
Abstract
Epigenetic effects such as gene silencing and variable expression are unintended consequences of plant transformation, a problem that is present in the transformation of all plant species. There is not yet a reliable way to prevent epigenetic silencing; however, the probability of epigenetic effects may be reduced by choosing an appropriate method of transgene introduction into a plant cell. Most methods used in plant biotechnology, such as direct gene transfer and particle bombardment, result in the introduction of multiple DNA molecules and, as a consequence, multi-copy multi-locus insertion patterns. These multiple insertions may lead to variations in transgene expression, epigenetic silencing being the most extreme. In contrast, Agrobacterium-mediated plant transformation procedures rarely cause such unintended effects. In this chapter, we present advantages and disadvantages of the Agrobacterium-mediated plant transformation method as well as protocols for transformation of Arabidopsis generative tissues and tobacco seedlings as the most classical techniques in these model plants, i.e., vacuum infiltration of explants and floral dip methods. Moreover, epigenetic effects of transgenes such as silencing related to the position and insertion effects as well as effects of the regeneration procedure causing somaclonal variation will be briefly discussed.
1 Introduction
The introduction of DNA into plant cells can be achieved by a variety of techniques, including polyethylene glycol (PEG) treatment, electroporation, microinjection, biolistic transformation as well as the use of Agrobacterium, plant viruses, and liposomes as DNA carriers (1-3).
The most commonly used method for obtaining transgenic plants involves the introduction of donor DNA into plant cells by a pathogenic bacterium Agrobacterium tumefaciens or Agrobacterium rhizogenes (4). Agrobacterium initiates T-DNA transfer from its tumor inducing (Ti) or root inducing (Ri) plasmid in response to phenolic signals produced by wounded plants. Plant phenolics then trigger the induction of the bacterial plasmid-encoded virulence (vir) genes, which in turn mediate the T-DNA transfer. The T-DNA (10-20 kb) on the Ti plasmid (Fig. 1) is flanked and delimited by border sequences (RB and LB - right and left border) that consist of 25 bp long, highly conserved direct DNA repeats. Under normal circumstances, the DNA sequence flanked by these borders is transferred to the plant, where it integrates randomly into the plant genome.

A general scheme of a pTi plasmid. T-DNA is located between LB (left border) and RB (right border), as indicated by arrowheads. Dots indicate other genetic elements, such as origin of replication, conjugal transfer region, and opine catabolism operon
The naturally occurring genes within the T-DNA are not necessary for the T-DNA transfer but are necessary for tumorigenesis and opine production. Thus, these naturally occurring T-DNA-encoded “oncogenes” can be deleted, thereby disarming the T-DNA. Such disarmed Ti plasmids are still capable of transferring T-DNA to plants, yet they will not cause disease. Thus, normal plants harboring novel gene(s) inserted between the T-DNA borders can be regenerated after cocultivation of plants with Agrobacterium carrying a modified Ti plasmid (vector). For species that are amenable to transformation and regeneration using A. tumefaciens, T-DNA-mediated gene transfer remains the method of choice because of the simplicity and efficiency of delivering a neat package of DNA that is integrated into the plant genome. In addition, this method mostly results in a single-copy, single-locus integration pattern, in contrast to other gene transfer methods (electroporation, microinjection, or particle bombardment), which suffer from a relatively low efficiency of transformation and frequent integration of catenated and/or rearranged DNA sequences (a multi-copy/multi-locus integration pattern).
Transformation methods used in plant biotechnology and modern agriculture have the potential to generate unintended genetic and epigenetic variations. (5-8). These unintended effects can be divided into three main groups: (1) insertion effects, (2) position effects, and (3) somaclonal variation. They are often observed when direct methods of DNA transfer are used; however, the Agrobacterium-mediated plant transformation method may also, although more rarely, cause unintended effects including those of epigenetic nature. This can be explained, at least partially, by the difference in transgene copy number in plants obtained by direct gene transfer (a multi-copy multi-locus integration pattern) and Agrobacterium-mediated transformation (a single-copy single locus integration pattern).
Insertion effects are defined as pleiotropic effects of integrated DNA on the host plant genome (8). The insertion effect can be mutagenic in nature, resulting in null, loss-of-function, gain-of-function, and other possible phenotypes depending on the specific region of transgene insertion into the plant DNA. Mutagenic gene disruption is not the only mechanism by which transgene insertion may affect the phenotype of a transgenic plant. Coding sequences surrounding insertion sites may fall under the influence of transgene regulatory elements (e.g., promoters) thus leading to sense and antisense transcripts, depending on the orientation of the effected host gene in relation to the transgene promoter. The sense transcripts may serve as mRNAs for recombinant (fusion) proteins, whereas the antisense transcripts may interfere with host gene expression via RNA interference (RNAi).
