
Abstract
Natural disturbances, or the lack thereof, contributed to shape Earth’s landscapes and maintain its diversity of ecosystems. In particular, natural fire dynamics and herbivory by wild megafauna played an essential role in defining European landscapes in pre-agricultural times. The advent of agriculture and the development of complex societies exacerbated the decline of European megafauna, leading to local and global extinctions of many species, and substantial alterations of fire regimes. Those natural phenomena were over time gradually and steadily replaced by anthropogenic disturbances. Yet, for the first time since the Black Death epidemic, agricultural land-use is decreasing in Europe. Less productive marginal areas have been progressively abandoned as crop and livestock production has become concentrated on the most fertile and easier to cultivate land. With little or no substitute for the anthropogenic disturbances associated with these abandoned agricultural practices, there is growing concern that disturbance-dependent communities may disappear, along with their associated ecosystem services. Nonetheless, rewilding can give an opportunity to tackle the issue of farmland abandonment. This chapter first depicts the historical European landscapes and the role of two natural disturbances, herbivory and fire. The importance of disturbance-dependent habitats is then highlighted by drawing attention to the alpha and beta diversity that they sustain. Finally, the chapter investigates options for rewilding abandoned land to maintain disturbance-dependent and self-sustained habitats for which we suggest active restoration in the early stages of abandonment. This may be achieved via prescribed burning and support or introduction, when necessary, of populations of wild mammals.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others

Keywords
1 Introduction
Disturbance can be defined as “a discrete event that disrupts the structure of an ecosystem’s community or population, and changes resources availability or the physical environment” (Turner 1998). Natural disturbances (i.e., not deriving from human-induced processes) are an essential process of ecosystem dynamics. Among other roles, disturbances contribute to the maintenance of ecosystem structure and nutrient cycling (Attiwill 1994; Turner 1998). More important than considering the impact of a disturbance event per se is to consider the regime underlying disturbances. The disturbance regime determines the landscape (Turner 1998), and is characterized by the disturbance frequency and return interval, spatial extent, intensity (energy flow per area per time) and severity (magnitude of impact).
For millennia, humans have modified ecosystems with varying intensity and over various spatial extents. These anthropogenic changes imply a modification in both the natural communities and the natural processes that cause disturbance. In particular, human activities often cause the disruption of natural regimes, either directly (e.g., livestock grazing, fire suppression) or indirectly (e.g., landscape fragmentation, introduction of exotic invasives or pests), or introduce new types of disturbance, such as pollution. Human activities can also mimic natural disturbance regimes and affect biotic communities in a similar way (Attiwill 1994). For example, the maintenance of traditional landscapes and the species-rich communities associated with them is implicitly linked with continuous ecosystem disturbance imposed by human activities.
If the regime of anthropogenic disturbances is altered, by a reduction or complete withdrawal of human activities, there is a concern that disturbance-dependent habitats and the associated communities may not be maintained. In particular, the maintenance of extensive farming systems in Europe is currently at stake due to farmland abandonment, which raises concerns about the potential effects of land-use changes on biodiversity (Rey Benayas et al. 2007). The trajectory of ecological succession after abandonment depends on several factors, but the probable shift from a moderate disturbance regime (i.e. traditional landscape mosaic) to a low or high disturbance regime is associated with the risk of habitat homogenization and decline of species richness. Thus, one of the challenges of rewilding abandoned farmland is to contribute to the maintenance of disturbance-dependent habitats.
Passive regeneration following farmland abandonment can be a long and complex process, specific to each area (see Chap. 1). It depends on the cultivation history, the time since abandonment, the availability of a “natural” seed bank, the proximity of sources of populations of species, and the requirements for natural disturbances, which will all take part in the self-sustained functioning of the restored ecosystem. When active restoration is needed, the choice of the baseline is also important (Corlett 2012), and in this regard, open land maintained quasi exclusively by (traditional) agricultural practices is a rather recent norm.
In this chapter, we first depict the European landscapes through time, from pre-modern human settlement to the progressive advent of agriculture and finally to the recent trends of agricultural abandonment. We then present two major disturbances: i.e. herbivory and fire, from both a natural history perspective and a restoration approach. We also look into the consequences of those disturbances on alpha and beta diversity levels in the landscape.
2 A Picture of Historical European Landscapes
2.1 An Ongoing Debate…
Describing the species, habitats, and interactions that would be present without the influence of modern humans, i.e. the pre-historical baseline, is an important step to understand natural dynamics and disturbances, and guide the restoration of self-sustaining systems (Svenning 2002; Gillson and Willis 2004; Willis and Birks 2006). However, the composition of the “pre-Neolithic landscape” (Hodder et al. 2009) is still the subject of active debate.
The Middle and Late Pleistocene interglacial can be used as proxies to describe the European pre-Neolithic landscapes, due to their similar climatic conditions and low human activity (Svenning 2002). Two contrasting pictures of lowland temperate European landscapes for these periods are described: (i) the “high-forest” hypothesis, where most of Europe was covered by forest and where the forest dynamics and the resulting availability of open-land influenced herbivore populations; (ii) the “wood-pasture” hypothesis, depicting the European landscapes as a mosaic of forest and open-land where herbivory was the main driver of openness (Vera 2000; Bradshaw et al. 2003; Birks 2005; Mitchell 2005).
Pollen records have been used to test both hypotheses and assess the degree of openness, or lack thereof, of European landscapes. Typically, the ratio between the percentage of tree pollen and non-arboreal pollen gives an indication of the openness of a landscape (Svenning 2002). Pollen records show that shade-intolerant species were present in areas both with and without evidence of large herbivores, which is in favor of the “high forest” hypothesis, in which grazers are not essential to maintaining those species (Mitchell 2005). Nonetheless, pollen and dung beetle fossil record support the idea that megaherbivores were the main keepers of openness, at least of the floodplains in Northwestern Europe (Svenning 2002), as a diverse community of dung-dependent beetles can be linked with the occurrences of large populations of herbivores (Sandom et al. 2014).
Yet, three other types of natural processes can also explain the occurrence of open areas: forest fires, windthrows and edaphic-topographic conditions (Svenning 2002; Fyfe 2007; Molinari et al. 2013). The most likely explanation is that the distribution of habitats was originally based on physical factors (Bradshaw et al. 2003), and was then enhanced and/or maintained by large herbivores, whose browsing and grazing impact delayed secondary successions.
2.2 Temporal Evolution of the European Landscape
The first hominids reached Europe from Africa in the Early Pleistocene, some 1.2–1.1 million years ago (Carbonell et al. 2008), while modern humans colonized the continent between 46,000 BP and 41,000 BP (Mellars 2006). The appropriation of new land coincided with changes in the European landscape . Nomadic hunter-gatherers started to actively manage their ecosystem with the use of fire during the Pleistocene: what started as a domestic tool (e.g. for cooking, heating, and for protection from predators) also became useful to draw game to hunting grounds, to clear travel routes, and to open space for grazers (Daniau et al. 2010; Kaplan et al. 2010; Pfeiffer et al. 2013).
