
Abstract
Background
Goodeid topminnows are live-bearing fishes endemic to the Mexican Highlands (Mesa Central, MC). Unfortunately, in the MC, environmental degradation and introduced species have pushed several goodeid species to the brink of extinction. Invasive fishes can introduce exotic parasites, and the most abundant goodeid, blackfin goodea Goodea atripinnis Jordan, is parasitised by six exotic helminths. Poeciliids are widely dispersed invasive fishes, which exert negative ecological effects on goodeids. Poeciliids host several species of the monogenean genus Gyrodactylus von Nordmann, 1832, including pathogenic, invasive parasites. Here, we looked for evidence of Gyrodactylus species switching hosts from poeciliids to goodeids.
Methods
Fish were collected in rivers draining the MC into both sides of the continental divide. Hosts were screened for gyrodactylid parasites in localities where G. atripinnis and poeciliids occurred sympatrically. Gyrodactylus specimens were characterised morphologically (attachment apparatus) and molecularly (internal transcribed spacer region, ITS). A Bayesian phylogenetic tree using ITS sequences established relationships between gyrodactylids collected from goodeid fishes and those from parasites infecting poeciliids.
Results
Gyrodactylids were collected from G. atripinnis in six localities on both sides of the watershed where exotic poeciliids occurred sympatrically. Morphological and molecular analyses indicated the presence of four undescribed species of Gyrodactylus infecting this goodeid host. Gyrodactylus tomahuac n. sp., the most abundant and geographically widespread species, is described here. The other three Gyrodactylus spp. are not described, but their ITS sequences are used as molecular data presented here, are the only available for gyrodactylids infecting goodeid fishes. Morphological and molecular data suggest that two distinct groups of gyrodactylids infect goodeids, one of which shares a common ancestor with gyrodactylids parasitizing poeciliids.
Conclusions
No evidence was found of gyrodactylids switching hosts from invasive poeciliids to endemic goodeids, nor vice versa. Moreover, considering that G. atripinnis is known to host both Gyrodactylus lamothei Mendoza-Palmero, Sereno-Uribe & Salgado-Maldonado, 2009 and Gyrodactylus mexicanus Mendoza-Palmero, Sereno-Uribe & Salgado-Maldonado, 2009, with the addition of G. tomahuac n. sp. and the three undescribed Gyrodactylus spp. reported, at least six gyrodactylids may infect this host. This would make monogeneans the second most abundant parasite group infecting G. atripinnis, which to date is known to harbour 22 helminth species: nine digeneans, five nematodes, four cestodes, three monogeneans and one acanthocephalan.
Similar content being viewed by others

Background
Human-mediated introduction of non-native (also referred to as alien or exotic) fishes has resulted in the establishment of breeding populations of exotic fishes in freshwater water bodies worldwide [1–4]. Introduced fishes may compete directly with native fishes, and several invasive fish species have been demonstrated to exert negative ecological effects following invasion [1–3, 5]. Alien fishes can additionally act as carriers of non-indigenous parasites, which can negatively affect native hosts, as has been shown for several flatworm parasites of the class Monogenea Carus, 1863, whose short, one-host direct life-cycle facilitates establishment and invasion. Two compelling examples involve monogeneans from the genus Gyrodactylus von Nordmann, 1832 introduced along with their commercially-important fish hosts: Gyrodactylus salaris Malmberg, 1957, introduced in the mid-1970’s from the Baltic Sea to Norway, which has caused a highly pathogenic epizootic among wild and farmed salmonid fishes [6]; and Gyrodactylus cichlidarum Paperna, 1968, a parasite which has achieved almost global distribution through the translocation of its “tilapia” (Oreochromis spp.) hosts and has caused mass mortalities of cultured fish stocks [7].
The ornamental fish trade is an important component of international commerce, with over 1.5 billion fish belonging to more than 4000 freshwater and 1400 marine species traded annually and generating revenue in excess of US $ 6 billion [8]. As is the case for species translocated for aquacultural purposes, anthropogenic introduction of exotic ornamental fishes has resulted in alien monogenean parasites causing epizootic outbreaks in confined fish populations, for instance in display aquaria [9]. Furthermore, monogeneans co-introduced to continental freshwater bodies with their exotic fish hosts can contribute to the loss of native fish biodiversity, following parasite host switches (HS) from the introduced, alien fish to indigenous fish hosts (reviewed in [10]). One group of ornamental fishes exemplifies the invasiveness of feral aquarium fishes and their role as vectors to monogenean parasites: the small, live-bearing freshwater fishes belonging to the cyprinodontiform family Poeciliidae, which were originally distributed in the Americas, Africa and Madagascar [11]. Poeciliids have been translocated worldwide with the aquarium trade (e.g. “guppies” and “mollies” from the genus Poecilia Bloch & Schneider, “swordtails” and “platys” from the genus Xiphophorus Heckel) and mosquito control programmes (“mosquito fish” from the genera Gambusia Poey and Poecilia), and now introduced, feral populations of exotic poeciliids are established in all continents except Antarctica [3]. Arguably, poeciliids are the most successful exotic fish family in Australia [12], a country with strictly enforced quarantine rules governing the import of live animals, with five ornamental species having established breeding populations in the continent: Eastern mosquito fish, Gambusia holbrooki Girard, green swordtail, Xiphophorus hellerii Heckel, Southern platyfish, Xiphophorus maculatus (Günther), guppy, Poecilia reticulata Peters, and dusky millions fish, Phalloceros caudimaculatus (Hensel) [13]. The same five poeciliid species plus mosquito fish, Gambusia affinis (Baird & Girard), have established reproductively in Indian continental waters [10]. In both cases, exotic poeciliid fishes have co-introduced their gyrodactylid parasites [10, 13]; nonetheless, no evidence was found in Australia of gyrodactylids switching hosts from exotic poeciliid to native fishes. Trade of ornamental poeciliids has also contributed to the wide dissemination of their monogenean parasites: several gyrodactylid species have been recorded to infect aquarium fishes in Canada, UK, Czech Republic, Singapore, Korea, India, Australia, etc. [10, 12, 14]. In fact, five of the 19 species of Gyrodactylus known to infect poeciliid fishes were originally obtained from exotic fishes reared in aquaria (reviewed in [14]). Although their potential pathogenic effects on native fish populations have not been fully demonstrated, some gyrodactylids known to infect poeciliid fishes are remarkable for the extent of their geographical distribution following anthropogenic translocation, and for the range of host species they infect. For instance, Gyrodactylus bullatarudis Turnbull, 1956 has been recorded from six poeciliid fish species from the genera Gambusia, Poecilia, Pseudoxiphophorus Bleeker (syn. Heterandria), and Xiphophorus, infecting wild, captive and feral populations in North America, Europe and Australia (reviewed in [14]), and has also been shown to infect the non-poeciliid, cyprinodontiform killifish, Anablepsoides hartii (Boulenger) (syn. Rivulus hartii) [15].
Poeciliid fishes are native to the southern, Neotropical regions of Mexico, but have been extensively introduced to the central and northern, Nearctic highlands (Mesa Central, MC) of the country [4, 5, 16], where endemic goodeid fishes occur. Thus, mosquito fish, G. affinis, originally from north-eastern Mexico and the USA, has been introduced to the MC; as well as several poeciliid species originally restricted to the Gulf of Mexico watershed, such as shortfin molly Poecilia mexicana Steindachner, porthole livebearer Poeciliopsis gracilis (Heckel), two-spot livebearer Pseudoxiphophorus bimaculatus Heckel (syn. Heterandria bimaculata), green swordtail X. hellerii, and variable platyfish Xiphophorus variatus (Meek) [4, 17], and invasive Trinidadian guppies Poecilia reticulata Peters [2, 4, 18].