Position effects represent the influence of the integration site and transgene architecture on transgene expression level and stability. Most of the variations in transgene expression are attributed to differences in transgene copy number and its integration sites. This may lead to changes in locus configuration and/or induction of silencing mechanism (5). Silencing resulting from interactions among multiple copies of transgenes and related endogenous genes involves homology-based mechanisms that function at both the transcriptional and post-transcriptional levels (RNA interference) (9, 10). Surrounding sequences can also influence transgene expression, and they do it in many ways (8). First, transgene expression may be enhanced by strong promoter and/or enhancer elements, if it integrated in their vicinity. Second, specific S/MARs (scaffold/matrix attachment regions) may have a beneficial influence on transgene expression. Third, the cytosine methylation status of the insertion site may influence transgene expression leading to transcriptional silencing of transgene.
Somaclonal variation is defined as the effect of various stresses related to tissue handling, regeneration, and clonal propagation. Most of the transformation protocols apply an in vitro selection and regeneration step, and these procedures always induce somaclonal variation, which leads to changes in numerous plant characteristics, most often as an unintended effect. Somaclonal variations can be manifested as either somatically or meiotically stable events. They are exhibited as cytological abnormalities, frequent qualitative and quantitative phenotypic mutations, sequence changes, and gene activation or silencing (e.g., transposable elements and retrotransposons) (7). The main factors contributing to somaclonal variation are generally classified as stress factors. Stress response can be potentially mutagenic, and it leads to: (1) genetic changes such as polyploidy, aneuploidy, chromosome rearrangements, somatic recombination, gene amplifications, point mutations, excisions and insertions of (retro)transposones and (2) epigenetic changes including DNA methylation and histone modifications (8, 11).
Agrobacterium-mediated plant transformation has found many applications in plant biotechnology for the creation of transgenic plants with improved characteristics such as insect and herbicide resistance, phytoremediation abilities, improved nutrient content, production of biopharmaceuticals, and many others. This method is also frequently used for research purposes through T-DNA mutagenesis and creation of gene over-expressing (OE) or knock-out (KO) lines to study gene function. The same strategy and Agrobacterium vectors are used for biotechnological applications and research purposes mentioned above. Although the effect of T-DNA mutagenesis is the same as that of creation of specific knock-out plant (gene inactivation), the mechanism of gene inactivation is different. T-DNA mutagenesis is a type of insertional mutagenesis in which T-DNA integration into a gene sequence (T-DNA tagging) usually causes its inactivation. In the case of KO lines, T-DNA carries an anti-sense sequence of the gene of interest or another sequence that will anneal to target mRNA molecules to prevent gene expression (RNA interference), including secondary structure-forming transcripts (so-called hairpin vectors). Overexpression vectors contain an expression cassette composed of an appropriate promoter (e.g., 35S promoter from CaMV), a gene of interest, and a transcription termination signal (e.g., nos terminator). Introduction of such cassette into the plant genome results in overproduction of gene products that may also exhibit a phenotypic effect, shedding more light on gene function.
Thus, modified A. tumefaciens strains and T-DNA vector systems have been generated to perform multiple functions. However, there is still a need to improve the Agrobacterium-mediated plant transformation method in order to enrich the arsenal of techniques for plant genetic engineering.
2 Materials
2.1 Materials for Transformation of Tobacco
2.1.1 Sterilization of Nicotiana tabacum Seeds
-
1.
Tobacco seeds: N. tabacum cv. SR1 (see Note 1).
-
2.
10% (v/v) bleach solution (1.2-1.4% sodium hypochlorite).
-
3.
Sterile distilled water.
2.1.2 In Vitro Cultivation of N. tabacum Plants
-
1.
Petri dishes containing standard ½ MS solid medium (see Note 2) or water-soaked filter papers (see Note 3).
2.1.3 Preparation of Agrobacterium Suspension
-
1.
A. tumefaciens strain, for example: GV3101[pPM6000; pTd33] grown on solid YEB medium containing rifampicin (100 mg/L) and gentamycin (20 mg/L).
-
2.
Liquid YEB medium (see Note 4) containing 100 mg/L rifampicin (Sigma) and 20 mg/L gentamycin (Bioshop).
