
Abstract
Dickins has made some thoughtful suggestions as to why the important contributions of inclusive fitness theory have not been more celebrated by the proponents of the Extended Evolutionary Synthesis, considering the extent to which inclusive fitness theory has accommodated and illuminated—and, indeed, been motivated by—their “laundry list” of supposedly neglected evolutionary factors. We agree that this oversight could be explained, in part, by their seeing inclusive fitness as a “monist” alternative to a more “pluralist” multilevel selection that was not part of the Modern Synthesis. Here we clarify that multilevel selection and inclusive fitness are not competing explanations, but rather they address orthogonal issues, concerning the process of selection and the purpose of adaptation, respectively. We discuss the sense in which inclusive fitness is “monist” in providing the only generally correct adaptive maximand, but also “pluralist” in the sense of accommodating a diversity of adaptive agents. We also emphasise that multilevel selection was, in fact, part of the Modern Synthesis and, indeed, its inadequacies as a theory of organismal adaptation provided a crucial motivation for the concept of inclusive fitness.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others


Keywords
- Levels of biological organisation
- Maximisation principle
- Meta-science
- Natural selection
- Purpose of adaptation
- Shifting balance
Dickins has made some thoughtful suggestions as to why the important contributions of inclusive fitness theory have not been more celebrated by the proponents of the Extended Evolutionary Synthesis, considering the extent to which inclusive fitness theory has accommodated and illuminated—and, indeed, been motivated by—their “laundry list” (Welch 2017) of supposedly neglected evolutionary factors. We agree that this oversight could be explained, in part, by their seeing inclusive fitness as a “monist” alternative to a more “pluralist” multilevel selection that was not part of the Modern Synthesis.
Here we clarify that multilevel selection and inclusive fitness are not competing explanations, but rather they address orthogonal issues, concerning the process of selection and the purpose of adaptation, respectively. We discuss the sense in which inclusive fitness is “monist” in providing the only generally correct adaptive maximand, but also “pluralist” in the sense of accommodating a diversity of adaptive agents. We also emphasize that multilevel selection was, in fact, part of the Modern Synthesis and, indeed, its inadequacies as a theory of organismal adaptation provided crucial motivation for the concept of inclusive fitness.
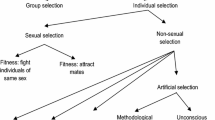
As discussed in our chapter, the kin selection and multilevel selection approaches concern different ways of separating the overall action of natural selection into its component parts (Hamilton 1975). The kin selection approach separates the total action of natural selection into its direct versus indirect components, and the multilevel selection approach separates the total action of natural selection into its within-group versus between-group components. These two approaches are empirically equivalent, in the sense that in principle they should always yield the same predictions; but they can be considered as competing explanations at a meta-scientific level, for example, in relation to how readily each approach facilitates the derivation of testable predictions (West et al. 2008).
In contrast, inclusive fitness concerns the design rationale for Darwinian adaptation (Hamilton 1964; West and Gardner 2013). As a consequence of natural selection—whether one chooses to frame this in terms of kin selection or multilevel selection—individual organisms are expected to appear designed to maximize their inclusive fitness. Only this maximand yields optima that correctly correspond to the action of natural selection. Alternative maximization principles, such as those in which individual organisms are viewed as maximizing the overall fitness of their social group, do not generally yield optima that correctly correspond to the action of natural selection; and in those scenarios in which they do yield the correct optima—for example, in the context of clonal groups in which all group mates are genetically identical—these exactly coincide with the inclusive fitness optima (Gardner and Grafen 2009; Rodrigues and Taylor 2018).
In this sense, inclusive fitness is a “monist” concept. However, it does allow “pluralism” in terms of permitting some flexibility as to the level of biological organization at which it is applied. One may view the immune system as an adaptation at the level of the individual organism, functioning to maximize inclusive fitness; or alternatively view the white blood cell as an adaptive agent in its own right, striving to maximize its own inclusive fitness; or alternatively view each of the genes that reside in the white blood cell as striving to maximize their own inclusive fitness (Gardner 2015). To the extent that these different viewpoints all yield exactly the same predictions they are all scientifically valid.
