
Abstract
Titanium dioxide nanoparticles (TiO2 NPs) are used in cosmetics, sunscreens, paints, and toothpaste, among other applications. These NPs are very stable and can be transported and dispersed in wastewater and biosolids. Animal species have shown negative reactions to TiO2 NPs. However, little is known about their toxicity in plants, specifically the possibility of genotoxic effects. In this study, we used a random amplified polymorphic DNA (RAPD) technique to study the genotoxic effects of TiO2 NPs on hydroponically cultivated zucchini (Cucurbita pepo) plants. Seeds were allowed to germinate for 7 d and plants were selected at random for individual and population studies. Four plants were selected for the individual study and 18 for the population study. RAPD profiles of TiO2 NPs treated plants showed differences in band intensity, loss of bands, or appearance of new bands, compared to untreated plants. To the authors’ knowledge, this is the first report of the genotoxic potential of TiO2 NPs in zucchini.
概要
研究目的
二氧化钛 (TiO2) 纳米颗粒已经广泛应用于化妆品、 防晒霜、 涂料和牙膏等。 这些纳米颗粒性质非常稳定, 能在废水和生物固体中转移和分散。 现有研究表明, TiO2 纳米颗粒对动物正常生理活动具有毒性等负面作用。 但是, 它们对植物是否具有毒性特别是是否会产生植物基因毒性至今尚不清楚。 因此, 本文使用随机扩增多态性 DNA 技术研究 TiO2 纳米颗粒是否对西葫芦具有基因毒性, 为 TiO2 纳米颗粒排放进入环境后的潜在植物毒性风险评价提供依据。
创新要点
首次发现了 TiO2 纳米颗粒对西葫芦具有基因毒性。
重要结论
采用随机扩增多态性 DNA 技术, 发现 TiO2 纳米颗粒污染处理的西葫芦样品与未处理样品的基因组 DNA 图谱相比, 不仅在谱带强度有明显差异, 而且存在谱带消失和新谱带产生现象, 表明 TiO2 纳米颗粒对西葫芦具有基因毒性。
Similar content being viewed by others

1 Introduction
Nanotechnology is considered an innovative field. The emergence of nanotechnologies has contributed to great progress in various industries, similar to the introduction of electricity and biotechnology (Roco et al., 2010). Nanomaterials are used in the production and enhancement of hundreds of consumer products such as computers, gas sensors, stainresistant clothing, sunscreens, cosmetics, and agricultural products (Williams and Adams, 2007; Johnson et al., 2011; Hong et al., 2013). Since 2001, the demand for nanotechnologybased products has increased at great pace (Roco, 2011). In 2009, the market value of products using nanotechnology reached about $254 billion (Roco et al., 2010). However, the mass production of nanotechnology based products has raised questions about their environmental impact.
Titanium dioxide (TiO2) is one of the most widely used materials at the nanoscale, but its interaction with environmental components is not well understood (Darlington et al., 2009). According to Robichaud et al. (2009), the estimated annual global production of TiO2 will reach almost 2.5 million metric tons by 2025. New studies confirm that TiO2, compared to nanoparticulate zinc oxide, silver, and carbon nanotubes, is more likely to be released into the environment in larger quantities (Gottschalk et al., 2009). The fate of the nanoparticles (NPs) after production is unknown (Keller et al., 2013), therefore, it is important to understand the potential impacts these NPs may have on living organisms, especially plants, since they are a critical link in the food chain.
Several reports indicate that NPs affect several plants at different levels. Genotoxic effects of ZnO and cerium oxide (CeO2) NPs have been reported in soybean (López Moreno et al., 2010). Previous studies have shown that TiO2 NPs affect the genomic DNA of Arabidopsis thaliana (Landa et al., 2012), but to the authors’ knowledge, there are no reports on the genotoxic effects of these NPs on zucchini (Cucurbita pepo) plants.