-
3.
10 mM MgSO4 solution (filter sterilized).
-
4.
Liquid MS medium (see Note 3).
-
5.
Optional: 100 mM acetosyringone (3′,5′-Dimethoxy-4′-hydroxyacetophenone; Aldrich) in alcohol solvent (isopropanol or ethanol).
2.1.4 Transformation by Infiltration
-
1.
10-14-day-old tobacco seedlings or 1-2 month-old tobacco plants cultured in vitro (see Subheading 3.1.2).
-
2.
Agrobacterium suspension (see Subheading 3.1.3).
-
3.
MSH1 solid medium (for modified MS media composition see Note 5).
-
4.
MSH1T or MSH1C solid medium (see Note 5).
2.1.5 β-Glucouronidase Histochemical Assay
-
1.
β-glucouronidase (GUS) staining solution: 100 mM Na-phosphate buffer, pH 7.0; 0.05% X-Gluc (5-Bromo-4-chloro-3-indoxyl-beta-D-glucuronide, GBT) dissolved in DMF (dimethylformamide, Sigma); 0.1% (w/v) Na-azide.
-
2.
99.6% and 70% (v/v) ethanol.
2.1.6 Selection and Regeneration of Transformants
-
1.
MSH1TK or MSH1CK solid medium (see Note 5).
-
2.
MSH2 solid medium (see Note 5).
2.2 Materials for Dipping Arabidopsis Flowers
2.2.1 Cultivation of Arabidopsis thaliana plants in soil
-
1.
Arabidopsis seeds: A. thaliana ecotype Columbia (see Note 6).
-
2.
Standard horticultural soil.
-
3.
Artificial carrier for plant soil culture, e.g., Vermiculite.
-
4.
Dry sand.
2.2.2 Preparation of Agrobacterium Suspension
-
1.
The same materials are required as described in Subheading 2.1.3.
2.2.3 Transformation by Flower Dipping/Infiltration
-
1.
Flowering Arabidopsis plants (see Subheading 3.2.1).
-
2.
Agrobacterium suspension (see Subheading 3.2.2).
2.2.4 Selection of Putative Transformants Using a Kanamycin Resistance Marker
-
1.
70% (v/v) ethanol.
-
2.
10% (v/v) bleach solution (1.2-1.4% sodium hypochlorite) containing detergent, e.g., 0.05% (v/v) Tween 20.
-
3.
MS (or ½ MS) medium containing kanamycin (50 mg/L).
3 Methods
The Agrobacterium-mediated plant transformation method employs a variety of different techniques for infecting plants to permit T-DNA transfer. In this chapter, we present the most commonly used techniques: vacuum infiltration of explants and floral dipping. Of the two, only vacuum infiltration requires plant regeneration steps (Fig. 2). In both cases, however, the same strain of binary vector containing A. tumefaciens can be used.

A general scheme of Agrobacterium-mediated transformation of explants using vacuum infiltration technique
In general, two types of Agrobacterium vectors are commonly used, co-integrate vectors and binary vectors. In both types of vectors, Agrobacterium plasmids are disarmed by deleting naturally-occurring T-DNA encoded “oncogenes” and replacing them with a gene of interest. In the co-integrative vector system, a donor vector containing the gene of interest is integrated with the disarmed Ti plasmid. The binary vector system consists of two autonomously-replicating plasmids within Agrobacterium, a binary vector that contains gene(s) of interest between T-DNA borders and a helper Ti plasmid that provides vir gene products required for T-DNA transfer to plant cells. Binary vectors have been recently developed, and are most commonly used now because of the ease of DNA manipulation both in vivo and in vitro, and for their higher transformation efficiencies.
The Agrobacterium strain used in the protocols described in the present paper is called GV3101[pPM6000; pTd33] and is a binary vector system (Fig. 3). GV3101 is a non-virulent Agrobacterium strain, a derivative of the wild type C58 strain but lacking the Ti plasmid (C58 is a virulent strain containing the nopaline Ti plasmid) (12).

A binary system of the Agrobacterium GV3101[pPM6000; pTd33] strain (see text for details)
Both stains are characterized by their resistance to the rifampicin antibiotic. pPM6000 is a derivative of the octopine plasmid pTiAch5 lacking the T-DNA region (12), and it is used as the helper plasmid, whereas pTd33 is the binary vector (13). This binary vector contains several important sequences: (1) T-DNA with genes encoding β-glucuronidase (uidA = GUS gene) and kanamycin resistance (nptII = KanR gene) under the control of plant regulatory elements (CaMV 35S promoter and nos terminator), (2) two origins of replications allowing plasmid amplification in both Escherichia coli and Agrobacterium cells (ori pBR322 and ori Ri), and (3) a gentamicin resistance gene (GmR) for bacteria selection.
