
Abstract
The rapid orienting of attention to potential threats has been proposed to proceed outside of top-down control. However, paradigms that have been used to investigate this have struggled to separate the rapid orienting of attention (i.e. capture) from the later disengagement of focal attention that may be subject to top-down control. Consequently, it remains unclear whether and to what extent orienting to threat is contingent on top-down goals. The current study manipulated the goal-relevance of threat distractors (spiders), whilst a strict top-down attentional set was encouraged by presenting the saliently colored target and the threat distracter simultaneously for a limited time. The goal-relevance of threatening distractors was manipulated by including a spider amongst the possible target stimuli (Experiment 1: spider/cat targets) or excluding it (Experiment 2: bird/fish targets). Orienting and disengagement were disentangled by cueing attention away from or towards the threat prior to its onset. The results indicated that the threatening spider distractors elicited rapid orienting of attention when spiders were potentially goal-relevant (Experiment 1) but did so much less when they were irrelevant to the task goal (Experiment 2). Delayed disengagement from the threat distractors was even more strongly contingent on the task goal and occurred only when a spider was a possible target. These results highlight the role of top-down goals in attentional orienting to and disengagement from threat.
Similar content being viewed by others

An organism’s odds of survival can be increased by focusing the limited-capacity processing resources of the visual system on potential threats in the environment (Öhman & Mineka, 2001; Tamietto & De Gelder, 2010; West, Anderson, & Pratt, 2009). For example, if a visual scene contained both a potentially poisonous spider and a harmless butterfly, orienting attention to the spider would facilitate its processing and allow us to take appropriate actions. According to evolutionary models of threat processing, such prioritized processing of potential threats is largely stimulus-driven and reliant on a fast, subcortical pathway that is unaffected by cognitive influences such as current task goals or intentions (LeDoux, 1996; Öhman & Mineka, 2001).
One potential issue is that such an encapsulated threat system could conflict with the allocation of attention in accordance with an organism’s immediate top-down goals and may even endanger survival (Chun, Golomb, & Turk-Browne, 2011; Mathews & Mackintosh, 1998; Mogg & Bradley, 1998; Yiend, 2010). For example, while you are driving, the reflexively orienting of attention to a spider on your leg and the goal-contingent orienting of attention to a traffic light that turns red at exactly the same time would be at odds with each other. Under conditions of stimulus competition (i.e., when multiple stimuli vie for attention), current theories of attention propose that orienting is the convergent result of goal-driven (i.e., top-down) and stimulus-driven (i.e., bottom-up) processes (Desimone & Duncan, 1995; Wolfe, 1994). For example, according to the Guided Search model, salient stimuli can guide orienting but be further up- or down-modulated depending on their relevance to current top-down goals (Wolfe, 1994). Given that the allocation of attention is strongly determined by top-down goals (Ansorge, Kiss, & Worschech, 2010; Awh, Matsukura, & Serences, 2003; Folk, Remington, & Johnston, 1992; Serences et al., 2005; Wolfe, Butcher, Lee, & Hyle, 2003), this raises the question of whether top-down task-goals can also affect orienting to threat.
To date, it is not clear whether prioritized orienting to threat is purely automatic, as claimed by the evolutionary account, or whether it is susceptible to cognitive processes, as would be more in line with current theories of visual search (even though such theories do not explicitly model attention to threat). Previous studies may seem to suggest that threat stimuli are prioritized for attention (over neutral stimuli) regardless of their relevance to the current task goal (Lipp & Waters, 2007; Mogg & Bradley, 1998). For example, goal-relevant threat targets (snakes) are detected more rapidly than neutral targets (frogs and flowers) in visual search tasks (LoBue & DeLoache, 2008, 2011), and goal-irrelevant threat distractors (snakes, spiders) also slow target responses relative to neutral distractors (e.g., butterflies and leaves, Devue, Belopolsky and Theeuwes, 2011; lizards and cockroaches, Lipp & Waters, 2007). Furthermore, studies using the dot-probe task have demonstrated that goal-irrelevant threat distractors that preceded a neutral target (irrelevant cue), speed responses when the threat distractor and target are presented in the same location and slow responses when presented in a different location (Koster, Crombez, & Verschuere, & De Houwer, 2004). Such results indicate that threat distractors can compel attention irrespective of the current task goals (i.e., when they are goal-relevant and goal-irrelevant).
An issue in previous studies is that they often struggle to distinguish early attentional orienting (i.e., capture) and later processes that commence after the selection of a stimulus (i.e., disengagement of focal attention; Yiend, 2010). Very few studies have been able to provide evidence in support of rapid orienting to threat (Stein, Seymour, Hebart, & Sterzer, 2014; West, Anderson, & Pratt, 2009). Delays in attentional disengagement have been observed more readily, mainly in studies employing the dot-probe paradigm with a neutral baseline (Fox, Russo, & Dutton, 2002). For example, Koster and colleagues (2004) showed that the time to locate a neutral target (dot) was longer when the target followed in the opposite location from a threat distractor (high threat IAPS picture) than when it appeared on a trial with neutral distractors only (non-threatening IAPS pictures). One issue with this approach is that attention is at fixation at the beginning of each trial and thus the threat will need to attract attention before it can influence disengagement. Belopolsky, Devue, and Theeuwes (2011) eliminated such a confound by presenting the (threat-) distractors at fixation and requiring an eye-movement to be made away from fixation to a target in the periphery. As it is unclear to what extend people engage with a stimulus at fixation, Rudaizky, Basanovic, and MacLeod (2014) have furthered this issue by using an exogenous cue to direct attention towards or away from the threat distractor and observed evidence for both rapid orienting towards and delayed disengagement from threats.
Disentangling the orienting and disengagement of attention is important, because both processes may differ in their susceptibility to goal-driven modulation (Carrasco, 2011). For instance, according to LeDoux’ (1996) account, it is plausible that only the initial rapid orienting response would be determined by subcortical processes that are encapsulated from cognitive top-down control (Low Road; also see Luo et al., 2010). By contrast, maintaining or disengaging attention commences at a later stage and could be influenced by top-down goals, because potential threats are eventually also cognitively evaluated in the cortex (High Road; see also Carrasco, 2011, for a similar view). To assess this, a paradigm is needed that can manipulate both the locus of attention prior to the threat presentation and the goal relevance of a threat (see Vromen, Lipp, & Remington, 2015, for a similar critique). To study rapid attentional orienting to threat, it is imperative to ensure that when the threat distractor appears, attention is allocated elsewhere (i.e., the threat will need to capture attention to interfere with target responding). Also, to obtain a clear measurement of attentional disengagement, it is imperative that attention is already at the threat location (thus eliminating the need for attentional orienting), such that the time needed to disengage attention from the threat stimulus can be compared to disengagement from other, neutral stimuli.
