
Abstract
In Experiment 1, two groups of female rats were trained in a triangular pool to find a hidden platform whose location was defined in terms of a single a landmark, a cylinder outside the pool. For one group, the landmark had only a single pattern (i.e., it looked the same when approached from any direction), while for the other, the landmark contained four different patterns (i.e., it looked different when approached from different directions). The first group learned to swim to the platform more rapidly than the second. Experiment 2 confirmed this difference when female rats were trained in a circular pool but found that male rats learned equally rapidly (and as rapidly as females trained with the single-pattern landmark) with both landmarks. This second finding was confirmed in Experiment 3. Finally, in Experiment 4a and 4b, male and female rats were trained either with the same, single-pattern landmark on all trials or with a different landmark each day. Males learned equally rapidly (and as rapidly as females trained with the unchanged landmark) whether the landmark changed or not. We conclude that male and female rats learn rather different things about the landmark that signals the location of the platform.
Similar content being viewed by others

Sex differences in spatial cognition or navigation have been observed in many animals, including humans (Mackintosh, 2011) and a variety of nonhuman species (Chamizo & Rodríguez, 2012; Coluccia & Louse, 2004; Jones, Braithwaite, & Healy, 2003). It is not just that males often learn to solve a spatial problem more rapidly than females, but that males and females may use different strategies to solve the same problem. In both rats and people, males seem more likely to rely on geometrical information to learn a route to a goal, while females are more likely to use landmarks (Choi & Silverman, 2003; Galea & Kimura, 1993; Williams & Meck, 1991). We have observed a similar difference in a number of experiments, in which rats were trained to find a hidden platform in a triangular-shaped pool; the platform was located in one corner of the pool, next to a single landmark or beacon. The rats were then given a test trial, without the platform, with these two sources of information put into opposition (the landmark was placed next to a different corner of the pool). We have consistently found that male rats tend to search in the correct corner of the pool, while females search next to the landmark (Rodríguez, Chamizo, & Mackintosh, 2011, 2013; Rodríguez, Torres, Mackintosh, & Chamizo, 2010). We should stress that these are differences in preference only: When tested with only one source of information available (either the landmark or the location), both male and female rats showed that they had learned something about both sources. Other studies have indeed shown that male rats may, in some circumstances, prefer to use the visual features of a landmark rather than its spatial location (e.g., Arain & Cohen, 2013).
In a preliminary study with the same general design, we were surprised to find that the behavior of female rats on the test trial, but not that of males, was affected by the nature of the landmark (Torres, Rodríguez, Chamizo, & Mackintosh, 2014). When the landmark was a cone, females, as usual, spent more time near the landmark, but when it was a pyramid, they spent more time in the originally correct corner of the pool. Males, on the other hand, always spent more time in the distinctive corner of the pool. In a second experiment, two identical cylinders were used as landmarks, one plain white and the other divided into four vertical segments, each patterned differently. On the test trial where the two sources of information (landmark and geometrical cue) were pitted against one another, female rats preferred the plain white cylinder to the geometrical cue, but this preference was reversed when the cylinder was divided into four different patterns.
We interpreted these findings to mean that the landmark would win out over the geometrical cue for females only when it looked the same from all perspectives. Thus, the sex difference we had observed in rats’ preferences between a landmark and geometrical information depended on the nature of the landmark. When sex differences have been explored, similar inconsistencies related to the material used have also being found in humans (Heil & Jansen-Osmann, 2008; Jansen-Osmann & Heil, 2007) using mental rotation tasks and different stimuli.
If the critical factor determining whether females relied on the landmark in preference to geometry in the study by Torres et al. (2014) was whether the landmark had the same appearance when viewed from different directions, we might have expected to see a difference between different groups of females in the speed with which they learned to use different landmarks to locate the platform during initial training: A landmark that appears to be different from one trial to the next must surely be a less effective cue to the location of the platform than one that can be seen to be the same on every trial. However, there was never any difference between groups during the course of initial training—or on the test trials with the landmark alone.
One possible solution to this apparent dilemma is to note that in all these experiments, animals always received initial training with the platform not only next to a landmark, but also in one distinctive corner of the pool. Under these circumstances, if animals found the landmark less effective, they could have used the geometrical information to locate the platform. If so, it would be necessary to train female rats with only the landmark relevant in order to detect any difference in the effectiveness of different landmarks.
The main aim of Experiment 1 of the present study was to test this possibility. Subsequent experiments explored whether males and females differed in the type of landmark that was most effective.