3.1 Agrobacterium-Mediated Transformation of Plant Explants (e.g., Tobacco Seedlings or Leaf Discs)
3.1.1 Sterilization of N. tabacum Seeds
-
1.
Open seed capsules of N. tabacum plants, and place the seeds in a sterile 50 ml Falcon tube.
-
2.
Fill the tube with bleach solution, and mix it for 10 min.
-
3.
Sediment the seeds by pouring the suspension through a sterile 50 μm sieve.
-
4.
Rinse the seeds 3-5 times with sterile distilled water.
-
5.
Place the sieve with sterilized seeds in a plastic Petri dish, and leave in a laminar flow cabinet till the seeds are dry (overnight).
-
6.
Close the Petri dish with Parafilm, and store the seeds at room temperature until use.
3.1.2 In Vitro Cultivation of N. tabacum Plants
-
1.
Sow ∼ 100 sterilized seeds uniformly onto the surface of ½ MS solid medium or moisten filter paper (see Subheading 2.1.2 and Note 3) in a 9 mm Petri dish by using a sterile scalpel or spatula.
-
2.
Close the plate with Parafilm, and place it at 4°C for 3-7 days for seed stratification (see Note 7).
-
3.
Transfer the plate to a plant in vitro culture cabinet under the following culture conditions: photoperiod - 16 h light/8 h dark (or 24 h light), temperature - 22-24°C, humidity - 60%.
-
4.
Cultivate the culture for 10-14 days to obtain seedlings.
-
5.
Optional: transfer 2-week-old seedlings into glass jars containing solid MS medium, and cultivate the plat cultures in vitro under the same conditions for additional 1-2 months to obtain young plants with fully developed leaves.
3.1.3 Preparation of Agrobacterium Suspension
-
1.
Inoculate 10.5 mL of liquid YEB medium containing 100 mg/L rifampicin, 20 mg/L gentamycin, and 0.5 mM acetosyringone (optional, see Note 8) with a single colony of the A. tumefaciens strain GV3101[pPM6000; pTd33].
-
2.
Incubate the culture on a rotary shaker at a speed of 120 rpm at 28°C for 24-48 h.
-
3.
Centrifuge the Agrobacterium culture at 5,500×g for 15 min. at 4°C.
-
4.
Discard the medium and resuspend the bacterial pellet in 35 mL of sterile 10 mM MgSO4.
-
5.
Repeat steps 3 and 4.
-
6.
Measure the suspension density (see Note 9).
-
7.
Centrifuge the Agrobacterium suspension at 5,500×g for 15 min at 4°C.
-
8.
Discard the supernatants and resuspend the bacterial pellet in liquid MS medium to bring the OD600 to 0.1 (see Note 10). Optional: add sterile acetosyringone (the final concentration: 0.5 mM).
3.1.4 Transformation by Infiltration
-
1.
If you use young tobacco plants (1-2 months old), cut out leaf discs (∼1 cm in diameter) under sterile conditions using a sterile cork-cutter.
-
2.
Place tobacco seedlings (optimum: 10-14-day-old) or leaf discs in tubes containing the Agrobacterium suspension or sterile MS medium (control).
-
3.
Infiltrate the plant material with the Agrobacterium suspension for 5 min (minimum pressure 130 mbar) in a sterile desiccator connected to a vacuum pump (see Note 11). Repeat the infiltration once more.
-
4.
Take out the infiltrated seedlings/leaf discs and dry them shortly (approximately for 1 min) on a sterile filter paper to remove excess of Agrobacterium cells.
-
5.
Transfer the infiltrated seedlings onto the surface of MSH1 medium and tightly seal the Petri dishes with Parafilm.
-
6.
Co-cultivate the plants with Agrobacterium for 3 days in a plant in vitro growth chamber to allow Agrobacterium to infect explants and transfer the T-DNA carrying transgene(s) to the plant tissue.
-
7.
After 3 days of co-cultivation, transfer the seedlings to MSH1T or MSH1C solid medium containing a bacteriostatic agent (see Note 12), and culture again for a week under the same conditions.
-
8.
Perform a test of transient expression of the GUS reporter gene, and/or continue with selection and regeneration.
3.1.5 β-Glucouronidase Histochemical Assay
-
1.