In a previous study, Vromen, Lipp, and Remington (2015) used a modified spatial cueing paradigm to exert tight control over attention and disentangle orienting and disengagement (Posner, 1980). The observer’s task was to search for a green animal silhouette among three grey irrelevant animal silhouettes (distractors). Prior to target presentation, attention was controlled by a green pre-cue. Previous studies have shown that such a target-matching cue will reliably attract attention even when it is non-predictive of the target (Folk & Remington, 1998). The results confirmed that the green cue successfully controlled attention, as reflected in significantly faster responses on target-cued versus distractor-cued trials. Orienting was assessed on trials where the threat distractor (spider) was presented at a non-cued location, whereas disengagement was assessed on trials where the spider distractor was cued.
Vromen and colleagues (2015) showed that the goal-irrelevant spider silhouette only led to significant disengagement costs when spiders were among the possible target stimuli (i.e., potentially goal-relevant), not when the spider was completely task-irrelevant. Attention was not oriented to the spider silhouette, regardless of its relevance to the task-goal. These findings contrast with previous studies that showed large costs by task-irrelevant spider stimuli (Lipp & Waters, 2007; Öhman et al., 2012; Ouimet et al., 2012; Rinck et al., 2005). Such a discrepancy could be due to the fact that the modified spatial cueing paradigm encourages the implementation of a strict top-down target set due to the salient target color and brief, simultaneous presentation of target and distractor. Alternatively, it is possible that the spider stimulus failed to attract attention because the silhouette shape was not realistic enough to elicit fear or trigger a threat or arousal response. For instance, Mather and Sutherland (2011) proposed that the effects of high priority stimuli (e.g., threat) on attention are amplified by arousal. Similarly, Phelps and LeDoux (2005) have proposed that arousal is the key factor that determines the speed of processing different stimuli as well as how strongly they are represented (see also Lang & Bradley, 2010).
The purpose of the present study was to test whether attention would be oriented to more naturalistic threat distractors within the rigorous conditions of the modified spatial cueing paradigm. Moreover, the present study examined to what extent the early covert orienting of attention to threat would be modulated by top-down processes, by assessing the effects of photorealistic spider distractors on attention when a spider was among the possible targets (Experiments 1) versus when it was a completely task-irrelevant distractor (Experiment 2). Thus, the current study assessed orienting to and disengagement from identical spider distractors with their relevance to the task-goal being manipulated across experiments.
Experiment 1
The goal of the first experiment was to investigate whether prioritized orienting to and delayed disengagement from threatening photorealistic spider distractors (compared with non-threatening cat distractors) was evident when both were goal-relevant (i.e., part of the target-set). The observer’s task was to identify the green target singleton as a spider or cat, while ignoring the three grey-scale distractors (bird, fish, and spider or cat). On half of the trials a grey-scale spider was present amongst the distractors, whereas on the other half of the trials a grey-scale cat distractor was present (Fig. 1 displays an example of the stimulus displays). To assess whether the spider pictures were perceived as more threatening than the other animal pictures, the photorealistic animal pictures were rated in terms of their fearfulness and arousal. Moreover, to ensure generalizability of the results, the participant population was assessed in terms of their spider fear with the Spider Phobia Questionnaire (SPQ; Klorman, Hastings, Weerts, Melamed, & Lang, 1974).

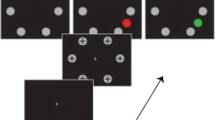
Procedure for the spatial cueing task (diagonally from bottom left to top right) showing an example of a foil-cued trial with a spider distractor from Experiment 1. The two cueing displays at the top left exemplify a target- and distractor-cued trial respectively. The foil-cued trials allowed examining delayed disengagement from the spider distractor (RT cued spider distractor – RT cued cat distractor), whereas target- and distractor-cued trials allowed us to examine orienting to the spider distractor (RT non-cued spider distractor – RT non-cued cat distractor)
Prior to presenting the target display, attention was cued to one of the four potential stimulus locations by a target-colored cue (four green dots), so that the spider or cat distractor was initially attended on some trials (disengagement trials) and unattended on others (orienting trials; Fig. 1). The effectiveness of the green cue in controlling attention was confirmed by faster target identification on trials on which the cue had been presented at the target location compared to trials where the cue had been presented elsewhere (Fig. 3). Pre-cueing of a threatening spider or a non-threatening cat distractor allowed measuring disengagement, as attention did not have to be oriented to the stimulus anymore, only to be disengaged and re-oriented to the target. Pre-cueing a different stimulus (e.g., neutral distractor or target) allowed measuring whether attention would be oriented to the threatening spider or non-threatening cat distractor, as they would need to capture attention in order to interfere with target identification. Based on the previous study by Vromen et al. (2015), threatening spider distractors would be expected to hold attention over and above non-threatening cat distractors, but only when relevant to the current task-goal. Due to the increased threat or arousal value of the photorealistic spider stimuli we also hypothesized that attention would be oriented more readily to threatening spider distractors than nonthreatening cat distractors.
Method
Participants
Twenty-four students from the University of Queensland, Australia (14 women; M = 19.29 years, SD = 3.42 years) participated in the experiment for course credit. One subject was excluded because of a 100 % error rate in one condition.
Participants’ total SPQ score ranged from 1 to 23, with a mean of 9.27 (SD = 6.63). This is indicative of a wide range of nonclinical spider fear, with nonfearful samples’ total SPQ score usually centered around 4 and arachnophobic samples’ around 23 (Fredrickson, 1983; Muris & Merckelbach, 1996).
Apparatus
Stimulus presentation was controlled by DMDX (Forster & Forster, 2003) running on a Dell Optiplex 780 computer. Stimuli were presented on a 19-in color monitor with a resolution of 1,024 × 768 pixels and a refresh rate of 85 Hz. Responses were collected through the left and right shift keys on a QWERTY keyboard.
Materials
Spatial cueing task
The fixation display consisted of a central fixation cross and four placeholder boxes (Fig. 1). Placeholders (2.9° × 2.5°) were positioned in a cross-like configuration equidistantly from the central fixation cross (5.7° from the nearest corner). The cueing display consisted of the fixation display with the addition of a spatial cue that consisted of a set of four green squares (CIE luv values = 80, −71, 92), each measuring (0.5° × 0.5°). The target display consisted of the fixation display with the addition of four animal pictures presented, one within each of the four placeholders. All target displays contained a spider, cat, bird, and fish, whereby either the spider or cat were presented in green (target), and the bird and fish were always presented in grey (nontargets).
In total, 24 photorealistic animal pictures were used, consisting of 6 spider, cat, bird, and fish pictures each. They were drawn from previous experiments conducted in the Emotion, Learning, and Psychophysiology Laboratory at The University of Queensland, from the Internet, and from the International Affective Picture System (IAPS; Lang, Bradley, & Cuthbert, 2005). The original picture backgrounds were replaced by a uniform white background and the animals were equated in size (M pixels = 2011) and average grey value (M CIEluv values = 54, 0, 0). The 12 animal target pictures used in Experiment 1 were created by transforming the color balance of each spider and cat picture so that bright green-scale images were created (M CIEluv values = 80, −71, 92). A pilot-test with 20 participants was conducted to confirm that spider and cat targets do not differ in time to be identified in a set-up where they were presented among three neutral animals, bird, fish, and horse nontargets, and in the absence of a cat or spider distractor). The results of a paired t test comparing trials with the spider versus cat target revealed no significant difference in target identification time (t(20) = 1.411, p = 0.174).
