
Abstract
Background
Groundnut, Arachis hypogaea L., crop is an important oil seed crop in India and is prone to attack by numerous fungal and viral diseases, among the soil-borne diseases, stem rot caused by Sclerotium rolfsii Sacc. is economically important.
Main body of abstract
Incidence (86.6%) of the stem rot disease was recorded in Coimbatore district, Tamil Nadu, India, during the roving survey. For its management, an attempt was made to isolate bacterial endophyte from peanut seeds of different groundnut cultivars by culture-dependent analysis. Totally, 16 bacterial endophytes (endophytes groundnut (EGN) 1 to EGN 16 along with standards TNAU-Pf1 and EPC 5) were obtained and characterized through morphological, biochemical and molecular studies and also phytostimulation activities were performed. Among the isolates, EGN 1 and EGN 4 showed positive results for indole acetic acid (IAA), siderophore, phosphate solubilization and protease tests in vitro. The dual culture analysis showed inhibition rates of 60.1% (dual plate assay), 68.23% (filter paper disc assay) and 100% (triangle method of streaking and culture filtrate assay) for EGN 1 against S. rolfsii. Further, the crude metabolite assay showed 97.7% inhibition in EGN 1, followed by 87.7% in EGN 4. The roll towel study showed a high vigour index of 4286.7 in EGN 1; hence, this isolate was chosen. Further, thin-layer chromatography (TLC) analysis showed various bands at 0.72 Rf, whereas GC-MS analysis indicated the prominent peaks of hexadecanoic acid and cis-vaccenic acid that may responsible for antifungal activity. In a molecular approach, the genomic DNA of EGN 1 strain was used to amplify a 1200 bp PCR-fragment and sequenced.
Short conclusion
The overall outcome of this study showed that Pseudomonas spp. EGN 1 had a great potential as a bio-stimulant and biocontrol agent to manage effectively the stem rot in peanut.
Similar content being viewed by others


Background
Groundnut crop known as “King of oilseeds” is an important oil seed crop in India and the fourth most important edible oil source in the world. It originated in South America and contains 48–50% oil and 26–28% protein (Janila et al., 2013). In India, Gujarat is the largest producer contributing 25% of the total production, followed by Tamil Nadu (22.48%). It is valued as fourth most important source of edible oil and third most important source of vegetable protein. It is also an important agricultural export commodity from India. Several factors are responsible for lower productivity among which leaf spot, collar rot, stem rot and bud necrosis, etc. are very important. Out of them, stem rot caused by Sclerotium rolfsii Sacc. is a major problem and is an important soil-borne pathogen. Characteristic symptoms include production of brown-coloured mustard-sized sclerotia, resting structure (0.1 to 3.0 mm) and causes pod yield losses up to 80% under favourable conditions (Deepthi, 2014). As an alternative to judicious use of fungicides, which is creating adverse effect on environment, eco-friendly disease management strategies have to be evolved, especially using endophytes from seeds. Seed endophytes resulting in colonization of embryo and endosperm are gaining importance. Some of these bacteria can benefit plant growth and defence against abiotic and biotic stress, as well as vertical transmission of endophytic bacteria in seeds has also been reported (Frank et al., 2017). This process of vertical transmission results in a weakening of microbial pathogenic strength in order to support plant growth and development. Truyens et al. (2015) reviewed studies on seed endophytes and reported that the bacteria found in seeds mostly belong to the genera, especially Bacillus and Pseudomonas, and also Paenibacillus, Micrococcus, Staphylococcus, Pantoea and Acinetobacter. Many seed-borne bacteria produce plant hormones such as auxins, cytokinins and gibberellins; others produce ACC deaminase. This confers plant protection by the induction of plant defence mechanisms and production of lytic enzymes and antibiotics. Moreover, alongside their vital role in plant growth and defence, these seed-borne bacterial and fungal endophytes benefit the host plants through providing their progeny with valuable endosymbionts. Plant growth promotion by endophytic bacteria-inoculated plants has been frequently reported and this has attracted research toward their use in phytoremediation or as bioinoculants in agriculture.
The study aimed to isolate and characterize endophytes that naturally colonized peanut seeds and their effect on soil-borne plant pathogen.
Main text
Materials and methods
Survey and stem rot pathogen
Survey was conducted in stem rot-infected fields during the growing seasons in different groundnut cultivars, namely COG 0549, VRI 7, C0G 0539, TMV 7, TMV 2, C0 6, TAG 24, CG 20, AK 303, TG 37, K6, COG 0537, Dharani, TMV 14, BSR 2, CO 7 and VRI 8. The cultivars were grouped into group 1 (resistant), group 2 (moderately resistant), group 3 (susceptible) and group 4 (highly susceptible cultivars) based on percent disease incidence (PDI) in groundnut cultivars. The pathogen namely Sclerotium was isolated from the infected stem regions of groundnut through tissue segmentation method (Adhilakshmi et al., 2014) from Aliyar, Coimbatore, Sivaganga and Madurai region.