Transient or stable expression of GUS can be tested using this assay (14) (see Note 13).
-
2.
Place the plant material in 15 ml Falcon tubes, and soak it in the GUS staining solution.
-
3.
Infiltrate the plant material with the GUS staining solution for 10 min. using a desiccator connected to a vacuum pump (minimal pressure 130 mbar).
-
4.
Remove the tubes from desiccator, and incubate them for 2-3 days at 37°C; leave lids open to permit oxidation.
-
5.
Pour off the staining solution, and wash the plant material with distilled water.
-
6.
Wash out chlorophyll from plant tissues by incubating them for several hours in 99.6% ethanol; this can be accelerated by gentle shaking and warming. Exchange ethanol until plants are white.
-
7.
Replace 99.6% ethanol with 70% ethanol solution (this makes the plant material less brittle). These samples can be kept indefinitely.
-
8.
Place the plant material in a plastic Petri dish, and observe GUS staining with a binocular microscope. The presence of blue spots indicates GUS expression (Fig. 4).
Fig. 4. 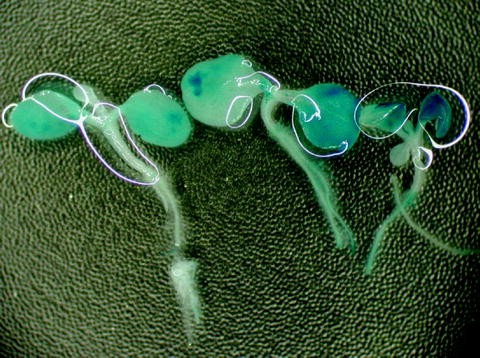
Transformed tobacco seedlings after the histochemical β-glucuronidase (GUS) assay

3.1.6 Selection and Regeneration of Transformants
-
1.
After a week (see Subheading 3.1.4, step 7), transfer the seedlings/leaf discs onto the surface of the selection medium containing kanamycin (MSH1TK or MSH1CK). Culture for 3 weeks under the same conditions.
-
2.
Transfer surviving tissue sectors on MSH1TK (or MSH1CK) medium for the next cycle of selection (see Note 14). Perform 3-4 cycles of selection. Green shoots will start to emerge from the tissue (see Note 15).
-
3.
Transfer regenerating shoots to MSH2 solid medium for root induction. If needed, add bacteriostatic antibiotic to MSH2 medium (timentin, carbenicillin or cefotaxime).
-
4.
The selected plants (survivals) are putative transgenic plants. For further procedures with putative transformants (see Notes 16 and 17).
3.2 Agrobacterium-Mediated Transformation of Female Reproductive Tissues of A. thaliana (Floral-Dip Method)
3.2.1 Cultivation of A. thaliana Plants in Soil
-
1.
Mix standard horticultural soil with an artificial carrier (volume ratio 3:1), sterilize the mixture by autoclaving, and place it in small plastic flower-pots.
-
2.
Put the pots on saucers (supports) filled with water, and leave them for 4-24 h.
-
3.
Mix A. thaliana seeds with small amount of dry sand, and sow them in the soil.
-
4.
Cover the pots with foil, and place them at 4°C for 3-7 days for seed stratification.
-
5.
Remove foil, and transfer the pots to a culture cabinet. Culture conditions: photoperiod - 16 h light/8 h darkness, temperature - 22-24°C, 60% - humidity.
-
6.
Culture the seeds/plants for 2-3 weeks until the plants reach the stage of a rosette with 2-4 leaves. Transfer the plants to individual pots (see Note 18).
-
7.
Continue cultivation for the next 1-2 weeks to reach the flowering stage.
3.2.2 Preparation of Agrobacterium Suspension
-
1.
Prepare the Agrobacterium suspension according to the protocol described in Subheading 3.1.3. The Agroabcterium suspension used for the floral dip method may have a higher density (OD600 = 0.6-1.0).
-
2.
Transfer the Agrobacterium suspension to a sterile glass beaker.
3.2.3 Transformation by Flower Dipping/Infiltration
-
1.
Put the pot with flowering Arabidopsis plants upside down, and place flower shoots into the Agrobacterium suspension (see Note 19) for up to 30 min. (dipping). Alternatively, dipping may be combined with vacuum infiltration (see Note 20).
-
2.
Remove the flower-pot containing the plant from the beaker, and place it on a support.
-
3.
Cover the flower-pot with a transparent plastic lid (see Note 21) to maintain humidity. Leave plants in a low light or dark location overnight. Keep the domed plant out of direct light.