The development of agriculture was the next step in humans’ appropriation and management of their environment (Pereira et al. 2012). The spread of agriculture from the northern Levant and northern Mesopotamian area towards Europe has been calculated to have started between 11,550 and 9000 BP and expanded at a rate of 0.6 to 1.3 km/year, with agriculture reaching north-western Europe in 3000 years (Pinhasi et al. 2005; Ruddiman 2013). Such spread of agriculture led to a fivefold increase in the human population (Gignoux et al. 2011), which had considerable consequences on the landscape. Several models have been designed to investigate the historical evolution of this human impact. Models that do not assume a direct linear link between human density and deforestation, but also consider other factors, such as technological change, show that the rate of land appropriation was much higher in the distant than in the recent past (Ruddiman 2013; Kaplan et al. 2010). First of all, as time passed and deforestation occurred, less and less forest was left to clear. Most of all, technological improvements allowed people to produce the same amount of food on less land, which contradicts a direct link between population density and deforestation (Ruddiman 2013). Following these non-linear concepts, Kaplan et al. (2010) presented model scenarios of Holocene anthropogenic land cover change. At 8000 BP, only Mesopotamia and Turkey were showing signs of human use of the land, but by the beginning of the Iron Age at 3000 BP, up to 40 % of European land could have been cleared for extensive agriculture and pastures (Fig. 8.1). Between 8000 and 3000 BP, Kaplan et al (2010) suggest that land use in Western Europe ranged from 5.5 to 6.5 ha per capita and was relatively stable. By 2500 BP, increasing populations in most of Western Europe triggered intensification of land use (Fig. 8.1) and decrease in per capita values. Later, decreases in population resulted in major land abandonment episodes, during the Migration Period following the fall of the Western Roman Empire, and after the Black Death epidemic of AD 1350. By AD 1850, the latest preindustrial time, most of the European landscapes usable for intensive crop or pasture were deforested, and land use had dropped to values close to 0.5 ha per capita.

Anthropogenic land cover change in Europe over the Holocene. Intensive land use includes land completely deforested and used for agriculture and pasture, while extensive land use includes forest-pasture, coppice and other managed forestlands. Open rangeland occurs on land that is either too cold or too dry for non-irrigated agricultural land uses. Land use is driven by estimates of past population at the country-level. (Based on Kaplan et al. 2009, 2010; Kaplan 2012)
After the Industrial Revolution, the relationship between population and land use had become largely uncoupled. Beginning in the late eighteenth century, these “forest transitions” (e.g. Mather et al. 1998) led to abandonment of unproductive agricultural and pasturel and in most European countries. Since the early 1960s, the rural population decreased by 17 % in Europe (FAOSTAT 2010), with repercussions for agricultural land-use, and both are projected to continue decreasing in the decades to come. By 2030, up to 15 % of the land cultivated in 2000 could be abandoned (e.g. Verburg and Overmars 2009; Eickhout et al. 2007) which represents 10–29 million ha of land. The areas facing the greatest likelihood of rural abandonment are remote and/or mountain areas, classified as “least favored”, with marginal value for agriculture (e.g. MacDonald et al. 2000). With the withdrawal of human activities, those abandoned areas are often left without the artificial disturbances that had replaced the natural ones, centuries or millennia ago.
3 The Role of Natural Disturbances
Investigating the history of natural disturbances can inform researchers and managers on guidelines for restoration (Donlan et al. 2006). We identified two types of disturbances as fundamental in the maintenance of European landscapes, prior to human appropriation of the land: large herbivores and natural fire dynamics.
3.1 The Pre-Neolithic Ecosystem Engineers
Ecosystem engineers are organisms that create and/or maintain habitats, either directly or indirectly (Jones et al. 1994; Wright and Jones 2006) and thus create niches for other species. The fact of grazing and browsing is not enough to be qualified as engineering (Wright and Jones 2006). Nonetheless, the consequences of herbivory , trampling, and fertilizing, especially by large herds of megafauna, have a direct impact on the distribution of habitats (Vera 2000; Birks 2005). Small mammals are also known to have an important impacts on the vegetation, for example by disturbing the soil and modifying its physical and chemical properties (Jones et al. 1994), but this goes beyond the scope of this chapter.
During the interglacial cycles of the late Quaternary, and prior to massive extinctions , the landscapes of Europe were characterized by a rich megafauna (Bradshaw et al. 2003). The available fossil evidence can attest the presence of species in a given region, while using the impact of similar extant species as a proxy can inform on the role of extinct megaherbivores on the landscape (Corlett 2012). Nonetheless, in contrast with pollen, there are too little fossil records of pre-Neolithic large herbivores to allow for an estimate of their past densities (Bradshaw et al. 2003; Mitchell 2005), and we still lack precise knowledge regarding their behavior (Hodder et al. 2009).
The late Pleistocene megafauna of Europe (Table 8.1) resembles the one currently found in savannas, with herbivores such as proboscidae and rhinocerotidae, and large carnivores such as hyaenidae and felidae (Blondel and Aronson 1999; Vera 2000; Bradshaw et al. 2003). Globally, the group of large herbivores suffered more prehistoric extinctions than other taxa (Johnson 2009). Cyclic climatic change had typically been responsible for a regular faunal turnover, and was later combined with increased human pressure (Corlett 2012; Morrison et al. 2007), leading to several of these megafauna becoming regionally (e.g. hippopotamus), globally (e.g. woolly mammoth), and often functionally, extinct (Blondel and Aronson 1999; Bradshaw et al. 2003). Some species also suffered large range contractions, such as the elk (Morrison et al. 2007). Additionally, humans domesticated animals in the Fertile Crescent, about 10,000 years ago (Zeder 2008; Pereira et al. 2012), and as herders migrated west, increasing the area of pasture in Europe, wild herbivores were replaced by domesticated species. Since AD 1, most of the open rangeland in Europe has been under human land-use (Fig. 8.1) .
Extinct and extant large herbivores can be classified according to their feeding behavior (Vera 2000; Svenning 2002; Bullock 2009): browsers (e.g. elk, straight-tusked elephants) are typically associated with tree rich areas; grazers (e.g. hippopotamus, aurochs) are in contrast associated with the occurrence of grass-rich habitat; finally, mixed feeders (e.g. red deer, wild goats) alternate between browsing and grazing (Table 8.1). The European bison, a mixed feeder, has for example been associated historically with both closed forest and semi-open habitats (Kuemmerle et al. 2012). The social structure of the herbivores (i.e. solitary, groups or herds) also provides information on the grazing and browsing pressure on the landscape (Table 8.1).
As a result, one of the most direct impacts of large herbivores on the landscape is the limitation and variation in the spatial distribution of secondary successions (Laskurain et al. 2013; Kuiters and Slim 2003). Yet, the role of herbivores goes beyond the direct impacts of browsing and grazing. For example, elephants are known to create large physical disturbances via the trampling of trees and shrubs (Jones et al. 1994), which changes their habitat, the fuel load and the local fire regime , and in return benefits light demanding plant species. The disturbance induced by the rooting behavior of wild boars favors natural forest regeneration, while being considered as damaging to grasslands (Schley et al. 2008; Sandom et al. 2013a). Large herbivores also have a role as seed-dispersers via their consumption of large quantities of forage: the low oral process of the fruits contained in this forage allows the dispersal of undamaged seeds in feces (Corlett 2012; Johnson 2009). Some seeds even need to pass through a digestive track to trigger germination. Finally, herbivore dung is important for nutrient cycling and soil fertilization (Zimov 2005) .
