
Abstract
This study examined whether encoding conditions that encourage thoughts about the environment of evolutionary adaptation (EEA) are necessary to produce optimal recall in the adaptive memory paradigm. Participants were asked to judge a list of words for their relevance to personal survival under two survival-based scenarios. In one condition, the EEA-relevant context was specified (i.e., you are trying to survive on the savannah/grasslands). In the other condition, no context was specified (i.e., you are simply trying to stay alive). The two tasks produced virtually identical recall despite participants in the former condition reporting significantly more EEA context-relevant thoughts (i.e., the savannah) than did participants in the latter condition (who reported virtually no EEA-related thoughts). The findings are discussed in terms of (1) survival as a target of natural selection and (2) the role of evolutionary theory in understanding memory in modern humans.
Similar content being viewed by others


Scientists adopting an evolutionary perspective on neural systems take, as a starting assumption, that systems exist in their present arrangement because that form solved certain recurrent problems the organism faced in its evolutionary history. Researchers attempt to use knowledge of those problems to reverse-engineer the design of the system or systems of interest (e.g., Barkow, Cosmides, & Tooby, 1992: Dawkins, 1976; Glenberg, 1997; Klein, Cosmides, Tooby, & Chance, 2002; Nairne, 2010; Sherry & Schacter, 1987; Suddendorf & Corballis, 1997; Williams, 1966).
From this perspective, memory systems can be viewed as the result of the complex interplay of a set of processes that enable an organism to draw on past experiences to guide current behavior and plan for future contingencies (for a discussion, see Adam, 2004; Bradley, 1887; Ingvar, 1985; Klein, 2007; Klein, in press-a; Klein et al., 2009; Klein et al., 2002; Klein, Robertson, & Delton, 2010; Suddendorf & Corballis, 1997). To the extent that these abilities affect the organism’s reproductive potential, they are acted on by natural selection.
In a recent series of studies adopting an evolutionary perspective, Nairne and his colleagues have examined the memorial advantages conferred by processing stimuli in relation to their relevance to personal survival (e.g., Nairne & Pandeirada, 2008a, 2008b, 2010; Nairne, Pandeirada, Gregory, & Van Arsdall, 2009; Nairne, Padeirada, & Thompson, 2008; Nairne, Thompson, & Pandeirada, 2007; Nairne, Van Arsdale, Pandierada, & Blunt, 2012). They repeatedly have demonstrated that encoding tasks that encourage thoughts of personal survival are associated with memorial benefits not found with tasks that do not encourage such thoughts. These findings have been widely replicated (e.g., Burns, Burns, & Hwang, 2011; Hart & Burns, 2012; Kang, McDermott, & Cohen, 2008; Otgaar, Smeets, & van Bergen, 2010; Soderstrom & McCabe, 2011; Weinstein, Bugg, & Roediger, 2008; however, see Butler, Kang, & Roediger, 2009; Klein et al., 2010; Klein, Robertson, & Delton, 2011; Klein, Robertson, Delton, & Lax, 2012).
As conceptualized by Nairne (e.g., 2005, 2010), the memorial advantage of survival processing can be explained by appeal to evolutionary considerations. According to this view, there are obvious fitness benefits from a memory system designed to take past events and extract information from them that increases one’s ability to survive long enough to enable transmission of his or her genetic endowment. It thus makes sense to consider the possibility that during the long period that natural selection “designed” hominid neural machinery, memory systems that evolved to enhance their owner’s ability to survive long enough to reproduce and care for offspring would exhibit a selective advantage.
Nairne and colleagues have suggested as well that memory mechanisms may work optimally when current encoding conditions prime or reinstate ancestrally-based problems, particularly those present in ancestral foraging environments (see Nairne & Pandeirada, 2010, p. 2). Consistent with this view, Nairne and colleagues repeatedly have demonstrated that memory is best when encoding not only primes thoughts of survival, but also primes thoughts of survival on the savannah (or African grasslands; i.e., the assumed environment of evolutionary adaptation [henceforth, EEA]; see also Butler et al., 2009; Kang et al., 2008; Kroneisen & Erdfelder, 2011; Weinstein et al., 2008). By contrast, thoughts of personal survival absent consideration of ancestral environment (e.g., survival in a modern city),Footnote 1 although sometimes associated with memory exceeding that of tasks that do not entail considerations of survival (e.g., rating words for pleasantness), typically fall short of memory for survival scenarios placed in the context of the EEA (e.g., Nairne & Pandeirada, 2010; Soderstrom & McCabe, 2011).
There are, however, a few studies that appear at odds with the idea that encoding conditions that encourage EEA-relevant thoughts necessarily promote optimal performance. For example, Klein and colleagues (Klein, in press-b; Klein et al., 2010, 2011; Klein et al., 2012) have argued that the mechanisms that mediate the excellent memory found with survival tasks used in the adaptive memory paradigm have more to do with factors such as planning and self-reference than with either survival context or even survival per se (for a review, see Klein, in press-a). For example, tasks that encouraged participants to plan for future contingencies but did not reference either the EEA or thoughts of survival (e.g., planning a birthday party) produced recall statistically indistinguishable from tasks promoting thoughts of survival on the savannah.
A finding by Soderstrom and McCabe (2011) also calls into question the causal potency of EEA priming. These investigators showed the type of predation (i.e., predation by zombies vs. unspecified predation) superseded considerations of encoding context: Regardless of whether context encouraged thoughts of a modern city or of ancestral grasslands, memory was better when the predator was a zombie. Two comments about this study are in order.
First, Soderstrom and McCabe (2011) did replicate Nairne’s finding that encoding conditions encouraging thoughts of personal survival lead to better retention than do conditions that do not promote thoughts of survival. Second, and more relevant to the present study, while the nature of predation had an independent effect on memory (i.e., memory was better when predators were identified as zombies than when the nature of predation was unspecified, regardless of whether encoding context was modern or ancestral), it is not clear how to interpret this finding. The superior performance associated with zombie-based predation may be attributable to the fact that for approximately 100 years, the figure of the “zombie” has maintained a very strong presence in popular culture and media. For example, zombies have been a favorite topic of horror movies, beginning with portrayals by Bela Lugosi in the 1930s and 1940s. Indeed, Wikipedia lists more than 300 films featuring zombies as central characters, at least 200 of which have appeared during the past 10 years! Today the character of the “zombie” predator is deeply embedded in our collective consciousness.