Seed bacterial endophytes
Seeds were surface disinfected in 5% commercial bleach and 0.01% Tween 20 for 10 min and rinsed 10 times with sterile distilled water. The efficiency of the procedure was checked using an aliquot of the water used and in the final wash was plated on nutrient agar. Surface-sterilized seeds were split into two and placed in a nutrient agar plates at an inverted position and incubated at 28 °C in the dark for 24–48 h. The rest of the isolates were obtained by plating dilutions of seed macerates on nutrient agar (NA). Surface-sterilized seeds were triturated with a pestle and mortar in PBS (phosphate-buffered saline, pH 7.0) and 60 μl of the suspension were placed onto NA, followed by incubation for 2 days at 28 ± 2 °C. After 24 h of incubation, colonies with distinct growth features were isolated (Xu et al., 2014).
In vitro antagonistic activity
All of the endophytic bacterial isolates were evaluated against S. rolfsii. A full loop of endophytic bacteria isolates (48 h old) grown on NA was streaked at the distance of 1 cm from the periphery of a Petri plate. Nine millimetre mycelial discs of S. rolfsii were taken from the 4-day-old culture and placed at the other end of the Petri plate containing potato dextrose agar (PDA). The plates were incubated for 7 days at 28 ± 2 °C (room temperature) and the radial growth of S. rolfsii was observed for the development of inhibition zone and the inhibition of mycelial growth was assessed by measuring the radial growth of S. rolfsii in the treated plate. The mycelial disc of S. rolfsii was also inoculated on the plate without endophytic bacteria as a control (Bodhankar et al., 2017). Four replicate plates were measured per isolate and the experiment was repeated twice to confirm the results. The percentage inhibition radial growth of treatments compared to the control was calculated as follows:
Where,: Rc is radial growth of S. rolfsii in the control plate and Rt is radial growth of S. rolfsii toward the antagonist in a dual culture plate.
Growth promotion assay by roll towel method
Effect of bacterial endophytes on seed germination was assessed by standard roll towel method (ISTA, 1993). Bacteria cultured in a LB broth for 48 h were centrifuged at 10,000 rpm for 5 min. The supernatant was discarded and the pellet was mixed with sterile water. Groundnut seeds were surface sterilized by 0.1% mercuric chloride for 5 min and rinsed with sterile distilled water. The surface-sterilized seeds were blot dried and soaked in the bacterial suspension for 4 h and sterile water was served as a control. Then, the seeds were placed in wet blotters and incubate at room temperature. Each treatment was replicated 3 times and at the 10th day, germination percentage, root length and shoot length was measured randomly (Agrawal and Agrawal, 2013). Plant growth promotion of groundnut seedlings was assessed using the vigour index (VI).
Further, total root length, root tips, segment and forks were assessed using a rhizoscanner instrument available at DARS, Chettinad, TNAU, Coimbatore.
Filter paper disc assay
Antimicrobial activity of crude extracts was examined using the disc diffusion method. Sterile filter paper discs of 6 mm diameter were soaked in crude extract and placed on the solidified PDA medium at the 4 corners of the plates at equal distance. Mycelial discs (9 mm) of S. rolfsii were collected from the 4-day-old culture and placed at the centre of the petriplate and incubated for 7 days at room temperature (Ramyabharathi and Raguchander, 2014). Diameters of the growth inhibition areas were measured along with control plate.
Morphological and biochemical characterizations
Seed endophytic bacteria were morphologically characterized based on colony formation, margin, elevation, texture, colour, surface and opacity. Effective strains were classified as gram positive or negative based on Gram staining. The antagonistic strains were identified to the species level based on the biochemical tests namely KOH, citrate utilization, indole tryptone, catalase, methyl red and vokes proskauer test, urease and gelatin liquefaction test (Bhoonobtong et al., 2012).
Molecular characterization
Genomic DNA extraction of the selected bacterial isolates was done using cetyltrimethylammoniumbromide (CTAB) method. The 16S rRNA gene was amplified using the universal primers Fd1 (5′AGAGTTTGATCCTGGCTCAG-3′) and Rd1 (5′AAGGAGGTGATCCAGCC-3′) (Dhouib et al., 2019) and the primers were obtained from Sigma Aldrich Chemicals Pvt. Ltd, Bangalore, India. The concentration of DNA was measured in NanoDrop followed by loading in 0.8% agarose gel electrophoresis and its concentration was observed using UV transilluminator (Inderiati and Franco, 2008). Thermocycler was used to perform PCR, using cycling conditions; initial denaturation at 94 °C for 5 min, denaturation at 94 °C for 30 s, annealing at 53 °C for 1 min, extension at 72 °C for 2 min and with a final extension at 72 °C for 7 min consisted of 30 cycles. Final products of PCR were analysed by 1.5% agarose gel electrophoresis. Based on the analysis, the amplified 16S rRNA PCR product of endophytes groundnut (EGN) 1 and EGN 4 was sent for custom sequencing (Barcode BioSciences, India). The obtained sequences were then compared for homology with those present in National Center for Biotechnology Information (NCBI), using Basic Local Alignment Search Tool (BLAST) analysis. The phylogenetic trees were generated using the neighbour-joining method by using MEGA 7.0 software.