Random amplified polymorphic DNA (RAPD) is a polymerase chain reaction (PCR)based technique that allows for quick and efficient comparisons of genomic DNA. This technique has been successfully used to study genotoxicity in different plants (Liu et al., 2005; Cenkci et al., 2009; Kumari et al., 2009; Liu et al., 2009). The lack of a published genomic DNA sequence for C. pepo prompted the use of this technique. Typically, PCR is performed with one forward and one reverse oligonucleotide primer, each of which binds to a specific DNA sequence flanking the area of interest within the genome, allowing amplification of the region between the primers. However, this method relies on the use of a known DNA sequence. When the DNA sequence is not known, it is possible to use oligonucleotide primers with random sequences to amplify regions of the genome. Often, several different random primers are examined to determine which primer pair exhibits the greatest binding affinity for the genomic template. Once the random primer pairs are selected, they are used in PCR with genomic DNA to amplify random DNA fragments, which can be analyzed by agarose gel electrophoresis (MacPherson and Gajadhar, 1992).
In ecotoxicology, toxic agents can show effects on individuals, but to generalize these effects and draw conclusions, it is necessary to do a population study (Amaral et al., 2012). Therefore, in this report we described two studies that we performed to evaluate the differences between a population study and an individual study. We determined whether TiO2 NPs have the potential to exert genotoxic effects on individual zucchini plants by exposing them to TiO2 NPs for 7 d and analyzing the genomic DNA by RAPD-PCR and gel electrophoresis immediately before and following exposure. For the population study, an experimental group of plants was exposed to TiO2 NP suspensions for 7 d while a replicate control group remained unexposed. Following treatment, RAPD analysis was performed on both groups.
2 Methods
2.1 Plant growth and TiO2 NP treatment
Zucchini seeds (Burpee®) were purchased at a local store. Seeds were stirred in 4% hypochlorite for 30 min, and then left overnight stirring in deionized water for cleaning and hydration. Subsequently, they were treated with an antibiotic/antimycotic solution (N-Acetyl-Asp-Gln-Met-Asp-7-amido-4-trifluoromethylcoumarin; Sigma-Aldrich A5595) to prohibit microbial growth, since the presence of microbial DNA could interfere with the results or interpretation of the study. For germination, the seeds were distributed on the border of sterile wet paper towels, rolled up, placed into Mason jars and incubated at 25 °C/20 °C day/night temperature in the dark for 5 d. The seedlings were then incubated for two more days at the same temperature with a photoperiod of 14 h. Subsequently, plants were selected at random for individual and population studies. Four plants were selected for the individual study and 18 for the population study. The selected plants were hydroponically grown in modified Hoagland nutrient solution for 7 d in an environmental growth chamber (Environmental Growth Chamber, Chagrin Falls, OH) with a 14-h photoperiod, illumination of 340 µmol/(m2·s), 25 °C/20 °C day/night temperature, and 65% relative humidity. Cultivation was performed as previously described by Servin et al.(2013).
The TiO2 NPs ((27±4) nm, Evonik Degussa C., USA) were obtained from the University of California Center for Environmental Implications of Nanotechnology (UC CEIN). These NPs were previously characterized as semispherical with a surface area 51.5 m2/g and formed by anatase (82%) and rutile (18%) crystalline phases (Keller et al., 2010).
A suspension of 50 mg/L TiO2 NPs in modified Hoagland solution (Peralta-Videa et al., 2002) was made by sonicating the NP suspension for 30 min to avoid aggregation. Plants were hydroponically incubated in the suspension for 7 d. For the individual plant studies, genomic DNA was extracted from each of the four plants before and after the 7-d treatment with TiO2 NPs. For the population studies, nine of the 18 plants were exposed for 7 d to the 50 mg/L TiO2 suspension, while the other nine were left in the modified Hoagland solution lacking TiO2 NPs. After treatment, genomic DNA was extracted from the roots of all 18 plants. In all cases, plant genomic DNA was extracted from the tip of the root, with minimum disturbance, using a Wizard® Genomic DNA Purification Kit (Promega). DNA extraction was performed following a previously described protocol (López Moreno et al., 2010).