Thus, in contrast to studies adopting modern survival scenarios featuring vaguely specified predation (i.e., nonspecific, city-based attackers; e.g., Nairne & Pandeirada, 2010; Weinstein et al., 2008), zombie-based predation has a very strong presence in popular culture and public awareness. As Klein and colleagues have demonstrated (e.g., Klein et al., 2010, 2011; Klein et al., 2012), personal familiarity with the context of encoding (whether ancestral or modern) makes an important, independent contribution to memory performance found in the adaptive memory paradigm (for a review, see Klein, in press-a). Adoption of the highly popular, and thus highly familiar, zombie predator leaves open the possibility that the reason zombie-based predation has memorial effects above and beyond those of temporal placement (i.e., ancestral vs. modern) may be attributable more to inclusion of this very familiar cultural figure than to any memorial impotency associated with priming thoughts of the EEA.
Several studies (e.g., Nairne & Pandeirada, 2011; Weinstein et al., 2008) have found that when survival scenarios are located in the modern times, memory is inferior to that found when the scenario is located in the EEA. While this might be taken, prima facie, as support for the memorial potency of the ancestral context, a critique similar to that addressed to the findings of Soderstrom and McCabes (2011) is applicable. Specifically, rather than simply eliminating thoughts of the EEA, these studies introduced novel and relatively unfamiliar factors into the modern survival scenario (e.g., fending off an unspecified assailant in a foreign land). In fairness, these studies were not focally concerned with demonstrating the idea that priming thoughts of the EEA are a necessary factor in producing optimal recall; rather, they primarily were interested in exploring the boundaries and mechanisms of the adaptive memory effect.
However, if one is interested in whether thoughts of the EEA constitute a necessary condition for obtaining optimal memory performance in the adaptive memory paradigm, a more convincing way of demonstrating its relevance is not by adding variables (e.g., types of predation) but, rather, by subtraction. Accordingly, work that, on the surface, appears consistent with the idea that reinstatement of EEA at encoding is not required to produce optimal retention (e.g., Klein et al., 2010, 2011; Nairne & Pandeirada, 2011; Soderstrom & McCabe, 2011) suffer from interpretive ambiguity. This is because they introduce a number of variables (e.g., use of different stimulus items [e.g., Hart & Burns, 2012; Klein et al., 2012]; alterations in the nature of predation [e.g., Soderstrom & McCabe, 2011; Weinstein et al., 2008]) that have the unintended effect of overdetermining the number of potentially causally relevant factors mediating memory performance, thereby compromising conclusions about the memorial potency of the EEA per se.
The present study
In the following study, I take the approach of stripping away, rather than adding to or altering, factors in an attempt to empirically discern the importance of reinstatement of ancestral context as a condition for optimal performance in the adaptive memory paradigm. Specifically, I compared four encoding tasks: Nairne’s and colleagues’ version of survival on the EEA, a survival task that did not specify context (either spatial or temporal), a mate selection task, and a word pleasantness judgment task.
With respect to the survival scenario pioneered by Nairne and colleagues (for discussions, see Nairne, 2005, 2010), the task I selected used the exact wording of the scenario employed by Nairne et al. (2007; also used by Butler et al., 2009; Klein, in press-b; Nairne & Pandeirada, 2008a, 2010; Weinstein et al., 2008, and many others). This task makes explicit mention of EEA context (i.e., the savannah; although in some versions, for purpose of explication, the term grasslands is substituted for savannah).
The temporally and spatially nonspecific survival task consisted in asking participants to imagine the relevance of a list of words to the task of staying alive. No context was mentioned. The mate selection task was chosen because of its clear fitness relevance (e.g., Williams, 1966), despite the fact that it makes no explicit mention of survival (indeed, some evolutionary theorists have argued that our brains evolved not as survival machines, but as courtship machines; e.g., Barkow et al., 1992; Miller, 2000). Finally, judging list words for pleasantness has served as a useful control task in a large number of adaptive memory studies (e.g., Kang et al., 2008; Nairne & Pandeirada, 2008a; Nairne et al., 2007) and, thus, has provided a base from which to assess the relative memorial potencies of the first three tasks.
My predictions, based on the findings from previous studies, were that Nairne’s EEA-specific version of the survival task would produce high levels of recall. I also predicted that if reinstatement of the EEA is not necessary to obtain optimal performance in the survival processing task (as suggested, albeit inconclusively, in work by Klein et al., 2010, 2011; Klein et al., 2012; Soderstrom & McCabe, 2011), then the survival task that lacked explicit mention of temporal or spatial context (and thus should not evoke thoughts of the EEA; evidence for this assertion is presented below) should yield recall approximately equivalent to the EEA-specific version of the task.
With regard to the mate choice scenario, I predicted that despite its obvious survival relevance, it would not equal the two survival-based scenarios in recall productivity. This is because planning plays a central role in survival-based tasks typically employed in the adaptive memory paradigm (for a review, see Klein, in press-a; for “potentially” conflicting findings, see Nairne et al., 2012). By contrast, mate choice, while likely to entail some planning, has far fewer requirements in that regard (evidence in support of this will be presented below in the section titled “Pretesting”). Finally, word pleasantness judgments have served well as “anchor” control tasks in the adaptive memory paradigm, and I fully anticipated that they would do so here.
Pretesting
Prior to conducting the main study, I felt it useful to pretest the materials being used. The predictions above are based on several assumptions, and it is advisable to determine whether evidence can be found in their support. Accordingly, 56 participants (who were not used in the main study) were recruited from an introductory psychology course to take part in the pretest phase.