Phytostimulation activities
IAA production
Salkowski reagent was used for IAA detection and quantification. The bacterial strains were grown in 100 ml nutrient broth with and without 0.5 g/l tryptophan (precursor of IAA) in darkness (Gordon and Paleg, 1957) for 3 days at 28 °C at 110 rpm. Bacterial culture suspensions were centrifuged (30 min at 8000 rpm) and 0.2 ml of the supernatant was mixed with 1 ml Salkowski’s reagent (50 ml 35% HClO4, 1 ml 0.5 M FeCl3). After 30 min, a pink colour was developed, which indicate IAA production. The absorbance of pink colour was read at 530 nm in spectrophotometer.
Siderophore production
Siderophore production of the endophytic bacterial strains was examined, using Chrome azurol S (CAS) agar medium following the protocol of Khamna et al. (2009). Five days post incubation at room temperature (28 ± 2 °C), orange halo around the colony indicated siderophore production. The experiment was carried out with 6 replications and repeated twice.
Phosphate solubilization
Pikovskayas medium was employed for detecting solubilization of tricalcium phosphate [Ca3(PO4)2] activity by antagonistic endophytic bacterial isolates. After 5 days of incubation at (28 ± 2 °C), phosphate-solubilizing activity (P-solubilizing positive) was observed by the development of a clear zone around the colonies (Pikovskaya, 1948). The experiment was carried out with 6 replications and repeated twice.
Protease production
Qualitative proteolytic activity of the selected endophytic bacterial strains was determined on skim milk agar medium (El-Deeb et al., 2012). Four days post incubation at (28 ± 2 °C), proteolytic activity was observed by the development of a halo clear zone around the colony of endophytic bacterial isolates. Six replicate plates were conducted and the experiment was repeated twice.
Cellulase production
Selected bacterial endophytes were spot inoculated on plates containing carboxy methyl cellulose (CMC) agar (0.05% K2HPO4, 0.025% MgSO4, 0.188% CMC sodium salt, 0.02% Congo red, 1.5% agar and 0.2% gelatin). The plates were incubated at 28 °C for 48 h. After incubation, CMC plates were observed for the presence of clear zone around the colonies (Kasana et al., 2008).
Culture filtrate test
Based on the in vitro test, selected endophytic bacterial isolates with high antagonistic activity were further investigated for their ability to produce inhibitory metabolites. The test was conducted using the method described by Intana et al. (2008). Erlenmeyer flask 250 ml containing 100 ml Luria-Bertani broth (LB) were inoculated with the selected bacterial isolates and incubated at 25 ± 2 °C on a rotary shaker (150 rpm) for 72 h. It was centrifuged at 10,000 rpm for 10 min and the pellets were discarded and the supernatant was filtrated with germ filter (d = 0.25 μm). The filtrate from each isolate was incorporated into sterilized PDA in ratio 2:1, and 25 ml of the amended agar was poured into each Petri plate and allowed to solidify. Mycelial disc of S. rolfsii (9 mm) was centrally inoculated on each plate. Sterilized water was used as the control. The diameter of the mycelial growth of S. rolfsii was measured over 7 days in comparison to the control plate (Alsultan et al., 2019).
Production of thermostable antifungal compounds
Protocol described by Ferraz et al. (2016) was used to assess the production of thermostable antifungal compounds. A loopful of 72-h-old culture inoculum was transferred to 250 ml Erlenmeyer flasks containing 100 ml PDA medium, followed by incubation at 150 rpm for 72 h. From that, 10 ml aliquots of each isolate were transferred to Erlenmeyer flasks containing 90 ml PDA medium. Then, it was subjected to autoclaving at 121 °C for 30 min. After sterilization, it was poured into a Petri plate and following solidification, a 9-mm culture disc of S. rolfsii was transferred to the centre of each plate. S. rolfsii that was grown on PDA medium containing sterile water instead of the metabolites is kept as a control. The plates were incubated in a BOD chamber at 25 ± 1 °C and constant light for 10 days. From two perpendicular directions, mycelial growth of S. rolfsii was measured in comparison to the control plate.
Effect of metabolites on the growth of Sclerotium rolfsii
Agar well diffusion assay modified by Islam et al. (2012) was used to determine the antagonistic activity of metabolites. Twenty millilitres of PDA medium was poured into sterilized Petri plates and after solidification, mycelia disc (9 mm) of test pathogen was placed at the centre. A well of 7 mm diameter was made by punching with a sterile cork borer on the solidified agar plate on the corner in 4 places at an equal distance. Then, the extracted metabolites were poured into the agar wells separately (100 ul) and incubated for 72 h at 28 ± 2 °C. After incubation, the inhibitory activity was assessed by measuring the radial growth of S. rolfsii in the treated plate compared to control.
