MO, Hawkins S, Inchausti P, Inglis C, Leslie H, Malmqvist B, Monaghan MT, Morin PJ, O’Mullan G (2004) Biodiversity effects on ecosystem functioning: emerging issues and their experimental test in aquatic environments. Oikos 104:423–436
Gyllström M, Hansson LA, Jeppesen E, Garcia-Criado F, Gross E, Irvine K, Kairesalo T, Kornijow R, Miracle MR, Nykanen M, Nõges T, Romo S, Stephen D, Van Donk E, Moss B (2005) The role of climate in shaping zooplankton communities of shallow lakes. Limnol Oceanogr 50:2008–2021
Hampton SE, Izmest’eva LR, Moore MV, Katz SL, Dennis B, Silow EA (2008) Sixty years of environmental change in the world’s largest freshwater lake – Lake Baikal, Siberia. Glob Change Biol 14:1947–1958
Hansson LA, Nicolle A, Granéli W, Hallgren P, Kritzberg E, Persson A, Björk J, Nilsson PA, Brönmark C (2012) Food-chain length alters community responses to global change in aquatic systems. Nature Clim Chan 3:228–233
Hargeby A, Blindow I, Hansson LA (2004) Shifts between clear and turbid states in a shallow lake: multi-causal stress from climate, nutrients and biotic interactions. Arch Hydrobiol 161:433–454
Harrod C, Griffiths D, Rosell R, McCarthy TK (2002) Current status of the pollan (Coregonus autumnalis Pallas 1776) in Ireland. Arch Hydrobiol 57:627–638
Hessen DO (1998) Food webs and carbon cycling in humic lakes. In: Hessen DO, Tranvik LJ (eds) Aquatic Humic Substances. Springer
Hessen DO (1999) Catchment properties and the transport of major elements to estuaries. In: Nedwell DB, Raffaelli DG (eds) Advances in Ecological Research 29: Estuaries. Elsevier
Hessen DO, Andersen T, Larsen S, Skjelkvåle BL, De Wit HA (2009) Nitrogen deposition, catchment productivity, and climate as determinants of lake stoichiometry. Limnol Oceanogr 54:2520–2528
Hessen DO, Daufresne M, Leinaas HP (2013) Temperature-size relations from the cellular-genomic perspective. Biol Rev 88:476–489
Hickling R, Roy DB, Hill JK, Thomas CD (2005) A northward shift of range margins in British Odonata. Glob Change Biol 11:502–506
Hillebrand H (2004) On the generality of the latitudinal diversity gradient. Amer Natur 163:192–211
Hillebrand H, Borer ET, Bracken MES, Cardinale BJ, Cebrian J, Cleland EE, Elser JJ, Gruner DS, Harpole WS, Ngai JT, Sandin S, Seabloom EW, Shurin JB, Smith JE, Smith MD (2009) Herbivore metabolism and stoichiometry each constrain herbivory at different organizational scales across ecosystems. Ecol Lett 12:516–527
Hillebrand H, Soininen J, Snoeijs P (2010) Warming leads to higher species turnover in a coastal ecosystem. Glob Change Biol 16:1181–1193
Hillebrand H, Burgmer T, Biermann E (2012) Running to stand still: temperature effects on species richness, species turnover, and functional community dynamics. Mar Biol 159:2415–2422
Hobbie SE, Miley TA, Weiss MS (2002a) Carbon and nitrogen cycling in soils from acidic and nonacidic tundra with different glacial histories in Northern Alaska. Ecosystems 5:761–774
Hobbie SE, Nadelhoffer KJ, Hogberg P (2002b) A synthesis: The role of nutrients as constraints on carbon balances in boreal and arctic regions. Plant Soil 242:163–170
Hofgaard A (1997) Inter-relationships between treeline position, species diversity, land use and climate change in the central Scandes Mountains of Norway. Glob Ecol Biogeogr 6:419–429
Hondzo M, Stefan HG (1993) Regional water temperature characteristics of lake subjected to climate change. Climatic Change 24:187–211
Hong Y, Andersen T, Dörch P, Tominaga K, Thrane JT, Hessen DO (2015) Greenhouse gas metabolism in Nordic boreal lakes. Biogeochemistry 126:211–225
Hongve D, Riise G, Kristiansen JF (2004) Increased colour and organic acid concentrations in Norwegian forest lakes and drinking water - a result of increased precipitation? Aqua Sci 66:231–238
Huber V, Adrian R, Gerten D (2008) Phytoplankton response to climate warming modified by trophic state. Limnol Oceanogr 53:1–13
Huber V, Adrian R, Gerten D (2010) A matter of timing: heat wave impact on crustacean zooplankton. Freshwater Biol 55:1769–1779
Huber V, Wagner C, Gerten D, Adrian R (2012) To bloom or not to bloom: contrasting responses of cyanobacteria to different heat waves explained by critical thresholds of abiotic drivers. Oecologia 169:245–256
Huisman J, Sharples J, Stroom JM, Visser PM, Kardinaal WEA, Verspagen JMH, Sommeijer B (2004) Changes In turbulent mixing shift competition for light between phytoplankton species. Ecology 85:2960–2970