Experiment 1
Experiment 1 had two groups of female rats (single pattern vs. four different patterns). Animals were trained with the triangular pool and a single landmark, although the pool’s geometrical cue was rendered irrelevant by moving the platform randomly from one corner of the pool to another. For one group of 9 rats (♀4-patterns group), the landmark was a cylinder with four different vertical patterns (plain white, vertical lines, horizontal lines, and dots), while the second group (♀1-pattern group) was divided into three subgroups of 3 rats, 1 trained with the landmark entirely covered with vertical lines, 1 with horizontal lines, and the third with dots. Would the 1-pattern group learn faster?
Method
Subjects
The subjects were 18 naive female Long Evans rats, approximately 3 months old at the beginning of the experiment. The animals were housed in standard cages, 25 × 15 × 50 cm, in groups of 2 and were maintained on ad lib food and water, in a colony room with a 12:12-h light:dark cycle. They were tested within the first 8 h of the light cycle.
Apparatus
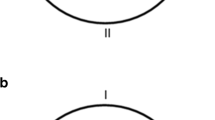
The apparatus was a circular swimming pool made of plastic and fiberglass and modeled after that used by Morris (1981). It measured 1.58 m in diameter and 0.65 m deep, and it was filled to a depth of 0.49 m, with water rendered opaque by the addition of 1 cl/l of latex. The water temperature was maintained at 22 + 1 °C. The pool was situated in the middle of a large room and mounted on a wooden platform 0.43 m above the floor. To create the triangular geometry, two acrylic boards forming an angle of 90° were inserted in the pool resting on platforms at the base, which supported them vertically. The boards were 39.5 cm high, 0.5 cm thick, and 112 cm long. The top of the boards was 9.5 cm above the water surface—that is, at the same height as the outer wall of the pool. The pool was surrounded by black curtains reaching from the ceiling to the base of the pool and forming a circular enclosure 2.4 m in diameter. A single object, landmark X, was suspended from a black false ceiling inside this enclosure, 35 cm above the surface of the water and with its midline directly above the wall of the pool. For one group of subjects, landmark X was always a cylinder, 8.5 cm in diameter and 30 cm in height, with four consecutive vertical segments, each 7 cm wide, with different patterns: One was plain white, one had four black horizontal lines (2 cm thick each line), one had eight black dots mixed up on the white surface (2-cm diameter each dot), and the fourth one had two vertical black lines (2 cm thick each line). The second group was divided into three subgroups: For a third of the rats, the cylinder was entirely patterned with vertical lines; for a second third, with dots; and for the final animals, with horizontal lines. The single landmark X was the only cue defining the location of the platform. A closed-circuit video camera with a wide-angle lens was mounted 1.75 m above the center of the pool inside the false ceiling, and its picture was relayed to recording equipment in an adjacent room. A circular platform 0.11 m in diameter and made of transparent Perspex was mounted on a rod and base that was placed 0.38 m from the point formed by any of the three corners of the pool, on a line that bisected the center of the pool, with its top 1 cm below the surface of the water. The hidden platform, P, landmark X, and the geometry of the pool were situated as shown in Fig. 1, top panel left.

Top: A schematic representation of the pool and the position of the landmark, X, and the hidden platform (P) during Experiment 1. Middle: A schematic representation of the pool and the position of the landmark, X, and the hidden platform (P) during Experiments 2 and 4. Bottom: A schematic representation of the pool and the position of the landmark, X, and the hidden platform (P) during Experiment 3. I, II, III, and IV indicate the four starting positions
Procedure
There were two types of trials: pretraining and training. Pretraining consisted of placing a rat into the circular pool without the landmark or boards, but with the hidden platform present. The rat was given 120 s to find the platform, and once the rat had found it, it was allowed to stay on it for 30 s. If it had not found the platform within the 120 s, it was picked up, placed on it, and left there for 30 s. The platform was moved from one trial to the next, and the rat was placed in the pool in a different location on each trial, as far as possible equally often on the same or opposite side of the pool from the platform, and with the platform to the right or to the left of where the rat was placed. Rats were given five such pretraining trials over 2 days, with two trials on day 1 and three on day 2. Rats were run in squads of eight and spent the intertrial interval (ITI; approximately 8–10 min) in small individual compartments.
The procedure for training was similar to that of pretraining, with two exceptions. The landmark, X (either of the two cylinders), was always present, as well as the two boards forming the triangular pool. In order to ensure that the rats used this landmark, X, as the source of information to locate the platform, rather than any inadvertently remaining static room cues (like noises from pipes and air conditioning), the landmark and the platform were semirandomly rotated with respect to the room (90°, 180°, 270°, or 360°), with the restriction that all four positions of the room were used each day. As in pretraining, the rat was placed in the pool in a different location on each trial, as far as possible equally often with the platform to the right, to the left, or in front of where the rat was placed (at I, II, III, IV of Fig. 1, top). Rats were given 8 trials per day over 3 days (a total of 24 trials). These trials had an ITI of 8–10 min, and the platform and the landmark were rotated between trials. Importantly, during training, the platform was randomly moved between the three corners of the triangular-shaped pool to prevent a permanent association between it and the geometry of the pool, as shown in Fig. 1, top panel. Only the landmark X maintained a fixed relationship with the position of the platform.