TLC and GCMS analysis
Bacterial strains were cultured on a LB broth and ethyl acetate was used for the extraction of crude antibiotics. In TLC analysis, methanol was used to dissolve the dried sample and the plates were developed with isopropanol: ammonia: water (8:1:1). Five microlitres of sample were spotted onto silica gel plate and the plate was visualized using UV transilluminator at short wave length (254 nm). The Rf values were calculated for specific antibiotics (Prabhukarthikeyan and Raguchander, 2016). Gas chromatography/mass spectrometry (GCMS) analysis was used for the selected endophytic bacterial isolates with high antifungal effects for the production of volatile antifungal metabolites as described by Hassi et al. (2012). The compounds were identified based on a comparison of their mass spectra data and retention time as well as with the existing analytical data of specially synthesized reference compounds database.
Statistical analysis
Statistical analyses were done using IRRISTAT version 92 developed by the International Rice Research Institute Biometrics Unit, the Philippines (Gomez and Gomez, 1984), and the percentage values of the disease index were arcsine transformed. Data were subjected to ANOVA at significant level (P < 0.05) and means were compared by Duncan’s multiple range test (DMRT).
Results and discussion
Survey and isolation of the pathogen
The survey of the pathogen revealed that the stem rot incidence of A. hypogaea was 86.6% in Coimbatore, 32% in Aliyar, 26% in Madurai and 21% in Sivaganga districts (Fig. 1 and Table 1). Also, 7 cultivars (COG 0539, TMV 14, CO 7, VRI 8, COG 0537, CO 6, Dharani) showed high resistance to the pathogen, 3 cultivars (VRI 7, BSR 2, TK 37) moderate resistance, 3 cultivars (COG 0549, CG 20, K6) moderately susceptible and 4 cultivars (TMV 7, TMV 2, TAG 24, AK 303) were highly susceptible under field conditions. Five isolates, obtained from 4 different regions of Coimbatore, Aliyar, Madurai and Sivaganga and the Sclerotium were isolated from the infected plant samples and the abundant whitish mycelium was observed in a distinctive fan-like pattern. It produced mustard-sized sclerotia of 0.1–0.3 mm after 3–4 days of incubation. Similarly, Rakh et al. (2011) reported that isolation of S. rolfsii from groundnut was carried out and maintained on PDA slants incubated at 28 °C.

Stem rot infected fields in (a). Coimbatore (b). Sivagangai (c). Madurai (d). Aliyar (e). Sclerotium in PDA
Isolation of seed bacterial endophytes
For the management of stem rot disease in groundnut, bacterial endophytes isolated from the inner tissues of seeds were used as an alternative eco-friendly strategy. Bacterial endophytes isolated from leaves, roots and stems have been reported by many authors (Saini et al., 2015 and Upreti et al., 2015) but endophytes isolated from the seeds have not been studied. Hence, the present study aimed to isolate seed bacterial endophytes and exploiting for the management of stem rot disease in groundnut. In this context, culture-dependent studies by split plot and serial dilution technique (Fig. 2) were undertaken on 17 groundnut cultivars VRI 2, JL 24, CO 6, VRI 8, TAG 24, CO 1, TMV 1, TG 37, CO 4, TMV 2, K 6, TMV 7, TMV 14, CO 7, ALR 3, BSR 9 and TMV 10 for bacterial endophytes isolation and the isolates namely EGN 1 to EGN 16 were isolated from seeds along with standards TNAU-Pf 1 and EPC 5 were used. Similarly, Sobolev et al. (2013) isolated the endophytic bacteria from groundnut seeds by split technique. Xu et al. (2014) identified 84 endophytic bacteria from the seeds of tomato by serial dilution technique.

Isolation of seed bacterial endophytes by (a) split and (b) serial dilution technique
Morphological, biochemical and molecular characterization
The colony of seed bacterial endophytes varied from circular to irregular with raised, flat and convex margin was observed and their texture varied from brittle to dry with rough, smooth and glistening surfaces. Based on morphological observation, the majority of colony colour ranged from whitish yellow to dull white and characterized biochemically through various biochemical tests (Table 2). The effective isolates namely EGN 1 and EGN 4 showed positive results for indole tryptone, gelatin liquefaction, urease, hydrogen peroxide and VP test and gram staining is also carried out for effective isolates along with standard EPC 5. The effective isolates showed negative reaction whereas standard EPC 5 showed positive. Similarly, Rajendran et al. (2007) characterized the endophytic species as Bacillus in cotton by various biochemical tests like Gram staining, KOH test, utilization of citrate, catalase test, starch hydrolysis, gelatin hydrolysis and methyl red test. Molecular characterization of effective isolates was carried out by extracting the DNA using CTAB method and the concentration of genomic DNA was measured, using NanoDrop. In a molecular approach, the genomic DNA of strains EGN 1 to EGN 8, TNAU-Pf 1 and EPC 5 were used to amplify a fragment coding its 16S rRNA and the isolates were amplified at 1.2 kb (approx). Erjaee et al. (2020) also reported that the isolates M11, TR1, TR12, TR13, TR14 and A11 obtained from herbal plants were 100% identical to B. pumilus. After amplification, the best isolate EGN 1 along with EGN 4 was sequenced and phylogenetic tree was generated in MEGA 7.0 software for 16S rRNA gene sequences for EGN 1 and the results revealed that Pseudomonas spp. EGN 1 isolate (MT526251) showed 55% similarity with Pseudomonas aeruginosa Mexico strain (Fig. 3). Whereas the sequence results of EGN 4 matched with Acinetobacter spp. Similarly, Staphylococcus is a plant-associated microbe, but recently various studies have reported it as endophytic bacteria (Haidar et al., 2018).

