
Abstract
Recent research using the Posner cuing paradigm to explore inhibition of return seems to suggest that repetition of a nonspatial feature modulates cue-related facilitatory and inhibitory aftereffects differently in detection and discrimination tasks. Because the cues were unrelated to the final response in the detection task but parallel to (not orthogonal with) the final response in the discrimination task, it is unclear whether the different patterns of results were caused by the complexity of the decision (detection vs. discrimination) or by the task relevance of the feature that might or might not repeat from cue to target. Using a paradigm modeled on previous work, in Experiment 1 (detection task) and 2 (discrimination task) we replicated the previous patterns: No early feature repetition benefit but reduced IOR for feature mismatch trials in the detection task; and large early feature repetition benefit but no effect of feature match upon the later IOR in the discrimination task. In Experiment 3 (discrimination task), we used an “orthogonal-cuing” method: the feature (color) that could repeat or not from cue to target was not on the dimension being discriminated (shape). The pattern of results was very similar to what is observed in detection tasks. These results demonstrate that it is not the task but the task relevance of the repeating feature that modulates facilitation and inhibition effects. The findings are generally consistent with a habituation account of the inhibitory aftereffects of orienting but do not rule out the contribution of other mechanisms.
Similar content being viewed by others
Interest in the inhibitory aftereffects of exogenous orienting was launched by Posner and Cohen’s (1984) paper, “Components of Visual Orienting.” Using uninformative peripheral cues and peripheral targets calling for a simple detection response, they discovered that following a period of facilitation at the cued location, the reaction time (RT) functions for cued and uncued targets crossed over such that the time to detect targets at the cued location was slower than at the uncued location when the cue-target stimulus onset asynchrony (CTOA) was greater than about 300 ms. In the following year, Posner, Rafal, Choate, and Vaughan (1985) named this inhibitory aftereffect “inhibition of return” (IOR) and, reinforcing Posner and Cohen’s suggestion that it might encourage orienting to novelty, described it as a bias that might operate in visual search.
In the 30 years since these seminal papers were published, IOR has received considerable attention (Klein, 2000; Lupiáñez, 2010; Lupiáñez, Klein & Bartolomeo, 2006; Wang & Klein, 2010). One of the many interesting questions about IOR is whether, and if so how, it might be modulated by cue-target relations on dimensions/attributes other than location (e.g., Fox & de Fockert, 2001; Hu, Fan, Samuel, & He, 2013; Hu & Samuel, 2011; Hu, Samuel & Chan, 2011; Hu, Zhan, Li, He, & Samuel, 2014; Kwak & Egeth, 1992; Tanaka & Shimojo, 1996; Taylor & Donnelly, 2002).
The present experiments on this topic were stimulated by, and are designed to replicate and extend, two recent publications (Hu & Samuel, 2011; Hu, Samuel & Chan, 2011) that used the fecund paradigm launched by Samuel and Weiner (2001). Although strongly rooted in the “uninformative cue-target paradigm” pioneered by Posner (1980; Posner & Cohen, 1984), the Samuel cuing paradigm is distinguished from it by the complexity of the display before any cues or targets are presented and by the nature of the cues and targets. In the standard, and most simple version of the Posner cuing paradigm, each trial begins with two boxes—one on either side of fixation. The cue is the brightening of either box, and the target is an asterisk that appears inside one of the boxes. In the Samuel paradigm, there are eight equi-eccentric gray discs spaced evenly around fixation. Some of these are filled with one or two stimuli that might differ in color, shape, or both. In the version of the paradigm that is the focus here, the cue is the appearance of a stimulus in one of the empty discs, and the target is the appearance of a stimulus in the same or a different disc. As in Posner’s paradigm, the key independent variables are the CTOA between the cue and target and whether the target is in the cued or a different location. The Samuel paradigm also allows the investigator to look, coarsely, at the distance between the cue and target. Importantly, and for present purposes, critically, the Samuel paradigm allows the investigator to present cues and targets that match or mismatch on a feature (color/shape) that is not a spatial location.
The present story starts with Hu et al.’s (2011) extension of Samuel and Weiner (2001). As in Samuel and Weiner, and as in Posner and Cohen, the participant’s task was simple, speeded detection of the occurrence of the target. An important difference between the Posner and Samuel paradigms is that in the Posner cuing paradigm cues and targets are usually different: brightening of a box and onset of a stimulus inside a box, whereas as in Hu et al. cues and targets are, aside from their time of occurrence, interchangeable. Although in both paradigms the cues can be used to prepare for responding to targets, in Hu et al. they must be, because if a cue in their paradigm were missed the participant would not know for sure if the target were a cue or target. While using a wide range of CTOAs (from 200 to 3,500 ms), Hu et al. manipulated whether the target did or did not match the cue in a nonspatial attribute: color or shape. As illustrated in Fig. 1a, targets that matched the cue’s nonspatial attribute suffered from location-based IOR (inhibition match) beginning by 350 ms from cue onset, reaching a maximum by 800 ms and decreasing only slightly at the longer CTOAs. Targets that did not match the cue’s nonspatial attribute behaved like those that did up until the 350 ms CTOA, and for all subsequent CTOAs suffered less IOR (if not none) at the remaining CTOAs (inhibition mismatch). Accordingly, this pattern (which is highlighted by the red arrow in Fig. 1a) can be described as greater IOR for targets that match rather than mismatch the cue on a nonspatial feature. We will use these two descriptions (“inhibition mismatch < inhibition match” and “inhibition match > inhibition mismatch”) interchangeably.