Hurrell JW (1995) Decadal trends in the North Atlantic Oscillation: regional temperatures and precipitation. Science 269:676–679
Hurrell JW, Kushnir Y, Ottersen G, Visbeck M (2003) The North Atlantic Oscillation: climatic significance and environmental impact. Geophysical Monograph Series, V 134. American Geophysical Union
Hussner A, Van Dam H, Vermaat JE, Hilt S (2014) Comparison of native and neophytic aquatic macrophyte developments in a geothermally warmed river and thermally normal channels. Fund Appl Limnol 185:155–165
IPCC (2013) Climate Change and Water. IPCC Technical Report VI. Intergovernmental Panel on Climate Change (IPCC)
Jacobsen D, Milner AM, Brown LE, Dangles O (2012) Biodiversity under threat in glacier-fed river systems. Nat Clim Chan 2:361–364
Janse JH (1997) A model of nutrient dynamics in shallow lakes in relation to multiple stable states. Hydrobiologia 342–343:1-8
Jansson M, Persson L, De Roos AM, Jones RI, Tranvik LJ (2007) Terrestrial carbon and intraspecific size-variation shape lake ecosystems. Trends Ecol Evol 22:316–322
Jennings E, Jones S, Arvola L, Steahr PA, Gaiser E, Jones ID, Weathers KC, Weyhenmeyer GA, Chiu CY, DeEyto E (2012) Effects of weather-related episodic events in lakes: an analysis based on high-frequency data. Freshwater Biol 57:589–601
Jensen HS, Andersen FØ (1992) Importance of temperature, nitrate, and pH for phosphate release from aerobic sediments of 4 shallow, eutrophic lakes. Limnol Oceanogr 37:577–589
Jensen OP, Benson BJ, Magnuson JJ, Card VM, Futter MN, Soranno PA, Stewart KM (2007) Spatial analysis of ice phenology trends across the Laurentian Great Lakes region during a recent warming period. Limnol Oceanogr 52:2013–2026
Jeppesen E, Søndergaard M, Jensen JP (2003) Climatic warming and regime shifts in lake food webs - some comments. Limnol Oceanogr 48:1346–1349
Jeppesen E, Kronvang B, Meerhoff M, Søndergaard M, Hansen KM, Andersen HE, Lauridsen TL, Beklioglu M, Ozen A, Olesen JE (2009) Climate change effects on runoff, catchment phosphorus loading and lake ecological state, and potential adaptations. J Env Qual 38:1930–1941
Jeppesen E, Moss B, Bennion H, Carvalho L, DeMeester L, Friberg N, Gessner MO, Lauridsen, TL, May L, Meerhoff M, Olafsson JS, Soons MB, Verhoeven JTA (2010a) Interaction of climate change and eutrophication. In: Kernan M, Battarbee R, Moss Blackwell B (eds). Climate Change Impacts on Freshwater Ecosystems. Publishing Ltd
Jeppesen E, Meerhoff M, Holmgren K, González-Bergonzoni I, Teixeira-de Mello F, Declerk SAJ, De Meester L, Søndergaard M, Lauridsen TL, Bjerring R, Conde-Porcuna JM, Mazzeo N, Iglesias C, Reizenstein M, Malmquist HJ, Balayla D, Lazzaro X (2010b) Impacts of climate warming on lake fish community structure and potential effects on ecosystem function. Hydrobiologia, 646:73–90
Jeppesen E, Kronvang B, Olesen JE, Søndergaard M, Hoffmann CC, Andersen HE, Lauridsen TL, Liboriussen L, Meerhoff M, Beklioglu M, Özen A (2011) Climate change effects on nitrogen loading from cultivated catchments in Europe: implications for nitrogen retention, ecological state of lakes and adaptation. Hydrobiologia 663:1–21
Jeppesen E, Mehner T, Winfield IJ, Kangur K, Sarvala J, Gerdeaux D, Rask M, Malmquist HJ, Holmgren K, Volta P, Romo S, Eckmann R, Sandstrom A, Blanco S, Kangur A, Stabo HR, Tarvainen M, Ventela AM, Sondergaard M, Lauridsen TL, Meerhoff M (2012) Impacts of climate warming on the long-term dynamics of key fish species in 24 European lakes. Hydrobiologia 694:1–39
Jöhnk KD, Huisman J, Sharples J, Sommeijer B, Visser PM, Stroom JM (2008) Summer heatwaves promote blooms of harmful cyanobacteria. Glob Change Biol 14:495–512
Jones SE, Solomon CT, Weidel BC (2012) Subsidy or subtraction: how do terrestrial inputs influence consumer production in lakes? Freshw Rev 5:37–49
Juutinen S, Rantakari M, Kortelainen P, Huttunen JT, Larmola T, Silvola J, Martikainen PJ (2009) Methane dynamics in different boreal lake types. Biogeosciences 6:209–223
Kalbitz K, Solinger S, Park JH, Michalzik B, Matzner E (2000) Controls on the dynamics of dissolved organic matter in soils: A review. Soil Sci 165:277–304
Kammer A, Hagedorn F, Shevchenko I, Leifeld J, Guggenberger G, Goryacheva T, Rigling A, Moiseev P (2009) Treeline shifts in the Ural mountains affect soil organic matter dynamics. Glob Change Biol 15:1570–1583