Phylogenetic tree of the 16S rRNA sequences of Pseudomonas spp.
In vitro study
The antagonistic effect of seed bacterial endophytes against S. rolfsii was assessed on dual culture technique. The results showed that seed bacterial endophytes recorded 25–60.1% of inhibition against S. rolfsii in vitro (Table 3). Further, triangle method of streaking showed EGN 1, EGN 2, EGN 3, EGN 4, EGN 5 and TNAU-Pf 1 and EPC 5 were maximum inhibition of 100%, whereas isolates EGN 6 and EGN 7 did not show any inhibition. These seed endophytes showed antagonistic potential against S. rolfsii through dual culture and filter paper disc assay. Correspondingly, Sahu et al. (2019) screened for the antagonistic potential of bacterial endophytes obtained from tomato against S. rolfsii, 2 endophytic isolates inhibited the radial growth of S. rolfsii by more than 50%. Kefi et al. (2015) also reported that the cell-free supernatant of the 4 strains inhibited the growth of Botrytis cinerea versus control through disc diffusion method and the strongest activity at a crude extract concentration of 7.81 g ml–1 was exhibited by BF11 strain.
Growth promotion by roll towel method
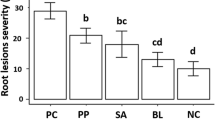
Results of growth promotion studies showed that EGN 1 increased the vigour index of groundnut seedlings up to 4286.7, followed by TNAU-Pf 1 (4040.0), EPC 5 (2599.74) and EGN 4 (2895.35) through roll towel method. The isolate EGN 1 also recorded high germination (86.67%) with increase root length (25.0 cm) and shoot length (24.5 cm). In the rhizoscanner study, total root length, root tips, segment and forks were higher in EGN 1 (3370 mm, 425 root tips, 501 forks) and EGN4 (2786 mm, 382 root tips, 501 forks) than the control (1129 mm, 184 root tips, 463 forks) (Table 4 and Fig. 4). Similarly, Sundaramoorthy et al. (2012) reported that Pf1 + EPCO16 considerably increased the vigour index of tomato seedlings through roll towel method up to 3840.70, followed by Pf1 (3143.4), Py 15 (2947.05), FP 7 (2989.30), EPC 5 (2814.3) and EPCO 16 (3225.7), respectively, when compared to control.

Growth promotion test by roll towel method and study using rhizoscanner
Filter paper disc assay
In this assay, EGN 4 showed the maximum inhibition (94.10), followed by EGN 1 (68.23%), TNAU-Pf 1 (57.60%), EGN 7 (52.94%), EPC 5 (40.20%) and EGN 2 (11.47%). The isolates, namely EGN 3, EGN 5 and EGN 6 did not show any antagonistic activity against S. rolfsii (Table 5). Antagonistic activity of effective seed bacterial endophytes (EGN 1 and EGN 4) against S. rolfsii through (I) dual plate assay, (II) triangle method of streaking and (III) filter paper disc assay is shown in Fig. 5. Similarly, Alsultan et al. (2019) reported that P. aeruginosa and C. proteolyticum were able to produce cellulase, lipase, protease and pectinase at varying levels and these enzymes participate in the antagonistic activity against Phytophthora palmivora causing black pod of cocoa.

(I) Dual plate assay (II) Triangle method of streaking and (III) Filter paper disc assay
Phytostimulation activities
The isolates namely EGN 1, EGN 3 and TNAU-Pf1 were high siderophore producers, whereas EGN 4, EGN 6, EGN 7 and EPC 5 were moderate producers and EGN 2 and EGN 5 were low producers. Four seed endophytic isolates EGN 1, EGN 3, EGN 4 and EPC 5 produced IAA (the hormone responsible of root growth), which was indicated by the production of pink to reddish pink colour in the broth after addition of Salkowski reagent. In case of phosphate solubilization, it was indicated by the formation of a clear halo zone. The isolates EGN 1 and TNAU-Pf 1 had the highest ability to solubilize the phosphate followed by EGN 2, EGN 4, EGN 6 and EGN 7. For the production of hydrolytic enzymes, cellulase and protease test was undertaken for the effective isolates. The results showed that 7 isolates had the ability to produce protease enzyme and 9 showed cellulase production, which indicated the hydrolytic enzyme production by endophytes (Table 6 and Fig. 6). Allu et al. (2014) also reported the siderophore production by the bacterial endophytes in chilli, while Wang et al. (2013) reported the growth promoting ability of endophytic bacteria, i.e. 48 isolates were able to produce 1AA and 3 isolates showed phosphate solubilization in peanut, nitrogen-fixing endophytic bacteria and their association in sugarcane (Cavalcante et al., 2007).