The effects on RT of Target Location and Target Feature Match (left panel) vs. Mismatch (right panel) as a function of CTOA. (a) Results adapted from Hu et al. (2011) using a target detection task. (b) Results adapted from Hu and Samuel (2011) using a nonorthogonal discrimination task. The solid green lines show the average RT for the two shortest CTOAs. The dashed green lines connecting the two panels allow us to visualize the benefit for feature match trials that is apparent in the discrimination task (lower panel) and illustrated by the vertical, green arrow. With no such benefit in the detection task it was not possible to display an arrow (upper panel). Orange lines in all the panels are plotted to represent the mean RT for the 4 longest CTOAs. These lines are solid for trials with targets at the cued location and dashed for uncued targets. The difference between these lines (vertical orange arrows) represents the IOR effects in each panel. The matching IOR effects are replotted on the right side of the figure where they can more easily be compared to the mismatching IOR effects. The vertical red arrow illustrates the reduction of IOR on mismatch trials that was observed in the detection task (upper panel). A similar arrow could not be plotted for the discrimination task (bottom panel) because here, there was no difference. Since here, “Same Loc” means that both target and cue appeared at the same location, whereas “Different Loc” indicates that they appeared at the different locations
The reduced cost (IOR) when spatially cued targets mismatched on a nonspatial feature with the cue was viewed as inconsistent with the traditional view of IOR, which, as implied by its name, is characterized by the inhibition of attention from returning to previously attended locations. Yet, it was viewed as consistent with two recent frameworks, often viewed as alternatives to the traditional account: Dukewich’s (2009) habituation account and Lupiáñez’s (2010) detection cost account. Unlike the traditional view of IOR, Dukewich reconceptualized IOR as habituation of the orienting response (Sokolov, 1960, 1963). In this view, habituation would be most robust when a sequence of identical stimuli is presented at the same location, whereas a target that differs from the preceding stimulus would be expected to benefit from dishabituation. Lupiáñez proposed a three-component framework to describe the time course of benefits and costs following an uninformative peripheral cue. The three components are (1) spatial selection benefit, (2) spatial orienting benefit, and (3) onset detection cost – the former two components facilitate performance whereas the latter component impairs it. It is further assumed that detection cost contributes to performance more in detection tasks than in discrimination tasks. From this perspective, the component that ought to accommodate IOR’s sensitivity to nonspatial features is the onset-detection cost, which is reduced by a cue-target featural mismatch.
Hu and Samuel (2011) used a very similar procedure to Hu et al. (2011) except their participants were performing a discrimination task. Importantly, as we will show, the discrimination task required a response that was parallel to (i.e., nonorthogonal with) the feature match/mismatch manipulation. When cues and targets were blue or red circles, the task was to make a blue/red discrimination about the targets’ color; and, when cues and targets differed in shape (filled circles vs. empty squares), participants made a shape discrimination. As illustrated in Fig. 1b, at the shortest CTOA there was a small location-based RT benefit (i.e., facilitation), and at the two shortest CTOAs there was a large feature-based RT benefit, which is highlighted by the green arrow in this Figure. At the longest two CTOAs there was location-based RT cost (i.e., IOR) that was insensitive to the non-spatial relation between the cue and target. At the two intermediate CTOAs these two effects combine roughly additively.
There were two remarkable differences in results between the discrimination task used by Hu and Samuel (2011) and the detection task used by Hu et al. (2011): First, using the discrimination task, Hu and Samuel found a strong, early (CTOA <400 ms) feature repetition benefit that was absent in Hu et al.; second, the extra IOR for stimuli whose nonspatial features matched between the cue and target with the detection task was eliminated in the discrimination task (CTOA >400 ms). These two differences are highlighted by the green and red arrows in Fig. 1 and are plotted in panels (a) and (b) of Fig. 2. Note that in Fig. 2 the early feature repetition benefit is calculated by [RT (feature match)–RT (feature mismatch)] (CTOA<400 ms), whereas the later extra IOR for feature match is indexed by the mean of [IOR (feature match)–IOR (feature mismatch)] (CTOA >400 ms).

The early feature repetition benefit and the later extra IOR for feature matches are plotted from 5 sources: (a) Hu et al. (2011); (b) Hu and Samuel (2011); (c) Experiment 1; (d) Experiment 2; (e) Experiment 3. The task was simple detection in Panels (a) and (c), and discrimination in the remaining panels. Panels (c) and (d) are our close replications of (a) and (b). Although Experiment 3 (panel e) used a discrimination task (as in panels b and d), the response was orthogonal to the possibly repeating feature whereas response and feature were confounded in the previous studies
Hu and Samuel favored the three-component framework over the habituation account because, while admittedly less parsimonious, it is “more complex” and therefore was assumed to be better able to handle complex patterns of results, like those of Hu and colleagues. Yet, our further analysis indicates that some of the attributions made by Hu and Samuel seem flawed. For example, Hu and Samuel (2011) explained the feature repetition benefit at the early CTOAs by appealing to a second component of the Lupiáñez (2010) framework: the spatial selection benefit. According to the three-component framework, a target that appears in close spatiotemporal proximity to the cue is likely to be integrated into the same object file, which is facilitated on cued trials by helping select the target location before it is presented. The benefit starts high at short cue-target CTOAs, diminishes over time, and is enhanced when task-relevant features of the cue and target overlap. A problem for this explanation is that these benefits ought to be confined to the cued location, yet they were approximately equal for the cued and uncued locations. In a similar vein, the three-component framework predicts a reduction of IOR on featural mismatch (vs. match) trials, as was observed in Hu et al. (2011). Yet, contrary to this prediction, the magnitude IOR was almost identical for featural match and mismatch conditions in Hu and Samuel (2011).