Rating task
To assess whether spider pictures were indeed more threatening than cat, bird, and fish pictures, respectively, animal fear ratings were collected using a visual analogue scale (VAS; 170-mm long) labelled with “Not at all fearful” on the left-hand side to “Very fearful” on the right-hand side, with no intermediate labels. Picture fear and arousal ratings for each animal category were also collected, with the VAS ranging from “Not at all scary” to “Very scary” for the former and from “Not at all arousing” to “Very arousing” for the latter.
As shown in Fig. 2, spider pictures had higher fear-ratings (M = 94.02, SD = 46.45) than cat, bird, and fish pictures (cats: M = 27.68, SD = 36.76, t(21) = 7.343, p < 0.001; birds: M = 42.00, SD = 42.39, t(21) = 4.058, p = 0.001; fish: M = 28.64, SD = 36.68, t(21) = 7.178, p < 0.001), whereas the others did not differ in terms of their fear ratings (cats-birds: t(21) = 1.436, p = 0.166; cats-fish: t(21) = 0.125, p = 0.902; birds-fish: t(21) = 1.259, p < 0.222). The results showed that spider and cat pictures had higher arousal ratings (spider: M = 55.64, SD = 46.05; cat: M = 44.05, SD = 44.05) than bird and fish pictures (birds: M = 26.57, SD = 27.04; fish: M = 28.07, SD = 29.97; spider vs. bird: t(21) = 2.909, p = 0.008; spider vs. fish: t(21) = 2.895, p = 0.009; cat vs. bird: t(21) = 2.370, p = 0.027; cat vs. fish: t(21) = 2.124, p = 0.046). Arousal ratings for spider and cat pictures did not differ from one another (t(21) = 1.104, p = 0.282) nor did arousal ratings for bird and fish pictures (t(21) < 1).

Mean fear and arousal ratings (and standard errors of the means) for the spider, cat, bird, and fish pictures in Experiment 1 (panel A) and Experiment 2 (panel B). In both experiments, spiders were rated as scarier than cats, birds, and fish. In Experiment 1, spiders and cats were rated equally arousing and as more arousing than birds and fish. In Experiment 2, fish were rated as less arousing than cats, whereas no other differences in arousal were observed. **p < 0.001; *p < 0.05
Design
The experiment consisted of the 3 × 2 within subjects conditions Cued Picture [target-, distractor-, foil-cued] and Distractor Identity [spider, cat]. Contingent on which picture was displayed in the cued placeholder, three trial types can be distinguished: target-cued trials (25 %; 72 trials) where the green cue was presented at the location of the green target picture (spider or cat); distractor-cued trials (50 %; 144 trials), in which the cue was presented at the location of a grey-scale picture with a nontarget identity (bird or fish); and foil-cued trials (25 %; 72 trials), where the cue was presented at the location of the grey distractor with a target identity (spider or cat). For half of the foil trials, the distractor was a spider and the other half was a cat.
Each picture was displayed equally often and each animal category was displayed equally often in each of the four placeholders. Each participant completed 12 practice trials and 288 test trials. Trial presentation order was randomized for each participant and allocation of target animal to response key was counterbalanced across participants.
Procedure
All participants first completed a spatial cueing task that consisted of a practice phase and a test phase. Immediately afterwards they completed the pen-and-paper version of the SPQ (Klorman et al., 1974) as well as the animal and picture VASs.
Spatial cueing task
At the start of the spatial cueing, task participants were informed that each target display would contain four animal pictures (spider, cat, bird, and fish). They were instructed to identify, as quickly and accurately as possible, whether the single green picture on each trial depicted a spider or a cat by pressing the left or right shift keys. Participants were told to try to ignore all grey-scale pictures and the green cue presented prior to the target display, because these stimuli were all irrelevant to their task.
Each trial started with the presentation of the fixation display (882 ms), which was followed immediately by the cue display (82 ms; Fig. 1). After the cue display, the fixation display was presented again (82 ms), followed by the target display (47 ms). The target display was followed by a white screen (882 ms). The next trial then started again with the presentation of the fixation display. Accuracy feedback was only presented on practice trials.
Results
Data
Reaction times from incorrect responses and extreme scores, defined as values more than three standard deviations above or below each participant’s mean reaction time, were excluded from the analyses leading to a loss of 10 % of all data.
Green cue
First, to assess whether the color cue directed attention, mean RT on target- and nontarget- (distractor- and foil-) cued trials were assessed. One-way ANOVA showed a significant effect of Cued Picture [target, distractor, foil] on target identification time, F(2,22) = 70.499, p < 0.001, partial η 2 = 0.865 (Fig. 3). Follow-up paired t tests showed faster target identification on target-cued trials than on distractor-cued trials (by 83 ms, t(23) = 11.613, p < 0.001) and foil-cued trials (by 96 ms, t(23) = 10.897, p < 0.001) and slower target identification on foil-cued trials compared with distractor-cued trials (by 13 ms, t(23) = 2.221, p = 0.037).

Mean target identification time (and standard error from the means) on target-, distractor-, and foil-cued trials in Experiment 1 (averaged over spider and cat distractor trials). Target identification is faster on target-cued trials than on distractor- and foil-cued trials. **p < 0.001; *p < 0.05
Orienting
To assess attentional orienting to spider distractors, we employed two paired t tests to compare target identification time on trials with a noncued spider versus cat distractor. The first paired t test showed that on distractor-cued trials, the spider slowed target identification by 52 ms more than the cat distractor, t(23) = 4.691, p < 0.001 (Fig. 4a). The second paired t test showed that even on target-cued trials where attention was already allocated to the target, the spider distractor slowed target identification, by 47 ms, compared with the cat-distractor, t(23) = 3.559, p < 0.05. Error rates did not differ between trials with a spider or cat distractor, neither on distractor-cued trials, t(23) = .277, p = 0.784, not on target-cued trials, t(23) = 1.045, p = 0.307).

a Mean target identification time (and standard errors of the means) on distractor- and target-cued trials with a spider versus cat distractor in Experiment 1. b Mean target identification time (and standard errors of the means) on foil-cued trials with a spider versus cat distractor in Experiment 1. **p < 0.001; *p < 0.05
Disengagement
To assess whether spider distractors delayed disengagement, we used a paired t test to compare target identification time on trials with a cued spider versus a cued cat distractor. Cueing the spider distractor slowed target identification on average by 72 ms compared with cueing of the cat distractor, t(23) = 5.253, p < 0.001 (Fig. 4b). The mean errors showed the same results, with significantly more errors when the spider distractor had been cued (M = 21.64, SD = 18.24) compared with when the cat distractor had been cued (M = 15.39, SD = 19.40; t(23) = 3.250, p = 0.004). We also conducted a t test to compare target identification time on trials were the spider distractor was cued (foil trials; 728 ms) or noncued (distractor trials; 702 ms). Cueing the spider distractor slowed target identification by 26 ms compared with presenting the spider distractor in a non-cued location, t(23) = 3.086, p = 0.005. The same t test for cat distractor trials (650 ms vs. 656 ms) showed no such difference, t(23) = 1.013, p = 0.321.