3.2 Fire Dynamics
Fire is a critical component in the functioning of many ecosystems. It maintains and shapes vegetation structure and biotic communities, promotes natural regeneration and habitat diversity, takes part in biogeochemical cycles, and can influence soil properties and water functions (Thonicke et al. 2001; Bond and Keeley 2005). Unlike grazing, fires consume both dead and living material and do not discriminate between edible and non-edible plants (Bond and Keeley 2005), but may act as a selective pressure over fire resistant traits (Pausas and Bradstock 2007; Pausas et al. 2006).
Fire-dependent systems cover about 53 % of the world’s terrestrial surface (Shlisky et al. 2007). These systems evolved in the presence of fire and depend on this disturbance to maintain their structure and composition (e.g., Mediterranean forests and boreal forests), with fire regimes characterized by their frequency, intensity, seasonality, and specific to each ecosystem. In addition, 22 % of the world’s terrestrial area is covered by fire-sensitive ecosystems, where fire plays a minor role in maintaining ecosystem structure and composition (e.g., broadleaved and mixed forests in the Alps), 15 % is covered by fire-independent ecosystems, where fire is not an evolutionary force due to the scarcity of fuel or ignition sources (e.g. tundra), and the remaining 10 % are not yet classified (Shlisky et al. 2007).
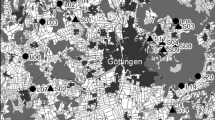
In Europe, natural fire regimes are mainly of two types: (i) intense and large, and (ii) cool and small (Archibald et al. 2013). The former type is typical of Mediterranean and boreal ecosystems, where large crown fires of high intensity return at intervals that can span from a decade, in particular in Mediterranean regions, to more than a century (Archibald et al. 2013). The latter type occurs interspersed with the first type, in the same biomes, and is associated with surface fires burning litter fuels (Archibald et al. 2013). However, due to a long history of human presence, many ecosystems in Europe, including fire-sensitive systems, present altered fire-regimes resulting from land-use changes and anthropogenic fire management (Shlisky et al. 2007; Archibald et al. 2013; Molinari et al. 2013). Current yearly fire occurrence in Europe ranges from less than five per NUTS3Footnote 1 to nearly a hundred in areas of the Mediterranean region, which also presents the largest average of area burned yearly, with over 10,000 ha/year in some NUTS (European Commission 2010). Four types of areas can be identified in Europe, based on their fire regimes, when combining both the occurrences of fire and the average area burned in each NUTS3 (Fig. 8.2). Central France, North-Eastern Germany, and most of Romania present small fire regimes , with few fires (< 20 per year) and little area burned (< 35 ha). Poland, most of the Baltic and Scandinavian countries are areas with relatively high occurrences of fire (> 50 per year) but small area burned (< 35 ha). In contrast, most of Bulgaria and Greece are regions where a small number of fires (< 20 per year) are sufficient to burn large areas (> 115 ha). Finally, Southern Italy, Croatia and the Iberian Peninsula are areas with both high fire frequency (> 50 per year) and large areas burned (> 115 ha).

Occurrence and intensity of fires in Europe over the 2005–2010 period. The average yearly occurrence of fire and average area burned (ha) for the 2005–2010 period, per NUTS3 administrative unit were calculated, only including NUTS3 for which data were available for at least 4 years. For both metrics, the data were split in two groups around the median value. The double color ramp allows to identify areas with high number of fire but low area burned (yellow), areas with low occurrence of fire but large burned areas (orange), areas with few fire and small areas burned (green), and areas with both high occurrence of fire and large burned areas (red). (Source: EFFIS for the fire data (European Commission 2010) and © EuroGeographics for the map of administrative boundaries)
Fire suppression is a common land management policy implemented to protect human communities and land (Shlisky et al. 2007; Fernandes 2013) but it also promotes fuel accumulation in fire-dependent systems and increases the risk of large and intense fires (Proença et al. 2010; Fernandes 2013). On the other hand, fire has also been extensively used as a tool to clear landscapes and reduce fire risk. In Europe, anthropogenic fires are often more frequent than natural fires. High frequency fire regimes can cause species community impoverishment, through the exclusion of fire sensitive species and the promotion of fire resilient species that can endure frequent fires, and it can also cause extensive soil degradation and nutrient loss (Thonicke et al. 2001). This is particularly true for Mediterranean ecosystems, where 93 % of fire regimes are considered to be in a degraded or very degraded state (Shlisky et al. 2007).
Today, farmland abandonment is driving further changes in fire regimes across Europe, particularly in Southern Europe, with potential impacts for biodiversity and ecosystem services (Mouillot et al. 2005; Bassi et al. 2008; Proença and Pereira 2010). Where the number of ignitions is not a limiting factor, which is true in many regions under farmland abandonment (Bassi et al. 2008; Ganteaume et al. 2013), climate and fuel availability will be the main determinants of future changes to the fire regime. In high-productivity ecosystems with a high level of humidity, such as temperate broadleaved forests, fires will be limited by climate and humidity level, and less responsive to changes in fuel accumulation, since fuel is already a non-limiting factor (Pausas and Ribeiro 2013). Vegetation will be more susceptible to fire during warmer seasons following droughts, when the existing fuel is more flammable (Proença et al. 2010; Pausas and Ribeiro 2013). In low-productivity ecosystems, such as arid Mediterranean scrublands, fuel is the main limiting factor and will be the main driver of shifts in the fire regime (Pausas and Fernández-Muñoz 2012; Pausas and Ribeiro 2013). Recent trends in the Western Mediterranean Basin support the above predictions (Pausas and Fernández-Muñoz 2012). In this region, fields used to be grazed, frequently burned (small scale) and cleared for farming and timber (Proença and Pereira 2010), limiting fuel availability. The rural exodus since the mid-twentieth century led to shrub encroachment and afforestation with fire-prone species, and resulted in more frequent, more intense and larger fires. Today, increased fuel load and spatial continuity are driving a shift in the fire regime, which is becoming more responsive to drought, similar to high-productivity ecosystems (Pausas and Fernández-Muñoz 2012). In the future, the response of fire regime to changes in climatic variables, such as precipitation, is expected to be non-linear (Batllori et al. 2013): while a small decrease in annual precipitation may increase probability of fire, a large decrease may lead to the inverse response due to a drop in ecosystem productivity, leading the system back to a fuel-limited fire regime .