(I) Phosphate solubilisation : 1- EGN2, 2- EGN1, 3- EGN4, 4- EGN3, 5- EGN6, 6- EGN5,7- EPC5, 8- TNAU Pf1, 9- EGN7 (II) Cellulase test and (III) Protease test : 1- EGN 1, 2- EGN 2, 3- EGN 3, 4- EGN 4, 5- EGN 6, 6- EGN 7, 7- TNAU Pf 1, 8- EPC 5 (IV) Siderophore production : 1- EGN 1, 2- EGN 4
Culture filtrate assay
The culture filtrate assays showed that the isolates EGN 1, EGN 2, EGN 3, EGN 4, EGN 5, EGN 6, EGN 7, TNAU Pf1 and EPC 5 had a maximum inhibition rate of 100% through amendment of cell-free supernatants to agar plates. They exhibited the effectiveness of culture filtrate on stem rot pathogen (S. rolfsii) (Fig. 7).

Culture filtrate test – (I) (EGN 1) and (II) (EGN 4), Production of thermostable compounds - (III) (EGN 1) and (IV) (EGN 4), (V)- Control
Thermostable compounds and secondary metabolite production
The effective isolates EGN 1, EGN 4 and EPC 5 were analysed for their presence of thermo stable compounds after autoclaving at 121 °C for 30 min. The mycelial growth was measured in 2 perpendicular directions. It showed that the isolate EGN 1 and EGN 4 had the presence of thermo stable compounds (Fig. 7). Similarly, Kupper et al. (2020) reported that 8 isolates (ACB-AP3, ACB-83, ACB-76, ACB-70, ACB-84, ACB-82, ACB-72 and ACB-69) of Bacillus spp. produced thermo-resistant metabolites produced inhibitions ranging from 25 to 42%. The secondary metabolite production of effective isolates was assessed through agar well diffusion technique. The isolate EGN 1 showed 97.7% inhibition, followed by EPC 5 (94.4%), EGN 4 (87.7%), TNAU Pf1 (84.4%), EGN 2 (82.2%) and EGN 3 (76.6%). Similarly, Prabhukarthikeyan and Raguchander (2016) tested the antimicrobial activity of Pseudomonas fluorescens against Pythium and reported that crude antibiotics of P. fluorescens FP 7 inhibited the mycelial growth of 40 mm accounted for 55.55% followed by P. fluorescens Pf1 (52.22% inhibition).
TLC and GC-MS analysis
Thin-layer chromatography (TLC) was carried out for effective isolates and it showed different bands (Fig. 8). The Rf values were as follows: EGN 1 (0.33, 0.44, 0.52 and 0.72), EGN 3 (0.29, 0.53), EGN 4 (0.46, 0.53), TNAU-Pf 1 (0.46, 0.53) and EPC 5 (0.46, 0.53 and 0.73). The compounds which were identified through GC-MS analysis in EGN 1 were octadecanal, pyrrol, hexadecanoic acid, oleic acid, cis-vaccenic acid, harmine and pyrrolo compounds and EPC 5 had Pyrrolo[1,2-a]pyrazine-1,4-dione, hexahydro-3-(2-methylpropyl), pentadeconoic acid, hexadecanoic acid, harmine and pyrrol (Fig. 9). Alike, Pratiwi et al. (2017) determined metabolite compound of P. aeruginosa through TLC analysis, the 11th fraction showed antibacterial activity with the Rf value of 0.53.

Thin-layer chromatography (TLC) analysis

Identification of compounds through GC-MS analysis
Conclusion
From the culture-dependent analysis, the effective isolate Pseudomonas spp. EGN 1 proved to be the most promising bio agent for the management of the stem rot (Sclerotium rolfsii) in groundnut.
Availability of data and materials
All the data in this study have been presented in this manuscript.