Kangur A, Kangur P, Kangur K, Möls T (2007) The role of temperature in the population dynamics of smelt Osmerus eperlanus eperlanus m. spirinchus Pallas in Lake Peipsi (Estonia/Russia). Hydrobiologia 584:433–441
Kangur K, Kangur P, Ginter K, Orru K, Haldna M, Möls T, Kangur A (2013) Long-term effects of extreme weather events and eutrophication on the fish community of shallow lake Peipsi. J Limnol 72:376–387
Karlsson J, Bystrom P, Ask J, Ask P, Persson L, Jansson M (2009) Light limitation of nutrient-poor lake ecosystems. Nature 460:506–580
Kernan M, Battarbee RW, Moss B (eds) (2010) Climate Change Impacts on Freshwater Ecosystems. Wiley-Blackwell
Kirillin G (2010) Modeling the impact of global warming on water temperature and seasonal mixing regimes in small temperate lakes. Boreal Environ Res 15:279–293
Kosten S, Kamarainen A, Jeppesen E, Van Nes EH, Peeters ETHM, Mazzeo N, Sass L, Hauxwell J, Hansel-Welch N, Lauridsen TL, Sondergaard M, Bachmann RW, Lacerot G, Scheffer M (2009) Climate-related differences in the dominance of submerged macrophytes in shallow lakes. Glob Change Biol 15:2503–2517
Kosten S, Huszar VLM, Bécares E, Costa LS, van Donk E, Hansson L-A, Jeppesen E, Kruk C, Lacerot G, Mazzeo N, De Meester L, Moss B, Lürling M, Nõges T, Romo S, Scheffer M (2012) Warmer climate boosts cyanobacterial dominance in Lakes. Glob Change Biol 18:118–126
Lamb HH (1950) Types and spells of weather around the year in the British Isles. Q J Roy Met Soc 76:393–438
Larsen S, Andersen T, Hessen DO (2011a) Predicting organic carbon in lakes from climate drivers and catchment properties. Glob Biogeochem Cyc 25:doi:10.1029/2010GB003908
Larsen S, Andersen T, Hessen DO (2011b) Climate change predicted to cause severe increase of organic carbon in lakes. Glob Change Biol 17:1186–1192
Larsen S, Andersen T, Hessen DO (2011c) The pCO2 in boreal lakes: Organic carbon as a universal predictor. Glob Biochem Cyc 25:doi:10.1029/2010GB003864
Lehtonen H (1996) Potential effects of global warming on northern European freshwater fish and fisheries. Fish Man Ecol 3:59–71
Lehtonen H, Lappalainen L (1995) The effects of climate on the year-class variations of certain freshwater fish species. In: Beamish RJ (ed). Climate Change and Northern Fish Populations, Vol. 121. Canadian Special Publication of Fisheries and Aquatic Sciences
Leibold MA, Holyoak M, Mouquet N, Amarasekare P, Chase JM, Hoopes MF, Holt RD, Shurin JB, Law R, Tilman D, Loreau M, Gonzalez A (2004) The metacommunity concept: a framework for multi-scale community ecology. Ecol Lett 7:601–613
Liboriussen L, Lauridsen TL, Sondergaard M, Landkildehus F, Sondergaard M, Larsen SE, Jeppesen E (2011) Effects of warming and nutrients on sediment community respiration in shallow lakes: an outdoor mesocosm experiment. Freshwater Biol 56:437–447
Lindholm M, Stordal F, Moe SJ, Hessen DO, Aass P (2012) Climate-driven range retraction on an Arctic freshwater crustacean. Freshwater Biol 57:2591–2601
Livingstone DM (2000) Large-scale climatic forcing detected in historical observations of lake ice break-up. Verhandlungen der Internationalen Vereiningung der theoretischen und angewandten Limnologie 27:2775–2783
Livingstone DM (2003) Impact of secular climate change on the thermal structure of a large temperate central European lake. Climatic Change 57:205–225
Livingstone DM (2008) A change of climate provokes a change of paradigm: taking leave of two tacit assumptions about physical lake forcing. Int Rev Hydrobiol 93:404–414
Livingstone DM, Adrian R (2009) Modeling the duration of intermittent ice cover on a lake for climate-change studies. Limnol Oceanogr 54:1709–1722
Livingstone DM, Kernan M (2009) Regional coherence and geographical variability in the surface water temperatures of Scottish Highland lochs. Fund Appl Limnol 62:367–378
Livingstone DM, Padisák J (2007) Large-scale coherence in the response of lake surface-water temperatures to synoptic-scale climate forcing during summer. Limnol Oceanogr 52:896–902
Livingstone DM, Adrian R, Arvola L, Blenckner T, Dokulil MT, Hari RE, George G, Jankowski T, Järvinen M, Jennings E, Nõges P, Nõges T, Straile D, Weyhenmeyer GA (2010a) Regional and supra-regional coherence in limnological variables. In: George DG (ed) The Impact of Climate Change on European Lakes. Aquatic Ecology Series 4, Springer