4 Disturbances and Diversity
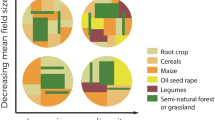
Traditional landscapes in Europe, in particular High Nature Value (HNV) farmland areas, are acknowledged for their high species richness and conservation value (Blondel and Aronson 1999; MacDonald et al. 2000; EEA 2004). Species diversity patterns in traditional landscapes are likely to be different from what would be found in non-modified (primary) landscapes (Blondel and Aronson 1999). When the total number of species is considered, a higher richness of species at the habitat patch scale (i.e., α-diversity) is expected in traditional landscapes due to species being able to use more than one habitat and due to the high density of habitat edges, which facilitates inter-patch movements and therefore leads to a higher species turnover in space and time (Proença and Pereira 2013; Guilherme and Pereira 2013). Note that even with inter-patch movements, each habitat type will support a distinct community of species due to differences in species abundances and due to the existence of strict habitat specialists. As a result, the α-diversity is probably lower in the case of specialist species in traditional mosaics due to the effect of habitat fragmentation and their low tolerance to the conditions found in other habitats (Proença and Pereira 2013). For instance, the diversity of forest species is lower in fragmented forest patches than in an area of similar size in continuous habitat (Proença 2009). Regarding species turnover (i.e., β-diversity) , traditional landscapes can have a higher turnover than former undisturbed land (Blondel and Aronson 1999), due to their mosaic structure. However, the soundness of this assumption depends on the scale of the analysis (see Chap. 6). For example, one can predict that the replication of the traditional habitat mosaic across large spatial scales results in a higher similarity of (modified) habitats, which promotes the presence of similar communities across large areas. Finally, the effect of these changes on the total number of species found in the landscape (i.e., γ-diversity) is less straightforward. Indeed, whilst several species suffered declines or even extinctions due to habitat destruction or modification (e.g., bear, auroch) , other species benefited from these changes and proliferated in the human-modified habitats (e.g., farmland birds). Moreover, starting in the earliest Neolithic, farmers continually and intentionally introduced new species to European ecosystems (Blondel and Aronson 1999). They also did so unintentionally as a result of species dispersal by animal herds along transhumance routes (e.g. Poschlod et al. 1998). Both of these activities thus increased the regional species pool, though globally richness declined due to extinctions .
4.1 Diversity and Intermediate Disturbance
The intermediate disturbance hypothesis (Connell 1978) and the diversity-disturbance hypothesis (Huston 1979) are often used to explain the ecological mechanisms determining the high diversity of species found in traditional landscapes (e.g. Blondel 2006): species diversity peaks when communities are exposed to moderate disturbance, in terms of frequency, extent and intensity. This occurs because moderate disturbance (e.g., moderate grazing) creates discontinuities in the ecosystem that allow the maintenance of early successional species while preventing dominance of more competitive species, hence keeping the ecosystem in a transitional state between early and steady-state communities. The management of traditional landscape mosaics (Fig. 8.3), with low intensity farming, moderate grazing and maintenance of forest patches, is often described as an example of intermediate disturbance, and therefore as a promoter of species diversity (Ostermann 1998; Henle et al. 2008). Nonetheless, peaked relationships between species richness and disturbance are not the rule across ecology studies (Mackey and Currie 2001). Peaked curves are more commonly reported by studies covering small spatial scales and in the presence of natural disturbance regimes (Mackey and Currie 2001). In addition, the relationship between taxa richness and the intensity of anthropogenic disturbance regimes is often non-significant (Mackey and Currie 2001), increasing the challenge of predicting the impacts of altered regimes of disturbances on biodiversity.

Minifundia system in a mountain landscape in Northwestern Portugal. (Photo credit: Vânia Proença)
4.2 Effects of Land-Use Change on Disturbance Regimes
Land-use changes caused by rural abandonment can create the conditions for an increase in the frequency and intensity of disturbance events, in particular higher fire risk due to fuel accumulation and shrub encroachment, but may also result in fewer disturbances if disturbance agents, such as domestic grazers or browsers, become residual or even disappear. The trajectory of secondary succession after abandonment depends on several interacting factors and ecological filters, such as the pool of colonizer species in the surrounding landscape, their ability to colonize abandoned patches, soil quality, and, of course, disturbance regime (Cramer 2007). Disturbances will not only exert a selective pressure on community assembly, but will also respond to community structure and composition.
In landscapes where tree density is very low, such as some Mediterranean landscapes, there is a high probability of shrub encroachment after farmland abandonment due to seed limitation, predatory pressure over oak acorns and deficient abiotic conditions, such as poor soils (Acácio et al. 2007). Wildfire will further promote shrub dominance, due to many shrubs’ resprouting ability. Wildfires may hence establish a reinforcing feedback loop, leading to community homogenization and a decline in diversity at all scales (Proença and Pereira 2010).
A different trajectory can be anticipated in landscapes with a higher tree density, such as semi-natural grasslands in northern Europe (Eriksson et al. 2002). There, seed availability and dispersal are not limiting factors and forest is able to colonize and regenerate in relatively short time. With an expected low disturbance regime, forest can expand, which would decline habitat heterogeneity . Some species, such as grassland specialists, will show strong reductions in abundance or even go locally extinct. Impacts at the landscape level will depend on species ability to persist in alternative habitats such as forest edges or heathlands (Proença and Pereira 2013).
The above examples describe abandoned patches in a fairly homogenous landscape matrix with either a low or high tree density. In a heterogeneous landscape with a more balanced cover of different habitats and a variety of edaphic-topographic conditions, scenarios would be different given the diversity of local responses to changes in disturbance regime. Habitat diversity will not only counteract landscape homogenization, but also provide alternative habitats for species affected by farmland abandonment, thus reducing the impact of land-use change on species diversity. The persistence of those species in the landscape will then depend on the maintenance of those alternative habitats, either by natural processes, such as herbivory by wild ungulates , or through assisted processes, such as prescribed fire or herbivore re-introduction. Such restoration approaches, either passive or assisted, are an important open question in rewilding research.
5 Maintaining Disturbance-Dependent Habitats
5.1 Wild Herbivores: Natural (Re)colonization or (Re)introduction?
Today, only 16 % of the Palearctic region, including Europe, contains areas occupied by relatively undisturbed large mammal faunas, i.e., species that have not undergone major changes in range between AD 1500 and the present (Morrison et al. 2007) . This figure does not even consider the number of species that went extinct early in the Holocene (Table 8.1). There is also a clear regional difference when looking at the current species richness of large herbivores in Europe (Fig. 8.4) : countries of central Europe present the highest diversity, while the Westernmost countries have low richness. Species rich areas, with lower human densities and less pressure on the land, could become “sources” for natural re-colonization. This has already been documented for some species of large herbivores that show substantial increases in their populations since the 1960s (Table 8.2). Though legislation and conservation measures largely contributed to it (Deinet et al. 2013), rural depopulation and the associated reduced human pressure, both direct (e.g. less hunting) and indirect (e.g. more land available), can also explain the phenomena (Table 8.2). Wild populations can also benefit from the absence of competitor and predator species (Bradshaw et al. 2003), though unregulated population growth could become an issue, e.g. if their pressure on the land is too high .