The three-component framework of Lupiáñez (2010) specifically asserts that “onset detection processes contribute mostly to detection tasks, spatial selection processes are tapped mainly by discrimination tasks” (p. 27). These assertions were at the core of the rationale for Hu and Samuel’s (2011) repeat of Hu et al.’s (2011) detection task using a discrimination task. First, they asserted that the habituation framework could not accommodate detection/discrimination differences and went on to say this about the three-component model:
To the extent that performance turns out to be task-dependent (e.g., detection vs discrimination) and stimulus property-dependent (e.g., location vs nonspatial feature), the habituation model does not appear to offer a basis for such dependencies; it certainly does not predict them. . . . A key strength of the model is that it allows differential predictions for detection versus discrimination tests: Detection experiments should be most affected by the detection cost factor’s time course, whereas discrimination tasks that presumably benefit more from properly focused attention should be driven more by the time course for the spatial factors. This feature of the model, while coming with a cost in parsimony, gives it an advantage over the other two models in explaining why discrimination tasks produce different results than those for detection. (Hu & Samuel, 2011, p. 787)
It is critically important that in Hu and Samuel (2011), the cuing dimension (color in Experiment 1; form in Experiment 2) was not orthogonal to the response dimension. For example, if the cue stimulus was a blue (or red) circle, then the target was a blue (or red) circle calling for a red/blue discrimination. This untoward design feature allows for the possibility that some of the observed effects were not due to the mechanisms hypothesized by Posner, Lupiáñez, or Dukewich to explain the time course of exogenous cuing, but rather were caused by cue-elicited automatic response activation. That is, when participants are responding differentially to two values (e.g., blue/red) of a dimension (color) of the target, and when these values are present in a nontarget stimulus (the cue), the habitual responses are likely to be activated, at least briefly (e.g., see Neill, 1978), thereby untowardly contributing to performance.
To avoid this possibility (as recommended, for example, by Spence & Driver, 1994, 1997) the cuing and response dimensions should be orthogonal rather than parallel. We thus hypothesized that the RT benefit for a feature repetition that was uniquely present in Hu and Samuel (2011) was caused by cue-elicited response activation rather than the enhanced spatial selection benefit attributed to the mere use of a discrimination task. We will directly test this hypothesis in Experiment 3 by conducting an orthogonal discrimination task in which the nonspatial feature of the cue that might match or mismatch with that of the target is orthogonal to the dimension being discriminated. To provide the appropriate baselines against which to compare the findings from Experiment 3 (and to ensure replicability of the key results illustrated in Figs. 1 and 2), we conducted approximate replications (i.e., Experiment 1 and Experiment 2) of the key methods from Hu et al.’s (2011) detection, and Hu and Samuel’s (2011) nonorthogonal discrimination, tasks. Importantly, if Hu and Samuel correctly attributed their early feature-repetition RT benefit to the mere use of a discrimination task, then this effect should be observed in both Experiments 2 and 3, but not in Experiment 1. Conversely, if the early RT benefit is only observed in Experiment 2, then the early feature-repetition benefit reported by Hu and Samuel (2011) can be confidently attributed to cue-elicited response activation. In that case, Hu and Samuel’s rationale for favoring the Lupiáñez account over the Dukewich account would be undermined.
General method
Participants
Participants in all three experiments were undergraduate and graduate students from Peking University. All had normal or corrected-to-normal color vision and were naïve to the purpose of the experiment. Participants were tested individually; each received 20 RMB (about U.S. $3) for participating. All participants were treated in accordance with the Declaration of Helsinki and its latest amendments, and provided a written informed consent before participating in the study.
Apparatus, stimuli and procedure
The experiments were conducted on a Pentium IV computer running E-Prime software (Schneider, Eschman, & Zuccolotto, 2002), with participants viewing the screen from a distance of approximately 65 cm. A computer keyboard was directly in front of the subject, and its space bar or Numlock pad was used as the response device.
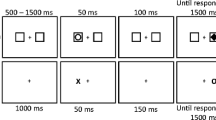
The apparatus, stimuli, and procedures were based on those used by Samuel and Weiner (2001; Samuel & Kat, 2003). Figure 3 shows the sequence of events on a trial. Each trial began with a gray fixation square (1° of visual angle; VA) displayed for 250 ms. This was followed by a display that consisted of eight white discs (diameter: 3.7° VA) presented for 750 ms. The white discs were arranged in a circular fashion around the fixation square (radius: 6.8° VA). Half of the white discs were filled with two smaller objects, either a red/blue circle, or a black/red/blue cross/plus. Empty discs alternated with filled discs. The cue, which was either a red or blue circle, then appeared in one of the four empty discs. Finally, a pseudo-randomly selected red/blue cross or plus serving as the target was presented, either within the same disc as the cue (Same Location condition), or within one of the other originally empty circles (Different Location condition). As Fig. 3 shows, the cue remained visible in the display once it appeared. The cue – target CTOAs were pseudo-randomly selected to be 200; 800; 1,600; or 3,200 ms. The longer CTOA was used as our previous work suggested that feature-based effects may require relatively long CTOAs. To look for feature repetition effects, half of the target objects matched the cues in color (Feature Match condition), and the other half mismatched the cues in color (Feature Mismatch condition). Targets were equally likely to be presented at each possible location. In this aspect, the present experiment was slightly different from the corresponding experiment in Hu et al. (2011), as in that study, on one-third of the trials, the target was presented in the same circle as the cue; on one-third of the trials, the target was displayed in a circle 90° away from the cue, either clockwise or counterclockwise; and on one-third of the trials, the target was displayed in the circle 180° away from the cue. Note as before that on a Same Location trial the cue and the target appeared within the same circle but were always in slightly different positions. In all experiments, trials were terminated by the subject’s response or after a time-out period of 3,000 ms.

Example of the sequence of events for a sample trial in Experiments 1, 2, and 3 (not drawn exactly to scale). Each trial begins with the fixation, then initial display, cue, CTOA (not illustrated here) target, and ends with the response. The cue is a small red circle that appears in a disc near the top of the third frame and the target is a small red plus sign that appears in the same disc. This example depicts a Feature Match/Same Location trial
Each participant was presented with four blocks of 140 trials. The 128 experimental trials within each block included the factorial crossing of 4 CTOAs × 4 possible cue locations × 4 possible target locations × 2 target color conditions (Feature Match or Mismatch). In total there were 512 experimental trials. Target shapes (i.e., cross or plus) were also balanced across the trial conditions. Thus, for each participant, there were 16 observations for each combination of CTOA, Target Location, and Target Feature.Footnote 1 There were also 12 catch trials per block, in which a cue but no target appeared. A rest was offered after each block.
The participant was instructed to fixate on the central fixation square throughout the experiment. The task varied between experiments, as described below.
Each participant was given a practice block of 30 trials that were not analyzed. Both speed and accuracy were emphasized. If a participant responded incorrectly, or failed to respond, a tone alarm was presented as feedback. Participants were tested individually in a darkened, sound-attenuated room.