Results and discussion
Latencies (SEMs) to find the platform decreased over the course of the five initial pretraining trials. The 4-patterns group decreased from a mean of 119.4 s (0.60) on trial 1 to a mean of 44.4 s (11.20) on trial 5, and the 1-pattern group from a mean of 85.2 s (12.52) on trial 1 to a mean of 38.9 s (8.79) on trial 5. An ANOVA conducted on these data taking into account the variables trials (1–5) and group (4-patterns, 1-pattern) showed that the only significant variable was trials, F(4, 64) = 9.02, p < .01, MSE = 11,263.72, η 2 p = .36. No other main effect or interaction was significant (Fs < 1.5).
Latencies (SEMs) to find the platform also decreased over the course of the training days (see Fig. 2). An ANOVA conducted on these data taking into account the variables days (1–3) and group (4-patterns, 1-pattern) showed that the variables days, F(2, 32) = 66.91, p < .01, MSE = 2,864.80, η 2 p = .81, and group, F(1, 16) = 18.00, p < .01, MSE = 1,500.16, η 2 p = .53, were significant, as well as the days × group interaction, F(2, 32) = 5.68, p = .008, MSE = 243.02, η 2 p = .26. The analysis of the interaction showed that females in the 1-pattern group reached the platform faster than females in the 4-patterns group on all three training days, [Fs(1, 16) =11.82, (p = .003, MSE = 1,465.75, η 2 p = .42), 11.95 (p = .003, MSE = 469.61, η 2 p = .43), and 9.01 (p = .008, MSE = 50.84, η 2 p = .36) on days 1, 2, and 3, respectively].

Mean escape latencies for the groups of Experiment 1. Error bars denote standard errors of the means
Experiment 2
The results of Experiment 1 showed that females learned more rapidly when the landmark had only a single pattern (and so looked the same when approached from any direction) than when the landmark contained four different patterns (and so looked different when approached from different directions). However, one could argue that because the procedure used in Experiment 1 was not optimal, these deficiencies could, somehow, be affecting our results. Specifically, the two groups in this experiment were given the opportunity to learn not only that the landmark cue signaled the location of the platform, but also that the other class of cue, the geometry of the pool, was irrelevant (i.e., a case of “learned irrelevance”; for a demonstration, see Mackintosh, 1973). To prevent this possibility, Experiment 2 (as well as the remaining experiments in this article) was conducted in a circular pool.
In Experiment 2, four groups of rats (two groups of males and two groups of females; single pattern vs. four different patterns in each sex) were trained in the presence of a single landmark to provide the needed within-sex comparison. For 24 rats, 12 males and 12 females, the landmark was the cylinder with four different patterns (4-patterns group), while the remaining 24 rats, 12 males and 12 females, were divided into three subgroups of four rats of each sex, one trained with the landmark entirely covered with vertical lines, one with horizontal lines, and the third with dots (1-pattern group; the plain white cylinder was not used, to avoid a greater contrast between this landmark and the background, the black curtains, than between the patterned cylinders and the background, as addressed in Torres et al., 2014, Experiment 3). On the basis of the results of Experiment 1 and those of Torres et al., we predicted that females would learn faster when trained with a single pattern landmark but that males would learn equally rapidly whether trained with a single pattern or a 4-pattern landmark.
Method
Subjects and apparatus
The subjects were 48 naive Long Evans rats, 24 males and 24 females, approximately 3 months old at the beginning of the experiment. The animals were kept and maintained as in Experiment 1.
Procedure
The general procedure was the same as that used in Experiment 1, with the exception that during training, the pool maintained the original circular shape instead of the triangular shaped pool used in Experiment 1 (as shown in Fig. 1, middle panel).
Results and discussion
Latencies (SEMs) to find the platform decreased over the course of the five initial pretraining trials. In the 4-patterns group, males decreased from a mean of 105.4 s (5.65) on trial 1 to a mean of 53.6 s (12.87) on trial 5, and females from a mean of 102.7 s (8.67) on trial 1 to a mean of 49.4 s (12.4) on trial 5. In the 1-pattern group, males decreased from a mean of 94.8 s (10.83) on trial 1 to a mean of 42.8 s (10.09) on trial 5, and females from a mean of 90.7 s (11.99) on trial 1 to a mean of 44.1 s (12.09) on trial 5. An ANOVA conducted on these data taking into account the variables trials (1–5), group (4-patterns, 1-pattern), and sex showed that the only significant variable was trials, F(4, 176) = 17.14, p < .01, MSE = 21,395.57, η 2 p = .28. No other main effect or interaction was significant (Fs < 2.0).