Goodeid topminnows or splitfins are a family of live-bearing cyprinodontoid fishes that evolved in the MC, and occur in Central Mexico and North America [17]; the subfamilies Goodeinae and Empetrichthyinae are principally distributed across these two geographic regions, respectively [19]. About 45 species from the MC are classified within the sub-family Goodeinae [20]. Goodeines were originally distributed in drainages along both the Pacific Ocean and Gulf of Mexico slopes; unfortunately, the MC is one of the most densely populated regions in Mexico and habitat destruction, water pollution, and introduction of exotic species have pushed several endemic fishes to the brink of extinction, with some goodeid species effectively extirpated from their natural habitats [21]. Only two goodeid species distributed in Central Mexico can be considered to be in no conservation risk category: the blackfin goodea Goodea atripinnis Jordan, and the dark-edged splitfin Girardinichthys multiradiatus (Meek) [22]. Goodea atripinnis remains common in many areas and is probably still the most abundant goodeid species overall, but its distribution and abundance have steadily declined over the last 30 years [20, 21, 23].
Goodea atripinnis has been recorded to be infected by 22 helminth parasite species [24], including 16 native and six alien parasite taxa: nine digeneans, five nematodes, four cestodes, three monogeneans and one acanthocephalan. Of these, the following are invasive, exotic parasites [18, 24]: the monogeneans Cichlidogyrus sclerosus Paperna & Thurston, 1969 and Dactylogyrus extensus Mueller & Van Cleave, 1932, the digenean Centrocestus formosanus Nishigori, 1924, the cestodes Schyzocotyle (syn. Bothriocephalus) acheilognathi (Yamaguti, 1934) Brabec, Waeschenbach, Scholz, Littlewood & Kuchta, 2015 and Proteocephalus ambloplitis (Leidy, 1887), and the nematode Pseudocapillaria tomentosa (Dujardin, 1843). Only three species of most likely indigenous monogeneans have been formally described from Mexican goodeids, all of which are known to infect G. atripinnis: the dactylogyrid Salsuginus angularis (Mueller, 1934) Beverley-Burton, 1984, and the gyrodactylids Gyrodactylus lamothei Mendoza-Palmero, Sereno-Uribe & Salgado-Maldonado, 2009 and Gyrodactylus mexicanus Mendoza-Palmero, Sereno-Uribe & Salgado-Maldonado, 2009 (Table 1), plus several records of undescribed species of Gyrodactylus from 11 goodeid fish species (Table 2). To date, 22 valid Gyrodactylus species are known to infect native and introduced fishes in Mexico (Table 1), including 11 species infecting poeciliid fishes [14, 25]. Some of the species of Gyrodactylus recorded in the country are capable of infecting several different genera of fish hosts (Table 1). For instance, both G. lamothei and G. mexicanus have been recorded on goodeids from the genera Allotoca, Girardinichthys and Goodea [24]; and Gyrodactylus pseudobullatarudis García-Vásquez, Razo-Mendivil & Rubio-Godoy, 2015 and Gyrodactylus xtachuna García-Vásquez, Razo-Mendivil & Rubio-Godoy, 2015 are able to infect poeciliids from the genera Poecilia, Poeciliopsis, Pseudoxiphophorus (syn. Heterandria) and Xiphophorus [14], which overlaps with the known host range of G. bullatarudis (see above).
Poeciliids and goodeids are morphologically similar viviparous, cyprinodontiform fishes that exhibit comparable habitat use [17], and are now sympatric following the human-mediated translocation of poeciliids to the MC. Exotic poeciliids have been shown to modify the ecological structure, function and native species abundance of water bodies following invasion [1], which is usually attributed to competition for habitat and/or food [3]. Additionally, invasive fishes may prey on native species, as shown for Xiphophorus spp., which feed on eggs and juveniles and have been implicated in the extinction of the goodeids golden skiffia, Skiffia francesae Kingston, and banded allotoca, Allotoca goslinei Smith & Miller [5]. A further negative impact of invasive poeciliids that has been recorded in the MC is the heterospecific harassment of native goodeids; in particular, male guppies (P. reticulata) have been recorded to attempt forced copulations of female twoline skiffia Skiffia bilineata (Bean) [2].
Given that the parasite fauna of the relatively abundant and widely distributed blackfin goodea, G. atripinnis is well known [24], with the exception of the Monogenea, and considering that Gyrodactylus sp. has been recorded to infect this goodeid in several states located in central Mexico (Table 2), a first objective of this study was to characterise gyrodactylids infecting this host in the MC. This included trying to collect specimens of G. lamothei and G. mexicanus, to characterise them molecularly, as their description contained morphometric data only [26]. A second objective was to ascertain whether any species of Gyrodactylus had switched host from introduced, alien poeciliids to endemic G. atripinnis. This was considered given that, as mentioned above, some species of Gyrodactylus have been shown to be extremely invasive following co-introduction with their fish hosts; and that poeciliids and goodeids are now sympatric in much of the MC and are likely to interact as they are morphologically and ecologically similar. We hypothesised HS were more likely from poeciliid to goodeid fishes than vice versa, because the first are very successful invaders and generally outnumber the endemic, endangered goodeids [4, 16], although evidently HS could occur in both directions. Potential HS from poeciliids to goodeids could in principle be recognised, considering that all 19 gyrodactylid species known to infect poeciliid fishes have been characterised morphologically [25], and parts of the genome have been sequenced for others [14]. To test the HS hypothesis, several localities on rivers draining the MC into both the Gulf of Mexico and the Pacific Ocean were sampled, and gyrodactylid parasites recovered from both G. atripinnis and poeciliids were analysed by morphological and molecular-based approaches from those localities where this goodeid fish and poeciliid fishes were found in sympatry. Finally, analysing the gyrodactylid fauna of non-native poeciliid fishes in the MC enabled a preliminary assessment of the enemy release hypothesis, which posits that upon translocation, introduced species lose some of their natural enemies such as parasites, thereby gaining a fitness advantage - a topic which was recently addressed studying the Gyrodactylus fauna of native and introduced minnow populations in Norway [27].
Methods
Specimen collection
Fishes were collected by electrofishing at different localities in the Río Pánuco and the Río Lerma basins, which flow from the MC into the Gulf of Mexico and the Pacific Ocean, respectively. Different species of goodeid [G. atripinnis, G. multiradiatus, S. bilineata and Xenotoca variata (Bean)] and poeciliid [P. bimaculatus, P. gracilis, P. mexicana, P. reticulata, Poeciliopsis infans (Woolman) and Pseudoxiphophorus jonesii (Günther)] fishes were collected. In the present study, only precise information is given for localities where blackfin goodea, G. atripinnis and poeciliid fish species were collected simultaneously (Table 3). Specimens were collected in April 2008 in the Río Moctezuma, Vega de Ramírez, Querétaro (21°03′31.01″N, 99°28′03.68″W); and in May 2014, in streams in Araro, Michoacán (19°54′27.52″N, 100°50′23.36″W); El Fresno, Guanajuato (20°16′39.07″N, 100°29′09.69″W); San Miguel Tlaxcaltepec (20°06′23.34″N, 100°07′36.74″W) and Santiago Mezquititlán (20°04′37.01″N, 100°04′29.38″W), both in Querétaro; in a reservoir at San Nicolás Peralta, Estado de México (19°21′26.75″N, 99°29′38.66″W); and along the northern shore of Lago de Chapala, Chapala, Jalisco (20°17′18.4″N, 103°11′35.9″W). Live fish were kept in buckets fitted with battery-operated aerators whilst electrofishing; an effort was made to keep poeciliid and goodeid species in separate buckets, to reduce the possibility of gyrodactylid parasite HS between non-related, cohabiting fishes [28]. After euthanasia, fish were preserved in groups separated by species and locality, and placed in labelled plastic bottles containing 96% ethanol until they were screened microscopically.