All participants received a booklet consisting of three pages. Page 1 contained task instructions. Page 2 contained 30 stimulus words along with a rating scale that ranged from 1 = very irrelevant to 5 = very relevant (to ensure task-rating coherence, the scale values in the pleasantness rating condition were changed to 1 = very unpleasant to 5 = very pleasant). All participants viewed the same list of 30 target words, selected from Nairne et al. (2007, Experiments 1 and 2). These also were the words used in the main study. Half the participants in each experimental condition were randomly assigned to receive one of two random orderings of the list words. On the final page, participants were asked to answer a set of questions (described below).
The 56 participants were randomly divided among the four experimental groups. Instructions for participants in the survival with context group read the following:
In this task, I would like you to imagine you are stranded on the grasslands in a foreign land, without any basic survival material. Over the next few months, you’ll need to find steady supply of food and water and protect yourself from predators. I am going to show you (on the next page) a list of words, and I would like you to rate how relevant each of these words would be for you in this survival situation. Some words may be relevant and others may not—it is up to you to decide.
The survival without context group received instructions that highlighted survival, but removed all mention of temporal or geographical context:
In this task, I want you to imagine that you are trying to stay alive. I am going to show you (on the next page) a list of words, and I would like you to rate how relevant each of these words might be to your staying alive. Some words may be relevant and others may not—it is up to you to decide.
The mate choice group was told the following:
In this task, I want you to imagine that you are trying to select a mate. I am going to show you (on the next page) a list of words, and I would like you to rate the relevance of each word to mate selection. Some of the words may be relevant and others may not—it’s up to you to decide.
Finally, the pleasantness judgment group received the following instructions:
In this task, I am going to show you (on the next page) a list of words, and I would like you to rate the pleasantness of each word. Some of the words may be pleasant and others may not—it’s up to you to decide.
Participants were allotted 30 s to read task instructions. They then were instructed to turn the page and were given 3 min to rate the 30 stimulus words in accordance with their respective task requirements (previous research has shown that this interval is sufficient to enable participants to complete their respective tasks at a comfortable pace). Finally, participants were asked to turn to page 3 of their booklet and answer three questions. All ratings were made on a 4-point scale accompanied by verbal designators: 1 = not at all, 2 = somewhat, 3 = quite a bit, and 4 = always. One minute was allotted to complete the questions. The questions, randomly varied among participants within each encoding condition, were the following:
-
1.
On the Rating Scale provided below, please rate the extent to which thoughts of survival played a part in your ratings of the items during performance of the item rating task
-
2.
On the Rating Scale provided below, please rate the extent to which thoughts of planning played a part in your ratings of the items during performance of the item rating task.
-
3.
On the Rating Scale provided below, please rate the extent to which thoughts of the savannah or grasslands played a part in your ratings of the items during performance of the item rating task.
The mean ratings for thoughts of survival during task performance were 3.71, 3.14, 1.57, and 1.36, for survival with context, survival without context, mate choice, and pleasantness judgments, respectively. A one-way analysis of variance (ANOVA) on these means was significant, F(3, 55) = 33.60, p < .001, MSE = .57, η 2 = .65, with Tukey tests (p < .05) showing that both survival tasks (i.e., with or without mention of EEA context) were associated with significantly more survival-relevant thoughts than was either the mate choice or the pleasantness judgment task. Thoughts of survival did not differ reliably between the two survival tasks or between the mate choice and pleasantness judgment tasks.
Turning to thoughts of planning, consistent with expectations (see the discussion above), participants in both survival groups (i.e., survival with context and survival without context) reported frequent thoughts of planning during task performance. By contrast, participants in the pleasantness judgment condition reported virtually no planning thoughts. Mate choice fell between these extremes (M = 3.14, 3.00, 2.21, and 1.14 for survival with context, survival without context, mate choice, and pleasantness judgments, respectively). A one-way ANOVA on these means was reliable, F(3, 55) = 28.12, p < .001, MSE = .42, η 2 = .61. As can be seen, thoughts of planning were virtually identical in the two survival conditions.Footnote 2 Thoughts of planning in the mating choice condition were statistically less frequent than were those found for the two survival tasks but exceeded thoughts of planning associated with pleasantness judgments. These observations were confirmed by a Tukey analysis (p < .05).
Finally, thoughts of savannah/grasslands were more frequent in the survival with context condition than in any of the other encoding conditions. Specifically, means of 2.00, 1.14, 1.07, and 1.14 were obtained for survival with context, survival without context, mate choice, and pleasantness judgments, respectively. Submitting these data to a one-way ANOVA produced a reliable effect, F(3, 55) = 6.89, p < .01, MSE = .39, η 2 = .29. Tukey tests (p < .05) revealed that, as was expected, thoughts of the savannah/grasslands in the survival with context condition reliably exceeded these thoughts in any of the remaining three conditions (none of which differed reliably, all clustered around the scale value not at all).
It is worth noting that, in absolute numbers, the amount of savannah-relevant thought in the survival with context condition is not high (i.e., around the scale midpoint). This was anticipated on the basis of Klein and colleagues’ (e.g., Klein, in press-b; Klein et al., 2011; Klein et al., 2012) demonstrations that participants in this condition often imagine the “savannah” in terms of more familiar scenarios, such as meadows, forests, and plains (for a discussion, see Klein, in press-a).
Main study
Method
Participants
One hundred four undergraduates in an introductory psychology class participated in the study. The experiment was conducted in small sessions averaging between 12 and 18 participants. Participants in each session were randomly assigned to experimental conditions. Each session lasted approximately 25 min.
Materials
Participants were randomly assigned to the survival with context, survival without context, mate choice, or pleasantness judgment condition (n = 26 per condition). All participants viewed the same list of 30 target words selected from the list used by Nairne et al. (2007, Experiments 1 and 2). Half the participants in each experimental condition were randomly assigned to receive one of two random orderings of the list words. This list was identical to that used during pretesting.