Does the dismissiveness of the proponents of the Extended Evolutionary Synthesis concerning inclusive fitness theory owe to a view that inclusive fitness is a natural entailment of the Modern Synthesis whilst multilevel selection represents a wholly different approach to understanding evolution? If so, then this view is historically unfounded. Sewall Wright , indisputably a core architect of the Modern Synthesis, placed multilevel selection firmly in the centre of his understanding of the evolutionary process, in the form of the “shifting balance ” view of adaptive evolution (Wright 1932). This concerned the rate—rather than the purpose—of adaptation. Indeed, there appears to have been a quite pervasive belief among evolutionary biologists through the first half of the twentieth century that what is best for the individual is also necessarily best for the population, such that the issue of the purpose of adaptation would have seemed trivial and scientifically uninteresting.
To his credit, Wynne-Edwards (1962) understood that selection at the within-population and between-population levels would often be in opposition. However, believing that between-population selection must always trump within-population selection, his application of multilevel selection logic led him to the erroneous view that the action of natural selection will always result in individual organisms behaving for the good of the population. The explicitness with which Wynne-Edwards set out his faulty logic—in contrast to others, such as Lorenz (1963), who simply and unreflectively assumed that animal behaviour must be understood as being for the good of the species—ignited a controversy from which there fortunately emerged a correct understanding of the purpose of adaptation. Today we understand that, irrespective of the relative strengths of different levels of selection, the purpose of adaptation is to maximize inclusive fitness (Hamilton 1964; Hamilton 1975).
References
Gardner A (2015) More on the genetical theory of multilevel selection. J Evol Biol 28:1747–1751
Gardner A, Grafen A (2009) Capturing the superorganism: a formal theory of group adaptation. J Evol Biol 22:659–671
Hamilton WD (1964) The genetical evolution of social behaviour. I. J Theor Biol 7:1–16
Hamilton WD (1975) Innate social aptitudes of man: an approach from evolutionary genetics. In: Bischof N, Fox R (eds) Biosocial anthropology. Malaby, London, pp 133–153
Lorenz K (1963) Das sogenannte Böse: zur Naturgeschichte der Aggression. Dr. G. Borotha-Schoeler, Wien
Rodrigues AMM, Taylor TB (2018) Ecological and demographic correlates of cooperation from individual to budding dispersal. J Evol Biol 31:1058–1070
Welch JJ (2017) What’s wrong with evolutionary biology? Biol Philos 32:263–279
West SA, Gardner A (2013) Adaptation and inclusive fitness. Curr Biol 23:R577–R584
West SA, Griffin AS, Gardner A (2008) Social semantics: how useful has group selection been? J Evol Biol 21:374–385
Wright S (1932) The roles of mutation, inbreeding, crossbreeding, and selection in evolution. Proceedings of the Sixth International Congress on Genetics 1:356–366
Wynne-Edwards VC (1962) Animal dispersion in relation to social behaviour. Oliver and Boyd, London
Author information
Authors and Affiliations
Corresponding authors
Editor information
Editors and Affiliations
Rights and permissions
Open Access This chapter is licensed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license and indicate if changes were made.
The images or other third party material in this chapter are included in the chapter's Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the chapter's Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
Copyright information
© 2023 The Author(s)
About this chapter

Cite this chapter
Rodrigues, A.M.M., Gardner, A. (2023). On Monism and Pluralism: A Reply to Dickins, T. E.. In: Dickins, T.E., Dickins, B.J. (eds) Evolutionary Biology: Contemporary and Historical Reflections Upon Core Theory. Evolutionary Biology – New Perspectives on Its Development, vol 6. Springer, Cham. https://doi.org/10.1007/978-3-031-22028-9_22
Download citation
DOI: https://doi.org/10.1007/978-3-031-22028-9_22
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-031-22027-2
Online ISBN: 978-3-031-22028-9
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)