Livingstone DM, Adrian R, Blenckner T, George G, Weyhenmeyer GA (2010b) Lake ice phenology. In: George DG (ed) The Impact of Climate Change on European Lakes. Aquatic Ecology Series 4, Springer
Lo TT, Hsu HH (2010) Change in the dominant decadal patterns and the late 1980 s abrupt warming in the extratropical Northern Hemisphere. Atmos Sci Lett 11:210–215
Loreau M, Mouquet N, Gonzalez A (2003) Biodiversity as spatial insurance in heterogeneous landscapes. Proc Nat Acad Sci USA 100:12765–12770
Magnuson JJ, Benson BJ, Kratz TK (1990) Temporal coherence in the limnology of a suite of lakes in Wisconsin, USA. Freshwater Biol 23:145–149
Magnuson JJ, Robertson DM, Benson BJ, Wynne RH, Livingstone DM, Arai T, Assel RA, Barry RG, Card V, Kuusisto E, Granin NG, Prowse TD, Stewart KM, Vuglinski VS (2000) Historical trends in lake and river ice cover in the Northern Hemisphere. Science 289:1743–1746 and Errata 2001. Science 291:254
MacKay MD, Neale PJ, Arp CD, De Senerpont Domis LN, Fang X, Gal G, Jöhnk KD, Kirillin G, Lenters JD, Litchman E, MacIntyre S, Marsh P, Melack J, Mooij WM, Peeters F, Quesada A, Schladow SG, Schmid M, Spence C, Stokes SL (2009) Modeling lakes and reservoirs in the climate system. Limnol Oceanog 54:2315–2329
McKee D, Hatton K, Eaton JW, Atkinson D, Atherton A, Harvey I, Moss B (2002) Effects of simulated climate warming on macrophytes in freshwater microcosm communities. Aquat Bot 74:71–83
McKee D, Atkinson D, Collings SE, Eaton JW, Gill AB, Harvey I, Hatton K, Heyes T, Wilson D, Moss B (2003) Response of freshwater microcosm communities to nutrients, fish, and elevated temperature during winter and summer. Limnol Oceanogr 48:707–722
Meerhoff M, Teixeira-de Mello F, Kruk C, Alonso C, González-Bergonzoni I, Pacheco JP, Lacerot G, Arim M, Beklioğlu M, Brucet S, Goyenola G, Iglesias C, Mazzeo N, Kosten S, Jeppesen E (2012) Environmental warming in shallow lakes: a review of potential changes in community structure as evidenced from space-for-time substitution approaches. Adv Ecol Res 46:259–349
Milbrink G, Vrede T, Tranvik LJ, Rydin E (2011) Large-scale and long-term decrease in fish growth following the construction of hydroelectric reservoirs. Can J Fish Aquat Sci 68:2167–2173
Moe TF, Brysting AK, Andersen T, Schneider SC, Kaste O, Hessen DO (2013) Nuisance growth of Juncus bulbosus: the roles of genetics and environmental drivers tested in a large-scale survey. Freshwater Biol 58:114–127
Monteith DT, Stoddard JL, Evans CD, de Wit HA, Forsius M, Høgåsen T, Wilander A, Skjelkvåle BL, Jeffries DS, Vuorenmaa J, Keller B, Kopácek J, Vesely J (2007) Dissolved organic carbon trends resulting from changes in atmospheric deposition chemistry. Nature 450:537–540
Mooij WM, Hülsman S, De Senerpont Domis LN, Nolet BA, Bodelier PLE, Boers PCM, Dionisio Pires LM, Gons HJ, Ibelings BW, Noordhuis R, Portielje R, Wolfstein K, Lammens EHRR (2005) The impact of climate change on lakes in the Netherlands: a review. Aquat Ecol 39:381–400
Mooij WM, Janse JH, De Senerpont Domis, L, Hülsmann S, Ibelings B (2007) Predicting the effect of climate change on temperate shallow lakes with the ecosystem model PCLake. Hydrobiologia 584:443–454
Mooij WM, De Senerpont Domis LN, Janse JH (2009) Linking species- and ecosystem-level impacts of climate change in lakes with a complex and a minimal model. Ecol Modell 220:3011–3020
Mormul RP, Ahlgren J, Ekvall MK, Hansson L-A, Brönmark C (2012) Water brownification may increase the invisibility of a submerged non-native macrophyte. Biol Invas 14:2091–2099
Moss B (2010) Climate change, nutrient pollution and the bargain of Dr Faustus. Freshwater Biol 55:175–187
Moss B, Hering D, Green AJ, Adoud A, Becares E, M. Beklioglu M, Bennion H, Boix D, Brucet S, Carvalho L, Clement B, Davidson TA, Declerck S, Dobson M, van Donk E, Dudley B, Feuchtmayr H, Friberg N, Grenouillet G, Hillebrand H, Hobaek H, Irvine K, Jeppesen E, Johnson R, Jones I, Kernan M, Lauridsen T, Manca M, Meerhoff M, Olafsson J, Ormerod S, Papastergiadou E, Penning W, Ptacnik R, Quintana X, Sandin L, Seferlis M, Simpson G, Trigal C, Verdonschot P, Verschoor A, Weyhenmeyer G (2009) Climate change and the future of freshwater biodiversity in Europe: a primer for policy-makers. Freshw Rev 2:103–130
Moss B, Kosten S, Meerhoff M, Battarbee RW, Jeppesen E, Mazzeo N, Havens K, Lacerot G, Liu Z, De Meester L, Paerl H, Scheffer M (2011) Allied attack: climate change and eutrophication. Inland Waters 1:101–105