In cases where the local richness of wild herbivores is low, as for example in Western European countries (Fig. 8.4), species can be introduced to restore ecosystem functioning (Sandom et al. 2013b). That is, provided that their functional role is left unattended (Lipsey and Child 2007), and that the abandoned land meets their requirement in natural resources. A study on fenced populations of wild boar showed that their rooting behavior can create germination niches (Sandom et al. 2013a) and contribute to forest regeneration. However, they can also be detrimental to the established trees when bark stripping and uprooting (Sandom et al. 2013b). Reintroducing ecosystem engineers to restore and/or maintain disturbance dependent habitats can also be more time and cost effective than man-made restoration (Byers et al. 2006; Sandom et al. 2013a). Moreover, provided that the re-introduced species present charismatic values, their presence could facilitate the acceptation of a rewilding project by the public (Lipsey and Child 2007; Kuemmerle et al. 2012). The reintroduction of wild grazers can also be assessed positively from the standpoint of ecosystem services, based on the existence value of the megafauna (Proença et al. 2008) and associated cultural services (e.g. tourism, hunting, and see Chap. 3) .
Nonetheless, a balance must be maintained when considering the (re)introduction of herbivores and many potential challenges should be raised and discussed (Seddon et al. 2007; Corlett 2012; IUCN 2013). First, which species should be reintroduced? When taxon substitutions are needed for ecological replacement (IUCN 2013), researchers’ opinions are divided, ranging from releasing breeds of domesticated animals, to the reintroduction of extant relatives of long gone species (e.g. Donlan et al. 2006). Releasing animals also raises the question of increasing the risk of conflicts between local human populations and “wildlife” (e.g. Enserink and Vogel 2006; Goulding and Roper 2002), which could be more easily accepted if the species was progressively, and naturally, recolonizing an area. For reintroduced domestic species (e.g. horses), a legal framework on the liability of the organization that performed the reintroduction is also missing, for instance in cases of damages or accidents. Finally, an overabundance of certain species can have detrimental effects, especially when the natural predator guild is absent and cannot regulate the populations (see Chap. 4), yet no specific guidelines are designed for the natural control of reintroduced populations (IUCN 2013). For instance, the large populations of browsers in the Scottish Highlands, where large carnivores have been extinct for centuries, currently limit the natural forest regeneration (Sandom et al. 2013b) .
5.2 Prescribed Burning
Fire can be used as a tool in landscape management for two main intents: to control fire risk and the intensity of wildfires, and to manage landscape structure and biodiversity. Prescribed fires are often used as a preventive measure to control fuel load and fire intensity (e.g. Fernandes 2013). In addition, the combination of different fire regimes can be used to maintain landscape heterogeneity and habitat for species dependent on different ecosystem successional stages (Driscoll et al. 2010). In regions where fire risk and shrub encroachment are paired threats to biodiversity conservation, fire can be used as a tool to approach both problems (Moreira and Russo 2007) .
Nonetheless, the use of prescribed fires can also raise some conservation issues. For instance, prescribed fires are performed during the wet season (winter to spring) when there is a low risk of fire spreading, while natural fires occur during dry days, especially in summer. This divergence in fire season can negatively impact species that reproduce in spring (van Andel and Aronson 2012), such as ground nesting birds, but also the persistence of plant species, for example by causing premature seed release, or by destroying seedlings of annual plants before they create a seed bank (Whelan 1995; Bowman et al. 2013). Another issue is the implications of prescribed fires for climate change mitigation. Large scale prescribed fires may aggravate climate change, due to the emission of greenhouse gases and aerosol particles (Russell-Smith et al. 2009; Fernandes et al. 2013). While more research is needed to understand the effects of prescribed burning on carbon cycle (Fernandes 2013), it is also accepted that well planned prescribed burning prevents larger losses of carbon to the atmosphere by reducing wildfire risk (Bowman et al. 2013; Fernandes 2013). Finally, defining the regime of prescribed fires can be challenging (Whelan 1995; van Andel and Aronson 2012). Replicating natural fire regimes may not be possible, due to the lack of historic information. It may even not be advisable, given changes in landscape structure and, in some areas, in local climate, which may lead to unpredicted responses to fire (Driscoll et al. 2010). Therefore induced fire regimes should be planned to meet the desired outcomes instead of trying to mimic the parameters of natural fire regimes (Whelan 1995). In particular, in a rewilding context, fire dynamics should only be managed, or “assisted” in the early stages post-abandonment in order to facilitate the restoration of natural fire regimes .
6 Concluding Remarks
Millennia of human activities have progressively replaced natural disturbances, such as herbivory and fire, to shape the European landscapes. Maintaining disturbance-dependent habitats after the withdrawal of those human activities is a difficult restoration process. It can be guided by knowledge of the past (Vera 2000; Gillson and Willis 2004; Willis and Birks 2006; Sandom et al. 2013b), and by improving our ability to understand ecosystem dynamics and projecting potential restoration pathways. This means identifying the most desirable outcome in terms of both biodiversity and resilience. Nonetheless, besides human impacts on the landscapes, other biotic and abiotic alterations have also led to the current ecosystem composition. The climate has changed during the past millennium and some species have gone extinct while others have invaded, all these changes influencing ecological processes (Gillson and Willis 2004; Hodder et al. 2009). The interaction between human pressure and natural changes (e.g. non-anthropogenic climatic changes) could also have led to the crossing of tipping points (Gillson and Willis 2004; Kaplan et al. 2010; Leadley et al. 2014). Returning the landscapes to their historical conditions would thus be unachievable, if even desirable. This means that the baseline must shift, not only for the policy makers and the public who attribute cultural values to a relatively recent landscape (Vera 2009), but also for scientists and conservationist, some of which, on the contrary, having too long of a memory of the European landscape.
An additional concern emerges with farmland abandonment when herbivores also become functionally extinct, following a decrease in agricultural activities (Donlan et al. 2006), while the artificial fire regime is altered. Hence, in the early stages after land abandonment, the “restoration goals” must be defined to determine the set of biotic and abiotic factors that might be managed (Byers et al. 2006). Supporting local populations of wild herbivores, reintroducing them in places where they are absent and using prescribed burning can constitute the first steps towards restoring ecological processes.
For instance, the choice between natural recolonizations, reinforcement of local populations or reintroductions will depend on the current distribution and abundances of the herbivore communities. In areas of Central Europe, one might expect that the diversity of herbivores is high enough to allow for recolonizations, while in Western and Southern Europe, active introduction might be needed (Fig. 8.4). In all cases, conservation measures, legislation and reduced human pressure are necessary for the establishment of viable populations (Table 8.2).
When rewilding is meant to lead to ecological restoration , reintroductions should be one of the tools rather than a goal per se. Moreover, historical baselines should be treated as guidelines, not as objectives. In other words, rather than focusing on the conservation of a given set of species or habitats, rewilding will focus on the restoration and conservation of natural processes, with human intervention reduced to its minimum.
Acknowledgments
We thank Thomas Merckx and Christopher Sandom for insightful comments on earlier versions of the manuscript. We are also thankful to the Societas Europaea Mammalogica for sharing their Atlas data. L.M.N. and V.P. were supported by fellowships from the FCT (SFRH/BD/62547/2009 and SFRH/BPD/80276/2011). JOK was supported by a grant from the European Research Council (COEVOLVE, 313797).
Notes
- 1.
Third level of the EU Nomenclature of Territorial Units for Statistics.
References
Acácio, V., Holmgren, M., Jansen, P.A., & Schrotter, O. (2007). Multiple recruitment limitation causes arrested succession in Mediterranean Cork Oak Systems. Ecosystems, 10(7), 1220–1230.