Design and procedure
At the start of the study, each participant received a seven-page booklet containing the experimental material appropriate to his or her condition. Participants were instructed to remain on the page on which they currently were working and not to turn from that page until explicitly requested to do so (the word STOP was prominently displayed at the bottom-center of each booklet page). To ensure compliance, four research assistants monitored participant performance during each testing session. There were no reports of participants who failed to comply with the “stop” instructions.
The first page of the booklet was left blank, and participants were instructed not to open the booklet until so requested. On receiving instructions, participants turned to the second page of the booklet, which contained instructions describing one of the four encoding tasks used in the study. The exact wording of these conditions has already been presented in the Pretesting section of this article and will not be repeated here. Participants were allowed 30 s to read the instructions from their respective conditions. They then were asked to turn to the third page in their booklet, which presented the 30 target words, 1 per line. Participants were asked to judge each word in accordance with their instructions from the previous page. Each word was accompanied by a 5-point scale relevant to their task (i.e., 1 = very irrelevant or 5 = very relevant for the two survival and the mate selection groups; 1 = very unpleasant to 5 = very pleasant for the pleasantness judgment group). Three minutes were allotted to complete this portion of the study (pretesting indicated that this interval was sufficient for participants to read instructions and make ratings at a comfortable pace; see Klein et al., 2010, 2011). Participants were instructed not to review their ratings once they had completed work. (The monitors identified 3 participants who failed to comply with this instruction These participants were removed from the study and replaced with 3 additional participants selected from the same undergraduate pool.)
Following the rating task, participants were instructed to turn to page 4, which asked them to “Please use the scale below to indicate how hard you found the rating task. Circle the most appropriate scale value.” The scale ranged from 1 (very hard) to 5 (very easy). Twenty seconds were provided to perform this rating. The next page contained a series of anagram completions and served as a 3-min distractor task. Participants were asked to complete as many anagrams as they could in the time provided.
Participants then were asked to continue to page 6, which contained instructions for the recall portion of the study. The instructions read as follows: “I now would like you to try to recall the words you rated in the first part of the study. Please write the words, one per line, in the spaces provided below. You may recall the words in any order they come to mind.” The page contained 30 blank lines. Five minutes were provided for recall.
On completion of recall, participants were instructed to turn to the final page of the booklet. This page contained questions designed as checks on the extent to which participants had survival- and savannah-relevant thoughts when rating stimulus items. The questions and rating scales were identical to those described in the Pretesting section of this article. One minute was allowed to complete the ratings, after which all the booklets were collected, and participants were debriefed.
Results
Recall
Out of the hundreds of items remembered, only 7 were extra-list intrusions. They were approximately evenly distributed among treatment conditions (i.e., 2, 2, 2, and 1 for the survival with context, survival without context, mate choice, and pleasantness judgment conditions, respectively). Errors were not counted as part of a participant’s recall score.
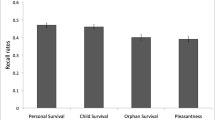
A one-way ANOVA conducted on the mean number of words correctly recalled yielded a reliable effect of task, F(3, 103) = 8.92, p < .001, MSE = 10.16, η 2 = .211. Tukey testing (p < .05) revealed that participants in both survival conditions recalled more words (M = 14.46 and 14.35 for the survival with context and survival without context groups, respectively) than did participants in either the mate choice or the pleasantness judgment group (M = 11.23 and 11.11, respectively). No reliable differences in recall were found between the two survival groups or between the mate choice and pleasantness groups.
Rating difficulty
I also examined whether participants found some rating tasks easier to perform than others. Although differences in ease of rating potentially could influence recall, it is hard to predict whether such an effect necessarily would be beneficial or harmful (e.g., Brown, 1976; Craik & Tulving, 1975).
The mean ease of rating scores were 4.04, 3.58, 4.23, and 4.40 for the survival with context, survival without context, mate choice, and pleasantness judgment groups, respectively. A one-way ANOVA performed on these scores failed to yield a significant effect of condition, F(3, 103) = 2.61, p > .05, MSE = .77, η 2 = .08: Participants in all four conditions found their tasks relatively easy to perform (the means ranged between 4.40 and 3.58 on a 5-point scale).
Thoughts of survival
Thoughts of survival during performance of the item-rating task yielded means of 3.66, 3.46, 1.50, and 1.38 for the survival with context, survival without context, mate choice, and pleasantness groups, respectively. A one-way ANOVA performed on these means yielded a reliable effect, F(3, 103) = 82.98, p < .001, MSE = .47, η 2 = .65. Not surprisingly, both survival tasks reliably encouraged more thoughts of survival than did either mate choice or pleasantness judgments. Furthermore, replicating the pretest findings, the two survival tasks did not differ in the high number of personal survival thoughts they produced. By contrast, the mate choice and pleasantness judgment tasks showed statistically comparable, low levels of survival-related thought. All observations were confirmed by Tukey testing (p < .05).
Thoughts of the savannah
Replicating the results of pretesting, the survival with context condition produced a greater number of thoughts of the EEA than did the survival without context, mate choice, or pleasantness judgment conditions (M = 2.23, 1.12, 1.04, and 104, respectively). These observations were confirmed by a one-way ANOVA, F(3, 103) = 38.29, p < .001, MSE = .23, η 2 = .52, with Tukey testing (p < .05) revealing that survival with context promoted reliably more thoughts of the savannah/grasslands than did any of the other encoding conditions (all three of which, as was found during pretesting, failed to differ reliably, hovering around the scale value not at all).
Study 1a
While the results of the first study offer support for the contention that survival processing does not require either imagination of the savannah or escape from predation to yield optimal recall performance, I felt it prudent to replicate this finding.
In addition, there is a potentially important design difference between the ancestral and modern survival experiments that have shown a memory advantage associated with ancestral context (e.g., Nairne & Pandeirada, 2010; Weinstein et al., 2008) and those that have not (e.g., Soderstrom & McCabe, 2011). In the former experiments, the survival scenario always was compared with a control condition within the same subject. For example, one group would receive both the ancestral scenario and pleasantness ratings; the other group would get a modern survival scenario and pleasantness ratings. The point is that in studies demonstrating the mnemonic superiority of ancestral survival processing, the ancestral/modern comparison is between subjects, but the survival/control comparison always is done within subjects. In my first study (as well as in Soderstrom & McCabe, 2011), all conditions were between subjects.