2.2 RAPD analysis
The extracted plant DNA was quantitated using a NanoDrop 1000 spectrophotometer (Thermo Scientific) and diluted to 50 ng/μL. PCR reactions were prepared using 50 ng of genomic DNA as a template, GoTaq® Green Master Mix (Promega), and 10 pmol of OPB-8 primer (sequence: GTCCACACGG). Afterwards, electrophoretic separation of the PCR products was carried out in a 1.6% agarose gel in trisborate EDTA (TBE) buffer. The gel was visualized and photographed using a Gel Doc XR System running Quantity One Software (BioRad).
3 Results and discussion
In this study, we focused on the potential genotoxic effects exerted on zucchini plants after exposure to TiO2 NPs. Genotoxicity was inferred from changes in the genomic DNA amplicon pattern, which can be interpreted as direct DNA alteration, such as recombination, mutagenesis, or other deleterious effects. The resulting changes to genomic DNA were visualized on an agarose gel after a RAPD assay, where they manifested as the appearance or disappearance of amplicon DNA or as changes in the abundance of amplicon DNA. These were indicators of genetic dissimilarities between controls and treatments due to TiO2 NP exposure. In this work, any change in the genomic DNA amplicon pattern was scored as a genotoxic change.
Genomic DNA was extracted from four individual plants and assayed by RAPD and agarose gel electrophoresis, both before (Fig. 1, lanes 1, 3, 6, and 8) and after (Fig. 1, lanes 2, 4, 7, and 9) NP exposure. A 100 base pairs (bp) marker was used as a control to determine the length of the DNA fragments (Fig. 1, lane 5). In the gel shown in Fig. 1, changes in the DNA banding pattern can be seen at multiple places. One such altered band appears at about 950 bp (Fig. 1, green rectangle) and is conserved among the four plants. While this band appears at similar intensities in untreated and treated samples from plant 1 (Fig. 1, lanes 1 and 2), it appears to decrease in abundance after treatment in the samples from plants 2 through 4 (Fig. 1; compare lanes 3, 6, and 8 to lanes 4, 7, and 9). Other changes in genomic DNA amplicon profiles can be observed in individual plants (indicated in Fig.1 by arrows). However, the question of whether any of the observed changes are deletions, inversions or some other genetic alteration can be answered only by a deeper analysis of a specific amplicon band. Several studies suggest that TiO2 NPs are toxic. Kang et al. (2008) suggested that TiO2 NPs induce DNA breakage in peripheral blood lymphocytes. Therefore, the changes we observed could indicate such breakage, although further analysis is required to determine the nature of the change. TiO2 NPs have also been reported to be toxic to plants. Servin et al. (2013) reported that TiO2 NPs induced macromolecular changes in cucumber fruit. In addition, there is evidence that TiO2 in the nanoanatase form increases antioxidant stress in spinach chloroplasts (Zheng et al., 2008). Moreover, in A. thaliana, TiO2 induced changes in gene expression at the mRNA level, as measured by microarray analysis (Landa et al., 2012). Landa et al. (2012) reported that TiO2 had a ‘mild influence’ on the expression of stressresponse genes. The ‘mild influence’ of TiO2 exposure on A. thaliana underscores this plant’s ability to defend its transcriptome against the genotoxic nature of TiO2 and possible DNA damage induced by TiO2. TiO2 has been shown to induce the formation of reactive oxygen species (ROS) in cells (Xue et al., 2010), which can lead to changes in gene expression in response to stress. TiO2 has also been shown to physically damage DNA (breaking strands and oxidizing purines) and upregulate stress response gene expression as a consequence of ROS generation (Petković et al., 2011). These TiO2induced changes to the cell will have downstream effects on the RAPD patterns generated and are considered genotoxic (causing alteration to genomic DNA integrity, which could lead to cell death or carcinogenesis).