Netten JJC, Arts GHP, Gylstra R, van Nes EH, Scheffer M, Roijackers RMM (2010) Effect of temperature and nutrients on the competition between free-floating Salvinia natans and submerged Elodea nuttallii in mesocosms. Fund Appl Limnol 177:125–132
Netten JJC, Van Zuidam J, Kosten S, Peeters ETHM (2011) Differential response to climatic variation of free-floating and submerged macrophytes in ditches. Freshwater Biol 56:1761–1768
Nielsen A, Trolle D, Me W, Luo L, Han B, Liu Z, Olesen JE, Jeppesen E (2013) Assessing ways to combat eutrophication in a Chinese drinking water reservoir using SWAT. Mar Freshw Res 64:1–18
Nilsson C, Jansson R, Kuglerova L, Lind L, Strom L (2013) Boreal riparian vegetation under climate change. Ecosystems 16:401–410
Norton L, Elliott JA, Maberly SC, May L (2012) Using models to bridge the gap between land use and algal blooms: An example from the Loweswater catchment, UK. Env Modell Softw 36:64–75
Ojala A, Lopez Bellido J, Tulonen T, Kankaala P, Huotari J (2011) Carbon gas fluxes from a brown-water and a clear-water lake in the boreal zone during a summer with extreme rain events. Limnol Oceanogr 56:61–76
O’Reilly CN, Sharma S, Gray DK et al. (2015) Rapid and highly variable warming of lake surface waters around the globe. Geophys Res Lett 42, doi: 10.1002/2015GL066235
O’Reilly CM, Alin SR, Plisnier P-D, Cohen AS, McKee BA (2003) Climate change decreases aquatic ecosystem productivity of Lake Tanganyika, Africa. Nature 424:766–768
Özen A, Šorf M, Trochine C, Liboriussen L, Beklioglu M, Søndergaard M, Lauridsen TL, Johansson LS, Jeppesen E (2013) Long-term effects of warming and nutrients on microbes and other plankton in mesocosms. Freshwater Biol 58:483–493
Padisak J (1997) Cylindrospermopsis raciborskii (Woloszynska) seenayya et Subba Raju, an expanding, highly adaptive cyanobacterium: worldwide distribution and review of its ecology. Arch. Hydrobiol Suppl 107:563–593
Paerl HW (1988) Nuisance phytoplankton blooms in coastal, estuarine and inland waters. Limnol Oceanogr 33:823–847
Parmesan C, Yohe G (2003) A globally coherent fingerprint of climate change impacts across natural systems. Nature 421:37–42
Peeters F, Livingstone DM, Goudsmit G-H, Kipfer R, Forster R (2002) Modeling 50 years of historical temperature profiles in a large central European lake. Limnol Oceanogr 47:186–197
Peeters F, Straile D, Lorke A, Livingstone DM (2007) Earlier onset of the spring phytoplankton bloom in lakes of the temperate zone in a warmer climate. Glob Change Biol 13:1898–1909
Posch T, Koster O, Salcher MM, Pernthaler J (2012) Harmful filamentous cyanobacteria favoured by reduced water turnover with lake warming. Nature Clim Change 2:809–813
Ptacnik R, Solimini AG, Andersen T, Tamminen T, Brettum P, Lepisto L, Willen E, Rekolainen S (2008) Diversity predicts stability and resource use efficiency in natural phytoplankton communities. Proc Nat Acad Sci USA 105:5134–5138
Quayle WC, Peck LS, Peat H, Ellis-Evans JC, Harrigan PR (2002) Extreme responses to climate change in Antarctic lakes. Science 295:645
Reid PC, de Fatima Borges M, Svendsen E (2001) A regime shift in the North Sea circa 1988 linked to changes in the North Sea horse mackerel fishery. Fish Res 50:163–171
Reynolds C (1984) The Ecology of Freshwater Phytoplankton. Cambridge University Press
Robertson DM, Ragotzkie RA (1990) Changes in the thermal structure of moderate to large sized lakes in response to changes in air temperature. Aquat Sci 52:360–380
Rodionov SN, Overland JE (2005) Application of a sequential regime shift detection method to the Bering Sea ecosystem. ICES J Mar Sci 62:328–333
Rolinski S, Horn H, Petzoldt T, Paul L (2007) Identifying cardinal dates in phytoplankton time series to enable the analysis of long-term trends. Oecologia 153:997–1008
Rosenzweig C (2007) Assessment of observed changes and responses in natural and managed systems. In: Parry ML, Canziani OF, Palutikof JP, van der Linden PJ, Hanson CE (eds) Climate change 2007. Impacts, adaptation and vulnerability. Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press
Ruuhijärvi J, Rask M, Vesala SA, Westermark A, Olin M, Keskitalo J, Lehtovaara A (2010) Recovery of the fish community and changes in the lower trophic levels in a eutrophic lake after a winter kill of fish. Hydrobiologia 646:145–158