Methods of analysis
As there were no systematic differences between targets in the Different Location conditions ( 90° clockwise, 90° counterclockwise, or 180° from the cued location), these conditions were collapsed into a single “Different Location” condition.Footnote 2 Prior to analysis, trials with reaction times of 2.5 standard deviations above or below the arithmetic mean were excluded for each participant. This resulted in the loss of 3.94 %, 3.08 %, and 3.19 % of the trials in Experiments 1, 2, and 3, respectively. Mean RTs were then submitted to a 2 (Target Feature: Match vs. Mismatch) × 4 (CTOA: 200, 800, 1600, and 3200 ms) × 2 (Target Location: Same vs Different) analysis of variance (ANOVA), with degrees of freedom corrected for any violations of the sphericity assumption (Jennings & Wood, 1976). For every reported F ratio, the uncorrected MSE is also reported along with the uncorrected degrees of freedom if those reported with the F had been corrected. Error rate data were analyzed in the same way, when necessary. To determine the similarity of the pattern of results to that of the studies reviewed in the introduction, for each experiment we calculated the early feature repetition benefit at short CTOAs (CTOA <400 ms), as well as the later extra IOR for featural matching at later CTOAs (CTOA >400 ms), as described in the introduction.
Experiment 1: Detection task
With minor procedural changes, Experiment 1 was a replication of Hu et al.’s (2011) Experiment 2.
Method
There were 36 participants (age range = 19–25, mean 22; 19 males, all right-handed). Participants were instructed to press the spacebar as soon as they saw the second item (i.e., the target) appear, and were asked to refrain from responding on catch trials.
Results and discussion
The miss rates and false alarm rate on catch trials were very low (less than 1 %), and therefore no further analyses were conducted. As can be seen in Fig. 4, the results in this experiment closely match those observed in Hu et al. (2011).

Mean RT results from Experiment 1, using a detection task. The effects of Target Location as a function of Target Feature Match (left panel) and Feature Mismatch (right panel) across various CTOAs
In the analysis of mean RT, the main effect of Target Location was significant, F(1, 35) = 6.97, MSE =1,700, p = .012, as was the two-way interaction of Target Location × CTOA, F(2, 72) = 32.45, MSE = 1,802(105), p < .001. This latter interaction reflects a transition from early attentional capture, as seen in the 50.6 ms of early facilitation at the cued location, CTOA 200 ms, t(35) = 5.14, p < .001, to 29.0 ms of IOR at the later CTOAs, smallest t(35) = 4.70, p < .001.
The main effect of Target Feature, F(1, 35) = 9.70, MSE = 948, p = .004, and the interaction of Target Location × Target Feature, F(1, 35) = 7.73, MSE = 1,444, p = .009, were significant. The IOR effect was significant for each CTOA greater than 200 ms in both the cue-target match condition (smallest t(35) = 3.58, p = .001) and mismatch condition (smallest t(35) = 2.06, p = .047). Importantly, IOR was reduced when cue-target features mismatched (see below). No other significant effects were detected.
The pattern of results (see Fig. 2c) is remarkably similar to that from the detection task of Hu et al. (2011): essentially, there was no early feature repetition benefit and at the longer CTOAs there was substantially more IOR on trials with cues and targets matching in color (40.4 ms) compared with the color mismatching trials (17.5 ms), t(35) = 2.50, p = .017.
Experiment 2: Parallel discrimination task
In Experiment 2, we used the same stimuli as Experiment 1 while changing the task from simple detection to a two-choice color discrimination. The goal of this experiment was to replicate Hu and Samuel (2011) in which featural match between cue and target generated an early benefit and did not modulate the later IOR effect.
Method
Thirty-eight new undergraduate and graduate students from Peking University were recruited (age range = 18–29, mean 22; 23 males, all right-handed). Experiment 2 was identical to Experiment 1 with the following exceptions. Instead of using a simple target detection response, participants were instructed to make a two alternative forced choice (2-AFC) response on each trial regarding the color of the target. The NumLock pad was used as a response device. Participants pressed either Key 1 or Key 2 to indicate whether the target was red or blue. The key-response mapping was counterbalanced across participants. We refer to this variation of the paradigm as a parallel discrimination task, as the feature on which the target can match or mismatch the cue (color) is parallel (nonorthogonal) to the judgment that participants were asked to make about the target (color).
Results and discussion
The miss rates and false-alarm rate on catch trials were both under 1 %, and therefore no further analyses were conducted. Error rates were also low (mean 1.98 %). The ANOVA of error rates only showed a significant two-way interaction of Target feature x CTOA, F(3, 111) = 3.93, MSE = .001, p = .010. The simple effect of featural matching was not significant for any CTOA.
As can be seen in Fig. 5, the results of Experiment 2 closely match those observed in Hu and Samuel (2011). In the analysis of mean RT, the main effect of Target Location was marginally significant, F(1, 37) = 3.59, MSE = 2,113, p = .066, and, reflecting the transition from attentional capture to IOR, the interaction of Target Location with CTOA, F(2, 77) = 10.09, MSE = 2271(111), p < .0001, was significant. At the shortest CTOA (200 ms), the appearance of the cue and target at the same location produced a significant RT benefit, 29 ms, t(37) = 3.18, p = .003, while for CTOAs of 800 ms and longer, there were RT costs for Same Location trials (overall 19 ms). This was not significant for the 800 ms CTOA condition, t(37) = 1.49, p = .14), but was for the two longest CTOAs, 1,600 ms, t(37) = 2.56, p = .015; 3,200 ms, t(37) = 4.50, p < .001. When compared with Experiment 1, the time course difference is consistent with Lupiáñez, Milán, Tornay, Madrid, and Tudela (1997) in that IOR appeared later when the task was discrimination than when it was simple detection.