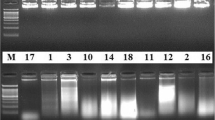
RAPD profiles from the roots of individual zucchini plants treated with TiO2 NPs at 50 mg/L
Genomic DNA from individual plants was extracted before (controls; lanes 1, 3, 6, and 8) and after (50 mg/L; lanes 2, 4, 7, and 9) treatment with TiO2 NPs suspension; RAPD profiles were generated from DNA extracts using primer OPB-3; lane 5 is a 100-bp DNA marker (100–1500 bp) for DNA size estimation
Next, we examined the effects of TiO2 NPs exposure on a population of zucchini plants. Fig. 2 shows the results from RAPD analysis and agarose gel electrophoresis. Nine control samples (Fig. 2, columns 2–10) and only seven treated samples (Fig. 2, lanes 11–17) are shown, because genomic DNA extraction for two of the treated plants did not yield enough DNA to analyze, based on quantitation by UV spectroscopy. A 100 bp marker (Fig. 2, lane 1) was included to determine the approximate size of the DNA fragments.

RAPD profiles from the roots of a population of zucchini plants treated with TiO2 NPs at 0 (control) and 50 mg/L
RAPD profiles were generated using primer OPB-3; first lane from bottom to top in all gels is a 100-bp DNA marker (100–1500 bp)
Bands that appeared or disappeared due to the NP treatment are indicated by colored rectangles. One band of interest, about 1200 bp in length, was present in the control samples (Fig. 2, lanes 2–10, green rectangle) but absent from the treated plants. A second band of about 400 bp was also present in the controls and absent from the NPtreated plants (Fig. 2, lanes 2–10, blue rectangle). Finally, a new band of about 650 bp did not appear in the control samples but was present in all the treated plants (Fig. 2, lanes 11–17, red rectangle). For all the samples, the only difference between the treatment conditions of these 18 plants was the presence or absence of NPs; thus, we can conclude that the changes in the DNA are being produced as an effect of the TiO2 NPs at the population level. The reason for the difference in RAPD profiles among the control plants is that each plant will have different DNA sequences.
Although there is overwhelming evidence of DNA change shown in these gels, it is not yet possible to interpret the results in terms of specific genetic alterations to the plant genomic DNA or its expression pattern. Additional analysis, such as cloning and sequencing of the bands of interest, will be required to determine the exact nature of these changes. Due to the lack of genomic sequence information available for C. pepo, it is not yet feasible to perform such techniques on this plant. However, it might be instructive to compare the sequences of the altered genes to the genomic sequences of other plants present in databases, in case the alterations occur in conserved regions of the genome.
Acknowledgements
This study was based upon work supported by the National Science Foundation and the Environmental Protection Agency under Cooperative Agreement (No. DBI0830117). Any opinions, findings, and conclusions or recommendations expressed in this study are those of the author(s) and do not necessarily reflect the views of the National Science Foundation or the Environmental Protection Agency. This work has not been subjected to Environmental Protection Agency review and no official endorsement should be inferred. We thank the UTEP Border Biomedical Research Center (BBRC) Genomic Analysis Core Facility and Ana Betancourt for DNA sequencing. Vincent U. GANT Jr. was supported by the National Institutes of Health (NIH)funded Research Scholars Program Grant (No. 2R25GM069621-09) to Dr. Renato J. AGUILERA (UTEP). This project was also supported by the National Institute on Minority Health and Health Disparities (NIMHD) Grant (No. 8G12MD007592), a component of the NIH. The contents of this study are solely the responsibility of the authors and do not necessarily represent the official views of NIMHD or NIH. The authors also acknowledge the USDA (No. 2011-38422-30835) and the National Science Foundation (No. CHE-0840525). Dr. Jorge L. GARDEATORRESDEY acknowledges the Dudley family for the Endowed Research Professorship, the Academy of Applied Science/US Army Research Office, Research and Engineering Apprenticeship Program (REAP) at UTEP (No. W11NF-10-2-0076, subgrant 13-7), and Science and Technology Acquisition and Retention Program of the University of Texas System.