Latencies (SEMs) to find the platform also decreased over the course of the training days (see Fig. 3). An ANOVA conducted on these data taking into account the variables days (1–3), group (4-pattern, 1-pattern), and sex showed that the variables days, F(2, 88) = 90.64, p < .01, MSE = 2,325.60, η 2 p = .67, group, F(1, 44) = 25.17, p < .01, MSE = 1,444.25, η 2 p = .36, and sex, F(1, 44) = 13.14, p = .001, MSE = 754.14, η 2 p = .23, were all significant, as well as the days × group, F(2, 88) = 10.43, p < .01, MSE = 267.58, η 2 p = .19, days × sex, F(2, 88) = 5.88, p = 0.004, MSE = 150.97, η 2 p = .12, and group × sex, F(1, 44) = 7.27, p = .010, MSE = 417.18, η 2 p = .14, interactions. The triple days × group × sex interaction was also significant, F(2, 88) = 5.81, p = .004, MSE = 149.06, η 2 p = .12. To address the triple interaction, one independent ANOVA was conducted each day to see whether the group × sex interaction was significant. These analyses revealed that the group × sex interaction was significant on day 1 only, F(1, 44) = 11.31, p = .002, MSE = 649.37, η 2 p = .20. The analysis of the group × sex interaction on day 1 revealed that females in the 1-pattern group reached the platform faster than females in the 4-pattern group, F(1, 22) = 30.77, p < .01, MSE = 2,137.22, η 2 p = .58, while the two groups of male rats did not differ (Fs < 2.5). In addition, in the 4-pattern group, males reached the platform faster than females, F(1, 22) = 23.52, p < .01, MSE = 1,368.97, η 2 p = .52, while males and females did not differ in the 1-pattern group (Fs < 0.5).

Mean escape latencies for the groups of Experiment 2. Error bars denote standard errors of the means
Experiment 3
Experiment 3 was designed to see whether the males’ previous null result could be replicated under slightly different conditions. The experiment had two groups of male rats (single pattern vs. four different patterns). Animals were trained with the circular pool and a single landmark. For one group of 9 rats (♂4-patterns group), the landmark was the previous cylinder with four different patterns, while the second group (♂1-pattern group) was divided into three subgroups of 3 rats, 1 trained with the landmark entirely covered with vertical lines, 1 with horizontal lines, and the third with dots. In this experiment, the platform was placed slightly closer to the center of the pool, as compared with Experiment 2, to facilitate the vision of the landmark, in an attempt to obtain a difference between the two groups. Under these new conditions, would the 1-pattern group learn faster?
Method
Subjects and apparatus
The subjects were 18 naive male Long Evans rats, approximately 3 months old at the beginning of the experiment. The animals were kept and maintained as in Experiments 1 and 2.
Procedure
The general procedure was the same as that used in Experiment 2, with the exception that the platform was placed 10 cm farther from the wall of the pool (i.e., 48 cm instead of 38 cm) in order to enhance the visibility of the landmark (as shown in Fig. 1, bottom panel).
Results and discussion
Latencies (SEMs) to find the platform decreased over the course of the five initial pretraining trials. The 4-patterns group decreased from a mean of 84.9 s (13.23) on trial 1 to a mean of 44.4 s (12.28) on trial 5, and the 1-pattern group from a mean of 90.3 s (14.52) on trial 1 to a mean of 49.6 s (13.95) on trial 5. An ANOVA conducted on these data taking into account the variables trials (1–5) and group (4-patterns, 1-pattern) showed that the only significant variable was trials, F(4, 64) = 4.64, p = .002, MSE = 6,755.95, η 2 p = .22. No other main effect or interaction was significant (Fs < 1.0).
Latencies (SEMs) to find the platform also decreased over the course of the training days (see Fig. 4). An ANOVA conducted on these data taking into account the variables days (1–3) and group (4-patterns, 1-pattern) showed that the only significant variable was days, F(2, 32) = 17.61, p < .01, MSE = 980.65, η 2 p = .52. No other main effect or interaction was significant (Fs < 0.5).