Mean RT results from Experiment 2, using a parallel discrimination task
The main effects for Target Feature, F(1, 37) = 5.56, MSE = 6,968, p = .024, and CTOA, F(2, 92) = 7.45, MSE = 2,980(111), p < .0001, were also significant, as was the two-way interaction of Target Feature x CTOA, F(3, 94) = 3.35, MSE = 2,103(111), p = .029. This interaction reflects the finding of a robust, 36 ms, t = 3.83, p < .0001, feature repetition benefit at the 200 ms CTOA which was absent at the later CTOAs. Neither the two-way interaction of Target Location x Target Feature, F(1, 37) = .30, MSE = 1,739, p = .590, nor the three-way interaction, F (3, 111) = .67, MSE = 1,822, p = .574, were significant. The nonsignificance of these higher order interactions means that the early feature repetition benefit was similar at both the cued and uncued location and that the later IOR effect was not (in contrast to Experiment 1) modulated by whether or not there was a feature change. As can be seen in Fig. 2d, the pattern of results is remarkably similar to that from the nonorthogonal discrimination task of Hu and Samuel.
Experiment 3: Orthogonal discrimination task
Primarily to evaluate whether the early RT benefit for the repetition of a nonspatial feature between the cue and target was due to cue-elicited response activation, Experiment 3 used an orthogonal discrimination task, wherein the feature repetition was orthogonal to, rather than parallel with, the dimension being discriminated. If the effect of nonspatial feature match/mismatch between the cue and target in Experiment 2 was due to response priming, we would expect the early RT benefit for the Feature Match condition to be eliminated in Experiment 3.
Method
Another 36 undergraduate and graduate students from Peking University were recruited (age range = 18–26, mean 22; 12 males, all right-handed). Experiment 3 was identical to Experiment 2 with the following exceptions. Instead of discriminating the color of the target, participants performed a 2-AFC task in which they discriminated whether the shape of the target was a plus, or a cross. We refer to this variation of the paradigm as an orthogonal discrimination task because the feature on which the target can match or mismatch the cue (color) is orthogonal to the judgment that participants were asked to make about the target (shape).
Results and discussion
The miss rates and false alarm rate on catch trials were under 1 %. The error rates (mean = 2 %) were low. No effects were significant when the errors were submitted to an ANOVA.
As can be seen in Fig. 6, the results of the present orthogonal discrimination task (Exp. 3) closely match those from a simple detection task (see Figs. 1a and 4) and not those from the nonorthogonal discrimination task (see Figs. 1b and 5). In the analysis of mean RT, the main effects of Target Location and CTOA were significant, F(1, 35) = 21.94, MSE = 2,907, p < .001, and F(1, 35) = 7.27, MSE = 5,012, p = .001, respectively. Reflecting the transition from attentional capture to IOR, the interaction between Target Location and CTOA was also significant, F(2, 83) = 19.74, MSE =3,648(105), p < .0001. At the shortest CTOA (200 ms), the appearance of the cue and target at the same location produced a significant RT benefit, 45.9 ms, t(35) = 4.39, p < .0001, while for CTOAs of 800 ms and longer, there were RT costs for Same Location trials, overall 43.3 ms, the smallest t(35) = 4.15, p < .0001.

Mean RT results from Experiment 3, using an orthogonal discrimination task
The main effect of Target Feature, F(1, 35) = 3.65, MSE = 1,185, p = .064, was marginally significant. Critically, the two-way interaction of Target Feature x Target Location, F(1, 35) = 10.25, MSE = 1,160, p = .003, and the three-way interaction of CTOA x Target Feature x Target Location, F(3, 105) = 3.95, MSE = 2,333, p = .010, were significant. The interaction of CTOA and Target Feature was marginally significant, F(3, 105) = 2.56, MSE = 1,606.48, p = .059. The IOR effect was significant for all CTOAs greater than 200 ms when cue-target feature matched, smallest t(35) = 2.83, p = .008. Regarding the cue-target feature mismatch condition, the IOR effect was significant for two longer CTOAs, 1,600 ms, t(35) = 3.36, p = .002; 3,200 ms, t(35) = 3.40, p = .002. At CTOA 800 ms, the inhibitory effect was not significant, t(35) = 1.73, p = .093. As in Experiment 1, IOR was significantly reduced (from 57.3 ms to 29.3 ms) when the cue-target features mismatched, t(35) = 2.87, p = .007.
As can be seen in Fig. 2e, even though a discrimination task was used here, the results of Experiment 3 closely match those of Experiment 1: essentially there was no early feature repetition benefit (1.4 ms), and at longer CTOAs there was substantially more IOR (28.0 ms) on trials with cues and targets matching in color.
General discussion
In the present study, we investigated the effects of repeating nonspatial features of the cue and target in the spatial cuing paradigm, with a particular focus on the potential relationship between the nonspatial feature being repeated and the target response. We equated all stimulus factors and experiment settings across our three experiments. Using a detection task, in Experiment 1 we successfully replicated Hu et al. (2011): IOR was greater when the cue and target were the same color than when the color changed. Using what we call a parallel discrimination task, in Experiment 2 we successfully replicated Hu and Samuel (2011): at a 200 ms CTOA there was robust facilitation when the cue and target shared the nonspatial feature of color and the later IOR was unaffected by this match. Importantly, in these papers by Hu, Samuel and colleagues, when cues and targets matched in color they were identical stimuli. In contrast, in our experiments whether or not cues and targets matched in color they were never identical (see Fig. 3). From an empirical point of view, then, our fairly close replication of the pattern of results from Hu et al.’s studies demonstrates that those findings do not depend on an identical match.
The purpose of Experiment 3, in which we maintained the discrimination requirement of Experiment 2 while making the cued dimension (color) orthogonal to the discriminated dimension (shape), was to determine if the different patterns were due to the nature of the task (detection vs. discrimination) or to the task-relevance of the nonspatial (color) feature of the cue and target. The results from Experiment 3 reproduced the key findings from Experiment 1, indicating that none of the differences between Experiment 1’s detection task and Experiment 2’s discrimination task are due to the change from detection to discrimination.