Salmaso N (2005) Effects of climatic fluctuations and vertical mixing on the interannual trophic variability of Lake Garda, Italy. Limno Oceanogr 50:553–565
Saloranta TM, Andersen T (2007) MyLake—A multi-year lake simulation model code suitable for uncertainty and sensitivity analysis simulations. Ecol Model 207:45–60
Schabhuettl S, Hingsamer P, Weigelhofer G, Hein T, Weigert A, Striebel M (2013) Temperature and species richness effects in phytoplankton communities. Oecologia 171:527–536
Schär C, Vidale P, Luthi D, Frei C, Haberli C, Liniger M, Appenzeller C (2004) The role of increasing temperature variability in European summer heatwaves. Nature 427:332–336
Scharfenberger U, Mahdy A, Adrian R (2013) Threshold-driven shifts in two copepod species: testing ecological theory with observational data. Limnol Oceanogr 58:741–752
Scheffer M, Carpenter SR (2003) Catastrophic regime shifts in ecosystems: linking theory to observation. Trends Ecol Evol 18:648–656
Scheffer M, Straile D, van Nes EH, Hosper H (2001) Climatic warming causes regime shifts in lake food webs. Limnol Oceanogr 46:1780–1783
Scheffer M, van Geest GJ, Zimmer K, Jeppesen E, Sondergaard M, Butler MG, Hanson MA, Declerck S, De Meester L (2006) Small habitat size and isolation can promote species richness: second-order effects on biodiversity in shallow lakes and ponds. Oikos 112:227–231
Schindler DW (2001) The cumulative effects of climate warming and other human stresses on Canadian freshwaters in the new millenium. Can J Fish Aquat Sci 58:18–29
Schindler DW, Beaty KG, Fee EJ, Cruikshank DR, DeBruyn ER, Findlay DL, Linsey GA, Shearer JA, Stainton MP, Turner MA (1990) Effects of climatic warming on lakes of the central boreal forest. Science 250:967–970
Schindler DE, Rogers DE, Scheuerell MD, Abrey CA (2005) Effects of changing climate on zooplankton and juvenile sockeye salmon growth in southwestern Alaska. Ecology 86:198–209
Schneider P, Hook SJ (2010) Space observations of inland water bodies show rapid surface warming since 1985. Geophys Res Lett 37:L22405, doi:10.1029/2010GL045059
Schneider C, Laize CLR, Acreman MC, Florke M (2013) How will climate change modify river flow regimes in Europe? Hydrol Earth Sys Sci 17:325–339
Seebens H, Straile D, Hoegg R, Stich HB, Einsle U (2007) Population dynamics of a freshwater calanoid copepod: Complex responses to changes in trophic status and climate variability. Limnol Oceanogr 52:2364–2372
Seebens H, Einsle U, Straile D (2009) Copepod life cycle adaptations and success in response to phytoplankton spring bloom phenology. Glob Change Biol 15:1394–1404
Shatwell T, Köhler J, Nicklisch A (2008) Warming promotes cold-adapted phytoplankton in temperate lakes and opens a loophole for Oscillatoriales in spring. Glob Change Biol 14:1–7
Shurin JB, Winder M, Adrian R, Keller W, Matthews B, Paterson AM, Paterson MJ, Pinel-Alloul B, Rusak JA, Yan ND (2010) Environmental stability and lake zooplankton diversity - contrasting effects of chemical and thermal variability. Ecol Lett 13:453–463
Sobek S, Algesten G, Bergstrom AK, Jansson M, Tranvik LJ (2003) The catchment and climate regulation of pCO2 in boreal lakes. Glob Change Biol 9:630–641
Solomon CT, Bruesewitz DA, Richardson DC, et al. (2013) Ecosystem respiration: Drivers of daily variability and background respiration in lakes around the globe. Limnol Oceanogr 58:849–866
Søndergaard M, Jensen JP, Jeppesen E (2003) Role of sediment and internal loading of phosphorus in shallow lakes. Hydrobiologia 506:135–145
Straile D (2002) North Atlantic Oscillation synchronizes food web interactions in central European lakes. Proc Roy Soc Lond 269:391–395
Straile D, Adrian R (2000) The North Atlantic Oscillation and plankton dynamics in two European lakes - two variations on a general theme. Glob Change Biol 6:663–670
Straile D, Livingstone DM, Weyhenmeyer GA, George DG (2003) The response of freshwater ecosystems to climate variability associated with the North Atlantic Oscillation. In: Hurrell JW, Kushnir Y, Ottersen G, Visbeck M (eds) The North Atlantic Oscillation: Climatic significance and environmental impact. Geophysical Monograph Series 134. American Geophysical Union
Straile D, Adrian R, Schindler DE (2012) Uniform temperature dependency of the phenology of keystone herbivore in lakes of the Northern Hemisphere. PloSONE 7:345497. doi:10.1371/journal.pone.0045497