Specimen preparation
Fish fins, body and gills were inspected under a dissection microscope, and parasites found were dislodged gently using acupuncture needles. Representative gyrodactylid specimens were prepared as whole mounts in ammonium picrate glycerine following the procedure detailed by Malmberg [29] to study taxonomic features of the haptor (= opisthaptor, or terminal attachment organ), male copulatory organ (MCO) and pharynx. Further specimens had their haptors excised using a scalpel and were subjected to proteolytic digestion as described previously [30], to release the attachment hooks from enclosing tissue. The corresponding anterior portions of bisected bodies were stored at -20 °C in 96% ethanol, individually labelled for subsequent molecular analyses. The hooks were mounted in a semi-permanent 1:1 formalin : glycerin solution and the edges of the coverslip were then sealed with the permanent mounting medium Pertex (Histolab Products AB, Gothenburg, Sweden).
Morphological analysis
For the morphological study, images of the haptoral attachment hooks of proteolytically-digested specimens were captured using a Zeiss AxioCam MRc digital camera interfacing with an Olympus BH2 compound microscope using a ×0.75 lens and MRGrab 1.0.0.4 (Carl Zeiss Vision GmbH, 2001) software. Each gyrodactylid specimen was subjected to morphometric analysis taking 24 point-to-point measurements on the haptoral hooks using a JVC KY–F30B 3CCD video camera mounted on an Olympus BH2 microscope using a ×2.5 interfacing lens at ×100 oil immersion and the gyrodactylid-specific Point-R macro (Bron & Shinn, University of Stirling) written within the KS300 (ver.3.0) (Carl Zeiss Vision GmbH, 1997) image analysis software. The 24 morphometric measurements are given in micrometres or in degrees (for angles) as the range followed by the mean in parentheses, and were selected from those described in Malmberg [29], Shinn et al. [31] and Paladini et al. [32]. Holotypes and voucher specimens of G. lamothei (CNHE 6310, 6749-6753 and 7120) and G. mexicanus (CNHE 6745-6749 and 7128) were borrowed from the Colección Nacional de Helmintos (CNHE), Mexico City, and analysed morphometrically.
To describe and compare the aspect ratio of the hamuli of different Gyrodactylus spp., the quotient between the mean hamulus proximal shaft width (HPSW) and the hamulus total length (HTL) was calculated. Measurements used to calculate the aspect ratio of G. lamothei, G. mexicanus and those of gyrodactylids recovered from G. atripinnis are new. Morphometric data of Gyrodactylus spp. infecting poeciliid fishes were taken from [25] and [14].
Molecular analyses
Individual genomic DNA of 17 ethanol-fixed bodies of excised specimens of Gyrodactylus spp. collected from G. atripinnis, and of three worms collected from G. multiradiatus were extracted using a DNeasy® Blood & Tissue Kit (Qiagen, Valencia, California, USA) following the manufacturer’s instructions. The target region spanning the 3′ end of the 18S rRNA gene, ITS1, 5.8S rRNA gene, ITS2 and the 5′ end of the 28S rRNA gene subunit failed to amplify using the primers employed previously by Rubio-Godoy et al. [33]. New primers ITS1-fm 5′-TAG AGG AAG TAC AAG TCG-3′ and ITS2-rm 5′-CGC TYG AAT CGA GGT CAG GAC-3′ were designed to facilitate amplification of the target using the following PCR conditions: initial denaturation for 4 min at 95 °C followed by 35 cycles of: 94 °C for 30 s, 54 °C for 45 s, 72 °C for 1 min, with a final extension at 72 °C for 7 min. PCR amplicons were visualised on GelRed (Biotium, San Francisco, California, USA) stained 1% agarose gels and then purified with ExoSap-IT (USB Corporation, Cleveland, Ohio, USA). Sequencing reactions were carried out with the use of Big Dye Terminator chemistry, incorporating the same primers as those used in PCR, and cleaned by filtration with Sephadex G50. The sequenced products were read on an ABI PRISM 3100 automated DNA sequencer (Applied Biosystems, Foster City, California, USA). Electropherograms were visually inspected with the use of FinchTV (Geospiza Inc., Seattle, Washington, USA), and overlapping fragments of forward and reverse sequences were assembled with the use of the computer program BioEdit v. 7.0.9 [34]. Sequences generated in the present study were deposited in the GenBank database.
New sequences of the ITS1, 5.8S rRNA gene, and ITS2 of the Gyrodactylus spp. infecting goodeid fishes were compared with the following Gyrodactylus sequences available from GenBank: Gyrodactylus actzu García-Vásquez, Razo-Mendivil & Rubio-Godoy, 2015; Gyrodactylus apazapanensis García-Vásquez, Razo-Mendivil & Rubio-Godoy, 2015; Gyrodactylus arcuatus Bychowsky, 1933; G. bullatarudis; Gyrodactylus gondae Huyse, Malmberg & Volckaert, 2004; Gyrodactylus jarocho Rubio-Godoy, Paladini, García-Vásquez & Shinn, 2010; Gyrodactylus lhkauhili García-Vásquez, Razo-Mendivil & Rubio-Godoy, 2015; Gyrodactylus microdactylus García-Vásquez, Razo-Mendivil & Rubio-Godoy, 2015; Gyrodactylus pakan Razo-Mendivil, García-Vásquez & Rubio-Godoy, 2016; Gyrodactylus pictae Cable, van Oosterhout, Barson & Harris, 2005; Gyrodactylus poeciliae Harris & Cable, 2000; G. pseudobullatarudis; Gyrodactylus stephanus Mueller, 1937; Gyrodactylus takoke García-Vásquez, Razo-Mendivil & Rubio-Godoy, 2015; Gyrodactylus teken Razo-Mendivil, García-Vásquez & Rubio-Godoy, 2016; Gyrodactylus turnbulli Harris, 1986; Gyrodactylus xalapensis Rubio-Godoy, Paladini, García-Vásquez & Shinn, 2010; and G. xtachuna. All sequences were aligned using MUSCLE v. 3.5 [35], implemented in the software SEAVIEW v. 4.2 [36]. Phylogenetic hypotheses of ITS sequences were inferred with maximum likelihood (ML) and Bayesian inference (BI). Prior to ML and BI analyses, the optimal model of evolution and parameter settings for the ITS dataset was selected in jModeltest 2.1.10 [37, 38] using the Akaike information criterion (AIC) [39] and the TVM+I+G model was selected. The likelihood program GARLI 2.0 [40] was used for topology reconstruction under a GTR model, allowing the program to estimate the I and G parameters. Two independent likelihood analyses were conducted for each data set to ensure convergence. Analyses were terminated after 100,000 generations, with an additional 1000 bootstrap replicates used for evaluating nodal support; each replicate was terminated after 10,000 generations. Bayesian phylogenetic relationships were inferred using MrBayes v.3.2.1 [41], running 10,000,000 generations and sampling one tree every 100 generations. Parameter settings used were nst = 6 and rates = invgamma. Posterior probabilities for supported clades were determined by a 50% majority-rule consensus of the 180,002 trees retained after 10% ‘burn-in’. Trees resulting of ML and BI analyses were rooted with G. pakan and G. teken, parasites of Astyanax aeneus, which occur sympatrically with poeciliids in their native distribution range in Veracruz. Finally, estimates of evolutionary divergence between ITS1, 5.8S and ITS2 sequences (number of base substitutions per site between sequence pairs) among species of Gyrodactylus parasitising goodeid and poeciliid fishes were obtained with MEGA7 [42]. All positions containing gaps and missing data were eliminated; there was a total of 677 positions in the final dataset.