Mean escape latencies for the groups of Experiment 3. Error bars denote standard errors of the means
Experiment 4a
In Experiment 4a, we asked whether presenting different landmarks during training (although always maintaining the same or a very similar position relative to the platform) would affect female rats more than males. The results of Torres et al. (2014) clearly revealed that a landmark cue will win out over a geometrical cue (the shape of a triangular pool) in female rats only when it looks the same from all perspectives. Therefore, the specific characteristics of a landmark cue seem to play a crucial role in females’, but not males’, preference when solving a spatial task. Experiment 4 provided a more rigorous test of this suggestion. The experiment required four groups of animals, two groups of 12 males each and two groups of 12 females each. A set of males and another set of females (Same-L group) were trained with one and the same landmark (single pattern; specifically, the cylinder entirely covered with dots), while a second set of males and a second set of females (Diff-Ls group) were trained with changing landmarks (all of them, eight in total, single-pattern landmarks that looked the same when approached from any direction; most of these landmarks had been used in previous experiments in our laboratory and, when used as beacons, were all equally effective—i.e., did not lead to differences during acquisition). Considering our previous results (Torres et al., 2014), we expected that the two groups of females would differ, with group Same-L reaching the platform significantly faster than group Diff-Ls. No differences were expected in the two groups of male rats that should perform as well as the females of group Same-L.
Method
Subjects and apparatus
The subjects were 48 naive Long Evans rats, 24 males and 24 females, approximately 3 months old at the beginning of the experiment. The animals were kept and maintained as in the previous experiments. They were divided into two groups with equal number of males and females (12 males and 12 females per group), Same-L and Diff-Ls. For the group Same-L the landmark X was always the same, the cylinder patterned with black dots (like that used in the previous experiments). The group Diff-Ls had eight different landmarks: the cylinder dots, the cylinder with vertical lines (identical to that used in the previous experiments), a ninepin (6 cm in diameter at the base and 16.5 cm in height, with the wider part measuring 26 cm in circumference, with blue and yellow segments), a ball (32 cm in diameter and mixed colors), an inverted cone (16 cm in diameter and 59 cm in height, with 1-cm-thick black horizontal stripe spaced 3.5 cm apart), three mopheads attached together, a vertical cone (16 cm in diameter and 59 cm in height, with 1-cm-thick black horizontal stripe spaced 3.5. cm apart and 1-cm-thick black vertical stripe spaced 3.5 cm apart), and three inverted white pots, varying in size from 13.5 cm in diameter and 9 cm in height to 20 cm in diameter and 12 cm in height and 26 cm in diameter and 20 cm in height. The three pots were tied together with the smallest pot at the bottom and the largest at the top.
Procedure
The general procedure was the same as that used in Experiments 2 and 3, with the exception that the cylinder with black dots was presented on all trials for group Same-L, while for group Diff-Ls, the eight different landmarks described above were presented as follows: The inverted cone, the ninepin, the cylinder with vertical lines, and the three mopheads attached together were presented twice (in a semirandom order) on days 1 and 3, while the cylinder with black dots, the vertical cone, the ball, and the three inverted pots were presented (in a semirandom order) on day 2 only.
Results and discussion
Latencies (SEMs) to find the platform decreased over the course of the five initial pretraining trials. In group Same-L, males decreased from a mean of 83.6 s (10.09) on trial 1 to a mean of 43.1 s (12.22) on trial 5, while females decreased from a mean of 94.0 s (12.80) on trial 1 to a mean of 74.5 s (13.38) on trial 5. In group Diff-Ls, males decreased from a mean of 98.1 s (11.24) on trial 1 to a mean of 40.6 s (9.82) on trial 5, while females decreased from a mean of 84.5 s (12.45) on trial 1 to a mean of 45.3 s (11.98) on trial 5. An ANOVA conducted on these data taking into account the variables trials (1–5), group (Same-L, Diff-Ls), and sex showed that the only significant variable was trials, F(4, 176) = 6.49, p < .01, MSE = 9,748.42, η 2 p = .13. No other main effect or interaction was significant (Fs < 1.5).
Latencies (SEMs) to find the platform also decreased over the course of the training days (see Fig. 5, top). An ANOVA conducted on these data taking into account the variables days (1–3), group (Same-L, Diff-Ls), and sex showed that the variables days, F(2, 88) = 34.20, (p < .01, MSE = 2,884.77, η 2 p = .44, group, F(1, 44) = 5.34, p = .026, MSE = 1,034.96, η 2 p = .11, and sex, F(1, 44) = 20.74, p < .01, MSE = 4,020.09, η 2 p = .31, were significant, as well as the days ×group, F(2, 88) = 3.97, p = .022, MSE = 335.27, η 2 p = .07, days × sex, F(2, 88) = 5.52, p = .005, MSE = 466.00, η 2 p = .10, and group × sex, F(1, 44) = 8.15, p = .007, MSE = 1,580.13, η 2 p = .16, interactions. The triple days × group × sex interaction was also significant, F(2, 88) = 3.16, p = .047, MSE = 266.83, η 2 p = .16. To address the triple interaction, one independent ANOVA was conducted each day to see whether the group × sex interaction was significant. These analyses revealed that the group × sex interaction was significant on days 1 and 2 only, Fs(1, 44) = 5.03 (p = .030, MSE = 1,196.5, η 2 p = .10) and 8.40 (p = .006, MSE = 898.99, η 2 p = .16), respectively. On both day 1 and day 2, males reached the platform faster than females in group Diff-Ls, Fs(1, 22) = 11.57 (p = .003, MSE = 4,060.68, η 2 p = .34) and 15.33 (p = .001, MSE = 2,533.4, η 2 p = .41), respectively, while the two sexes did not differ in group Same-L, Fs < 2.0 and 1.5, respectively. Moreover, females in group Same-L reached the platform faster than females in group Diff-Ls, Fs(1, 22) = 5.64 (p = .027, MSE = 2,394.2, η 2 p = .20) and 7.74 (p = .011, MSE = 1,376.83, η 2 p = .26), respectively, while males in the two groups learned equally rapidly (Fs < 0.0 and 1.0, respectively).