Discussion
The current experiment shows rapid orienting to photorealistic spider distractors in the modified cueing paradigm with a spider and cat target set. Rapid orienting to the threat distractor was observed even when attention was already at the target location and thus there was no need to shift attention. The experiment also shows evidence for delayed disengagement from the goal-relevant spider distractors when they were presented in a cued location. The current findings contrast with a previous study by Vromen and colleagues (2015), which showed that simplistic threat silhouettes delayed disengagement when they were goal-relevant, but did not capture attention, and suggests that the preattentive orienting to and the subsequent disengagement of attention from the same threat may be differentially susceptible to stimulus characteristics.
Experiment 2
Experiment 1 showed that spider distractors can both capture attention and delay disengagement when the current target-set includes spiders. To assess whether the rapid orienting of attention to spider distractors is contingent on top-down attentional control, Experiment 2 assessed orienting to the same spiders when they were not relevant to the top-down target-set (bird and fish targets). Previous studies have shown that spider distractors can bias attention even when they are completely irrelevant to the current task-goal, which has been interpreted as evidence for the evolutionary hypothesis that attentional orienting is not subject to top-down modulation. If this hypothesis holds in the present design, which 1) allows distinguishing between attentional orienting and disengagement and 2) encourages the adoption of a strict top-down set, then Experiment 2 should yield similar results as Experiment 1. However, a previous study has provided evidence that disengagement from threat can be affected by top-down modulation (Vromen et al., 2015). If orienting to threat also is contingent on top-down control, then we would expect Experiment 2 to show evidence of reduced attentional orienting to spider distractors. As in Experiment 1, if attention is oriented preferentially to spider distractors then noncued spider distractors should interfere with target responses, whereas delayed disengagement should manifest in prolonged RTs only when spider distractors are cued.
Method
Participants
Twenty-five students from the University of Queensland, Australia (13 women; M = 18.17 years, SD = 3.84) participated in the experiment for course credit. Participants’ total SPQ score ranged from 0 to 21, with a mean of 8.36 (SD = 5.09; Klorman et al., 1974). The SPQ scores in Experiment 2 did not differ from those in Experiment 1, as assessed by an independent-samples t test: t(45) = 0.533, p = 0.597.
Apparatus
The apparatus was identical to Experiment 1.
Materials
The materials were identical to Experiment 1, with the following exceptions.
Spatial cueing task
The targets in Experiment 2 were either a green fish or a green bird (identical to the grey-scale bird and fish pictures except in color; never a spider or cat). Each target display consisted of a green bird or fish (target), grey bird or fish (foil), spider or cat, and a horse (distractors). Six horse pictures were added to the overall stimulus set.
Rating task
Similar to Experiment 1, spider pictures had higher fear ratings (M = 88.13, SD = 55.35) than cat, bird, and fish pictures (cats: M = 16.00, SD = 20.52, t(23) = 6.116, p < 0.001; birds: M = 19.13, SD = 24.88, t(23) = 6.284, p < 0.001; fish: M = 20.79, SD = 35.79, t(23) = 6.676, p < 0.001; Fig. 2, right panel). Arousal ratings were comparable for all pictures (spiders: M = 48.12, SD = 48.81; fish: M = 27.00, SD = 35.21; all ts < 1.4), except for cats (M = 43.88, SD = 47.90) being rated as more arousing than fish (M = 27.00, SD = 35.21; t(21) = 2.118, p = 0.045).
Design
The design was identical to Experiment 1, with four stimuli displayed on each trial. The following changes were implemented. On each trial a bird, fish (one as the target, one as a target-similar distractor), either spider or cat (as target-dissimilar distractors), and a horse distractor were presented. To compare cat and spider distractor trials when both were irrelevant to the task, the targets in Experiment 2 were a green bird or a green fish (never a cat or spider). When the target was a bird, the fish was included in the display as the foil, and when the target was a fish, the bird acted as the foil. Moreover, each display contained a single irrelevant spider or cat distractor. Because we wanted to keep the number of stimuli within a display identical to Experiment 1 (4 items), we added a neutral horse distractor to each display. Thus, each display contained a green target (bird/fish), foil distractor (fish/bird), irrelevant threatening or nonthreatening distractor (spider/cat), and a neutral distractor (horse). With this, the conditions and displays in Experiment 2 were maximally similar to those used in Experiment 1, with all changes relating to the necessity of presenting the spider and cat distractors in separate displays (as these were the critical comparison categories). Attentional orienting and disengagement were measured by comparing trials with a spider versus cat distractor when they were not pre-cued and cued (by the green cue), respectively. Thus, four different trial types can be distinguished: (1) target-cued trials (25 %; 72 trials) where the green cue was presented at the location of the green target (green-scale bird or fish); (2) spider/cat distractor-cued trials (25 %; 72 trials) where the cue was presented at the location of the spider or cat distractor; (3) horse distractor-cued trials (25 %; 72 trials) where the cue was presented at the location of a grey-scale horse picture; and (4) foil-cued trials (25 %; 72 trials) where the cue was presented at the location of a grey-scale target-similar distractor (bird or fish).
Procedure
The procedure was identical to Experiment 1. The only difference was that participants were instructed to identify whether the single green picture on each trial depicted a bird or a fish (rather than a spider or cat as in Experiment 1).
Results
Green cue
Fifteen percent of all data was lost due to errors and extreme scores. The one-way ANOVA showed a significant effect of Cued Picture [target-, horse distractor-, spider/cat distractor-, and foil-cued] on target identification time, F(3,22) = 48.246, p < 0.01, partial η 2 = 0.868 (Fig. 5). Paired t tests showed that target identification was faster on target-cued trials than on distractor-cued trials (spider/cat cued: 101 ms, t(24) = 12.192, p < 0.001; horse cued: 98 ms, t(24) = 12.123, p < 0.001) and foil-cued trials (103 ms, t(24) = 10.935, p < 0.001), indicating that the color cue directed attention. There was no difference between the other trial types (horse-cued trials vs. spider/cat-cued: 3 ms, t(24) = 0.905, p = 0.374; spider/cat-cued vs. foil-cued: 2 ms, t(24) = 0.347, p = 0.732; horse-cued vs. foil-cued: 5 ms, t(24) = 0.839, p = 0.410).

Mean target identification time (and standard error of the means) for target-, horse distractor-, spider/cat distractor-, and foil-cued trials in Experiment 2 (averaged over spider and cat distractor trials). Target identification is faster on target-cued trials than on both the distractor-cued trials and the foil-cued trials. **p < 0.001
Orienting
To assess whether the spider distractors captured attention, we conducted three paired t tests to compare spider and cat distractor trials when the cue had been presented in a location other than that of the spider or cat (Fig. 6a). When the cue had been presented at the location of the horse (horse cued trial), target identification was 18-ms slower with a spider distractor than with a cat distractor (t(24) = 2.227, p = 0.036). When the cue had been presented at the location of the foil (fish or bird), there were no differences between spider and cat distractor trials, t < 1. Similarly, on target-cued trials, when the cue directed attention directly to the green target, there were no differences between spider and cat distractor trials, t(24) = 0.714, p = 0.482). The results for the mean error rates were in the same direction, with more errors on spider than cat distractor trials only observed on horse-cued trials, t(24) = 3.079, p = 0.005), and no differences on foil-cued trials, t(24) = 1.270, p = 0.994, or target-cued trials, t(24) = 0.175, p = 0.863.