Results
Goodeid and poeciliid fishes were found to be sympatric in several locations in both the Atlantic Ocean and Pacific Ocean watersheds of Mexico. In general, invasive poeciliids were more numerous than endemic goodeids at localities sampled in the MC (Table 3). In the current study, only cases where gyrodactylids were recovered from blackfin goodea, G. atripinnis in localities where both goodeid and poeciliid fishes were collected simultaneously are presented (Table 3). As reported previously [14], poeciliids (P. mexicana) captured within their native distribution range in the Río Pánuco basin on the Gulf of Mexico slope were infected with G. pseudobullatarudis and G. xtachuna. None of the gyrodactylid species known to infect poeciliids were found on invasive fishes at the localities sampled within the MC (Table 3). Nonetheless, two invasive poeciliid species (P. mexicana and P. bimaculatus) collected at Araro, Michoacán, within the Río Lerma basin were infected with the cichlid fish pathogen G. cichlidarum (García-Vásquez A, Razo-Mendivil U, Rubio-Godoy M, unpublished). Further, undescribed species of Gyrodactylus were found infecting poeciliid fishes on localities draining into both sides of the continental divide (Table 2); these will be described following collection of further samples. Specimens of G. lamothei were recovered from the goodeid G. multiradiatus collected at San Nicolás Peralta, Estado de México, a locality within the MC in the Río Lerma basin. No specimens of G. mexicanus were recovered.
Morphological and molecular analyses indicated that four undescribed species of Gyrodactylus infect the blackfin goodea G. atripinnis (Table 3). In the current study, only one species is described, i.e. the most abundant and geographically widespread gyrodactylid species, which was collected from both sides of the continental divide (Fig. 1). The other three undescribed species of Gyrodactylus (referred to as Gyrodactylus sp. 1, sp. 2 and sp. 3) were found in limited numbers (i.e. 1, 2 and 4 specimens, respectively) and in only a few localities (for details see Table 3); these latter species are not described taxonomically in the present work. The findings of the study, however, broadly illustrate that the three undescribed species differ morphologically from other known gyrodactylids infecting goodeid and poeciliid fishes; and molecular data obtained from them are presented, because given the scarcity of genetic data on Mexican Gyrodactylus spp., these new sequences inform the phylogenetic relationships between gyrodactylid parasites infecting goodeid and poeciliid fishes. A summary of the specimens prepared from each locality and analysed by morphological and molecular means is presented in Table 3.

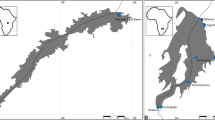
Map of Central Mexico, showing locations where Gyrodactylus tomahuac n. sp. was collected. The red star shows the type-locality of G. tomahuac n. sp. at Vega de Ramírez, Río Moctezuma, Querétaro (Qro). Río Moctezuma is a tributary of the Río Pánuco (shown in blue), the only major river from the Mesa Central (MC) that drains into the Gulf of Mexico. Locations where G. tomahuac n. sp. and other gyrodactylids were collected from Goodea atripinnis in the Río Lerma-Santiago basin, which drains the MC into the Pacific Ocean, are shown with letters: A, Araro, Michoacán; C, Lago de Chapala, Jalisco; F, El Fresno, Guanajuato; M, San Miguel Tlaxcaltepec, Qro; S, Santiago Mezquititlán, Qro
Gyrodactylus tomahuac n. sp.
Type-host: Goodea atripinnis Jordan (“blackfin goodea”, “Tiro”) (Cyprinodontiformes: Goodeidae).
Type-locality: Río Moctezuma (21°03′31.01″N, 99°28′03.68″W), at Vega de Ramírez, Cadereyta de Montes, Querétaro, Mexico.
Other localities: Streams in Araro, Michoacán (19°54′27.52″N, 100°50′23.36″W); El Fresno, Guanajuato (20°16′39.07″N, 100°29′09.69″W); and San Miguel Tlaxcaltepec, Querétaro (20°06′23.34″N, 100°07′36.74″W).
Type-material: Fifteen specimens were studied for light microscopy, nine from the type-locality and six from other localities (Table 2). Holotype (CNHE accession no. 9991), paratype (CNHE no. 9992) and voucher specimens (CNHE nos. 9930 through 9932, and 9993 through 9995) are deposited in the Colección Nacional de Helmintos, Instituto de Biología, Universidad Nacional Autónoma de México (UNAM), Mexico City, Mexico. In addition, three voucher specimens (CMNPA accession no. 2015-0009, CMNPA 2015-0010 and CMNPA 2015-0011) have been deposited in the Canadian Museum of Nature, Parasite Collection (CMNPA), Ontario, Canada.
Site on host: Fins.
Representative DNA sequences: Partial ITS1 and ITS2 sequences and complete 5.8S ribosomal RNA gene sequences, with a length of 904 bp, have been deposited in GenBank under accession numbers KJ621983 (type-locality specimen, n = 1) and KR815847–KR815852 (Río Lerma basin paratypes/voucher specimens, n = 6).
ZooBank registration: In accordance with the regulations set out in article 8.5 of the International Code of Zoological Nomenclature (ICZN) [43], details of the new species have been submitted to ZooBank. The Life Science Identifier (LSID) of the article is urn:lsid:zoobank.org:pub:375D2AD9-11F8-4705-B966-A1EA19C2CA5B. The LSID for the new name Gyrodactylus tomahuac is urn:lsid:zoobank.org:act:73980450-57E3-4A03-9F17-70BD0E392ECB. The electronic edition of this work is published in a journal with an ISSN, and has been archived and is available from the following digital repositories: PubMed Central, LOCKSS. In addition, a species profile including taxonomic traits, host details and additional metadata is provided on www.gyrodb.net [44].
Etymology: From the Náhuatl (Aztec) word “tomahuac” meaning thick, robust, which describes the nature of the hamuli.
Description
[Body measurements based on six whole worms, partially proteolytic-digested ammonium picrate glycerine, coverslip-flattened specimens (i.e. not processed for morphometric analysis of haptoral structures); see Table 4.] Body 223–345 (273) long, 70–76 (73.7) wide at uterus mid-point. Anterior pharyngeal bulb 16 × 24, posterior pharyngeal bulb 10 × 30. Excretory bladders present. Gut not extending beyond posterior terminus of uterus, almost reaching limit of haptor; testes and ovary not discerned. Haptor circular, clearly delineated from body, 68 × 63. Male copulatory organ posterior to pharynx, visible on one specimen only, 7.9 × 9.3, armed with a single curved apical spine facing a single row of 6 equally sized spines, 1.8 long (Fig. 2f, i). Hamuli, proportionately stout (Fig. 2a, g); total length 42.2–46.4 (44.9); broad at dorsal bar attachment point with proximal shaft width of 4.0–5.1 (4.4); shaft length 24.9–26.4 (25.6) long; similarly-sized point 22.3–24.5 (23.5) long such that each hamulus tip extends towards ventral bar articulation point on each hamulus, creating small hamulus aperture distance 10.9–15.0 (12.6) long and narrow aperture angle of 25.5–36.1° (30°). Root portion of each hamulus narrows away from shaft (i.e. from dorsal and ventral bar attachment points) with both dorsal and ventral surfaces of root, 17.8–21.5 (19.3) long, following a shallow, recurved line. Root terminus of each hamulus slopes towards its anterior and inner extremity. Dorsal bar attachment points approximately rectangular; dorsal bar simple, 18.7–21.6 (20.2) long, attached to upper third of dorsal bar attachment point; bar broadens slightly away from its attachment to each hamulus and then narrows marginally at its mid-point, 1.8–2.3 (2.0) wide (Fig. 2a). Anterior edge of median portion of ventral bar proper straight, posterior edge curves gently (Fig. 2c). Ventral bar processes small, rounded, 1.6–2.5 (2) long, positioned laterally in anterior third of ventral bar extremities. Ventral bar membrane lingulate at its anterior portion, posterior portion ends as a narrow “V”, 9.2–14.4 (10.7) long. Total length of marginal hooks 29.5–31.7 (31.0) (Fig. 2b); marginal hook shaft 24.0–26.7 (25.9) long; marginal hook sickle proper 5.2–5.8 (5.5) long. Shaft of sickle proper, angled approximately forward 15° to the perpendicular, uniform along its length, before turning almost 90° into a short tip that terminates at a point in line with approximate anterior third of sickle toe (Fig. 2d, e, h). Sickle distal width 2.0–2.7 (2.3), proximal width 3.7–4.4 (4.0). Aperture of marginal sickle, 4.5–5.3 (4.9) wide, inner curve of sickle proper approximately rectangular. Base of sickle proper with a flat bridge, approximately one third the proximal width of base; triangular toe 1.2–1.8 (1.6) long, terminates below marginal hook shaft insertion point on sickle proper and in line with posterior edge of heel. Sickle instep height 0.4–0.8 (0.6). Heel of sickle proper, pronounced, approximately rhomboid in dimensions and drops 35° in a downwards direction from the perpendicular (Fig. 2d, e, h).