A summary of the results from Hu’s studies and the three experiments reported here, focusing on the early feature repetition benefit and the later reduced IOR for feature mismatches (increased IOR for feature matches), are presented in Fig. 2. Two patterns are evident here: (1) the RT benefit for the repetition of nonspatial features at short CTOAs depends on the response relevance of the repeated feature and is almost certainly generated by cue-elicited response activation; and (2) IOR is reduced by a color change from the cue to the target, but only if this color change is task irrelevant.
The most emphasized finding in Hu and Samuel (2011) was that the effects of repetition (of location and feature) varied dramatically with the type of task participants were performing. For the reasons outlined in the introduction, this finding led Hu and Samuel to side with the three-component explanation of IOR over alternative explanations, such as habituation. The similarity of results from our detection (Experiment 1) and orthogonal discrimination (Experiment 3) tasks demonstrates that neither the early RT benefit for nonspatial feature repetition, nor the reduction of IOR when a nonspatial feature changes is about the task type. Rather both of these findings are about whether the repeated feature is or is not task relevant. The finding that it was not task difficulty that was responsible for the early feature repetition facilitatory effect removes one reason provided by Hu and Samuel for favoring the three-component framework over the habituation account. When this finding is considered together with the complexity of the three-component framework, its failure to accommodate the nearly identical feature repetition benefits at the cued and uncued locations in Experiment 2 (and in Hu & Samuel, 2011) and the similar amount of IOR in our detection and orthogonal discrimination tasks, we are compelled to favor the habituation account. Although we will focus upon the habituation account in the remainder of this paper, for the interested reader we present in the appendix a summary of the findings reported here (and the similar studies of Hu and colleagues) along with our view of how each finding might be accommodated by or might be problematic for the habituation and three-component frameworks.
Another finding uncovered by our experiments is that the reduction of IOR when there was a color change from the cue to the target, which was present in the detection task and absent in the nonorthogonal discrimination task, was present in the orthogonal discrimination task. How might this pattern be explained by Dukewich’s habituation account? It is a general principle of habituation that it is modulated by task relevance: Habituation is about learning to ignore repeating stimuli that have no behavioral significance (Rankin et al., 2009). In a standard exogenous cuing paradigm, cues are objectively irrelevant because they do not predict the location or identity of the upcoming, task-relevant target. However, in the nonorthogonal discrimination task we used in Experiment 2 (and as was used by Hu & Samuel), the participant’s attentional system is set to make a red/blue color discrimination. As hypothesized by Klein (2000), to explain the sensitivity of the time course of IOR to different task parameters, the attentional control setting that is put in place for processing targets will apply to the cues. Hence, despite its objective task irrelevance, the cues in this discrimination task are less likely to suffer from habituation than in either the detection or orthogonal discrimination tasks for which color is completely irrelevant. Cast in broader terms, the modulation of IOR by the nonspatial feature seems to depend on that feature being irrelevant. Once it is attended (e.g., in the nonorthogonal discrimination task), there is little or no habituation. If this view is correct, and maintaining the habituation account, habituation will be observed most robustly when color is irrelevant and the cues and targets match in color (and location). As seen in Fig. 7, the two conditions with the most robust IOR scores fit this description. We see two broadly different ways to explain this pattern within the habituation framework. In the unimodal explanation all of these effects are akin to habituation, which is reduced when the cue and target differ in a salient, nonspatial feature or when the cues are attended because they share a task-relevant feature with the targets.

IOR scores for the 6 location repetition conditions derived from the 3 experiments by averaging the effects from the 3 longer CTOAs (800, 1,600, and 3,200 ms)
The unimodal view can accommodate most of the findings shown in Fig. 7, with the possible exception of the absence of an effect of color repetition in Experiment 2. An alternative, bimodal view accepts that habituation is occurring when the cues and targets match in color but only when the cues do not fit the observer’s attentional control setting (Experiments 1 and 3) and it assumes a second mechanism, perhaps IOR in the sense meant by Posner et al. (1985; see Hilchey, Klein & Satel, 2014) or perhaps another inhibitory aftereffect of cuing (Berlucchi, 2006; e.g., see Hilchey, Satel, Ivanoff, & Klein, 2013), that is operating, relatively equally, in all six conditions.
There is a growing interest (e.g., see Huber, 2014) around the notion, explored here and in the studies by Hu and colleagues, that sequential presentation of stimuli that share stimulus properties such as location, shape, and color can affect both behavioral and neural responses. For instance, it has been demonstrated that neuronal responses are diminished with stimulus repetition (e.g., Brown, Wilson, & Riches, 1987; Desimone, 1996; Fahy, Riches, & Brown, 1993; for a review, see Brown & Xiang, 1998). We have emphasized Dukewich’s habituation account because it figured prominently in our rationale for the present series of experiments. Sereno, Lehky, Patel, and Pang (2010) have proposed a similar idea for interpreting both location- and feature-based repetition effects in terms of neuronal response suppression effects in neurons of the dorsal and ventral cortical visual processing streams (also see Patel, Peng, & Sereno, 2010). In line with this, recent neurophysiological (e.g., Janssen, Srivastava, Ombelet, & Orban, 2008) and neuroimaging studies (e.g., Konen & Kastner, 2008) in monkeys and in humans, respectively, have demonstrated that their proposed neurophysiological mechanism underlying behavioral inhibition may be a common property of many neural networks. How this “world of stimulus repetition” might be related to IOR, which as conceived by Posner et al. (1985), and explored during and after searching (for a review, see Wang & Klein, 2010), does not require repeated stimuli, remains to be worked out in future behavioral and neuroscientific studies.
Notes
Due to a technical problem, the first 19 participants completed 476 experimental trials rather than the originally intended 512 trials. This led to a small difference in the number of trials per condition – several conditions had about 14 rather 16 observations. This also applied to the first 19 participants from Experiment 2. For the remaining participants in both experiments, the programs with exactly 512 main trials were obtained (each cell had 16 observations). Further data analysis did not show any significant differences between the balanced and unbalanced data sets, therefore the data from all participants were retained for the analyses reported here.