-
4.
Remove the lid 12-24 h after treatment (infiltration), and transfer the inoculated plant to a culture cabinet. Culture conditions: photoperiod - 16 h light/8 h dark or 24 h light, temperature - 22-24°C, humidity - 60%.
-
5.
Optional: repeat dipping (or infiltration) of Arabidopsis flower shoots in the Agrobacterium suspension 2-3 times at 5-6 day intervals (see Note 22).
-
6.
Grow plants for a further 3-5 weeks period until siliques are brown and dry. Stop watering the plants at this stage.
-
7.
Harvest seeds by gentle pulling groups of inflorescences through your fingers over a piece of clean paper. Remove the majority of the stem and pod material.
-
8.
Store seeds in Eppendorf tubes either at room temperature or at 4ºC under desiccation.
3.2.4 Selection of Putative Transformants Using a Kanamycin Resistance Marker
-
1.
Sterilization of Arabidopsis seeds:
-
-Place a desired amount of seeds in an Eppendorf tube, and soak them for 1-2 s in 1 mL of 70% ethanol
-
-Under sterile conditions, remove ethanol, and soak seeds in 1 mL of 10 % bleach solution containing a detergent (e.g. 0.05% (v/v) Tween 20) and incubate for 5-10 min
-
-Remove bleach solution
-
-Rinse seeds 5 times with 1 mL of sterile distilled water (leave them in water after the last wash)
-
-
2.
Transfer the suspension of sterilized seeds onto the surface of solid ½ MS medium containing kanamycin (50 mg/L). Leave the plates open in a laminar flow cabinet for 1 h or until excess water is evaporated.
-
3.
Close the plates, and seal them with Parafilm. Place at 4ºC for 3-7 days for seed stratification.
-
4.
Place the plates in a plant in vitro culture cabinet/growth chamber. Culture conditions: photoperiod - 16 h light/8 h dark or 24 h light, temperature - 22-24°C, humidity - 60%.
-
5.
Grow the plants for 1-2 weeks. Transformants should be identified as kanamycin-resistant seedlings that produce green leaves and a well-established root system within the selective medium (see Note 23).
-
6.
Grow some putative transformants to maturity by transplanting into heavily moistened potting soil (preferably after the development of 3-5 adult leaves). For further procedures with putative transformants, see Notes 16 and 17.
4 Notes
-
1.
All tobacco species, including N. tabacum, Nicotiana bentamiana, and others as well as all cultivars within each species (e.g., SR1 and Samsun of N. tabacum) can be used.
-
2.
Standard MS medium (15) is prepared using pre-mixed macro- and micro-elements (MS basal salts; 4.33 g/L; Sigma), 2-3% (w/v) sucrose and MS vitamins (1,000× stock, Sigma). pH 5.7-5.8 should be adjusted using 1 N KOH. ½ MS medium contains half the amount of MS basal salts and unchanged amounts of other compounds. Solid medium contains 0.8% (w/v) agar (Sigma) in addition.
-
3.
2-3 layers of filter paper (e.g., Whatmann paper, Sigma-Aldrich) may be used instead of ½ MS solid medium.
-
4.
YEB medium contains 0.1% (w/v) yeast extract (Difco), 0.5% (w/v) peptone (Difco), 0.5% (w/v) beef extract (Bioshop), and 0.5% (w/v) sucrose. pH 7.2 should be adjusted using 1 N NaOH. After autoclaving, MgSO4 (1 M filter sterilized solution) should be added to achieve the final concentration of 2 mM.
-
5.
Modified MS media are prepared based on the standard MS medium (see Note 2) supplemented with plant hormones and/or antibiotics. MSH1 solid medium is MS solid medium supplemented with the following phytohormones: BAP (6-Benzylaminopurine, Sigma; the final concentration: 1 mg/L) and NAA (1-Naphtaleneacetic acid, Sigma; the final concentration: 0.1 mg/L). MSH1T solid medium is MSH1 solid medium containing 100 mg/L timentin (Ticarcillin disodium + Potassium clavulanate 15:1, Duchefa Biochemie), whereas MSH1C solid medium is MSH1 solid medium containing 300 mg/L carbenicillin (Bioshop) or 300-500 mg/L cefotaxime (Sigma). MSH1TK (or MSH1CK) solid medium is MSH1T (or MSH1C) solid medium containing 50-100 mg/L kanamycin (Sigma). MSH2 solid medium is MS solid medium containing 0.5-1.0 mg/L IBA (indole-3-butyric acid, Sigma).