Archibald, S., Lehmann, C.E.R., Gómez-Dans, J.L., & Bradstock, R.A. (2013). Defining pyromes and global syndromes of fire regimes. Proceedings of the National Academy of Sciences, 110(16), 6442–6447.
Attiwill, P. M. (1994). The disturbance of forest ecosystems: The ecological basis for conservative management. Forest Ecology and management, 63(2), 247–300.
Bassi, S., Kettunen, M., Kampa, E., & Cavalieri, S. (2008). Forest fires: causes and contributing factors to forest fire events in Europe. Study for the European Parliament Committee on Environment, Public Health and Food Safety under contract IP/A/ENVI/FWC/2006-172/LOT1/C1/SC10. Institute for European Environmental Policy, London, United Kingdom. 56p.
Batllori, E., Parisien, M.-A., Krawchuk, M. A., & Moritz, M. A. (2013). Climate change-induced shifts in fire for Mediterranean ecosystems. Global Ecology and Biogeography, 22(10), 1118–1129.
Birks, H. J. B. (2005). Mind the gap: How open were European primeval forests? Trends in Ecology and Evolution, 20(4), 151–154.
Blondel, J. (2006). The “Design” of Mediterranean landscapes: A millennial story of humans and ecological systems during the historic period. Human Ecology, 34(5), 713–729.
Blondel, J., & Aronson, J. (1999). Biology and wildlife of the Mediterranean Region. Oxford University Press, New York, USA.
Bond, W. J., & Keeley, J. E. (2005). Fire as a global “herbivore”: The ecology and evolution of flammable ecosystems. Trends in Ecology and Evolution, 20(7), 387–394.
Bowman, D. M. J. S., O’Brien, J. A., & Goldammer, J. G. (2013). Pyrogeography and the global quest for sustainable fire management. Annual Review of Environment and Resources, 38(1), 57–80.
Bradshaw, R. H., Hannon, G. E., & Lister, A. M. (2003). A long-term perspective on ungulate-vegetation interactions. Forest Ecology and Management, 181(1–2), 267–280.
Bullock, D. J. (2009). What larger mammals did Britain have and what did they do? British Wildlife, 20(5), 16–20.
Byers, J.E., Cuddington, K., Jones, C.G., Talley, T.S., Hastings, A., Lambrinos, J.G., Crooks, J.A., & Wilson, W.G. (2006). Using ecosystem engineers to restore ecological systems. Trends in Ecology and Evolution, 21(9), 493–500.
Carbonell, E., Bermúdez de Castro, J.M., Parés, J.M., Pérez-González, A., Cuenca-Bescós, G., Ollé, A., Mosquera, M., Huguet, R., van der Made, J., Rosas, A., Sala, R., Vallverdú, J., García, N., Granger, D.E., Martinón-Torres, M., Rodríguez, X.P., Stock, G.M., Vergès, J.M., Allué, E., Burjachs, F., Cáceres, I., Canals, A., Benito, A., Díez, C., Lozano, M., Mateos, A., Navazo, M., Rodríguez, J., Rosell, J., & Arsuaga, J.L. (2008). The first hominin of Europe. Nature, 452(7186), 465–469.
Connell, J. H. (1978). Diversity in tropical rain forests and coral reefs. Science, 199(4335), 1302–1310.
Corlett, R. T. (2012). The shifted baseline: Prehistoric defaunation in the tropics and its consequences for biodiversity conservation. Biological Conservation, 163, 13–21.
Cramer, V. A. (2007). Old fields as complex systems: New concepts for describing the dynamics of abandoned farmland. In V. A. Cramer & R. J. Hobbs (Eds.), Old fields: Dynamics and restoration of abandoned farmland (p. 334). Washington, DC: Island Press.
Daniau, A. L., d’ Errico, F., & Goñi, M. F. S. (2010). Testing the hypothesis of fire use for ecosystem management by Neanderthal and Upper Palaeolithic modern human populations. PloS ONE, 5(2), e9157.
Deinet, S., Ieronymidou, C., McRae, L., Burfield, I.J., Foppen, R.P., Collen, B., & Bohm, M. (2013). Wildlife comeback in Europe: The recovery of selected mammal and bird species. Final report to Rewilding Europe by ZSL. London: BirdLife International and the European Bird Census Council.
Donlan, C.J., Berger, J., Bock, C.E., Bock, J.H., Burney, D.A., Estes, J.A., Foreman, D., Martin, P.S., Roemer, G.W., & Smith, F.A. (2006). Pleistocene rewilding: An optimistic agenda for twenty-first century conservation. American Naturalist, 168(5), 660–681.
Driscoll, D.A., Lindenmayer, D.B., Bennett, A.F., Bode, M., Bradstock, R.A., Cary, G.J., Clarke, M.F., Dexter, N., Fensham, R., Friend, G., Gill, M., James, S., Kay, G., Keith, D.A., MacGregor, C., Russell-Smith, J., Salt, D., Watson, J.E.M., Williams, R.J., & York, A. (2010). Fire management for biodiversity conservation: Key research questions and our capacity to answer them. Biological Conservation, 143(9), 1928–1939.
EEA. (2004). High nature value farmland: Characteristics, trends and policy challenges. Copenhagen: European Environment Agency.
Eickhout, B., Van Meijl, H., Tabeau, A., & Van Rheenen, T. (2007). Economic and ecological consequences of four European land use scenarios. Land use policy, 24(3), 562–575.
Enserink, M., & Vogel, G. (2006). The carnivore comeback. Science, 314(5800), 746.
Eriksson, O., Cousins, S. A. O., & Bruun, H. H. (2002). Land-use history and fragmentation of traditionally managed grasslands in Scandinavia. Journal of Vegetation Science, 13(5), 743–748.
European Commission. (2010). Forest Fires in Europe 2009. Luxemburg: Office for Official Publications of the European Communities.
FAOSTAT. (2010). Data. http://faostat.fao.org. Accessed 1 March 2011
Fernandes, P. M. (2013). Fire-smart management of forest landscapes in the Mediterranean basin under global change. Landscape and Urban Planning, 110, 175–182.
Fernandes, P.M., Davies, G.M., Ascoli, D., Fernández, C., Moreira, F., Rigolot, E., Stoof, C.R., Vega, J.A., & Molina, D. (2013). Prescribed burning in southern Europe: Developing fire management in a dynamic landscape. Frontiers in Ecology and the Environment, 11(s1), e4–e14.
Fyfe, R. (2007). The importance of local-scale openness within regions dominated by closed woodland. Journal of Quaternary Science, 22(6), 571–578.
Ganteaume, A., Camia, A., Jappiot, M., San-Miguel-Ayanz, J., Long-Fournel, M., & Lampin, C. (2013). A review of the main driving factors of forest fire ignition over Europe. Environmental Management, 51(3), 651–662.
Gignoux, C. R., Henn, B. M., & Mountain, J. L. (2011). Rapid, global demographic expansions after the origins of agriculture. Proceedings of the National Academy of Sciences, 108(15), 6044–6049.