a Mean target identification time (and standard errors of the means) on horse distractor-, foil-, and target-cued trials with a spider versus cat distractor in Experiment 2. b Mean target identification time (and standard errors of the means) on spider/cat distractor-cued trials in Experiment 2. **p < 0.001; *p < 0.05
Disengagement
To assess delayed disengagement, a paired t test compared target identification time on spider-cued trials with cat-cued trials. The results showed no difference between the two (t(24) = 0.971, p = 0.341; Fig. 6b). Similarly, the mean error rates showed no difference between spider and cat-cued trials (t(24) = 0.984, p = 0.335).
Orienting and disengagement compared across experiments
A direct comparison of the overall spider orienting scores (RT non-cued spider distractor trials minus RT non-cued cat distractor trials) between Experiments 1 (M = 49 ms) and 2 (M = 4 ms) with an independent samples t-test confirmed that orienting to spiders was significantly reduced in Experiment 2, t(46) = 3.731, p = 0.001. A direct comparison of the overall spider disengagement scores (RT cued spider distractor trials minus RT cued cat distractor trials) between experiments 1 (M = 73 ms) and 2 (M = 12 ms) with an independent samples t-test confirmed that spider disengagement also was significantly reduced in Experiment 2, t(46) = 3.308, p = 0.002.
Discussion
The results of Experiment 2 showed that orienting to spider distractors was much reduced when spiders were not included in the target-set. The spider distractor only attracted attention when attention had initially been misdirected to an entirely task-irrelevant stimulus but not when the target or target-similar distractor (foil) had been cued. Thus, even when the spider was task-irrelevant it still had the ability to capture attention, but only when the initially attended (cued) stimulus did not contain any task-relevant features (nontarget identity and color). These findings indicate that threatening spider stimuli did only capture attention away from task-relevant stimuli (foil, target; Experiment 1) when included in the target set (i.e., potentially task-relevant), which provides strong evidence for the view that attentional orienting to threatening stimuli is modulated by goal-driven processes. A second important finding of the current study was that delayed disengagement from the threat was eliminated when spiders were not included in the target-set. Of note, deviating from previous studies, capture by the spider was not necessary to observe delayed disengagement, as attention was directed to the spider by the green cue.
General discussion
The present study provides evidence that top-down processes modulate both attentional orienting and attentional disengagement when these processes are manipulated and measured independently from each other. The study also demonstrates that a task-relevant spider distractor can attract attention even when the locus of attention is tightly controlled. This contrasts with the findings of Vromen et al. (2015) where a potentially task-relevant spider silhouette did not attract attention. Consequently, rapid orienting to threat may require a richness in stimulus features that is observed only in more naturalistic stimuli. More naturalistic images are likely also evaluated as being more threatening or arousing. Support for perceived threat value, rather than arousal value, was observed in the current study as spider images were rated as more threatening, but not more arousing, than control stimuli (Fig. 2; e.g., Mather & Sutherland, 2011; Phelps & LeDoux, 2005; although a strong test of the arousal hypothesis may require a psychophysiological measurement of arousal).
It should be noted that the present study cannot rule out that the effects of the threatening spider were due to its salient perceptual features (e.g., its long legs). Previous studies have shown that attention can be automatically attracted to salient features that are embedded in complex stimuli (such as visible teeth in angry faces; Becker, Horstmann, & Remington, 2011; Horstmann & Becker, 2008; Horstmann, Lipp, & Becker, 2012). However, these findings do not necessarily contradict the evolutionary view, as certain feature detectors (e.g., for the detection of spiders) could have evolved because they were relevant for survival, thus rendering the visual system more sensitive to these features (Horstmann & Bauland, 2006). Moreover, the conclusion that the implied threat value plays a role in driving attention seems to be the most parsimonious explanation for the differences observed between spider silhouettes vs. more naturalistic spider stimuli (Vromen et al., 2015); specifically, the finding that only the more naturalistic spider attracted attention when spiders were completely task-irrelevant.
Of most interest to our research question, we observed that orienting to spider distractors was modulated by top-down goals such that orienting was strong when a spider was amongst the possible targets (i.e., potentially goal-relevant; Experiment 1) but much reduced when it was completely goal-irrelevant (Experiment 2). Specifically, when the spider was potentially goal-relevant, it attracted attention even when attention was pre-cued to the target and obviated the need for further search. By contrast, when the spider was goal-irrelevant and attention was cued to a target or foil, the spider distractor failed to attract attention. Only when attention had been pre-cued to another task-irrelevant and target-dissimilar distractor did the photorealistic spider attract attention. The results show that spider distractors have an ability to attract attention even when they are completely task-irrelevant.
This finding is in line with previous studies that have demonstrated evidence for rapid orienting to (simplistic) task-irrelevant threat (Carlson & Reinke, 2008). However, previous studies differ from the present study in that (1) the target was often not singled out by a salient perceptual feature (e.g., green colour) that would allow effective top-down tuning of attention to the target; (2) the target was often presented until the response, which contrasts with the data-limited displays used in the present study; and in that (3) the locus of attention was not as tightly controlled as in the present study (Lipp & Waters, 2007; Mogg & Bradley, 1998; Öhman & Mineka, 2001; Rinck et al., 2005; Schmidt, Belopolsky, & Theeuwes, 2012; Schmidt, Belopolsky, & Theeuwes, 2014). The modified cueing paradigm as employed in the current study may be less likely than previously employed paradigms to show orienting to goal-irrelevant threats due to the conditions encouraging a strong top-down bias to the target feature(s). Hence, the finding that task-irrelevant photorealistic spiders could still attract attention shows that threatening stimuli can attract attention quite strongly—viz., against an effective top-down set for completely different stimuli (i.e., green bird and fish targets) and despite the fact that data-limited displays require allocating attention swiftly to the target. That said, the theoretically more important finding is perhaps that orienting to threat was still modulated by top-down control (see comparison between Experiment 1 and 2). This shows that orienting to threat is not encapsulated from top-down control (LeDoux, 1996; Öhman & Mineka, 2001) but instead requires a model that takes both the (subjective) threat value of stimuli and their task relevance into account, and allows top-down modulation of threatening stimuli (e.g., competitive interaction model; Mathews & Mackintosh, 1998; Desimone & Duncan, 1995; Pessoa & Adolphs, 2010).
The current results also demonstrate top-down modulation of attentional disengagement, with a spider distractor delaying disengagement only when it was potentially goal-relevant (Experiment 1). Both orienting and disengagement have to be due to distractor-related effects and cannot be due to target-related effects, because a pilot study without spider and cat distractors showed that the identification speed was comparable for spider and cat targets. The observed top-down modulation of both orienting and disengagement may call into question a strict distinction between earl and late attentional processes, which has, for example, been proposed by Carrasco (2011). Instead it may indicate that both are largely determined by the same, cognitively penetrable mechanisms (Becker, 2011). However, this conclusion is certainly speculative and would require further research.