Experiment 4b
A problem in Experiment 4a is that the same landmark (i.e., the cylinder with black dots) was used for all the rats in group Same-L. It could be argued that this particular landmark had a greater salience than the average effectiveness of the eight landmarks in group Diff-Ls, which could have affected the results of the experiment. The aim of Experiment 4b, only with female rats, was to address this possibility. The experiment required two groups of 8 animals each, all of them trained with one and the same single-pattern landmark. One group (Same-L group) was trained with the cylinder entirely covered with dots (thus, it was identical to group Same-L in Experiment 4a), while rats in the second group (Same-8Ls group) were trained each of them with a different landmark (and the eight landmarks used in group Diff-Ls in Experiment 4a were employed). No difference between the two groups would indicate that the single landmark in group Same-L is not of particularly high salience.
Method
Subjects and apparatus
The subjects were 16 naive female Long Evans rats approximately 4 months old at the beginning of the experiment. The animals were kept and maintained as in the previous experiments. They were divided into two groups (8 females per group), Same-L and Same-8Ls. Every rat was trained with a single landmark. For group Same-L, the landmark, X, was the same for all subjects (i.e., the cylinder patterned with black dots, as in group Same-L in Experiment 4a). For group Same-8Ls, the landmark, X, was a different object for each rat (i.e., the cylinder dots, the cylinder with vertical lines, the ninepin, the ball, the inverted cone, the three mopheads attached together, the vertical cone, and the three inverted white pots—those used in Experiment 4a for group Diff-Ls). Would the two groups differ?
Procedure
The general procedure was the same as that used for group Same-L in Experiment 4a, with the exception that each rat in group Same-8Ls was trained with a different landmark (i.e., those used in group Diff-Ls in Experiment 4a), instead of all rats being trained with one and the same landmark, the cylinder dots.
Results and discussion
Latencies (SEMs) to find the platform decreased over the course of the five initial pretraining trials. The Same-L group decreased from a mean of 120.0 s (0) on trial 1 to a mean of 95.2 s (9.85) on trial 5, while the Same-8Ls group decreased from a mean of 120 s (0) on trial 1 to a mean of 98.8 s (10.35) on trial 5. An ANOVA conducted on these data taking into account the variables trials (1–5) group (Same-L, Same-8Ls), and sex showed that no main effect or interaction was significant (Fs < 1.0).
Latencies (SEMs) to find the platform also decreased over the course of the training days (see Fig. 5, bottom). An ANOVA conducted on these data taking into account the variables days (1–3) and group (Same-L, Same-8Ls) showed that the only significant variable was days, F(2, 28) = 81.50, p < .01, MSE = 11,044.93, η 2 p = .85. No other main effect or interaction was significant (Fs < 1.0).
The results of Experiment 4b challenge the notion that the results of Experiment 4a are a consequence of the special salience of the object used as the single landmark. In conclusion, Experiment 4 shows that different aspects or components of a landmark can control navigation in male and female rats, with position being crucial for males, while pattern or specific features are crucial for females (for a related result in domestic chicks, see Vallortigara, 1996). Future research should address these issues.
General discussion
In Experiments 1 and 2, when female rats were trained to find a hidden platform whose location in the pool was indicated only by a landmark next to the platform, they learned significantly faster when the landmark was covered in only a single pattern than when it had four different patterns. These results contrast sharply with those reported by Torres et al. (2014), where, although the preference test showed that females preferred the landmark to the geometrical cue only when the landmark had a single pattern, females trained with the single pattern learned to find the platform no more rapidly than did those trained with the 4-pattern landmark. As was noted above, the most plausible explanation of this difference is that in the Torres et al. experiments, the platform was always located in one distinctive corner of the triangular pool, as well as next to a landmark, while in the present experiments, only the landmark signaled the location of the platform. The addition of the geometrical cue in the Torres et al. experiments may have potentiated learning about the landmark and, thus, may have obscured any difference in learning about the single-pattern and 4-pattern landmarks. Potentiation, rather than overshadowing, has frequently been observed in experiments on spatial learning in the Morris pool (Cole, Gibson, Pollack, & Yates, 2011; Graham, Good, McGregor, & Pearce, 2006; Horne & Pearce, 2011; McGregor, Horne, Esber, & Pearce, 2009; Pearce, Graham, Good, Jones, & McGregor, 2006).