Abbreviations
- PDI:
-
Percent disease incidence
- PDA:
-
Potato dextrose agar
- NA:
-
Nutrient agar
- VI:
-
Vigour index
- CTAB:
-
Cetyltrimethylammoniumbromide
- NCBI:
-
National Center for Biotechnology Information
- EGN:
-
Endophytes groundnut
References
Adhilakshmi M, Latha P, Paranidharan V, Balachandar D, Ganesamurthy K, Velazhahan R (2014) Biological control of stem rot of groundnut (Arachis hypogaea L.) caused by Sclerotium rolfsii Sacc. with actinomycetes. Archiv Phytopathol Plant Protect 47(3):298–311
Agrawal DPK, Agrawal S (2013) Characterization of Bacillus sp. strains isolated from rhizosphere of tomato plants (Lycopersicon esculentum) for their use as potential plant growth promoting rhizobacteria. Int J Curr Microbiol App Sci 2(10):406–417
Allu S, Kumar NP, Audipudi AV (2014) Isolation, biochemical and PGP characterization of endophytic Pseudomonas aeruginosa isolated from chilli red fruit antagonistic against chilli anthracnose disease. Int J Curr Microbiol App Sci 3(2):318–329
Alsultan W, Vadamalai G, Khairulmazmi A, Saud HM, Al-Sadi AM, Rashed O, Nasehi A (2019) Isolation, identification and characterization of endophytic bacteria antagonistic to Phytophthora palmivora causing black pod of cocoa in Malaysia. Eur J Plant Pathol 155(4):1077–1091
Bhoonobtong A, Sawadsitang S, Sodngam S, Mongkolthanaruk W (2012) Characterization of endophytic bacteria, Bacillus amyloliquefaciens for antimicrobial agents production. Int Conference Biol Life Sci 40:6–11
Bodhankar S, Grover M, Hemanth S, Reddy G, Rasul S, Yadav SK, Srinivasarao C (2017) Maize seed endophytic bacteria: dominance of antagonistic, lytic enzyme-producing Bacillus spp. 3 Biotech 7(4):232
Cavalcante JJV, Vargas C, Nogueira EM, Vinagre F, Schwarcz K, Baldani JI, Hemerly AS (2007) Members of the ethylene signaling pathway are regulated in sugarcane during the association with nitrogen-fixing endophytic bacteria. J Exp Botany 58(3):673–686
Deepthi KC (2014) Effect of potential biocontrol agents against Sclerotium rolfsii causing stem rot of groundnut. Int J Life Sc Bt Pharm Res 2(2):58–65
Dhouib H, Zouari I, Abdallah DB, Belbahri L, Taktak W, Triki MA, Tounsi S (2019) Potential of a novel endophytic Bacillus velezensis in tomato growth promotion and protection against Verticillium wilt disease. Biol Control 139:104092. https://doi.org/10.1016/j.biocontrol.2019.104092
El-Deeb B, Bazaid S, Gherbawy Y, Elhariry H (2012) Characterization of endophytic bacteria associated with rose plant (Rosa damascena trigintipeta) during flowering stage and their plant growth promoting traits. Journal of plant interactions 7(3):248–253
Erjaee Z, Shekarforoush SS, Hosseinzadeh S, Dehghani A, Winter D (2020) Identification of antifungal intracellular proteins of endophytic Bacillus pumilus by LC–MS/MS analysis. Int J Pept Res Ther. https://doi.org/10.1007/00989-020-10040-8
Ferraz LP, da Cunha T, da Silva AC, Kupper KC (2016) Biocontrol ability and putative mode of action of yeasts against Geotrichum citri-aurantii in citrus fruit. Microbiol Res 188:72–79
Frank AC, Saldierna Guzmán JP, Shay JE (2017) Transmission of bacterial endophytes. Microorganisms 5(4):70
Gomez KA, Gomez AA (1984) Statistical procedures for agricultural research. New York: Wiley
Gordon SA, Paleg LG (1957) Quantitative measurement of indole acetic acid. Physiol Plant 10:37–48
Haidar B, Ferdous M, Fatema B, Ferdous AS, Islam MR, Khan H (2018) Population diversity of bacterial endophytes from jute (Corchorus olitorius) andevaluation of their potential role as bioinoculants. Microbiol Res 208:43–53
Hassi M, El Guendouzi S, Haggoud A, David S, Ibnsouda S, Houari A, Iraqui M (2012) Antimycobacterial activity of a Brevibacillus laterosporus strain isolated from a Moroccan soil. Brazilian Journal of Microbiology 43(4):1516–1522
Inderiati S, Franco CM (2008) Isolation and identification of endophytic actinomycetes and their antifungal activity. J Biotechnol Res Trop Reg 1:1–6
Intana W, Yenjit P, Suwanno T, Sattasakulchai S, Suwanno M, Chamswarng C (2008) Efficacy of antifungal metabolites of Bacillus spp. for controlling tomato damping-off caused by Pythium aphanidermatum. Walailak J Sci Technol 5(1):29–38
Islam MR, Jeong YT, Lee YS, Song CH (2012) Isolation and identification of antifungal compounds from Bacillus subtilis C9 inhibiting the growth of plant pathogenic fungi. Mycobiology 40(1):59–65
ISTA (1993) Proceedings of International Seed Test Association, International rules for seed testing. Seed Sci Technol 21:1–152
Janila P, Nigam SN, Manish K, Pandey P, Nagesh Rajeev K, Varshney (2013) Groundnut improvement: use of genetic and genomic tools. Front Plant Sci 4:23. https://doi.org/10.3389/fpls.2013.00023
Kasana RC, Salwan R, Dhar H, Dutt S, Gulati A (2008) A rapid and easy method for the detection of microbial cellulases on agar plates using Gram’s iodine. Current microbiology 57(5):503–507
Kefi A, Slimene IB, Karkouch I, Rihouey C, Azaeiz S, Bejaoui M, Limam F (2015) Characterization of endophytic Bacillus strains from tomato plants (Lycopersicon esculentum) displaying antifungal activity against Botrytis cinerea Pers. World J Microbiol Biotechnol 31(12):1967–1976
Khamna S, Yokota A, Lumyong S (2009) Actinomycetes isolated from medicinal plant rhizosphere soils: diversity and screening of antifungal compounds, indole-3-acetic acid and siderophore production. World Journal of Microbiology and Biotechnology 25(4):649
Kupper KC, Moretto RK, Fujimoto A (2020) Production of antifungal compounds by Bacillus spp. isolates and its capacity for controlling citrus black spot under field conditions. World Journal of Microbiology and Biotechnology 36(1):7
Pikovskaya RI (1948) Mobilization of phosphorus in soil in connection with the vital activity of some microbial species. Microbiology 17:362–70.