-
6.
Different Arabidopsis ecotypes, including Columbia (Col-0), C24, Wassilewskija (WS-0), Landsberg (La-0, La-1), and Landsberg erecta (Ler-0), may be used. Seeds are available from The European Arabidopsis Stock Centre (NASC; http://arabidopsis.info) or Arabidopsis Biological Resource Center (ABRS; http://www.biosci.ohio-state.edu/∼plantbio/Facilities/abrc/abrchome.htm).
-
7.
Some seeds need a period of moisture and cold after harvest before they will germinate; usually this is necessary to either allow the embryo to mature or to break dormancy. This period can be artificially stimulated by placing the moistened seed (e.g., on moist medium) in a refrigerator for a certain period of time. Seed stratification is also used for those seeds that do not require stratification, as this treatment allows synchronization of germination.
-
8.
Liquid YEB medium and/or MS medium used to prepare the Agrobacterium suspension may be supplemented with acetosyringone (the final concentration: 0.5 mM), especially when intact plant tissues are used for transformation. In case of explant transformation (leaf discs, cotyledons), the addition of acetosyringone is not required, since wounding causes the production of plant compounds (including acetosyringone) that induce vir gene expression in Agrobacterium. However, when the problem with low transformation efficiency occurs, addition of acetosyringone to bacterial culture or bacterial suspension (or even both) is also recommended for explants transformation.
-
9.
OD600 of diluted bacterial suspension should be measured. Dilute bacterial suspension in 10 mM MgSO4 at the ratio of 1:2-1:5. OD readings should be in the range of 0.2-0.5. Higher values indicate that the bacterial culture was overgrown and thus contains too many old bacterial cells, which are not likely to be competent for DNA transfer. Lower values indicate that bacteria did not grow properly and will not be able to transfer DNA either.
-
10.
The Agrobacterium suspension with higher density may be used (OD600 up to 1.0). However, the higher density increases the probability of multi-copies and/or multi-loci T-DNA integration patterns.
-
11.
For releasing the air pressure after infiltration, place a desiccator in a laminar-flow hood and only then open the desiccator’s valve. This will prevent contamination of infiltrated plant material.
-
12.
Any bacteriostatic agent that kills Agrobacterium (or at least inhibits its growth) can be used, including timentin, carbenicillin, and cefotaxime.
-
13.
Transient expression of GUS can be monitored 0-3 days after transfer of seedlings/leaf discs to the MSH1 medium. For analysis of the stable expression of GUS, plant material from T1 or next generations of transgenic plants selected for kanamycin resistance may be used.
-
14.
The bacteriostatic agent may be omitted at any later round of selection if no growth of Agrobacterium on the surface of plant tissue (or medium) is observed.
-
15.
The first shoots will appear during the first and second round of selection. It is recommended to transfer both explants containing emerging shoots and those which do not contain shoots but are still not dead (completely brown) to new selection media every 3 weeks. In the first case, it will help to eliminate false-positive lines, whereas in the second case - it will allow the recovery of late-appearing transgenic lines.
-
16.
Putative transformants should be further tested for stable integration of transgenes by the PCR or Southern hybridization method (16, 17).
-
17.
To obtain a homozygous transgenic line, perform self-crosses of primary transgenic plants. The purity of the homozygous line can be confirmed in back-crosses with a wild type non-transgenic line (18).
-
18.
The soil in the individual pots should be covered with a net to prevent loss of soil during dipping of Arabidopsis flowers in Agrobacterium suspension. As an alternative to a net, you may cover Arabidopsis leave rosettes with gauze (fixed to a pot with a rubber ring) at the time of the first flower shoot appearance.
-
19.
Remove any siliques that may have already developed. Infiltration of premature (not fully developed) flowers is recommended. Agrobacterium must be delivered to the interior of the developing gynoecium prior to locule closure if efficient transformation is to be achieved, as ovules were shown to be the site of productive transformation in the floral-dip method (19).
-
20.
Place the beaker with flower shoots dipped in the Agrobacterium suspension solution in a desiccator connected to a vacuum pump and infiltrate the plant material for 5 min (minimal pressure 130 mbar). Repeat infiltration once more (optional).
-
21.
Instead of using a commercial flower-pot lid, you may form a dome from a plastic bag or saran-wrap.
-
22.