Be this as it may, the present results add to those reported by Torres et al. (2014) in demonstrating that females rats are more likely to rely on the single-pattern landmark than on the 4-pattern landmark to locate the hidden platform in the pool. Why should this be? It will not to do say that this is because a cylinder covered with a single pattern is a more salient cue than one covered with four different patterns; the only grounds for saying that the cylinders differ in salience are that one is learned about more rapidly than the other. Difference in two cues’ salience can explain differences in what is learned about them only when there are independent grounds for claiming that they differ in salience—as, for example, when they differ in intensity. In the present case, we suggest that the explanation is relatively simple: A landmark that has the same appearance when approached from different directions will be more effective than one whose appearance differs, for example, if it is approached from the north or from the south. This may not matter very much in most maze experiments, where the animal’s movements are constrained by the structure of the maze, but it is surely important in the real world and is also likely to be important when rats are trained to find a hidden platform in a pool, since they are put into the pool at different points on its circumference on different trials and, once in the pool, are free to swim anywhere. It seems very probable that they will approach the platform from different directions on different trials.
In an attempt to provide evidence for this supposition, we performed a further analysis of the training results of Experiment 2 (males and females of group 4-Patterns). The pool can be divided into four imaginary quadrants: one centered on the platform, one to the left of the platform quadrant, one to its right, and one opposite. We have a record of the trajectory taken by each animal on every training trial, which made it possible to record the quadrant from which each animal entered the platform quadrant for the last time—that is, when it ended the trial by finding the platform. Ignoring the small number of trials on which a rat was placed into the pool in the platform quadrant and never left it and the equally small number of trials on which they entered the platform quadrant from the opposite quadrant (which required them to swim straight through the point at which the two quadrants touched), we were left with an average of 20.3 trials per rat on which they made their final entry into the platform quadrant either from the quadrant to its left or from the quadrant to its right. Of the total of 24 rats, 1 female entered the platform quadrant on 84 % of trials from the left, 1 male on 72 % of trials from the left, and another male on 72 % of trials from the right. No other rat entered the platform quadrant on more than 70 % of trials from the same side. The remaining 10 males averaged 57.4 % of their entries from their preferred side (whether this was the left or the right), and the remaining 11 females averaged 56.2 % of their entries from their preferred side (again whether this was the left or the right). Thus, it is clear that the large majority of rats made their final entry into the platform quadrant more or less equally often from the right and from the left. We do not believe any statistical analysis is needed to draw the conclusion that both male and female rats approached the platform from different directions on different trials.
Our suggestion, then, is that a landmark that looks the same when approached from different directions will be more effective than one whose appearance changes when approached from different directions. With the advantage of hindsight, this may not seem very surprising, but the nearest we can find to an earlier statement of this idea is in some papers by Vallortigara and his colleagues (e.g., Pecchia & Vallitorgara, 2012). They have written, for example, of the “advantage in both a visual discrimination task . . . and a visuo-spatial re-orientation task . . . from perceiving the stimuli in the scene from highly stereotyped vantage points” (Pecchia, Gagliardo, & Vallortigara, 2011, p. 6).
More important, this suggestion still leaves one unanswered question: Why did the nature of the landmark affect only the behavior of females, and not that of males? In the Torres et al. (2014) experiments, males always relied on geometry in preference to the landmark when the two cues were put in opposition. In Experiments 1 and 3 here, they learned about the 4-pattern cylinder as rapidly as about one with a single pattern. We can only suggest that male rats, being somewhat less likely than females to rely on landmarks to solve a spatial problem, do not attend very closely to the specific features of any landmark; in the present case, they simply learned that the platform was to be found next to the only large object suspended above the pool. Further research would be needed to test this idea.
References
Arain, M., & Cohen, J. (2013). Hierarchical use of cues in the missing object recognition task by rats. Behavioural Processes, 97, 41–52.
Chamizo, V. D., & Rodríguez, C. A. (2012). Qualitative sex differences in spatial learning. In S. P. McGeown (Ed.), Psychology of gender differences (pp. 267–281). Hauppauge, NY: Nova Science Publishers, Inc.
Choi, J., & Silverman, I. (2003). Processes underlying sex differences in route-learning strategies in children and adolescents. Personality and Individual Differences, 34, 1153–1166.