Temnerud J, Weyhenmeyer GA (2008) Abrubt changes in air temperature and precipitation: Do they matter for water chemistry? Glob Biochem Cyc 22:GB2008, doi:10.1029/2007GB003023
Thompson PL, Shurin JB (2012) Regional zooplankton biodiversity provides limited buffering of pond ecosystems against climate change. J Anim Ecol 81:251–259
Thompson DWJ, Wallace JM (1998) The Arctic Oscillation signature in the wintertime geopotential height and temperature fields. Geophys Res Lett 25:1297–1300
Thompson RM, Beardall J, Beringer J, Grace M, Sardina P (2013) Means and extremes: building variability into community-level climate change experiments. Ecol Lett 16:799–806
Thrane J-E, Hessen DO, Andersen T (2014) The absorption of light in lakes: negative impacts of dissolved organic carbon on primary productivity. Ecosystems 17:1040–1052
Tisseuil C, Cornu J-F, Beauchard O, Brosse S, Darwall W, Holland R, Hugueny B, Tedesco PA, Oberdorff T (2013) Global diversity patterns and cross-taxa convergence in freshwater systems. J Anim Ecol 82:365–376
Tockner K, Pusch M, Borchardt D, Lorang MS (2010) Multiple stressors in coupled river-floodplain ecosystems. Freshwater Biol 55:135–151
Tranvik LJ, Downing JA, Cotner JB, Loiselle SA, Striegl RG, Ballatore TJ, Dillon P, Finlay K, Fortino K, Knoll LB, Kortelainen PL, Kutser T, Larsen S, Laurion I, Leech DM, McCallister SL, McKnight DM, Melack JM, Overholt E, Porter JA, Prairie Y, Renwick WH, Roland F, Sherman BS, Schindler DW, Sobek S, Tremblay A, Vanni M J, Verschoor AM, von Wachenfeldt E, Weyhenmeyer GA (2009) Lakes and reservoirs as regulators of carbon cycling and climate. Limnol Oceanogr 54:2298–2314
Trochine C, Guerrero M, Liboriussen L, Meerhoff M, Lauridsen TL, Søndergaard M, Jeppesen E (2011) Filamentous green algae inhibit phytoplankton with enhanced effects when lakes get warmer. Freshwater Biol 56:541–553
Trolle D, Hamilton DP, Pilditch CA, Duggan IC, Jeppesen E (2011) Predicting the effects of climate change on trophic status of three morphologically varying lakes: Implications for lake restoration and management. Env Modell Softw 26:354–370
Trolle D, Hamilton DP, Hipsey MR, Bolding K, Bruggeman J, Mooij WM, Janse JH, Nielsen A, Jeppesen E, Elliott A, Makler-Pick V, Petzoldt T, Rinke K, Flindt MR, Arhonditsis GB, Gal G, Bjerring R, Tominaga K, Hoen J, Downing AS, Marques DM, Fragoso CR, Søndergaard M, Hanson PC (2012) A community-based framework for aquatic ecosystem models. Hydrobiologia 683:25–34
van Donk E, Santamaria L, Mooij W (2003) Climatic warming causes regime shifts in lake food webs: a re-assessment. Limnol Oceanogr 48:1350–1353
Verburg P, Hecky RE, Kling H (2003) Ecological consequences of a century of warming in Lake Tanganyika. Science 301:505–507
Wagner C, Adrian R (2009a) Exploring lake ecosystems: hierarchy responses to long-term change? Glob Change Biol 5:1104–1115
Wagner C, Adrian R (2009b) Cyanobacteria dominance: Quantifying the effects of climate change. Limnol Oceanogr 54:2460–2468
Wagner C, Adrian R (2011) Consequences of changes in thermal regime for plankton diversity and trait composition in a polymictic lake: a matter of temporal scale. Freshwater Biol 56:1949–1961
Wagner A, Hülsmann S, Paul L, Paul RJ, Petzoldt T, Sachse R, Schiller T, Zeis B, Benndorf J, Berendonk TU (2012) A phenomenological approach shows a high coherence of warming patterns in dimictic aquatic systems across latitude. Mar Biol 159:2543–2559
Watkins EM, Schindler DW, Turner MA, Findlay D (2001) Effects of solar ultraviolet radiation on epilithic metabolism, and nutrient and community composition in a clear-water boreal lake. Can J Fish Aquat Sci 58:2059–2070
Weyhenmeyer GA (2004) Synchrony in relationships between the North Atlantic Oscillation and water chemistry among Sweden’s largest lakes. Limnol Oceanogr 49:1191–1201
Weyhenmeyer GA, Karlsson J (2009) Nonlinear response of dissolved organic carbon concentrations in boreal lakes to increasing temperatures. Limnol Oceanogr 54:2513–2519
Weyhenmeyer GA, Blenckner T, Pettersson K (1999) Changes of the plankton spring outburst related to the North Atlantic Oscillation. Limnol Oceanogr 44:1788–1792
Weyhenmeyer GA, Meili M, Livingstone DM (2004a) Nonlinear temperature response of lake ice breakup. Geophys Res Lett 31:L07203, doi:10.1029/2004GL019530
Weyhenmeyer GA, Willén E, Sonesten L (2004b) Effects of an extreme precipitation event on water chemistry and phytoplankton in the Swedish Lake Mälaren. Bor Env Res 9:409–420