Potential alternative accounts
Our conclusion that the differences in orienting and disengagement between Experiments 1 and 2 were due to top-down influences could still be subject to two possible objections. First, it should be noted that if participants in Experiment 2 were generally less fearful of spiders, this could provide an alternative explanation for reduced attentional orienting to threat. Second, the mean target identification time in Experiment 2 (M = 718 ms) was longer than in Experiment 1 (M = 663 ms). One might argue that slower target identification could be correlated with reduced spider interference, if the process that is needed to override attention to spider distractors is an active, effortful process that takes time to implement (e.g., filtering; see Folk & Remington, 1998). If this was the case, spider interference would be expected to be more pronounced on trials with relatively faster RTs and less pronounced on trials with slower RTs within each of the experiments.
To assess whether attentional orienting was affected by spider fear, we first reanalysed the data from Experiments 1 and 2 separately for the relatively more and less spider fearful participants. Second, to probe whether prioritized attention to spider distractors and its top-down control differed over the response time distribution, we assessed orienting and disengagement separately for each decile of the RT distribution.
Effects of self-reported spider fear
Orienting
To assess whether prioritized orienting to spider distractors was contingent on spider fear we performed a median split analysis based on participants’ total SPQ scores (median SPQ score Experiment 1: 8; Experiment 2: 7). For Experiment 1, an independent samples t test showed that the orienting difference scores (RT noncued spider distractor minus RT noncued cat distractor) did not differ between the relatively more and less spider fearful group, neither on distractor-cued trials, t(22) = 0.282, p = 0.695, nor target-cued trials, t(22) = 1.513, p = 0.145. The same comparisons for the overall error rates showed no difference either between the relatively more and less fearful groups (distractor-cued: t(22) = 0.295, p = 0.771; target-cued: t(22) = 0.851, p = 0.404).
In a similar vein, no differences in orienting were observed between the relatively more and less spider fearful groups in Experiment 2. Target identification time was comparable for the relatively more and less fearful groups on horse distractor-cued trials: t(23) = 0.759, p = 0.456, and target-cued trials: t(23) = 1.225, p = 0.233). The overall difference between the relatively more and less fearful groups was significant (in the hypothesized direction) on foil-cued trials, t(23) = 2.691, p = 0.013, but follow-up t tests showed that neither group showed a delay in target identification that differed significantly from zero (higher fear: t(12) = 1.795, p = 0.098; lower fear: t(11) = 2.027, p = 0.068). No differences in error rates were observed either between the groups (horse distractor-cued trials: t(23) = 1.728, p = 0.097; foil-cued trials: t(23) = .982, p = 0.336; target-cued trials: t(23) = 1.635, p = 0.116).
Disengagement
To assess whether attentional disengagement was contingent on spider fear, we compared the disengagement score (RT cued spider distractor minus RT cued cat distractor) for the relatively more and less spider fearful groups with another independent samples t test. No differences in disengagement were observed between the relatively more and less spider fearful groups (Experiment 1: t(22) = 0.609, p = 0.549; Experiment 2: t(23) = 0.814, p = 0.424). No differences in error rates were observed either (Experiment 1: t(22) = 0.445, p = 0.661; Experiment 2: t(23) = 1.562, p = 0.132).
In sum, there were no differences in the mean SPQ score between experiments nor did the behavioural results differ between the relatively more and less spider fearful groups within both experiments. Combined, these results render it unlikely that the reduction in orienting to and rapid disengagement from spiders in Experiment 2 can be attributed to spider fear.
Results from the response time distributions
To assess whether the longer RTs in Experiment 2 compared with Experiment 1 could account for reduced attention to threat, we assessed orienting and disengagement separately for faster and slower responses. Specifically, we assessed these effects within each decile of the RT distributions.
Orienting
To test whether prioritized attentional orienting to the noncued spider distractors in Experiment 1 was observed across all points of the RT distribution, we binned distractor- and target-cued trials with a spider versus cat distractor based on RT (Leber, Lechak, & Tower-Ricardi, 2013; Fig. 7). The first decile (0.1) contains the fastest 10 % of response times, the second decile (0.2) the second fastest 10 % of response times, etc. Paired t tests comparing target identification time for spider and cat distractor trials showed that, for the distractor-cued trials (Fig. 7, first panel), the spider distractor slowed target identification more than the cat distractor in all deciles, with the smallest t value observed in the first decile (all t(23) > 2.612, all p < 0.017). For the target-cued trials (second panel), the spider distractor significantly slowed target identification in each of the deciles, except the last decile (t(23) = 1.270, p = 0.217; all other p < 0.018).

Mean target identification time for spider and cat trials for each RT decile in Experiment 1, with the first panel showing distractor-cued trials and the second panel showing target-cued trials
The same analyses conducted for Experiment 2 showed that on horse distractor-cued trials (Fig. 8, first panel) the spider distractor slowed target identification in the intermediate deciles (0.4: t(24) = 2.200, p < 0.038; 0.5: t(24) = 2.646, p < 0.014; 0.6: t(24) = 3.057, p = 0.005; 0.7: t(24) = 3.135, p = 0.004), whereas it failed to do so in both the earlier and later deciles (all t(24) < 1.979, all p > 0.150). For the foil-cued trials (Fig. 8, second panel), the spider distractor slowed target identification in the 4th and 5th deciles (0.4: t(24) = 2.629, p < 0.05; 0.5: t(24) = 2.060, p < 0.05) but not in any of the other deciles (all t(24) < 1.686, all p > 0.104). For the target-cued trials (Fig. 8, third panel), the spider distractor did not delay target identification in any decile (all t(24) < 1.527, all p > 0.140).

Mean target identification time for spider and cat trials for each RT decile in Experiment 2, with the first panel showing horse distractor-cued trials, the second panel showing foil-cued trials, and the third panel showing target-cued trials
Disengagement
The RT distribution analysis for Experiment 1 showed that the spider distractor delayed target identification more than the cat distractor in all 10 deciles, with the smallest t-value observed in the fourth decile (all t(23) > 3.520, all p < 0.003; Fig. 9). Conversely, for Experiment 2, the spider distractor did not slow target identification more than the cat distractor in any of the RT deciles (all t < 1.863, all p >0.074; Fig. 10).

Mean target identification time for spider and cat cued trials for each RT decile in Experiment 1

Mean target identification time for spider and cat cued trials for each RT decile in Experiment 2
The RT distribution analyses show that the difference in result between experiments cannot be attributed to overall RTs being longer in Experiment 2. Rather, when spiders were task-relevant (Experiment 1), rapid orienting and delayed disengagement were observed over the entire RT distribution, indicating spider interference to be robust and not disproportionally driven by trials with especially fast or slow responses. When spiders were task-irrelevant (Experiment 2), orienting and disengagement were not more pronounced on trials with shorter RTs. The RT distribution analysis revealed some residual orienting to spiders in intermediate deciles when attention had been initially deployed to a target-(dis)similar distractor but not when attention had been cued to the green target. These results are consistent with a model in which attention to threat is modulated by top-down mechanisms that boost target-similar but not target-dissimilar stimuli (or perhaps even down-modulates the latter. Mathews & Mackintosh, 1998; see also e.g. Desimone & Duncan 1995; Pessoa & Adolphs, 2010).