References
Amaral, M.J., Carretero, M.A., Bicho, R.C., et al., 2012. The use of a lacertid lizard as a model for reptile ecotoxicology studies—part 1 field demographics and morphology. Chemosphere, 87 (7):757–764. [doi:10.1016/j.chemosphere.2011.12.075]
Cenkci, S., Yildiz, M., Ciğerci, İ.H., et al., 2009. Toxic chemicalsinduced genotoxicity detected by random amplified polymorphic DNA (RAPD) in bean (Phaseolus vulgaris L.) seedlings. Chemosphere, 76 (7):900–906. [doi:10.1016/j.chemosphere.2009.05.001]
Darlington, T.K., Neigh, A.M., Spencer, M.T., et al., 2009. Nanoparticle characteristics affecting environmental fate and transport through soil. Environmental Toxicology and Chemistry, 28 (6):1191–1199. [doi:10.1897/08-341.1]
Gottschalk, F., Sonderer, T., Scholz, R.W., et al., 2009. Modeled environmental concentrations of engineered nanomaterials (TiO2, ZnO, Ag, CNT, fullerenes) for different regions. Environonmental Science & Technology, 43 (24): 9216–9222. [doi:10.1021/es9015553]
Hong, J., Peralta-Videa, J.R., Gardea-Torresdey, J.L., 2013. Nanomaterials in agricultural production: benefits and possible threats. Shamin, N., Sharma, V.K. (Eds.), Sustainable Nanotechnology and the Environment: Advances and Achievements. ACS Symposium Series, 1124:73–90.
Johnson, A.C., Bowes, M.J., Crossley, A., et al., 2011. An assessment of the fate, behaviour and environmental risk associated with sunscreen TiO2 nanoparticles in UK field scenarios. Science of the Total Environment, 409 (13): 2503–2510. [doi:10.1016/j.scitotenv.2011.03.040]
Kang, S.J., Kim, B.M., Lee, Y.J., et al., 2008. Titanium dioxide nanoparticles trigger p53 mediated damage response in peripheral blood lymphocytes. Environmental and Molecular Mutagenesis, 49 (5):399–405. [doi:10.1002/em. 20399]
Keller, A.A., Wang, H., Zhou, D., et al., 2010. Stability and aggregation of metal oxide nanoparticles in natural aqueous media. Environmental Science and Technology, 44 (6):1962–1967. [doi:10.1021/es902987d]
Keller, A.A., McFerran, S., Lazareva, A., et al., 2013. Global life cycle releases of engineered nanomaterials. Journal of Nanoparticle Research, 15 (6):1692. [doi:10. 1007/s11051-013-1692-4]
Kumari, M., Mukherjee, A., Chandrasekaran, N., 2009. Genotoxicity of silver nanoparticles in Allium cepa. Science of the Total Environment, 407 (19):5243–5246. [doi:10. 1016/j.scitotenv.2009.06.024]
Landa, P., Vankova, R., Andrlova, J., et al., 2012. Nanoparticlespecific changes in Arabidopsis thaliana gene expression after exposure to ZnO, TiO2, and fullerene soot. Journal of Hazardous Materials, 241– 242: 55–62. [doi:10.1016/j.jhazmat.2012.08.059]
Liu, W., Li, P.J., Qi, X.M., et al., 2005. DNA changes in barley (Hordeum vulgare) seedlings induced by cadmium pollution using RAPD analysis. Chemosphere, 61 (2):158– 167. [doi:10.1016/j.chemosphere.2005.02.078]
Liu, W., Yang, Y.S., Li, P.J., et al., 2009. Risk assesment of cadmiumcontaminated soil on plant DNA damage using RAPD and physiological indices. Journal of Hazardous Materials, 161 (2–3):878–883. [doi:10.1016/j.jhazmat. 2008.04.038]
López-Moreno, M.L., de la Rosa, G., Hernández-Viezcas, J.Á., et al., 2010. Evidence of the differential biotransformation and genotoxicity of ZnO and CeO2 nanoparticles on soybean (Glycine max) plants. Environmental Science and Technology, 44 (19):7315–7320. [doi:10.1021/ es903891g]