Haptoral armature and male copulatory organ of Gyrodactylus tomahuac n. sp. ex Goodea atripinnis. a Hamuli and dorsal bar. b Marginal hook. c Ventral bar. d-e Marginal hook sickles. f Male copulatory organ (MCO). g Line drawing of the hamulus complex. h Line drawing of the marginal hook sickle. i Line drawing of the MCO. Scale-bars: a-c, g, 5 μm; d-f, h-i, 3 μm
Remarks
Gyrodactylus tomahuac n. sp. is the third gyrodactylid described from G. atripinnis. This host has previously been found to harbour G. lamothei and G. mexicanus [24], as well as undescribed species of Gyrodactylus (Table 2). At the type-locality, Vega de Ramírez, Río Moctezuma, Querétaro, the only gyrodactylid species recorded was G. tomahuac n. sp., with 41 parasites found on ten hosts (Table 3). At other localities, G. tomahuac n. sp. burdens were lower; and G. tomahuac n. sp. was found concurrently infecting the same hosts as Gyrodactylus sp. 3 in two localities in the Río Lerma basin (Table 3). Gyrodactylus tomahuac n. sp. was found in four different localities in the Mexican highlands (Fig. 1) at considerable distances from each other: Vega de Ramírez, Río Moctezuma is c.150 km apart from the other three locations in the Río Lerma basin where the parasite was recorded (i.e. El Fresno, San Miguel Tlaxcaltepec, and Araro), and these three sites are c.60 km apart from each other. More importantly, the parasite was recorded in river basins flowing into both directions of the continental divide: the type-locality (Vega de Ramírez) at the Río Moctezuma is within the Río Pánuco basin, which drains into the Gulf of Mexico; while streams at El Fresno, San Miguel Tlaxcaltepec and Araro are part of the Río Lerma-Santiago basin and flow into the Pacific Ocean.
The morphology of the marginal hooks is considered the most informative character to differentiate between species of Gyrodactylus. The marginal hooks of G. tomahuac n. sp. clearly differ in shape from those of both G. lamothei and G. mexicanus (Fig. 3): the marginal hooks of G. lamothei possess a very long and forward-tilted shaft whose tip ends well beyond the toe of a rounded, almost elliptical base, in contrast to G. tomahuac n. sp. where the marginal hook shaft is upright and does not extend beyond the angular base of the toe; and the marginal hooks of G. mexicanus have an upright shaft ending in a downward-curved point almost as long as the shaft itself, and the marginal hook base has an extended bridge ending in a rounded toe, in contrast to the comparatively short shaft point and short-bridged, angular toe in G. tomahuac n. sp. Superficially, the marginal hooks of G. tomahuac n. sp. resemble those of two gyrodactylids infecting poeciliid fishes, G. bullatarudis and G. takoke (Fig. 4); however, features of the marginal hook base allow their discrimination. While the marginal hook base in G. tomahuac n. sp. has a clear bridge almost level with the union of the heel and the shaft which forms a sharp angle with an almost straight, downward inclined toe, in both G. bullatarudis and G. takoke the toe curves gently downward and there is only a very short bridge in the first, and no bridge in the latter.

Haptoral armatures of different species of Gyrodactylus von Nordmann, 1832 infecting goodeid fishes. Gyrodactylus sp. 1, sp. 2, and sp. 3 refer to the three undescribed but genetically characterised species reported in this work. Scale-bar: 10 μm

Marginal hook sickles of different species of Gyrodactylus von Nordmann, 1832 infecting poeciliid fishes. The marginal hook of Gyrodactylus tomahuac n. sp., a parasite of the goodeid fish Goodea atripinnis is shown for comparison. a G. actzu. b G. apazapanensis. c G. bullatarudis. d G. costaricensis. e G. cytophagus. f G. gambusiae. g G. lhkahuili. h G. jarocho. i G. microdactylus. j G. milleri. k G. pictae. l G. poeciliae. m G. pseudobullatarudis. o G. takoke. p G. tomahuac n. sp. q G. turnbulli. r G. unami. s G. xalapensis. t G. xtachuna. Scale-bar: 5 μm
Gyrodactylus tomahuac n. sp. can also be differentiated from gyrodactylids known to infect goodeid and poeciliid fishes, comparing the overall morphology of their haptoral sclerites. Based on the variation in the haptoral hook morphology of the Gyrodactylus species infecting goodeid fishes (Fig. 3), two groups can be roughly identified: G. lamothei and Gyrodactylus sp. 2 both have relatively slender hamuli and marginal hooks with forward-angled shafts extending well beyond the toe, while G. tomahuac n. sp., G. mexicanus and both Gyrodactylus sp. 1 and sp. 3 possess robust hooks, with comparatively wider hamulus proximal shafts (as shown by the ratio between the mean hamulus proximal shaft width (HPSW) to the hamulus total length (HTL; Table 5), and marginal hooks with upright, shorter sickle shafts. Further, G. tomahuac n. sp. can easily be distinguished from G. lamothei, G. mexicanus and Gyrodactylus sp. 2, as these have elongated sclerotised structures attached to the root of the hamuli (Fig. 3). The haptoral hook morphology of G. tomahuac n. sp. is quite distinctive from that of most gyrodactylid species infecting poeciliid fishes (Fig. 5), except for perhaps G. bullatarudis, G. gambusiae Rogers & Wellborn, 1965 and G. pictae, all of which have robust hamuli (Table 5); albeit these species can be easily separated from G. tomahuac n. sp. because they all have longer hamuli (HTL > 50 μm vs c.45 μm) and since the ventral bars of these gyrodactylids infecting poeciliid fishes are comparatively larger and have long processes, which are absent on the relatively smaller ventral bar of G. tomahuac n. sp. Some other gyrodactylids infecting poeciliids have stout hamuli (HPSW/HTL ratio = 0.16; Table 5), but the majority possess slender hamuli.

Hamulus complex, including dorsal and ventral bars of different species of Gyrodactylus von Nordmann, 1832 infecting poeciliid fishes. The hamulus complex of Gyrodactylus tomahuac n. sp., a parasite of the goodeid fish Goodea atripinnis is shown for comparison. Scale-bar: 10 μm
Phylogenetic analyses
The length of the new ITS sequences of 14 specimens of Gyrodactylus spp. (seven G. tomahuac n. sp., one Gyrodactylus sp. 1, two Gyrodactylus sp. 2, and four Gyrodactylus sp. 3) collected from G. atripinnis varied from 866 to 934 bp. ITS sequences obtained for G. lamothei were deposited in GenBank (GenBank acc. nos. KX555666–KX555668), and varied from 887 to 888 bp. One representative sequence of each of these mentioned taxa was compared with those downloaded from GenBank, producing an alignment composed by 24 sequences with a length of 1324 bp. Phylogenetic analyses did not include G. mexicanus, because no ITS sequences are available in GenBank for this species and we did not recover any specimens during the current survey.
ML and BI analyses of ITS sequences recovered identical phylogenetic relationships among the analysed species, with several well-supported nodes. ML analysis resulted in one tree (not shown) with a value of likelihood = -10,210.6007. The 50% majority-rule consensus tree (Fig. 6) obtained from the BI analysis recovered three main clades, in which the sequences of Gyrodactylus spp. infecting goodeid and poeciliid fishes do not constitute monophyletic assemblages; and the gyrodactylids from goodeid fishes appeared in two clades constituting five different lineages.