The RT distributions also indicated that disengagement was perhaps more strongly modulated by goal-driven processes (i.e., no residual disengagement delays from the spider distractor in any RT decile). The present results essentially indicate that, when a tight top-down set is encouraged, attentional disengagement depends only on whether the selected stimulus is similar to the target, regardless of the threat potential of a stimulus. These results are in line with the view that disengagement is entirely under the command of top-down goals, possibly because the perceptual analysis of selected stimuli can be limited to only task-relevant aspects and features (with identification initially proceeding on a “need to know” basis for the purpose of distractor rejection; Becker, 2011; Hahn & Gronlund, 2007).
The finding that individual spider fear did not modulate orienting or disengagement is at odds with previous studies showing, for instance, delayed disengagement in participants who are highly fearful of threat-related stimuli (e.g., in the dot probe task; Fox et al., 2001). Whereas the current study showed no modulation of threat interference or top-down control by spider fear, it is of note that we tested a nonphobic sample and that the median is a rather arbitrary divide. Thus, the present results should not be taken to mean that subjective fear cannot modulate orienting or disengagement. Further research is needed to investigate how top-down modulation of threat interference is affected in individuals with high (clinical) levels of spider fear and whether fear could affect top-down modulation of attentional orienting and disengagement differently.
Conclusions
The current study indicates that attentional orienting to threat may only be observed when nothing more relevant to one’s current goals is vying for attention. These findings provide a new perspective on the research domain of attention to threat by indicating that even the early orienting of attention to threat is contingent on current top-down goals, suggesting early orienting to threat to be more complex and cognitively involved than previously hypothesized.
References
Ansorge, U., Kiss, M., & Worschech, F. (2010). The initial stage of visual selection is controlled by top-down task-set: new ERP evidence. Attention, Perception, & Psychophysics, 73, 113–122. doi:10.3758/s13414-010-0008-3
Awh, E., Matsukura, M., & Serences, J. T. (2003). Top-down control over biased competition during covert spatial orienting. Journal of Experimental Psychology: Human Perception & performance, 29, 52–63. doi:10.1037/0096-1523.29.1.52
Becker, S. I. (2011). Determinants of dwell time in visual search: Similarity or perceptual difficulty? PloS One, 6(3), 1–5. doi:10.1371/journal.pone.0017740
Becker, S. I., Horstmann, G., & Remington, R. W. (2011). Perceptual grouping, not emotion, accounts for search asymmetries with schematic faces. Journal of Experimental Psychology: Human Perception and Performance, 37, 1739–1757. doi:10.1037/a0024665
Belopolsky, A. V., Devue, C., & Theeuwes, J. (2011). Angry faces hold the eyes. Visual Cognition, 19(1), 27–36. doi:10.1080/13506285.2010.536186
Carlson, J. M., & Reinke, K. S. (2008). Masked fearful faces modulate the orienting of covert spatial attention. Emotion, 8, 522–529. doi:10.1037/a0012653
Carrasco, M. (2011). Visual attention: The past 25 years. Vision Research, 51, 1484–1525. doi:10.1016/j.visres.2011.04.012
Chun, M. M., Golomb, J. D., & Turk-Browne, N. B. (2011). A taxonomy of external and internal attention. Annual Review of Psychology, 62, 73–101. doi:10.1146/annurev.psych.093008.100427
Desimone, R., & Duncan, J. (1995). Neural mechanisms of selective visual-attention. Annual Review of Neuroscience, 18, 193–222. doi:10.1146/annurev.ne.18.030195.001205
Devue, C., Belopolsky, A., & Theeuwes, J. (2011). The role of fear and expectancies in capture of covert attention by spiders. Emotion, 11, 768–775. doi:10.1037/a0023418
Folk, C. L., & Remington, R. W. (1998). Selectivity in distraction by irrelevant featural singletons: evidence for two forms of attentional capture. Journal of Experimental Psychology: Human Perception & Performance, 24, 847–858. doi:10.1037/0096-1523.24.3.847
Folk, C. L., Remington, R. W., & Johnston, J. C. (1992). Involuntary covert orienting is contingent on attentional control settings. Journal of Experimental Psychology: Human Perception & performance, 18(4), 1030–1044. doi:10.1037/0096-1523.18.4.1030
Forster, K. I., & Forster, J. C. (2003). DMDX: A windows display program with millisecond accuracy. Behavior Research Methods, Instruments, and Computers, 35(1), 116–124. doi:10.3758/BF03195503
Fox, E., Russo, R., Bowles, R. J., & Dutton, K. (2001). Do threatening stimuli draw or hold visual attention in subclinical anxiety? Journal of Experimental Psychology: General, 130(4), 681–700. doi:10.1037/0096-3445.130.4.681
Fox, E., Russo, R., & Dutton, K. (2002). Attentional bias for threat: Evidence for delayed disengagement from emotional faces. Cognition & Emotion, 16(3), 355–379. doi:10.1080/02699930143000527
Fredrickson, M. (1983). Reliability and validity of some specific fear questionnaires. Scandinavian Journal of Psychology, 24, 331–334. doi:10.1111/j.1467-9450.1983.tb00507.x
Hahn, S., & Gronlund, S. D. (2007). Top-down guidance in visual search for facial expressions. Psychonomic Bulletin & Review, 14(1), 159–165. doi:10.3758/BF03194044
Horstmann, G., & Bauland, A. (2006). Search asymmetries with real faces: Testing the anger-superiority effect. Emotion, 6, 193–207. doi:10.1037/1528-3542.6.2.193
Horstmann, G., & Becker, S. (2008). Attentional effects of negative faces: Top-down contingent or involuntary? Perception and Psychophysics, 70, 1416–1434. doi:10.3758/PP.70.8.1416
Horstmann, G., Lipp, O.V., & Becker, S.I. (2012). Of toothy grins and angry snarls? Open mouth displays contribute to efficiency gains in search for emotional faces. Journal of Vision, 12(5):7, 1–15. doi:10.1167/12.5.7
Klorman, R., Hastings, J., Weerts, T., Melamed, B., & Lang, P. (1974). Psychometric description of some specific fear questionnaires. Behavior Therapy, 5, 401–409. doi:10.1016/S0005-7894(74)80008-0
Koster, E. H. W., Crombez, G., Verschuere, B., & De Houwer, J. (2004). Selective attention to threat in the dot probe paradigm: Differentiating vigilance and difficulty to disengage. Behaviour Research & Therapy, 42(10), 1183–1192.
Lang, P. J., & Badley, M. M. (2010). Emotion and the motivational brain. Biological Psychology, 84, 437–450. doi:10.1016/j.biopsycho.2009.10.007
Lang, P. J., Bradley, M. M., & Cuthbert, B. N. (2005). International Affective Picture System (IAPS): Affective ratings of pictures and instruction manual. Technical Report A-6. Gainesville: University of Florida.