MacPherson, J.M., Gajadhar, A.A., 1992. Random amplified polymorphic DNA. Parasitology Today, 8 (7):235. [doi:10.1016/0169-4758(92)90120-Q]
Peralta-Videa, J.R., Gardea-Torresdey, J.L., Go-mez, E., et al., 2002. Effect of mixed cadmium, copper, nickel and zinc at different pHs upon alfalfa growth and heavy metal uptake. Environmental Pollution, 119 (3):291–301. [doi:10. 1016/S0269-7491(02)00105-7]
Petković, J., Zegura, B., Ste-vanović, M., et al., 2011. DNA damage and alterations in expression of DNA damage responsive genes induced by TiO2 nanoparticles in human hepatoma HepG2 cells. Nanotoxicology, 5(3):341–353. [doi:10.3109/17435390.2010.507316]
Robichaud, C.O., Uyar, A.E., Darby, M.R., et al., 2009. Estimates of upper bounds and trends in nanoTiO2 production as basis for exposure assessment. Environmental Science and Technology, 43 (12):4227–4233. [doi:10. 1021/es8032549]
Roco, M.C., 2011. The long view of nanotechnology development: the national nanotechnology initiative at 10 years. Journal of Nanoparticle Research, 13 (2):427–445. [doi:10. 1007/s11051-010-0192-z]
Roco, M.C., Mirkin, C.A., Hersam, M.C., 2010. Nanotechnology research directions for societal needs in 2020: retrospective and outlook. Available from http://www.nano.gov/node/797 [Accessed on Oct. 29, 2013].
Servin, A.D., Morales, M.I., Castillo-Michel, H., et al., 2013. Synchrotron verification of TiO2 accumulation in cucumber fruit: a possible pathway of TiO2 nanoparticle transfer from soil into the food chain. Environmental Science and Technology, 47 (20):11592–11598. [doi:10. 1021/es403368j]
Williams, L., Adams, W., 2007. Nanotechnology Demystified. McGraw-Hill, New York, USA, p.4–5.
Xue, C., Wu, J., Lan, F., et al., 2010. Nano titanium dioxide induces the generation of ROS and potential damage in HaCaT cells under UVA irradiation. Journal of Nanoscience and Nanotechnology, 10 (12):8500–8507. [doi:10. 1166/jnn.2010.2682]
Zheng, L., Su, M., Wu, X., et al., 2008. Antioxidant stress is promoted by nano-anatase in spinach chloroplasts under UVB radiation. Biological Trace Element Research, 121 (1):69–79. [doi:10.1007/s12011-007-8028-0]
Author information
Authors and Affiliations
Corresponding authors
Additional information
Project supported by the National Science Foundation (NSF) and the Environmental Protection Agency under Cooperative Agreement (No. DBI-0830117), the National Institutes of Health (NIH) Grant (No. 2R25GM069621-09), the National Institute on Minority Health and Health Disparities (NIMHD) Grant (No. 8G12MD007592), the United States Department of Agriculture (USDA) Grant (No. 2011-38422-30835), and the Research and Engineering Apprenticeship Program (REAP) Grant (No. W11NF-10-2-0076), USA
Rights and permissions
About this article

Cite this article
Moreno-Olivas, F., Gant, V.U., Johnson, K.L. et al. Random amplified polymorphic DNA reveals that TiO2 nanoparticles are genotoxic to Cucurbita pepo . J. Zhejiang Univ. Sci. A 15, 618–623 (2014). https://doi.org/10.1631/jzus.A1400159
Received:
Revised:
Published:
Issue Date:
DOI: https://doi.org/10.1631/jzus.A1400159
Key words
- Random amplified polymorphic DNA (RAPD)
- Titanium dioxide (TiO2)
- Nanoparticles (NPs)
- Genomic DNA
- Zucchini