Phylogenetic tree of different Gyrodactylus species infecting teleost fishes. Shaded boxes indicate parasite clades infecting the following host groups: Goodea atripinnis (red), other goodeid fishes (yellow), and poeciliid fishes (green and blue). Fifty percent majority-rule consensus tree obtained from the Bayesian Inference analysis of the ITS1, 5.8S and ITS2 sequences from 23 species of Gyrodactylus. Bayesian posterior probabilities are shown above the nodes, and ML bootstrap support below the nodes. The phylogram is rooted with gyrodactylids infecting the characid Astyanax aeneus
Sequences of G. tomahuac n. sp., Gyrodactylus sp. 1 and Gyrodactylus sp. 3 formed a main clade with very low branch support (0.5321 of pp). This clade is composed by one terminal branch (Gyrodactylus sp. 1) and one derived group containing G. tomahuac n. sp. and Gyrodactylus sp. 3 (Fig. 6; red box). A second main clade contains two subclades, one of them constituted by the goodeid-infecting parasites Gyrodactylus sp. 2 and G. lamothei (red and yellow boxes), which appear as sister group to a cluster of mostly well-supported derived branches containing 11 species of Gyrodactylus infecting poeciliid fishes (green box). Finally, a third main clade has one terminal and two derived branches containing parasites infecting several non-related fish hosts: G. gondae infecting Lozano’s goby Pomatoschistus lozanoi (de Buen); G. stephanus infecting common mummichogs Fundulus heteroclitus (L.); G. arcuatus infecting three-spined sticklebacks Gasterosteus aculeatus L.; and two gyrodactylids from poeciliids, G. xalapensis and G. takoke (blue box).
Intra- and inter-specific nucleotide variation
Comparison of ITS1, 5.8S and ITS2 sequences among the 24 species analysed showed a great amount of nucleotide variation, ranging from 0.4 to 32.7%. The species pairs that showed these contrasting values were, respectively, G. lamothei and Gyrodactylus sp. 2, and both G. arcuatus and Gyrodactylus sp. 2, as well as G. jarocho and G. stephanus. Likewise, new sequences of the gyrodactylids collected from G. atripinnis, representing G. tomahuac n. sp. and three unnamed species (Gyrodactylus sp. 1, sp. 2, and sp. 3) exhibited high sequence variation (Table 6). Nucleotide differences ranging from 6.8 to 31.3% were found among the 14 sequences of Gyrodactylus spp. infecting this goodeid fish. Values of nucleotide variation between the new species, G. tomahuac, and the other three unnamed species (Gyrodactylus sp. 1, sp. 2, and sp. 3) were 25.6 to 25.9%, 24.0 to 24.2%, and 6.8 to 7.1%, respectively.
Intra-specific nucleotide variation was detected in G. tomahuac n. sp. and in Gyrodactylus sp. 3. Of the seven sequenced specimens of G. tomahuac n. sp. and four Gyrodactylus sp. 3, four and one specimens showed nucleotide variation, which ranged from 0.13 to 0.27%, and from 0.12 to 0.13%, respectively. Intra-specific nucleotide variation ranging from 0.13 to 0.45% was detected in the three G. lamothei specimens sequenced.
Discussion
The first objective of this study was to characterise the gyrodactylid fauna of G. atripinnis within its native distribution range in the MC; and in doing so, assess whether HS have occurred to this endemic host from alien poeciliid fishes, which have been extensively introduced to central Mexico.
No evidence was found of gyrodactylid HS from poeciliids to goodeids, nor vice versa, in the six localities where fishes of these families occurred sympatrically. In all instances, poeciliid fishes were more abundant than goodeid fishes (Table 3), and the former included species known to harbour several gyrodactylids: P. mexicana has the richest gyrodactylid fauna associated with poeciliids, as this host has been recorded to harbour six parasite species in Mexico; P. bimaculatus is a translocated species known to be infected by four gyrodactylid species; and the invasive P. reticulata has been registered as host to three gyrodactylids [14]. In their native Río Pánuco basin, P. mexicana were infected by G. pseudobullatarudis and G. xtachuna, two gyrodactylid species known to have a wide geographical and host range [14]. In the MC, poeciliid fishes were not infected by any of the gyrodactylids known to parasitise them in their native distribution range [14, 25], some of which have been widely disseminated through the ornamental aquarium trade [10, 13]. Although the current study was not designed to test the enemy release hypothesis, results suggest that upon translocation to the MC, poeciliid fishes have lost their gyrodactylid parasite fauna; however, this finding is based on the limited sample sizes of poeciliid fishes found sympatrically with G. atripinnis in a few localities only, and are by no means representative of invasive poeciliid populations established in the MC. Surprisingly, invasive P. mexicana and P. bimaculatus collected in the MC were found to carry the alien tilapia parasite G. cichlidarum (García-Vásquez A, Razo-Mendivil U, Rubio-Godoy M, unpublished).
Goodea atripinnis was found to be infected by G. tomahuac n. sp. and three undescribed Gyrodactylus spp. This host was previously recorded to be infected by G. lamothei and G. mexicanum [24], suggesting that this endemic fish is parasitised by at least six gyrodactylids. The helminth fauna of goodeid fishes is well-characterised [24], and G. atripinnis is the host species known to harbour most parasites: nine digeneans, five nematodes, four cestodes, three monogeneans (including G. lamothei and G. mexicanum) and one acanthocephalan are recorded to infect this host. Considering six gyrodactylid species as parasites of this host, monogeneans represent the second most numerous helminth group known to infect this endemic fish. Bearing in mind that several reports have been made of the presence of undescribed Gyrodactylus sp. infecting G. atripinnis and several other goodeid fishes (Table 2); and the fact that along with G. multiradiatus, G. atripinnis remains a common and relatively abundant fish in much of the MC [22], it is foreseeable that several new species of Gyrodactylus infecting goodeid fishes will be found, potentially making monogeneans the most species-rich group of helminths infecting this host group.
Regarding the characterisation of the gyrodactylid fauna of G. atripinnis and other goodeid fishes, both morphological and molecular data support the hypothesis that two distinct groups of parasites infect these native fishes. Morphologically, some gyrodactylids possess robust hamuli, while others have slender hamuli with sclerotised plates located at the base of these sclerites. Very robust hamuli are the most salient feature of G. tomahuac n. sp., the species described here, a characteristic shared with G. mexicanus, and to a slightly lesser degree with the undescribed Gyrodactylus sp. 1 and Gyrodactylus sp. 3, both of which have stout hamuli (Table 5). Both G. lamothei and the undescribed Gyrodactylus sp. 2 possess slender hamuli, with elongated plates located at their roots; these plates were also drawn in the description of G. mexicanus [26]. It would be interesting to ascertain whether these structures are similar to the sclerotised plates found in Gyrodactylus proterorhini Ergens, 1967 [45], and to establish the phylogenetic relationships of G. mexicanus to other gyrodactylids once molecular markers are available.
The phylogenetic tree presented here is the most complete hypothesis of the relationships between gyrodactylids infecting goodeid fishes; to date, the only molecular data available for these parasites are those presented in this work (Fig. 6). Based on this hypothesis, Gyrodactylus spp. infecting goodeid fishes do not constitute a monophyletic group, as these parasites appear in two clades, which correspond to the two groups formed based on the morphology of the hamuli. The first, not well-supported clade includes G. tomahuac n. sp. as sister species to the undescribed Gyrodactylus sp. 3, and Gyrodactylus sp. 1 as a terminal branch; these three taxa all have relatively robust hamuli, albeit their marginal hooks differ (Fig. 3). Gyrodactylus lamothei and Gyrodactylus sp. 2 both have slender hamuli with plates at their roots, and appear in a separate clade as sister taxa to 11 gyrodactylid species known to infect poeciliid fishes, most of which also possess slender hamuli. Two gyrodactylids known to infect poeciliid fishes, G. takoke and G. xalapensis, appear in a further clade, which includes parasites infecting a variety of non-related fish hosts. Molecular characterisation of further gyrodactylids infecting goodeid and poeciliid fishes should shed light on the phylogenetic hypotheses outlined here: that two morphologically-distinct parasite lineages infect goodeids, one of which shares a common ancestor with several gyrodactylids infecting poeciliids.