While a theoretical basis for assuming that these methodological differences matter is not readily apparent, the data nonetheless indicate that these design features may be critical to obtaining an ancestral survival memory advantage. Accordingly, I chose to replicate my study using two groups (as per Nairne & Pandeirada, 2010): survival (with context) and pleasantness ratings versus survival (without context) and pleasantness ratings.
Method
Participants
Fifty-six undergraduates in an introductory psychology class participated in the study. To better ensure compliance with instructions, testing was limited to 4 participants per session. Two investigators were present in each session—one conducting testing and blind to participant condition, and one serving as a proctor to monitor compliance. No participants failed to act in accordance with instructions. Participants in each session were randomly assigned to experimental conditions. Each session lasted approximately 30 min.
Materials and design
Participants randomly were assigned to either the survival with context + pleasantness judgment or the survival without context + pleasantness judgment condition (n = 28 per condition). All participants viewed the same list of 32 target words: The words were the same 30 as those used in the first study plus 2 additional words randomly selected from the list presented in Nairne et al. (2007, Experiment 3). All participants rated and recalled the same list of 32 words presented in the same order.
Within a session, participants rated 16 words for survival relevance (S) and 16 for pleasantness (P). Rating condition was blocked in trials of 8 words in the form SPSP or PSPS; half the participants received each version to ensure that each word was rated equally under both judgment tasks. The content of the survival scenario was manipulated between subjects: One group received survival with context instructions, and the other received survival without context instructions. Both groups received the same pleasantness judgment instructions.
Procedure
The procedure was identical to that of Study 1, with the following changes: (1) Participants made their ratings on the first four pages of the booklet, each page containing instructions for assigned task (SPSP or PSPS) accompanied by eight words to be rated (45 s per page) in accordance with task instructions; (2) data were not collected on either “rating ease” or “thoughts during task performance” (e.g., of survival or of savannah), since the interleaving of tasks would have made it difficult for participants to provide a “pure” retrospective assessment of these variables; (3) since the mating task was not germane to the primary goal of the study, it was not tested.
Results and discussion
Participants produced nine extra-list intrusions. They were approximately evenly distributed among treatment conditions (four for the survival with context/pleasantness condition and five for the survival without context/pleasantness condition). Errors were not counted as part of a participant’s recall score.
A two-way mixed ANOVA on the recall data, with survival (with and without context) as the between-subjects variable and pleasantness as the within-subjects variable, yielded only a reliable effect of rating task, F(1, 54) = 34.98, p < .001, MSE = 5.14, η 2 = .557: The survival with context (M = 8.96) and survival without context (M = 9. 46) tasks produced significantly better recall than did the pleasantness judgment task (M = 6.46 and 7.59 for the pleasantness tasks paired with survival with context and survival without context groups, respectively). No other effects or interactions approached significance (all ps > .30).
This study replicates the findings of the first study using a design that better approximates the one used in previous studies finding that survival processing advantages are attenuated when the ancestral properties of the task are removed. In contrast to those studies, the present study failed to obtain any evidence suggesting that reinstating the EEA setting improves recall, relative to a survival without context task. It also is important to note that in both this and the previous study, the survival without context task did not include any reference to predation.
General Discussion
This set of studies is the first (that I am aware of) to demonstrate, in a relatively unambiguous manner (e.g., by using the same instructions, stimuli, etc. as those used by Nairne and colleagues, and without introducing potentially confounding variables such as zombies, inner-city assassins, or space travel),Footnote 3 that reinstatement of ancestral conditions at encoding is not necessary to produce optimal recall with survival processing. In addition to analysis of recall performance, this conclusion receives support from examination of such factors as rating difficulty, thoughts of planning, and thoughts of savannah/grasslands. In short, both the survival with context and the survival without context conditions produced virtually identical, high levels of retention. This equivalence was obtained despite the fact that the survival without context condition failed to specify a spatial or temporal context (multiple testing confirmed that participants in this condition reported few, if any, EEA-relevant thoughts while making survival judgments). In fact, on all dependent measures, save one, the two survival tasks performed identically. The one exception was the mean number of “thoughts of savannah/grasslands”: The survival with context task promoted more of these thoughts that did the survival without context task. This was both anticipated (on the basis of task design) and consistent with the hypothesis that thoughts relevant to the EEA are not required to produce optimal recall performance, at least not consciously. (Of course, one cannot rule out the possibility that such thoughts take place unconsciously. However, unconscious thought, while a possibility, if demonstrated to be a causally relevant factor would create interpretive havoc with both present and previous conclusions about a number of measures employed in adaptive memory studies—such as rating difficulty, item relevance, etc.—all of which rely on participants’ introspective judgments. I leave this as an open question.)
As an additional perspective on the recall equivalence of the two survival tasks, I compared participant ratings of the relevance of stimulus words to performance of their assigned scenario. As Nairne et al. (2007) pointed out, such data are important because memory can be influenced by congruity effects. For example, Craik and Tulving (1975) and others (e.g., Hyde & Jenkins, 1973; Rogers, Kuiper, & Kirker, 1977; for a discussion, see Nairne & Pandeirada, 2011) have shown that participants are more likely to remember items rated “yes” than items rated “no” during performance of an encoding task. If the mean relevance ratings in the survival without context condition exceeded those in the survival with context group, memory performance of the former could be attributed, at least in part, to a greater congruence between target–task relations, thereby compromising any conclusions about the effect of reinstatement of the EEA on recall performance. Fortunately, this was not the case: A test of the mean relevance ratings for the survival with context (M = 3.02) and survival without context (M = 3.28) tasks in Study 1 was not significant, t(50) = 1.58, p > .10 (one-tailed). The same pattern was obtained in Study 1a: The difference between survival with context (M = 2.80) and survival without context (M = 2.75) ratings fell well short of significance, t(54) = 0.39, p > .30 (one-tailed). Although not central to the present discussion, it is worth noting that the mean relevance rating for mate choice (Study 1) was a full scale point above those for the survival conditions (M = 4.19) While this may appear to imply that mate choice should enjoy a memorial advantage (based on considerations of congruity), as is discussed below, there are reasons pertaining to the memorial requirements of mate selection and evolutionary selection that work against this assumption.