Gillson, L., & Willis, K. J. (2004). “As Earth’s testimonies tell”: Wilderness conservation in a changing world. Ecology Letters, 7(10), 990–998.
Goulding, M. J., & Roper, T. J. (2002). Press responses to the presence of free-living Wild Boar (Sus scrofa) in southern England. Mammal Review, 32(4), 272–282.
Guilherme, J. L., & Pereira, H. M. (2013). Adaptation of bird communities to farmland abandonment in a mountain landscape. PLoS ONE, 8(9), e73619.
Henle, K., Alard, D., Clitherow, J., Cobb, P., Firbank, L., Kull, T., McCracken, D., Moritz, R.F.A., Niemelä, J., Rebane, M., Wascher, D., Watt, A., & Young, J. (2008). Identifying and managing the conflicts between agriculture and biodiversity conservation in Europe-a review. Agriculture, Ecosystems & Environment, 124(1-2), 60–71.
Hodder, K.H., Buckland, P.C., Kirby, K.J., & Bullock, J.M. (2009). Can the Pre-Neolithic provide suitable models for re-wilding the landscape in Britain? British Wildlife, 20(5), 4–15.
Huston, M., (1979). A general hypothesis of species diversity. American Naturalist, 113(1), 81–101.
IUCN. (2013). Guidelines for reintroductions and other conservation translocations. Version 1.0., Gland. Switzerland: IUCN Species Survival Commission.
Johnson, C. N. (2009). Ecological consequences of late Quaternary extinctions of megafauna. Proceedings of the Royal Society B: Biological Sciences, 276, 2509–2519.
Jones, C. G., Lawton, J. H., & Shachak, M., (1994). Organisms as ecosystem engineers. Oikos, 69, 373–386.
Kaplan, J. O. (2012). Integrated modeling of Holocene land cover change in Europe. Quaternary International, 279, 235–236.
Kaplan, J. O., Krumhardt, K. M., & Zimmermann, N. (2009). The prehistoric and preindustrial deforestation of Europe. Quaternary Science Reviews, 28(27–28), 3016–3034.
Kaplan, J.O., Krumhardt, K.M., Ellis, E.C., Ruddiman, W.F., Lemmen, C., & Goldewijk, K.K. (2010). Holocene carbon emissions as a result of anthropogenic land cover change. The Holocene, 21(5), 775–791.
Kuemmerle, T., Hickler, T., Olofsson, J., Schurgers, G., & Radeloff, V.C. (2012). Refugee species: Which historic baseline should inform conservation planning? Diversity and Distributions, 18(12), 1258–1261.
Kuiters, A. T., & Slim, P. A. (2003). Tree colonisation of abandoned arable land after 27 years of horse-grazing: The role of bramble as a facilitator of oak wood regeneration. Forest Ecology and Management, 181(1–2), 239–251.
Laskurain, N. A., Aldezabal, A., Olano, J. M., Loidi, J., & Escudero, A. (2013). Intensification of domestic ungulate grazing delays secondary forest succession: Evidence from exclosure plots. Journal of Vegetation Science, 24(2), 320–331.
Leadley, P., Proença, V., Fernández-Manjarrés, J., Pereira, H.M., Alkemade, R., Biggs, R., Bruley, E., Cheung, W., Cooper, D., Figueiredo, J., Gilman, E., Guénette, S., Hurtt, G., Mbow, C., Oberdorff, T., Revenga, C., Scharlemann, J.P.W., Scholes, R., Smith, M.S., Sumaila, U.R., & Walpole, M. (2014). Interacting regional-scale regime shifts for biodiversity and ecosystem services. BioScience, 64(8), 665–679.
Lipsey, M. K., & Child, M. F. (2007). Combining the fields of reintroduction biology and restoration ecology. Conservation Biology, 21(6), 1387–1390.
MacDonald, D., Crabtree, J.R., Wiesinger, G., Dax, T., Stamou, N., Fleury, P., Gutierrez Lazpita, J., & Gibon, A. (2000). Agricultural abandonment in mountain areas of Europe: Environmental consequences and policy response. Journal of Environmental Management, 59(1), 47–69.
Mackey, R. L., & Currie, D. J. (2001). The diversity-disturbance relationship: Is it generally strong and peaked? Ecology, 82(12), 3479–3492.
Mather, A. S., Needle, C. L., & Fairbairn, J. (1998). The human drivers of global land cover change: The case of forests. Hydrological processes, 12(13–14), 1983–1994.
Mellars, P. (2006). A new radiocarbon revolution and the dispersal of modern humans in Eurasia. Nature, 439(7079), 931–935.
Mitchell, F. J. G. (2005). How open were European primeval forests? Hypothesis testing using palaeoecological data. Journal of Ecology, 93(1), 168–177.
Mitchell-Jones, A.J., Amori, G., Bogdanowicz, W., Krystufek, B., Reijnders, P.J.H., Spitzenberger, F., Stubbe, M., Thissen, J.B.M., Vohralik, V., & Zima, J. (1999). The atlas of European mammals. http://www.european-mammals.org/. Accessed July 27th 2013.
Molinari, C., Lehsten, V., Bradshaw, R.H.W., Power, M.J., Harmand, P., Arneth, A., Kaplan, J.O., Vannière, B., & Sykes, M.T. (2013). Exploring potential drivers of European biomass burning over the Holocene: A data-model analysis. Global Ecology and Biogeography, 22(12), 1248–1260.
Moreira, F., & Russo, D., (2007). Modelling the impact of agricultural abandonment and wildfires on vertebrate diversity in Mediterranean Europe. Landscape Ecology, 22, 1461–1476.
Morrison, J.C., Sechrest, W., Dinerstein, E., Wilcove, D.S., & Lamoreux, J.F. (2007). Persistence of large mammal faunas as indicators of global human impacts. Journal of Mammalogy, 88(6), 1363–1380.
Mouillot, F., Ratte, J.-P., Joffre, R., Mouillot, D., & Rambal, S. (2005). Long-term forest dynamic after land abandonment in a fire prone Mediterranean landscape (central Corsica, France). Landscape Ecology, 20(1), 101–112.
Ostermann, O. P. 1998. The need for management of nature conservation sites designated under Natura 2000. Journal of Applied Ecology, 35(6), 968–973.
Pausas, J. G., & Bradstock, R. A. (2007). Fire persistence traits of plants along a productivity and disturbance gradient in mediterranean shrublands of south-east Australia. Global Ecology and Biogeography, 16(3), 330–340.
Pausas, J. G., & Fernández-Muñoz, S. (2012). Fire regime changes in the Western Mediterranean Basin: From fuel-limited to drought-driven fire regime. Climatic Change, 110(1–2), 215–226.
Pausas, J. G., & Ribeiro, E. (2013). The global fire-productivity relationship. Global Ecology and Biogeographyl, 22(6), 728–736.
Pausas, J. G., Keeley, J. E., & Verdú, M. (2006). Inferring differential evolutionary processes of plant persistence traits in Northern Hemisphere Mediterranean fire-prone ecosystems. Journal of Ecology, 94(1), 31–39.