Leber, A. B., Lechak, J. R., & Tower-Richardi, S. M. (2013). What do fast response times tell us about attentional control. Journal of Vision, 13(3), 1–12. doi:10.1167/13.3.31
LeDoux, J. E. (1996). The emotional brain: The mysterious underpinnings of emotional life. New York, NY, US: Simon & Schuster. doi:10.1016/B978-0-12-420170-5.00010-6
Lipp, O. V., & Waters, A. M. (2007). When danger lurks in the background: Attentional capture by animal fear-relevant distractors is specific and selectively enhanced by animal fear. Emotion, 7(1), 192–200. doi:10.1037/1528-3542.7.1.192
LoBue, V., & DeLoache, J. S. (2008). Detecting the snake in the grass: Attention to fear-relevant stimuli by adults and young children. Psychological Science, 19(3), 284–289. doi:10.1111/j.1467-9280.2008.02081.x
LoBue, V., & DeLoache, J. S. (2011). What’s so special about slithering serpents? Children and adults rapidly detect snakes based on their simple features. Visual Cognition, 19, 129–143. doi:10.1080/13506285.2010.522216
Luo, Q., Holroyd, T., Majestic, C., Cheng, X., Schechter, J., & Blair, R. J. (2010). Emotional automaticity is a matter of timing. Journal of Neuroscience, 30, 5825–5829. doi:10.1523/JNEUROSCI.BC-5668-09.2010
Mather, M., & Sutherland, M. R. (2011). Arousal-biased competition in perception and memory. Psychological Science, 6, 1–20. doi:10.1177/1745691611400234
Mathews, A., & Mackintosh, B. (1998). A cognitive model of selective processing in anxiety. Cognitive Therapy & Research, 22(6), 539–560. doi:10.1023/A:1018738019346
Mogg, K., & Bradley, B. (1998). A cognitive motivational analysis of anxiety. Behaviour Research Therapy, 36(9), 809–848. doi:10.1016/S0005-7967(98)00063-1
Muris, P., & Merckelback, H. (1996). A comparison of two spider fear questionnaires. Journal of Behaviour Therapy & Experimental Psychiatry, 27, 241–244. doi:10.1016/j.brat.2006.05.003. doi:10.1016/S0005-7916(96)00022-5
Öhman, A., & Mineka, S. (2001). Fears, phobias, and preparedness: Toward an evolved module of fear and fear learning. Psychological Review, 108(3), 483–522. doi:10.1037//0033-295X.108.3.483
Öhman, A., Soares, S. C., Juth, P., Lindström, B., & Esteves, F. (2012). Evolutionary derived modulations of attention to two common fear stimuli: Serpents and hostile humans. Journal of Cognitive Psychology, 24, 17–32. doi:10.1080/20445911.2011.629603
Ouimet, A. J., Radomsky, A. S., & Barber, K. C. (2012). Interrelationships between spider fear associations, attentional disengagement and self-reported fear: A preliminary test of a dual-systems model. Cognition & Emotion, 26, 1–17. doi:10.1080/02699931.2012.671175
Pessoa, L., & Adolphs, R. (2010). Emotion processing and the amygdala: From a ‘low road’ to ‘many roads’ of evaluating biological significance. Nature Reviews Neuroscience, 11, 773–782. doi:10.1038/nrn2920
Phelps, E. A., & LeDoux, J. E. (2005). Contributions of the amygdala to emotion processing: From animal models to human behaviour. Neuron, 48, 175–187. doi:10.1016/j.neuron.2005.09.025
Posner, M. I. (1980). Orienting of attention. Quarterly Journal of Experimental Psychology, 32, 3–25. doi:10.1080/00335558008248231
Rinck, M., Reinecke, A., Ellwart, T., Heuer, K., & Becker, E. S. (2005). Speeded detection and increased distraction in fear of spiders: Evidence from eye movements. Journal of Abnormal Psychology, 114(2), 235–248. doi:10.1037/0021-843X.114.2.235
Rudaizky, D., Basanovic, J., & MacLeod, C. (2014). Biased attentional engagement with, and disengagement from, negative information: Independent cognitive pathways to anxiety vulnerability? Cognition & Emotion, 28, 245–259. doi:10.1080/02699931.2013.815154
Schmidt, L., Belopolsky, A., & Theeuwes, J. (2012). The presence of threat affects saccade trajectories. Visual Cognition, 20, 284–299. doi:10.1080/13506285.2012.658885
Schmidt, L. J., Belopolsky, A. V., & Theeuwes, J. (2014). Attentional capture by signals of threat. Cognition & Emotion, 29, 687–694. doi:10.1080/02699931.2014.924484
Serences, J. T., Shomstein, S., Leber, A. B., Golay, X., Egeth, H. E., & Yantis, S. (2005). Coordination of voluntary and stimulus-driven attentional control in human cortex. Psychological Science, 16, 114–122. doi:10.1111/j.0956-7976.2005.00791.x
Stein, T., Seymour, K., Hebart, M. N., & Sterzer, P. (2014). Rapid fear detection relies on high spatial frequencies. Psychological Science, 25(2), 566–574. doi:10.1177/0956797613512509
Tamietto, M., & De Gelder, B. (2010). Neural bases of the non-conscious perception of emotional signals. Nature Reviews Neuroscience, 11, 697–709. doi:10.1038/nrn2889
Vromen, J.M.G., Lipp, O.V., & Remington, R.W. (2015). The spider does not always win the fight for attention: Disengagement from threat is modulated by goal set. Cognition & Emotion, 1–12. Advance online publication. doi:10.1080/02699931.2014.969198
West, G. L., Anderson, A. K., & Pratt, J. (2009). Motivationally significant stimuli show visual prior entry: Evidence for attentional capture. Journal of Experimental Psychology: Human Perception and Performance, 35(4), 1032–1042. doi:10.1037/a0014493
Wolfe. (1994). Guided search 2.0. A revised model of visual search. Psychonomic Bulletin & Review, 1, 202–238. doi:10.3758/BF03200774
Wolfe, J. M., Butcher, S. J., Lee, C., & Hyle, M. (2003). Changing your mind: On the contributions of top-down and bottom-up guidance in visual search for feature singletons. Journal of Experimental Psychology: Human Perception & performance, 29, 483–502. doi:10.1037/0096-1523.29.2.483
Yiend, J. (2010). The effects of emotion on attention: A review of attentional processing of emotional information. Cognition & Emotion, 24(1), 3–47. doi:10.1080/02699930903205698
Acknowledgments
This work was supported by the Australian Research Council grant DP120100750 to OVL and a grant from UQRS/UQIRTA to JMGV.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Vromen, J.M.G., Lipp, O.V., Remington, R.W. et al. Threat captures attention, but not automatically: Top-down goals modulate attentional orienting to threat distractors. Atten Percept Psychophys 78, 2266–2279 (2016). https://doi.org/10.3758/s13414-016-1142-3
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13414-016-1142-3