Survival as an evolved adaptation of the human mind
Although survival processing clearly captures an important function of human memory, from a conceptual standpoint, survival per se may not be the optimal starting point for exploring the effects of natural selection on an organism. As I have argued at length (see, Klein, in press-a, in press-b; Klein et al., 2010, 2011), it is not easy to enumerate a set of minimally necessary or sufficient behaviors that entail survival. (Nairne and colleagues have focused on survival processing rather than on a survival module per se. Considered in this context, there is no need to suggest that such a set of conditions must exist.) Indeed, as developed in biological theory, the concept of survival encompasses everything that is not growth or reproduction—clearly, an extremely heterogeneous set of activities and processes. Although natural selection can design distinct adaptations for distinct aspects of survival, it would be difficult for it to build a single adaptation for survival per se (see also Nairne & Pandeirada, 2008b).
I have argued, instead, that rather than being taken as a conceptual primitive, the concept of survival is best seen as consisting in a large collection of fitness-relevant variables, some of which obviously would benefit from enhanced memorial abilities (e.g., planning), whereas others would not (e.g., mate selection). Consistent with this proposal, a recent study by Sandry, Trafimow, Marks, and Rice (2012) found that although Nairne’s survival task produced high levels of recall, other survival-relevant tasks—for example, incest avoidance and cheater detection (e.g., Barkow et al., 1992)—failed to produce retention substantially better than that found for standard control conditions. These findings reinforce my argument that survival processing, per se, is too broad a construct to be targeted by natural selection.
The finding that mate choice does not enhance recall is consistent with my contention that the memorial consequences of the survival processing tasks employed in adaptive memory studies result primarily from the memorial demands of correlated factors—such as episodically based (1) self-referential processing and (2) personal planning (for a review, see Klein, in press-a). Survival-relevant tasks that can be performed in a proficient manner absent enhanced memorial capabilities should not be—and, as both Sandry et al., 2012, and present results suggest, have not been—selected by evolution to interface with mechanisms that enable superior memory. With respect to mate choice, it likely would not take much in the way of memory capability to successfully keep track of potential mates—particularly during the ancestral times on the savannah, when the size of one’s group numbered, at most, approximately 120 individuals (with many groups considerably smaller; e.g., Dunbar, 1996), only a subset of which would be relevant to one’s mating opportunities (due to considerations of age and gender). Accordingly, mate choice would not be expected to demonstrate special memorial abilities, despite its clear identification as a survival-relevant adaptation (e.g., Miller, 2000).Footnote 4 This, of course, is exactly what I found in the present study.
Components of survival processing and their effect on memory
In previous studies, my colleagues and I have demonstrated that factors such as planning and self-reference play a role in the survival processing memory advantage (e.g., Klein, in press-b; Klein et al., 2010, 2011; for a review, see Klein, in press-a). This is not to say, as we hopefully make clear, that these factors are conceptual primitives or that their presence necessarily is sufficient to promote optimal recall. Rather, our point is that survival processing admits to a number of memory-enhancing properties that can, and often do, work together to benefit recall (see also Burns et al., 2011).
As is the case for most mechanisms underlying behavioral outcomes, nearly all explanations of psychological processes require the assumption of multiple causality (e.g., Hanson, 1958; while one-to-one mappings of cause and effect are possible, they are highly unlikely in complex human behavior). Although the individual effects of causally relevant component processes can (and must) be taken into account, their individual effects seldom are sufficient to explain a particular outcome, and their joint effects are not necessarily linear or additive (e.g., Anderson, 1981).
The survival effect has been found to consist in several subcomponents (for a review, see Klein, in press-a), each of which, when present (this depends on a host of causally relevant factors, such as type of survival scenario, task instructions, specific memorial demands of the scenario, etc.), can have a positive effect on memory. But none, taken in isolation, necessarily is the critical factor mediating the effect. They are contributants, and their contributions depend on a number of additional factors (e.g., scenario, instructions, etc.).
Final thoughts
While survival encoding has an undeniable effect on memory proficiency, the reinstatement of the context of the EEA appears unnecessary for producing optimal recall in the adaptive memory paradigm (see also Klein et al., 2010, 2011; Soderstrom & McCabe, 2011). While this finding does not challenge the role of thoughts of personal survival in the enhancement of memory per se, it does place conceptual limitations on interpretation of this effect. Specifically, the present results call to mind Allport’s (1937) principle of functional autonomy, according to which “learning brings new systems of interests into existence just as it does new abilities and skills. At each stage of development these interests are always contemporary; whatever drives, drives now” (p. 155; emphasis added). Taken broadly as characterizing both phylogenetic and ontogenetic development (as I believe Allport intended), Allport’s principle appears at odds with the idea that imagining conditions of the EEA is necessary to maximize the effects of survival processing on memory now—that is, in its present form. Human evolution did not come to a halt 2 million years ago, 200 thousand years ago, 40 thousand years ago, or even 10 thousand years ago (e.g., Cochran & Harpending, 2009). Indeed, evidence suggests that the human brain currently is undergoing evolutionary change (e.g., Evans et al., 2005). On this view, memory is best understood in terms of its structure and function now, in the present. The findings reported herein, in conjunction with Allport’s insights and empirical evidence of ongoing changes in neural structures, suggest the assumption that reinstatement of the conditions present during the initial selection of ancestral forms of memory may not necessarily be a precondition for obtaining a survival memory effect.