Conclusions
No evidence was found of gyrodactylids switching host from introduced poeciliid fishes to endemic goodeid fishes in the MC; nor vice versa. Goodea atripinnis harbours at least six species of Gyrodactylus, including G. tomahuac n. sp., which is described here. Morphological and molecular data support the hypothesis that two groups of gyrodactylids infect goodeid fishes: one contains worms which possess robust hamuli including G. tomahuac n. sp., and the other encompasses parasites with slender hamuli and sclerotised plates at their roots such as G. lamothei - and phylogenetic analyses suggest that this second group shares a common ancestor with gyrodactylids infecting poeciliid fishes. Monogeneans may be the most species-rich group of helminth parasites infecting goodeid fishes.
Abbreviations
- AIC:
-
Akaike information criterion
- BI:
-
Bayesian inference
- CMNPA:
-
Canadian Museum of Nature Parasite Collection, Ontario
- CNHE:
-
Colección Nacional de Helmintos Mexico City
- HPSW:
-
Hamulus proximal shaft width
- HS:
-
Host switch(es)
- HTL:
-
Hamulus total length
- ITS:
-
Internal transcribed spacer
- MC:
-
Mexican central highlands Mesa Central
- MCO:
-
Male copulatory organ
- ML:
-
Maximum likelihood
- Qro:
-
State of Querétaro
References
Holitzki TM, MacKenzie RA, Wiegner TN, McDermid KJ. Differences in ecological structure, function, and native species abundance between native and invaded Hawaiian streams. Ecol Appl. 2013;23:1367–83.
Valero A, Macías-García C, Magurran AE. Heterospecific harassment of native endangered fishes by invasive guppies in Mexico. Biol Lett. 2008;4(2):149–52.
Pyke GH. Plague minnow or mosquito fish? A review of the biology and impacts of introduced Gambusia species. Annu Rev Ecol Evol Syst. 2008;39:171–91.
Contreras-MacBeath T, Gaspar-Dillanes MT, Huidobro-Campos L, Mejía-Mojica H. Peces invasores en el centro de México. In: Mendoza R, Koleff P, editors. Especies acuáticas invasoras en México. Mexico City: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad; 2014. p. 413–24.
Mendoza R, Luna S, Aguilera C. Risk assessment of the ornamental fish trade in Mexico: analysis of freshwater species and effectiveness of the FISK (Fish Invasiveness Screening Kit). Biol Invasions. 2015;17(12):3491–502.
Bakke TA, Cable J, Harris PD. The biology of gyrodactylid monogeneans: the “Russian-doll killers”. Adv Parasitol. 2007;64:161–376.
García-Vásquez A, Hansen H, Christison KW, Rubio-Godoy M, Bron JE, Shinn AP. Gyrodactylids (Gyrodactylidae, Monogenea) infecting Oreochromis niloticus niloticus (L.) and O. mossambicus (Peters) (Cichlidae): a pan-global survey. Acta Parasitol. 2010;55(3):215–29.
FAO yearbook. Fishery and aquaculture statistics. Rome: Food and Agriculture Organization of the United Nations; 2012. p. 1–76.
Thoney DA, Hargis WJ. Monogenea (Platyhelminthes) as hazards for fish in confinement. Annu Rev Fish Dis. 1991;1:133–53.
Tripathi A. The invasive potential of parasitic monogenoids (Platyhelminthes) via the aquarium fish trade: an appraisal with special reference to India. Rev Aquacult. 2014;6(3):147–61.
Froese R, Pauly D. FishBase. World Wide Web electronic publication [www.fishbase.org]. 2016. Accessed 3 Nov 2016.
Whittington RJ, Chong R. Global trade in ornamental fish from an Australian perspective: the case for revised import risk analysis and management strategies. Prev Vet Med. 2007;81(1-3):92–116.
Dove ADM. Richness patterns in the parasite communities of exotic poeciliid fishes. Parasitology. 2000;120:609–23.
García-Vásquez A, Razo-Mendivil U, Rubio-Godoy M. Morphological and molecular description of eight new species of Gyrodactylus von Nordmann, 1832 (Platyhelminthes: Monogenea) from poeciliid fishes, collected in their natural distribution range in the Gulf of Mexico slope, Mexico. Parasitol Res. 2015;114(9):3337–55.
Cable J, Archard GA, Mohammed RS, McMullan M, Stephenson JF, Hansen H, van Oosterhout C. Can parasites use predators to spread between primary hosts? Parasitology. 2013;140(9):1138–43.
Ruiz-Campos G, Varela-Romero A, Sánchez-Gonzales S, Camarena-Rosales F, Maeda-Martínez AM, González-Acosta AF, et al. Peces invasores en el noroeste de México. In: Mendoza R, Koleff P, editors. Especies acuáticas invasoras en México. México City: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad; 2014. p. 375–99.
Miller RR, Minckley WL, Norris SM. Freshwater fishes of México. Chicago: The University of Chicago Press; 2005. p. 1–652.
Salgado-Maldonado G, Rubio-Godoy M. Helmintos parásitos de peces de agua dulce introducidos. In: Mendoza R, Koleff P, editors. Especies acuáticas invasoras en México. Mexico City: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad; 2014. p. 269–85.
Integrated Taxonomic Information System. World Wide Web electronic publication [www.itis.gov]. 2016. Accessed 3 Nov 2016.
Domínguez-Domínguez O, Pedraza-Lara C, Gurrola-Sánchez N, Pérez-Rodríguez R, Israde-Alcántara I, Garduño-Monroy VH, et al. Historical biogeography of the Goodeinae (Cyprinodontiforms). In: Uribe-Aranzabal MC, Grier H, editors. Viviparous fishes vol II. Homestead: New Life Publications; 2010. p. 13–30.
De la Vega-Salazar MY, Macías-García C. Principal factors in the decline of the Mexican endemic viviparous fishes (Goodeidae: Goodeidae). In: Uribe-Aranzabal MC, Grier H, editors. Viviparous fishes vol II. Homestead: New Life Publications; 2005. p. 506–13.
De la Vega-Salazar MY. Conservation status of Goodeidae familiy fishes (Cyprinodontiformes) from the Mexican Central Plateau. Rev Biol Trop. 2006;54(1):163–77.
Mercado-Silva N, Lyons J, Díaz-Pardo E, Gutiérrez-Hernández A, Ornelas-García CP, Pedraza-Lara C, Vander Zanden MJ. Long-term changes in the fish assemblage of the Laja River, Guanajuato, central Mexico. Aquat Conserv. 2006;16(5):533–46.
Martínez-Aquino A, Mendoza-Palmero CA, Aguilar-Aguilar R, Pérez-Ponce de León G. Checklist of helminth parasites of Goodeinae (Osteichthyes: Cyprinodontiformes: Goodeidae), an endemic subfamily of freshwater fishes from Mexico. Zootaxa. 2014;3856:151–91.
Rubio-Godoy M, Paladini G, García-Vásquez A, Shinn AP. Gyrodactylus jarocho sp. nov. and Gyrodactylus xalapensis sp. nov. (Platyhelminthes: Monogenea) from Mexican poeciliids (Teleostei: Cyprinodontiformes), with comments on the known gyrodactylid fauna infecting poeciliid fish. Zootaxa. 2010;2509:1–29.
Mendoza-Palmero CA, Sereno-Uribe AL, Salgado-Maldonado G. Two new species of Gyrodactylus von Nordmann, 1832 (Monogenea : Gyrodactylidae) parasitizing Girardinichthys multiradiatus (Cyprinodontiformes: Goodeidae), an endemic fish from central Mexico. J Parasitol. 2009;95(2):315–8.