Repeating the dipping (or infiltration) is strongly recommended, as it increases the number of transgenic seeds obtained as a result of using the floral dip transformation method, mainly due to the higher number of treated flowers. While repeating the dipping/infiltration, do not remove siliques developed after previous dipping/infiltration, since they most likely contain developing transgenic seeds.
-
23.
In case of a high number of false-positive plants, increase the strength of selection by increasing the concentration of the selective agent (for kanamycin: 50-250 mg/L).
References
Birch RG (1997) Plant transformation: problems and strategies for practical application. Annu Rev Plant Physiol Plant Mol Biol 48:297-326
Chawla HS (2002) Gene transfer in plants. In: Introduction to plant biotechnology, 2nd edn. Science Publishers Inc., Enfield, NH, USA, pp 359-395
Slater A, Scott NW, Fowler MR (2008) Techniques for plant transformation. In: Plant biotechnology: the genetic manipulation of plants, 2nd edn. Oxford University Press, New York, USA pp 54-74
Tzfira T, Citovsky V (2008) Agrobacterium: from biology to biotechnology. Springer, NY
Matzke AJM, Matzke MA (1998) Position effects and epigenetic silencing of plant transgenes. Curr Opin Plant Biol 1:142-148
Matzke MA, Matzke AJM (1998) Epigenetic silencing of plant transgenes as a consequence of diverse cellular defense responses. Cell Mol Life Sci 54:94-103
Kaeppler SM, Kaeppler H, Rhee Y (2000) Epigenetic aspects of somaclonal variation in plants. Plant Mol Biol 43:179-188
Filipecki M, Malepszy S (2006) Unintended consequences of plant transformation: a molecular insight. J Appl Genet 47:277-286
Henderson IR, Jacobsen SE (2007) Epigenetic inheritance in plants. Nature 447:418-424
Gendrel A-V, Colot V (2005) Arabidopsis epigenetics: when RNA meets chromatin. Curr Opin Plant Biol 8:142-147
Madlund A, Comai L (2004) The effect of stress on genome regulation and structure. Ann Bot 94:481-495
Bonnard G, Tinland B, Paulus F, Szegedi E, Otten L (1989) Nucleotide sequence, evolutionary origin and biological role of a rearranged cytokinin gene isolated from a wide host range biotype III Agrobacterium strain. Mol Gen Genet 216:428-438
Tinland B, Schoumacher F, Gloeckler V, Bravo-Angel AM, Hohn B (1995) The Agrobacterium tumefaciens virulence D2 protein is responsible for precise integration of T-DNA into the plant genome. EMBO J 14:3585-3595
Rossi L, Escudero J, Hohn B, Tinland B (1993) Efficient and sensitive assay for T-DNA-dependent transient gene expression. Plant Mol Biol Rep 11:220-229
Murashige T, Skoog F (1962) A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol Plant 15:473
Chawla HS (2003) In vitro amplification of DNA by PCR: detection of transgenes. In: Plant biotechnology: a practical approach. Science Publishers Inc., Enfield, NH, USA, pp 215-222
Chawla HS (2003) Sothern blotting, Southern hybridization. In: Plant biotechnology: a practical approach. Science Publishers Inc., Enfield, NH, USA, pp 237-252
Koornneef M, Alonso-Blanco C, Stam P (2006) Genetic analysis. In: Salinas J, Sanchez-Serrano JJ (eds) Arabidopsis protocols, 2nd edn. Humana Press, Totowa, NJ, pp 65-77
Desfeux C, Clough SJ, Bent AF (2000) Female reproductive tissues are primary target of Agrobacterium-mediated transformation by the Arabidopsis floral-dip method. Plant Physiol 123:895-904
Acknowledgments
The author would like to thank Prof. Barbara Hohn and her co-workers for establishing the protocol for Agrobacterium-mediated transformation of tobacco seedlings, Dr. Igor Kovalchuk for his advice and encouragement, and Dr. Valentina Titova for language revision.
Author information
Authors and Affiliations
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2010 Springer Science+Business Media, LLC
About this protocol
Cite this protocol
Ziemienowicz, A. (2010). Plant Transgenesis. In: Kovalchuk, I., Zemp, F. (eds) Plant Epigenetics. Methods in Molecular Biology™, vol 631. Humana Press. https://doi.org/10.1007/978-1-60761-646-7_19
Download citation
DOI: https://doi.org/10.1007/978-1-60761-646-7_19
Published:
Publisher Name: Humana Press
Print ISBN: 978-1-60761-645-0
Online ISBN: 978-1-60761-646-7
eBook Packages: Springer Protocols