Cole, M. R., Gibson, L., Pollack, A., & Yates, L. (2011). Potentiation and overshadowing of shape by wall color in a kite-shaped maze using rats in a foraging task. Learning and Motivation, 42, 99–112.
Coluccia, E., & Louse, G. (2004). Gender differences in spatial orientation: A review. Journal of Environmental Psychology, 24, 329–340.
Galea, L. A. M., & Kimura, D. (1993). Sex differences in route learning. Personality and Individual Differences, 14, 53–65.
Graham, M., Good, M. A., McGregor, A., & Pearce, J. P. (2006). Spatial learning based on the shape of the environment is influenced by properties of the objects forming the shape. Journal of Experimental Psychology: Animal Behavior Processes, 32, 44–59.
Heil, M., & Jansen-Osmann, P. (2008). Sex differences in mental rotation with polygons of different complexity: Do men utilize holistic processes whereas women prefer piecemeal ones? Quarterly Journal of Experimental Psychology, 61, 683–689.
Horne, M. R., & Pearce, J. M. (2011). Potentiation and overshadowing between landmarks and environmental geometric cues. Learning & Behavior, 39, 371–382.
Jansen-Osmann, P., & Heil, M. (2007). Suitable stimuli to obtain (no) gender differences in the speedof cognitive processes involved in mental rotation. Brain and Cognition, 64, 217–227.
Jones, C. M., Braithwaite, V. A., & Healy, S. D. (2003). The evolution of sex differences in spatial ability. Behavioral Neuroscience, 117, 403–411.
Mackintosh, N. J. (1973). Stimulus selection: Learning to ignore stimuli that predict no change in reinforcement. In R. A. Hinde & J. S. Hinde (Eds.), Constraints on learning (pp. 75–96). London: Academic Press.
Mackintosh, N. J. (2011). IQ and human intelligence (2nd ed.). Oxford: Oxford University Press.
McGregor, A., Horne, M. R., Esber, G. R., & Pearce, J. M. (2009). Absence of overshadowing between a landmark and geometric cues in a distinctively shaped environment: A test of Miller and Shettleworth (2007). Journal of Experimental Psychology: Animal Behavior Processes, 35, 357–370.
Morris, R. G. M. (1981). Spatial localization does not require the presence of local cues. Learning and Motivation, 12, 239–260.
Pearce, J. M., Graham, M., Good, M. A., Jones, P. M., & McGregor, A. (2006). Potentiation, overshadowing and blocking of spatial learning based on the shape of the environment. Journal of Experimental Psychology: Animal Behavior Processes, 32, 201–214.
Pecchia, T., Gagliardo, A., & Vallortigara, G. (2011). Stable panoramic view facilitates snapshot like memories for spatial reorientation in homing pigeons. PLoS One, 6, e22657.
Pecchia, T., & Vallortigara, G. (2012). Spatial reorientation by geometry with freestanding objects and extended surfaces: A unifying view. Proceedings of the Royal Society B, 279, 2228–2236.
Rodríguez, C. A., Chamizo, V. D., & Mackintosh, N. J. (2011). Overshadowing and blocking between landmark learning and shape learning: The importance of sex differences. Learning & Behavior, 39, 324–335.
Rodríguez, C. A., Chamizo, V. D., & Mackintosh, N. J. (2013). Do hormonal changes that appear at the onset of puberty determine the strategies used by female rats when solving a navigation task? Hormones and Behavior, 64, 122–135.
Rodríguez, C. A., Torres, A. A., Mackintosh, N. J., & Chamizo, V. D. (2010). Sex differences in the strategies used by rats to solve a navigation task. Journal of Experimental Psychology: Animal Behavior Processes, 36, 395–401.
Torres, M. N., Rodríguez, C. A., Chamizo, V. D., & Mackintosh, N. J. (2014). Landmark vs. geometry learning: Explaining female rasts’ selective preference for a landmark. Psicológica, 35, 81–100.
Vallortigara, G. (1996). Learning of colour and position cues in domestic chicks: Males are better at position, females at colour. Behavioural Processes, 36, 289–296.
Williams, C. L., & Meck, W. H. (1991). The organizational effects of gonadal steroids on sexually dimorphic spatial ability. Psychoneuroendocrinology, 16, 155–176.
Acknowledgments
This research was supported by a grant from the Spanish ‘Ministerio de Ciencia e Innovación’ (Ref. PSI2010-20424) to V. D.Chamizo.
Author information
Authors and Affiliations
Corresponding authors
Rights and permissions
About this article

Cite this article
Chamizo, V.D., Rodríguez, C.A., Torres, I. et al. What makes a landmark effective? Sex differences in a navigation task. Learn Behav 42, 348–356 (2014). https://doi.org/10.3758/s13420-014-0152-z
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13420-014-0152-z