For each experiment we conducted an ANOVA on RTs from the Different Location conditions with distance (90° vs 180°), CTOA and feature match vs mismatch as factors. In the analyses of Experiment 1 (detection task) and Experiment 3 (orthogonal discrimination task) there were no effects or interactions involving distance. In the analysis of Experiment 2 distance interacted with feature match vs mismatch (the same color advantage for the 180° condition, 31 ms, was larger than that for the 90° condition, 10 ms). Because this advantage did not interact with CTOA, it does not impact any of the conclusions drawn.
References
Berlucchi, G. (2006). Inhibition of return: A phenomenon in search of a mechanism and a better name. Cognitive Neuropsychology, 23(7), 1065–1074.
Brown, M. W., & Xiang, J. Z. (1998). Recognition memory: Neuronal substrates of the judgement of prior occurrence. Progress in Neurobiology, 55(2), 149–189.
Brown, M. W., Wilson, F. A. W., & Riches, I. P. (1987). Neuronal evidence that inferomedial temporal cortex is more important than hippocampus in certain processes underlying recognition memory. Brain Research, 409(1), 158–162.
Desimone, R. (1996). Neural mechanisms for visual memory and their role in attention. Proceedings of the National Academy of Sciences, 93(24), 13494–13499.
Dukewich, K. R. (2009). Reconceptualizing inhibition of return as habituation of the orienting response. Psychonomic Bulletin & Review, 16, 238–251.
Fahy, F. L., Riches, I. P., & Brown, M. W. (1993). Neuronal activity related to visual recognition memory: Long-term memory and the encoding of recency and familiarity information in the primate anterior and medial inferior temporal and rhinal cortex. Experimental Brain Research, 96(3), 457–472.
Fox, E., & de Fockert, J. W. (2001). Inhibitory effects of repeating color and shape: Inhibition of return or repetition blindness? Journal of Experimental Psychology: Human Perception and Performance, 27(4), 798–812.
Hilchey, M. D., Satel, J., Ivanoff, J., & Klein, R. M. (2013). On the nature of the delayed “inhibitory” Cuing effects generated by uninformative arrows at fixation. Psychonomic Bulletin & Review, 20(3), 593–600.
Hilchey, M. D., Klein, R. M., & Satel, J. (2014). Returning to “inhibition of return” by dissociating long-term oculomotor IOR from short-term sensory adaptation and other nonoculomotor “inhibitory” cuing effects. Journal of Experimental Psychology: Human Perception and Performance, 40(4), 1603–1616.
Hu, F. K., & Samuel, A. G. (2011). Facilitation versus inhibition in non-spatial attribute discrimination tasks. Attention, Perception, & Psychophysics, 73(3), 784–796.
Hu, F. K., Samuel, A. G., & Chan, A. S. (2011). Eliminating inhibition of return by changing salient nonspatial attributes in a complex environment. Journal of Experimental Psychology: General, 140(1), 35–50.
Hu, F. K., Fan, Z., Samuel, A. G., & He, S. (2013). Effects of display complexity on location and feature inhibition. Attention, Perception, & Psychophysics, 75(8), 1619–1632.
Hu, K., Zhan, J., Li, B., He, S., & Samuel, A. G. (2014). Multiple cuing dissociates location- and feature-based repetition effects. Vision Research, 101, 73–81.
Huber, D. E. (2014). The rise and fall of the recent past: A unified account of immediate repetition paradigms. In B. H. Ross (Ed.), Psychology of learning and motivation (Vol. 60, pp. 191–226). Amsterdam: Elsevier.
Janssen, P., Srivastava, S., Ombelet, S., & Orban, G. A. (2008). Coding of shape and position in macaque lateral intraparietal area. The Journal of Neuroscience, 28(26), 6679–6690.
Jennings, J. R., & Wood, C. C. (1976). The e-adjustment procedure for repeated measure analyses of variance. Psychophysiology, 13, 277–278.
Klein, R. M. (2000). Inhibition of return. Trends in Cognitive Sciences, 4(4), 138–147.
Konen, C. S., & Kastner, S. (2008). Two hierarchically organized neural systems for object information in human visual cortex. Nature Neuroscience, 11(2), 224–231.
Kwak, H. W., & Egeth, H. (1992). Consequences of allocating attention to locations and to other attributes. Perception & Psychophysics, 51(5), 455–464.
Lupiáñez, J. (2010). Inhibition of return. In A. C. Nobre & J. T. Coull (Eds.), Attention and time (pp. 17–34). Oxford: Oxford University Press.
Lupiáñez, J., Milán, E. G., Tornay, F. J., Madrid, E., & Tudela, P. (1997). Does IOR occur in discrimination tasks? Yes, it does, but later. Perception & Psychophysics, 59(8), 1241–1254.
Lupiáñez, J., Klein, R. M., & Bartolomeo, P. (2006). Inhibition of return: Twenty years after. Cognitive Neuropsychology, 23(7), 1003–1014.
Neill, W. T. (1978). Decision processes in selective attention: Response priming in Stroop color-word task. Perception & Psychophysics, 23(1), 80–84.
Patel, S. S., Peng, X., & Sereno, A. B. (2010). Shape effects on reflexive spatial selective attention and a plausible neurophysiological model. Vision Research, 50(13), 1235–1248.
Posner, M. I. (1980). Orienting of attention. Quarterly Journal of Experimental Psychology, 32, 3–25.
Posner, M. I., & Cohen, Y. (1984). Components of visual orienting. In H. Bouma & D. Bouwhuis (Eds.), Attention and performance (Vol 10, pp. 531–556). Hillsdale, NJ: Erlbaum.
Posner, M. I., Rafal, R. D., Choate, L. S., & Vaughan, J. (1985). Inhibition of return: Neural basis and function. Cognitive Neuropsychology, 2, 211–228.
Rankin, C. H., Abrams, T., Barry, R. J., Bhatnagar, S., Clayton, D. F., Colombo, J., … & Thompson, R. F. (2009). Habituation revisited: An updated and revised description of the behavioral characteristics of habituation. Neurobiology of Learning and Memory, 92(2), 135–138.