Pereira, H. M., Navarro, L. M., & Martins, I. S. (2012). Global biodiversity change: The bad, the good, and the unknown. Annual Review of Environment and Resources, 37(1), 25–50.
Pfeiffer, M., Spessa, A., & Kaplan, J. O. (2013). A model for global biomass burning in preindustrial time: LPJ-LMfire (v1.0). Geoscientific Model Development, 6(3), 643–685.
Pinhasi, R., Fort, J., & Ammerman, A. J. (2005). Tracing the origin and spread of agriculture in Europe. PLoS Biology, 3(12), e410.
Poschlod, P., Kiefer, S., Tränkle, U., Fischer, S., & Bonn, S. (1998). Plant species richness in calcareous grasslands as affected by dispersability in space and time. Applied Vegetation Science, 1(1), 75–91.
Proença, V. (2009). Galicio-Portuguese oak forest of Quercus robur and Quercus pyrenaica: Biodiversity patterns and forest response to fire. Tese de Doutoramento. Lisboa: Faculdade de Ciências da Universidade de Lisboa.
Proença, V., & Pereira, H. M. (2010). Appendix 2: Mediterranean Forest (pp. 60–67). Leadley, P., Pereira, H.M., Alkemade, R., Fernandez-Manjarrés, J.F., Proença, V., Scharlemann, J.P.W., Walpole, M.J. (Eds.) Biodiversity Scenarios: Projections of 21st century change in biodiversity and associated ecosystem services. Secretariat of the Convention on Biological Diversity, Montreal. Technical Series no. 50, 132 pages.
Proença, V., & Pereira, H. M. (2013). Species-area models to assess biodiversity change in multi-habitat landscapes: The importance of species habitat affinity. Basic and Applied Ecology, 14, 102–114.
Proença, V. M., Pereira, H. M., & Vicente, L. (2008). Organismal complexity is an indicator of species existence value. Frontiers in. Ecology and the Environment, 6(6), 298–299.
Proença, V., Pereira, H. M., & Vicente, L. (2010). Resistance to wildfire and early regeneration in natural broadleaved forest and pine plantation. Acta Oecologica, 36(6), 626–633.
Rey Benayas, J.M., Martins, A., Nicolau, J.M., & Schulz, J.J. (2007). Abandonment of agricultural land: An overview of drivers and consequences. CAB reviews: Perspectives in Agriculture, Veterinary Science, Nutrition and Natural Resources, 2(57), 1–14.
Ruddiman, W. F. (2013). The Anthropocene. Annual Review of Earth and Planetary Sciences, 41(1), 1–24.
Russell-Smith, J., Murphy, B.P., Meyer, C.P., Cook, G.D., Maier, S., Edwards, A.C., Schatz, J., & Brocklehurst, P. (2009). Improving estimates of savanna burning emissions for greenhouse accounting in northern Australia: Limitations, challenges, applications. International Journal of Wildland Fire, 18(1), 1–18.
Sandom, C. J., Hughes, J., & Macdonald, D. W. (2013a). Rooting for rewilding: Quantifying wild boar’s Sus scrofa rooting rate in the scottish highlands. Restoration Ecology, 21(3), 329–335.
Sandom, C., Donlan, C.J., Svenning, J.C., & Hansen, D. (2013b). Rewilding. In D. W. Mcdonald & K. J. Willis (Eds.), Key Topics in Conservation Biology 2 (pp. 430–451). Oxford: John Wiley & Sons.
Sandom, C.J., Ejrnæs, R., Hansen, M.D.D., & Svenning, J.-C. (2014). High herbivore density associated with vegetation diversity in interglacial ecosystems. Proceedings of the National Academy of Sciences, 111(11), 4162–4167.
Schley, L., Dufrêne, M., Krier, A., & Frantz, A.C. (2008). Patterns of crop damage by wild boar (Sus scrofa) in Luxembourg over a 10-year period. European Journal of Wildlife Research, 54(4), 589–599.
Seddon, P. J., Armstrong, D. P., & Maloney, R. F. (2007). Developing the science of reintroduction biology. Conservation biology, 21(2), 303–312.
Shlisky, A., Waugh, J., Gonzalez, P., Gonzalez, M., Manta, M., Santoso, H., Alvarado, E., Nuruddin, A.A., Rodríguez-Trejo, D.A., & Swaty, R. (2007). Fire, ecosystems and people: Threats and strategies for global biodiversity conservation. Arlington: The Nature Conservancy.
Smith, F.A., Lyons, S.K., Ernest, S.K.M., Jones, K.E., Kaufman, D.M., Dayan, T., Marquet, P.A., & Haskell, J.P. (2003). Body mass of late Quaternary mammals. Ecology, 84, 3402.
Svenning, J. C. (2002). A review of natural vegetation openness in north-western Europe. Biological Conservation, 104(2), 133–148.
Thonicke, K., Venevsky, S., Sitch, S., & Cramer, W. (2001). The role of fire disturbance for global vegetation dynamics: Coupling fire into a dynamic global vegetation model. Global Ecology and Biogeography, 10(6), 661–677.
Turner, M. G. (1998). Landscape ecology, living in a mosaic. In S. I. Dodson et al., (Eds.), Ecology (pp. 78–122). New York: Oxford University Press.
Van Andel, J., & Aronson, J., (2012). Restoration ecology: The new frontier. Chichester: Wiley-Blackwell.
Vera, F. W. M. (2000). Grazing ecology and forest history. CABI Publishing, Oxon, UK.
Vera, F. W. M. (2009). Large-scale nature development-The Oostvaardersplassen. British Wildlife, 20(5), 28–36.
Verburg, P. H., & Overmars, K. P. (2009). Combining top-down and bottom-up dynamics in land use modeling: Exploring the future of abandoned farmlands in Europe with the Dyna-CLUE model. Landscape Ecology, 24(9), 1167–1181.
Whelan, R. J. (1995). The ecology of fire. Cambridge University Press, Cambridge, UK.
Willis, K. J., & Birks, H. J. B. (2006). What is natural? The need for a long-term perspective in biodiversity conservation. Science, 314(5803), 1261–1265.
Wright, J. P., & Jones, C. G. (2006). The concept of organisms as ecosystem engineers ten years on: Progress, limitations, and challenges. BioScience, 56(3), 203–209.
Zeder, M. A. (2008). Domestication and early agriculture in the Mediterranean Basin: Origins, diffusion, and impact. Proceedings of the National Academy of Sciences, 105(33), 11597–11604.
Zimov, S. A. (2005). Pleistocene park: Return of the mammoth’s ecosystem. Science, 308(5723), 796–798.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Open Access This chapter is distributed under the terms of the Creative Commons Attribution Noncommercial License, which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Copyright information
© 2015 The Author(s)
About this chapter
Cite this chapter
Navarro, L., Proença, V., Kaplan, J., Pereira, H. (2015). Maintaining Disturbance-Dependent Habitats. In: Pereira, H., Navarro, L. (eds) Rewilding European Landscapes. Springer, Cham. https://doi.org/10.1007/978-3-319-12039-3_8
Download citation
DOI: https://doi.org/10.1007/978-3-319-12039-3_8
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-12038-6
Online ISBN: 978-3-319-12039-3
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)