Prabhukarthikeyan SR, Raguchander T (2016) Antifungal metabolites of Pseudomonas fluorescens against Pythium aphanidermatum. J Pure Appl Microbiolo 10(1):579–584
Pratiwi RH, Hidayat I, Hanafi M, Mangunwardoyo W (2017) Antibacterial compound produced by Pseudomonas aeruginosa strain UICC B-40, an endophytic bacterium isolated from Neesia altissima. Journal of Microbiology 55(4):289–295
Rajendran L, Samiyappan R, Raguchander T, Saravanakumar D (2007) Endophytic bacteria mediate plant resistance against cotton bollworm. Journal of Plant Interactions 2(1):1–10
Rakh RR, Raut LS, Dalvi SM, Manwar AV (2011) Biological control of Sclerotium rolfsii, causing stem rot of groundnut by Pseudomonas monteilii 9. Recent Res Sci Technol 3(3):26–34
Ramyabharathi SA, Raguchander T (2014) Efficacy of secondary metabolites produced by EPCO16 against tomato wilt pathogen. J Mycol Plant Pathol 44(2):148
Sahu PK, Singh S, Gupta A, Singh UB, Brahmaprakash GP, Saxena AK (2019) Antagonistic potential of bacterial endophytes and induction of systemic resistance against collar rot pathogen Sclerotium rolfsii in tomato. Biological Control 137:1040–1014
Saini R, Dudeja SS, Giri R, Kumar V (2015) Isolation, characterization, and evaluation of bacterial root and nodule endophytes from chickpea cultivated in Northern India. Journal of basic microbiology 55(1):74–81
Sobolev VS, Orner VA, Arias RS (2013) Distribution of bacterial endophytes in peanut seeds obtained from axenic and control plant material under field conditions. Plant and soil 371(1-2):367–376
Sundaramoorthy S, Raguchander T, Ragupathi N, Samiyappan R (2012) Combinatorial effect of endophytic and plant growth promoting rhizobacteria against wilt disease of Capsicum annum L. caused by Fusarium solani. Biological Control 60(1):59–67
Truyens S, Weyens N, Cuypers A, Vangronsveld J (2015) Bacterial seed endophytes: genera, vertical transmission and interaction with plants. Environmental Microbiology Reports 7(1):40–50
Upreti R, Thomas P (2015) Root-associated bacterial endophytes from Ralstonia solanacearum resistant and susceptible tomato cultivars and their pathogen antagonistic effects. Front Microbiology 6:255
Wang S, Wang W, Jin Z, Du B, Ding Y, Ni T, Jiao F (2013) Screening and diversity of plant growth promoting endophytic bacteria from peanut. Afr J Microbiol Res 7(10):875–884
Xu M, Sheng J, Chen L, Men Y, Gan L, Guo S, Shen L (2014) Bacterial community compositions of tomato (Lycopersicum esculentum Mill.) seeds and plant growth promoting activity of ACC deaminase producing Bacillus subtilis (HYT-12-1) on tomato seedlings. World Journal of Microbiology and Biotechnology 30(3):835–845
Acknowledgements
We would like to acknowledge the DST-FIST Lab & Biocontrol Lab, Department of Plant Pathology, TNAU, Coimbatore, for providing the necessary facilities for the work.
This article does not contain any studies with human or animal participants performed by any of the authors.
Funding
Not applicable
Author information
Authors and Affiliations
Contributions
LR, SKM and GK designed the experiment. TA, LR and SKVP conducted the experiment and wrote the article and performed the statistical analysis. LR and GK revised the article and MP did the growth promotion, rhizoscanner study and its interpretation. All authors approved the final article after reading.
Corresponding author
Ethics declarations
Consent for publication
Not applicable
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Archana, T., Rajendran, L., Manoranjitham, S.K. et al. Culture-dependent analysis of seed bacterial endophyte, Pseudomonas spp. EGN 1 against the stem rot disease (Sclerotium rolfsii Sacc.) in groundnut. Egypt J Biol Pest Control 30, 119 (2020). https://doi.org/10.1186/s41938-020-00317-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s41938-020-00317-x