Samuel, A. G., & Kat, D. (2003). Inhibition of return: A graphical meta-analysis of its time course and an empirical test of its temporal and spatial properties. Psychonomic Bulletin & Review, 10(4), 897–906.
Samuel, A. G., & Weiner, S. K. (2001). Attentional consequences of object appearance and disappearance. Journal of Experimental Psychology: Human Perception and Performance, 27(6), 1433–1451.
Schneider, W., Eschmann, A., & Zuccolotto, A. (2002). E-prime user’s guide. Pittsburgh: Psychology Software Tools.
Sereno, A. B., Lehky, S. R., Patel, S., & Peng, X. (2010). A neurophysiological correlate and model of reflexive spatial attention. In N. Srinivasan, B. R. Kar, & J. Pandey (Eds.), Advances in cognitive science (Vol. 2, pp. 104–131). New Delhi: SAGE.
Sokolov, Y. N. (1960). Neuronal models and the orienting reflex. In M. A. Brazier (Ed.), The central nervous system and behavior. New York: Macy Foundation.
Sokolov, Y. N. (1963). Perception and the conditioned reflex. New York: MacMillan.
Spence, C. J., & Driver, J. (1994). Covert spatial orienting in audition: Exogenous and endogenous mechanisms. Journal of Experimental Psychology: Human Perception and Performance, 20(3), 555–574.
Spence, C., & Driver, J. (1997). Audiovisual links in exogenous covert spatial orienting. Perception & Psychophysics, 59(1), 1–22.
Tanaka, Y., & Shimojo, S. (1996). Location vs. feature: Reaction time reveals dissociation between two visual functions. Vision Research, 36(14), 2125–2140.
Taylor, T. L., & Donnelly, M. P. (2002). Inhibition of return for target discriminations: The effect of repeating discriminated and irrelevant stimulus dimensions. Perception & Psychophysics, 64(2), 292–317.
Wang, Z., & Klein, R. M. (2010). Searching for inhibition of return in visual search: A review. Vision Research, 50, 220–228.
Acknowledgments
Kesong Hu thanks Ray Klein for his traveling support to the Department of Psychology and Neuroscience, Dalhousie University, in May 2012 to make this collaborative project possible. This study was financially supported by research projects granted to Shuchang He (Natural Science Foundation of China, Grant No. 81271491/H0920), and Raymond Klein (Natural Science and Engineering Research Council of Canada, Discovery Grant). The methodological weakness (use of nonorthogonal cuing and response dimensions) of Hu and Samuel (2011) was first pointed out by R. M. K. to K. D. in e-mail correspondence in September 2011, and eventually led to this collaborative project with K. H. and his colleagues in China, where the research was conducted. We thank Arthur G. Samuel and an anonymous reviewer for helpful suggestions on an earlier draft of the paper, and all the participants for their contribution.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Appendix
Appendix
It was not our intention in this paper to provide a thorough analysis of the viability of the different frameworks for understanding IOR. It is worth noting, however, an important contrast in the scopes of these frameworks. Dukewich (2009) proposed that some forms of IOR might be spatial examples of a much more general phenomenon, habituation, which is about learning to ignore irrelevant stimuli. In contrast, Lupiáñez (2010) proposed his three-component framework to help shed light on all peripheral cuing effects. The findings from our three experiments and from those of Hu and colleagues are listed in the left-hand column of Table 1. In the remaining columns we note how we believe the two frameworks explain these findings, and we highlight where we think that they do not do so successfully.
-
(1)
Because both frameworks were developed in the context of the cross-over pattern discovered by Posner and Cohen (1984) that is so often seen in studies with uninformative peripheral cues, it is not surprising that the early facilitation of processing at the cued location and the later inhibition at this location are easily accommodated by both frameworks. Early facilitation is generated by the summation of orienting responses in the habituation framework and by the spatial orienting benefit in the three-component framework. Later costs are explained by habituation of the orienting response in the habituation framework and by onset detection cost in the three-component framework. A problem (1X) for the three-component framework, however, is our finding (which can be seen by comparing Figs. 4 and 6 and which is highlighted in Fig. 7) that the magnitude of the inhibition in our orthogonal discrimination task (Experiment 3) was not reduced relative to that in the detection task (Experiment 1). The problem arises because, as described above, according to this framework, onset detection costs play a reduced role in discrimination tasks.
-
(2)
Neither framework has a ready-made explanation for the early feature repetition benefit that was observed at both the cued and uncued locations in the nonorthogonal discrimination task (Experiment 2). Although the habituation account allows for early summation of activation in any pathway, which might generate an early feature repetition benefit, we can’t imagine how this type of summation would be restricted to the condition in which the nonspatial feature is task relevant. The “spatial selection benefit” in the three-process model accounts well for an early benefit for feature repetition at the cued location, but (and this was missed by Hu & Samuel, 2011) it specifically cannot handle such a benefit at the uncued location. The nonorthogonal nature of the cuing manipulation in this paradigm allows for cue-generated response activation, which we proposed might be responsible for this benefit. And the elimination of this benefit in the orthogonal discrimination task of Experiment 3 confirms this hypothesis. This proposal can be viewed as a “friendly” amendment to (and suggesting a mechanism external to) both frameworks.
-
(3)
Each framework can explain the finding of reduced inhibition when the colors of the cues and targets mismatch in Experiment 1. Dishabituation is the explanation that is part and parcel of the habituation framework. In the three-component model, this is explained by a decrease in onset detection cost that is associated with the change in color. Contrary to both of these mechanisms, however, identical amounts of inhibition was observed in Experiment 2. And because onset detection costs are supposed to be reduced in discrimination tasks, the similarity of the patterns of inhibition in Experiment 1 and Experiment 3 is problematic for the three-process framework.
Rights and permissions
About this article

Cite this article
Klein, R.M., Wang, Y., Dukewich, K.R. et al. On the costs and benefits of repeating a nonspatial feature in an exogenous spatial cuing paradigm. Atten Percept Psychophys 77, 2293–2304 (2015). https://doi.org/10.3758/s13414-015-0941-2
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13414-015-0941-2