Nature builds on existing structures, but those structures and their attendant processes are continually worked on by natural selection in harmony with changes in current environmental conditions. Simply put, there is no strong evolutionary principle or empirical evidence that would lead one to believe that reinstatement of the conditions present during the EEA is necessary (although it may be sufficient) for optimal survival-relevant memory performance in modern humans.
Evolutionary principles certainly serve an important heuristic role in formulating hypotheses about the functional design of biological systems (such as human memory; e.g., Glenberg, 1997; Klein et al., 2002; Nairne, 2010; Sherry & Schacter, 1987). While the origins of our ability to manifest superior recall of survival-relevant information may be traced to conditions located in the EEA (for a discussion, see Klein et al., 2010; Nairne, 2005; Soderstrom, & McCabe, 2011), the neural and cognitive underpinnings of this effect, as presently constituted and observed, are dependent to a large degree on systems and structures in the brain of modern Homo Sapiens—systems and structures that have continued to adapt as circumstance and context have changed over the past several million years of hominid evolution.
Notes
There is considerable variability in the memorial efficacy of such tasks. As will be briefly mentioned below, the nature of the “modern” (i.e., non-EEA based) survival scenario plays a central role in memory performances obtained with such tasks. Certain modern survival tasks encourage participants to imagine contexts that are relatively unfamiliar and occasionally bizarre (e.g., being pursued by assassins/attackers in a strange city), which have the unintended effect of inhibiting optimal memory performance by making it difficult for participants to engage in memory-based planning (e.g., Klein et al., 2010, 2011; for detailed discussion, see Klein, in press a).
An additional point of interest concerns the relatively high level of thoughts of planning in both the survival with context and survival without context conditions. This finding was anticipated on the basis of the demonstrated role planning plays in producing the excellent recall found with survival scenarios used in the adaptive memory paradigm (e.g., Klein, in press-a; Klein et al., 2010, 2011; Klein et al., 2012).
While working on the revision of this article, I came across an in-press article by Kostic, McFarlan, and Cleary that also finds that the survival processing memory advantage does not depend on reinstatement of ancestral context at encoding. Although this certainly is consistent with the findings of the present study, the Kostic et al. study employs as its survival sans ancestral context conditions rating tasks involving scenarios such as being stranded in space. By introducing such atypical (but familiar by virtue of popular literature and cinema) scenarios, the critique addressed to Soderstrom and McCabe (2011) holds for this study as well.
This is not to say that mate choice has no memory requirements. It does, but (1) they are minimal, in comparison with those needed to identify objects and their locations in a survival scenario of the type employed in adaptive memory paradigms, and (2) to the extent memory is functionally relevant, it is more likely to be the semantic than episodic component of long-term memory that plays the central role in impression formation (e.g., Klein et al. 2009; for reviews, see Klein & Loftus, 1993; Klein, Robertson, Gangi, & Loftus, 2008). This system of memory is not directly tested in the standard version of the adaptive memory paradigm.
References
Adam, B. (2004). Memory of futures. KronoScope, 4, 297–315.
Allport, G. (1937). The functional autonomy of motives. The American Journal of Psychology, 50, 141–156.
Anderson, N. H. (1981). Foundations of information integration theory. New York, NY: Academic Press.
Barkow, J. H., Cosmides, L., & Tooby, J. (Eds.). (1992). The adapted mind: Evolutionary psychology and the generation of culture. New York, NY: Oxford University Press.
Bradley, F. H. (1887). Why do we remember forwards and not backwards? Mind, 12, 579–582.
Brown, J. (Ed.). (1976). Recall and recognition. New York, NY: Wiley.
Burns, D. J., Hwang, A. J., & Burns, S. A. (2011). Adaptive memory: Determining the proximate mechanisms responsible for the memorial advantage of survival processing. Journal of Experimental Psychology: Learning, Memory, and Cognition, 37, 206–218.
Butler, A. C., Kang, S. H. K., & Roediger, H. L. I. I. I. (2009). Congruity effects between materials and processing tasks in the survival processing paradigm. Journal of Experimental Psychology: Learning, Memory, and Cognition, 35, 1477–1486.
Cochran, G., & Harpending, H. (2009). The 10,000 Year Explosion. New York, NY: Basic Books.
Craik, F. I. M., & Tulving, E. (1975). Depth of processing and the retention of words in episodic memory. Journal of Experimental Psychology: General, 11, 268–294.
Dawkins, R. (1976). The selfish gene. Oxford, UK: Oxford University press.
Dunbar, R. (1996). Grooming, gossip, and the evolution of language. Cambridge, MA: Harvard University Press.
Evans, P. D., Gilbert, S. L., Mekel-Bobrov, N., Vallender, E. J., Anderson, J. R., Vaez-Azizi, L. M., Tishkoff, S. A., Hudson, R. R., & Lahn, B. T. (2005). Microcephalin, a gene regulating brain size, continues to evolve adaptively in humans. Science, 309, 1717–1720.
Glenberg, A. M. (1997). What memory is for. Behavioral and Brain Sciences, 20, 1–55.
Hanson, N. R. (1958). Patterns of discovery. New York, NY: Cambridge University Press.
Hart, J., & Burns, D. J. (2012). Nothing concentrates the mind: Thoughts of death improve recall. Psychonomic Bulletin & Review, 19, 264–269.
Hyde, T. S., & Jenkins, J. J. (1973). Recall for words as a function of semantic, graphic, and syntactic orienting tasks. Journal of Verbal Learning and Verbal Behavior, 12, 471–480.
Ingvar, D. H. (1985). “Memory for the future”: An essay on the temporal organization of conscious awareness. Human Neurobiology, 4, 127–136.
Kang, S. H. K., McDermott, K. B., & Cohen, S. M. (2008). The mnemonic advantage of processing fitness-relevant information. Memory & Cognition, 36, 1151–1156.
Klein, S. B. (2007). Phylogeny and evolution: Implications for understanding the nature of a memory system. In H. L. Roediger, Y. Dudai, & S. M. Fitzpatrick (Eds.), Science of memory: Concepts (pp. 377–381). Oxford, UK: Oxford University Press.