Weyhenmeyer GA, Meili M, Livingstone DM (2005) Systematic differences in the trend towards earlier ice-out on Swedish lakes along a latitudinal temperature gradient. Verhandlungen der Internationalen Vereiningung der theoretischen und angewandten Limnologie 29:257–260
Weyhenmeyer GA, Jeppesen E, Adrian R, Blenckner T, Jankowski T, Jennings E, Jensen JP, Nõges P, Nõges T, Straile D (2007) Nitrate-depleted conditions on the increase in shallow northern European lakes. Limnol Oceanogr 52:1346–1353
Weyhenmeyer GA, Livingstone DM, Meili M, Jensen OP, Benson B, Magnuson JJ (2011) Large geographical differences in the sensitivity of ice-covered lakes and rivers in the Northern Hemisphere to temperature changes. Glob Change Biol 17:268–275
Wiedner C, Rücker J, Brüggemann R, Nixdorf B (2007) Climate change affects timing and size of populations of an invasive cyanobacterium in temperate regions. Oecologia 152:473–484
Wilhelm S, Adrian R (2007) Long-term response of Dreissena polymorpha larvae to physical and biological forcing in a shallow lake. Oecologia 151:104–114
Wilhelm S, Adrian R (2008) Impact of summer warming on the thermal characteristics of a polymictic lake and consequences for oxygen, nutrients and phytoplankton. Freshwater Biol 53:226–237
Wilhelm S, Hintze T, Livingstone DM, Adrian R (2006) Long-term response of daily epilimnetic temperature extrema to climate forcing. Can J Fish Aquat Sci 63:2467–2477
Williamson CE, Saros JE, Schindler DW (2009) Sentinels of change. Science 323:887–889
Winder M, Schindler DE, Essington TE, Litt AH (2009) Disrupted seasonal clockwork in the population dynamics of a freshwater copepod by climate warming. Limnol Oceanogr 54:2493–2505
Winfield IJ, Hateley J, Fletcher JM, James JB, Bean CW, Clabburn P (2010) Population trends of Arctic charr (Salvelinus alpinus) in the UK: assessing the evidence for a widespread decline in response to climate change. Hydrobiologia 650:55–65
Woodward G, Perkins DM, Brown LE (2010) Climate change and freshwater ecosystems: impacts across multiple levels of organization. Phil Trans Roy Soc 365:2093–2106
Wright RF, Bishop K, Dillon PJ, Erlandsson M, Evans CD, Hardekopf DW, Helliwell R, Hruška J, Hutchins M, Kaste Ø, Kopáček J, Krám P, Laudon H, Moldan F, Rogora M, Sjøeng AMS, de Wit HA (2010) Interaction of climate change and acid deposition. In: Kernan M, Moss B, Battarbee RW (eds) Climate Change Impacts on Freshwater Ecosystems. Wiley-Blackwell
Xenopoulos MA, Leavitt PR, Schindler DW (2009) Ecosystem-level regulation of boreal lake phytoplankton by ultraviolet radiation. Can J Fish Aquat Sci 66:2002–2010
Yoo J, D’Odorico P (2002) Trends and fluctuations in the dates of ice break-up of lakes and rivers in Northern Europe: the effect of the North Atlantic Oscillation. J Hydrol 268:100–112
Yvon-Durocher G, Jones JI, Trimmer M, Woodward G, Montoya JM (2010). Warming alters the metabolic balance of ecosystems. Phil Trans Roy Soc 365:2117–2126
Yvon-Durocher G, Montoya JM, Trimmer M, Woodward G (2011) Warming alters the size spectrum and shifts the distribution of biomass in freshwater ecosystems. Glob Change Biol 17:1681–1694
Author information
Authors and Affiliations
Corresponding authors
Editor information
Editors and Affiliations
Rights and permissions
Open Access This chapter is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits use, duplication, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license and indicate if changes were made.
The images or other third party material in this chapter are included in the work’s Creative Commons license, unless indicated otherwise in the credit line; if such material is not included in the work’s Creative Commons license and the respective action is not permitted by statutory regulation, users will need to obtain permission from the license holder to duplicate, adapt or reproduce the material.
Copyright information
© 2016 The Author(s)
About this chapter
Cite this chapter
Adrian, R. et al. (2016). Environmental Impacts—Lake Ecosystems. In: Quante, M., Colijn, F. (eds) North Sea Region Climate Change Assessment. Regional Climate Studies. Springer, Cham. https://doi.org/10.1007/978-3-319-39745-0_10
Download citation
DOI: https://doi.org/10.1007/978-3-319-39745-0_10
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-39743-6
Online ISBN: 978-3-319-39745-0
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)