Klein, S.B. (in press a). Evolution, memory, and the role of self-referent recall in planning for the Future. In Adaptive Memory and Survival Processing (Schwartz, B.L., Howe, M., Toglia, M., & Otgaar, H. Eds.). Oxford University Press.
Klein, S. B. (in press b). A role for self-referential processing in tasks requiring participants to imagine survival on the savannah. Journal of Experimental Psychology: Learning, Memory & Cognition.
Klein, S. B., Cosmides, L., Gangi, C. E., Jackson, B., Tooby, J., & Costabile, K. A. (2009). Evolution and episodic memory: An analysis and demonstration of a social function of episodic recollection. Social Cognition, 27, 283–319.
Klein, S. B., Cosmides, L., Tooby, J., & Chance, S. (2002). Decisions and the evolution of memory: Multiple systems, multiple functions. Psychological Review, 109, 306–329.
Klein, S. B., & Loftus, J. (1993). The mental representation of trait and autobiographical knowledge about the self. In T. K. Srull & R. S. Wyer (Eds.), Advances in Social cognition (Vol. 5, pp. 1–49). Hillsdale, N.J.: Erlbaum.
Klein, S. B., Robertson, T. E., & Delton, A. W. (2010). Facing the future: Memory as an evolved system for planning future acts. Memory & Cognition, 38, 13–22.
Klein, S. B., Robertson, T. E., & Delton, A. W. (2011). The future-orientation of memory: Planning as a key component mediating the high levels of recall found with survival processing. Memory, 19, 121–139.
Klein, S. B., Robertson, T. E., Delton, A. W., & Lax, M. L. (2012). Familiarity and personal experience as mediators of recall when planning for future contingencies. Journal of Experimental Psychology: Learning, Memory, and Cognition, 38, 240–245.
Klein, S. B., Robertson, T. E., Gangi, C. E., & Loftus, J. (2008). The functional independence of trait self-knowledge: Commentary on Sakaki (2007). Memory, 16, 556–565.
Kroneisen, M., & Erdfelder, E. (2011). On the plasticity of the survival processing effect. Journal of Experimental Psychology: Learning, Memory, & Cognition, 37, 1553–1562.
Miller, G. (2000). The mating mind: How sexual choice shaped the evolution of human nature. London, UK: Heineman.
Nairne, J. S. (2005). The functionalist agenda in memory research. In A. F. Healy (Ed.), Experimental cognitive psychology and its applications: A Festschrift in honor of Lyle Bourne, Walter Kintsch and Thomas Landaur. Washington DC: American Psychological Association.
Nairne, J. S. (2010). Adaptive memory: Evolutionary constraints on remembering. In B. H. Ross (Ed.), The psychology of learning and motivation (Vol. 53, pp. 1–32). London, UK: Academic Press.
Nairne, J. S., & Pandeirada, J. N. S. (2008a). Adaptive memory: Is survival processing Special? Journal of Memory and Language, 59, 377–385.
Nairne, J. S., & Pandeirada, J. N. S. (2008b). Adaptive memory: Remembering with a stone-age brain. Current Directions on Psychological Science, 17, 239–243.
Nairne, J. S., & Pandeirada, J. N. S. (2010). Adaptive memory: Ancestral priorities and the mnemonic value of survival processing. Cognitive Psychology, 61, 1–22.
Nairne, J. S., & Pandeirada, J. N. S. (2011). Congruity effects in the survival paradigm. Journal of Experimental Psychology: Learning, Memory, and Cognition, 33, 263–273.
Nairne, J. S., Pandeirada, J. N. S., Gregory, K. J., & Van Arsdall, J. E. (2009). Adaptive memory: Fitness relevance and the hunter-gatherer mind. Psychological Science, 20, 740–746.
Nairne, J. S., Pandeirada, J. N. S., & Thompson, S. R. (2008). Adaptive memory: The comparative value of survival processing. Psychological Science, 19, 176–180.
Nairne, J. S., Thompson, S. R., & Pandeirada, J. N. S. (2007). Adaptive memory: Survival processing enhances retention. Journal of Experimental Psychology: Learning, Memory, and Cognition, 33, 263–273.
Nairne, J. S., VanArsdall, J. E., Pandeirada, J. N. S., & Blunt, J. R. (2012). Adaptive memory: Enhanced location memory after survival processing. Journal of Experimental Psychology: Learning, Memory, and Cognition, 38, 495–501.
Otgaar, H., Smeets, T., & van Bergen, S. (2010). Picturing survival memories: Enhanced memory after fitness-relevant processing occurs for both verbal and visual stimuli. Memory & Cognition, 38, 23–28.
Rogers, T. B., Kuiper, N. A., & Kirker, W. S. (1977). Self-reference and the encoding of personal information. Journal of Personality and Social Psychology, 35, 677–688.
Sandry, J., Trafimow, D., Marks, M., , & Rice, S. (2012). Adaptive memory: Evaluating alternative forms of fitness-relevant processing in the survival processing paradigm. Unpublished manuscript.
Sherry, D. F., & Schacter, D. L. (1987). The evolution of multiple memory systems. Psychological Review, 94, 439–454.
Soderstrom, N. C., & McCabe, D. P. (2011). Are survival processing memory advantages based on ancestral priorities? Psychonomic Bulletin & Review, 18, 564–569.
Suddendorf, T., & Corballis, M. C. (1997). Mental time travel and the evolution of the human mind. Genetic, Social, and General Psychology Monographs, 123, 133–167.
Weinstein, Y., Bugg, J. M., & Roediger, H. L. (2008). Can the survival recall advantage be explained by basic memory processes? Memory & Cognition, 36, 913–919.
Williams, G. C. (1966). Adaptation and natural selection: A critique of some current evolutionary thought. Princeton: Princeton University Press.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Klein, S.B. Does optimal recall performance in the adaptive memory paradigm require the encoding context to encourage thoughts about the environment of evolutionary adaptation?. Mem Cogn 41, 49–59 (2013). https://doi.org/10.3758/s13421-012-0239-8
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13421-012-0239-8