Pettersen RA, Ostbye K, Holmen J, Vollestad LA, Mo TA. Gyrodactylus spp. diversity in native and introduced minnow (Phoxinus phoxinus) populations: no support for “the enemy release” hypothesis. Parasit Vectors. 2016;9:51.
Grano-Maldonado MI, Bron JE, Longshaw M, Shinn AP. The accidental transfer of Gyrodactylus (Monogenea) during short duration fish transportation. Fish Pathol. 2011;46(3):71–9.
Malmberg G. The excretory systems and the marginal hooks as a basis for the systematics of Gyrodactylus (Trematoda, Monogenea). Ark Zool. 1970;23:1–235.
Paladini G, Gustinelli A, Fioravanti ML, Hansen H, Shinn AP. The first report of Gyrodactylus salaris Malmberg, 1957 (Platyhelminthes, Monogenea) on Italian cultured stocks of rainbow trout (Oncorhynchus mykiss Walbaum). Vet Parasitol. 2009;165(3-4):290–7.
Shinn AP, Hansen H, Olstad K, Bachmann L, Bakke TA. The use of morphometric characters to discriminate specimens of laboratory-reared and wild populations of Gyrodactylus salaris and G. thymalli (Monogenea). Folia Parasitol. 2004;51(2-3):239–52.
Paladini G, Huyse T, Shinn AP. Gyrodactylus salinae n. sp (Platyhelminthes: Monogenea) infecting the south European toothcarp Aphanius fasciatus (Valenciennes) (Teleostei, Cyprinodontidae) from a hypersaline environment in Italy. Parasit Vectors. 2011;4:100.
Rubio-Godoy M, Paladini G, Freeman MA, García-Vásquez A, Shinn AP. Morphological and molecular characterisation of Gyrodactylus salmonis (Platyhelminthes, Monogenea) isolates collected in Mexico from rainbow trout (Oncorhynchus mykiss Walbaum). Vet Parasitol. 2012;186(3-4):289–300.
Hall T. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp Ser. 1999;41:95–8.
Edgar RC. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004;32(5):1792–7.
Galtier N, Gouy M, Gautier C. SEAVIEW and PHYLO_WIN: two graphic tools for sequence alignment and molecular phylogeny. Comput Appl Biosci. 1996;12(6):543–8.
Guindon S, Gascuel O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst Biol. 2003;52(5):696–704.
Darriba D, Taboada GL, Doallo R, Posada D. jModelTest 2: more models, new heuristics and parallel computing. Nat Methods. 2012;9(8):772.
Schwarz G. Estimating the dimension of a model. Ann Stat. 1978;6:461–4.
Zwickl DJ. Genetic algorithm approaches for the phylogenetic analysis of large biological sequence datasets under the maximum likelihood criterion. Ph.D. thesis, The University of Texas at Austin; 2006. p. 1–115
Ronquist F, Teslenko M, van der Mark P, Ayres DL, Darling A, Hohna S, et al. MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Syst Biol. 2012;61(3):539–42.
Kumar S, Stecher G, Tamura K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol Biol Evol. 2016;33(7):1870–4.
International Commission on Zoological Nomenclature. Amendment of articles 8, 9, 10, 21 and 78 of the International Code of Zoological Nomenclature to expand and refine methods of publication. Zootaxa. 2012;3450:1–7.
Shinn AP, Harris PD, Cable J, Bakke TA, Paladini G, Bron JE. GyroDb. A home for gyrodactylids on the web. World Wide Web electronic publication [www.gyrodb.net]. 2016. Accessed 3 Nov 2016.
Huyse T, Vanhove MPM, Mombaerts M, Volckaert FAM, Verreycken H. Parasite introduction with an invasive goby in Belgium: double trouble? Parasitol Res. 2015;114:2789–93.
Salgado-Maldonado G, Novelo-Turcotte MT, Vázquez G, Caspeta-Mandujano JM, Quiroz-Martínez B, Favila M. The communities of helminth parasites of Heterandria bimaculata (Teleostei: Poeciliidae) from the upper Rio La Antigua basin, east-central Mexico show a predictable structure. Parasitology. 2014;141(7):970–80.
Rubio-Godoy M, Muñoz-Córdova G, Garduño-Lugo M, Salazar-Ulloa M, Mercado-Vidal G. Microhabitat use, not temperature, regulates intensity of Gyrodactylus cichlidarum long-term infection on farmed tilapia-Are parasites evading competition or immunity? Vet Parasitol. 2012;183(3-4):305–16.
García-Vásquez A, Hansen H, Shinn AP. A revised description of Gyrodactylus cichlidarum Paperna, 1968 (Gyrodactylidae) from the Nile tilapia, Oreochromis niloticus niloticus (Cichlidae), and its synonymy with G. niloticus Cone, Arthur et Bondad-Reantaso, 1995. Folia Parasitol. 2007;54(2):129–40.
Paredes-Trujillo A, Velázquez-Abunader I, Torres-Irineo E, Romero D, Vidal-Martínez VM. Geographical distribution of protozoan and metazoan parasites of farmed Nile tilapia Oreochromis niloticus (L.) (Perciformes: Cichlidae) in Yucatán, México. Parasit Vectors. 2016;9:66.
Salgado-Maldonado G, Cabañas-Carranza G, Soto-Galera E, Caspeta-Mandujano JM, Moreno-Navarrete RG, Sánchez-Nava P, Aguilar-Aguilar R. A checklist of helminth parasites of freshwater fishes from the Lerma-Santiago river basin, Mexico. Comp Parasitol. 2001;68(2):204–18.
Sánchez-Nava P, Salgado-Maldonado G, Soto-Galera E, Cruz BJ. Helminth parasites of Girardinichthys multiradiatus (Pisces: Goodeidae) in the upper Lerma River sub-basin, Mexico. Parasitol Res. 2004;93(5):396–402.
Salgado-Maldonado G. Checklist of helminth parasites of freshwater fishes from Mexico. Zootaxa. 2006;1324:1–357.
Mendoza-Franco EF, Scholz T, Vivas-Rodríguez C, Vargas-Vázquez J. Monogeneans of freshwater fishes from cenotes (sinkholes) of the Yucatan Peninsula, Mexico. Folia Parasitol. 1999;46(4):267–73.
Salgado-Maldonado G, Aguilar-Aguilar R, Cabañas-Carranza G, Soto-Galera E, Mendoza-Palmero C. Helminth parasites in freshwater fish from the Papaloapan river basin, Mexico. Parasitol Res. 2005;96(2):69–89.
López-Jiménez S. Estudio parasitológico de los peces de aguas dulces del estado de Tabasco. Gaceta SIGOLFO Sistema de Investigación del Golfo de México; 2001. p. 8–10
Razo-Mendivil U, García-Vásquez A, Rubio-Godoy M. Spot the difference: Two cryptic species of Gyrodactylus von Nordmann, 1832 (Platyhelminthes: Monogenea) infecting Astyanax aeneus (Actinopterygii, Characidae) in Mexico. Parasitol Int. 2016;65:389–400.
Pérez-Ponce de León G, Rosas-Valdez R, Aguilar-Aguilar R, Mendoza-Garfias B, Mendoza-Palmero C, García-Prieto L, et al. Helminth parasites of freshwater fishes, Nazas River basin, northern Mexico. Check List. 2010;6:26–35.
García-Vásquez A, Hansen H, Christison KW, Bron JE, Shinn AP. Description of three new species of Gyrodactylus von Nordmann, 1832 (Monogenea) parasitising Oreochromis niloticus niloticus (L.) and O. mossambicus (Peters) (Cichlidae). Acta Parasitol. 2011;56(1):20–33.
Salgado-Maldonado G, Cabañas-Carranza G, Soto-Galera E, Pineda-López RF, Caspeta-Mandujano JM, Aguilar-Castellanos E, Mercado-Silva N. Helminth parasites of freshwater fishes of the Panuco River Basin, east Central Mexico. Comp Parasitol. 2004;71(2):190–202.
Vidal-Martínez VM, Aguirre-Macedo ML, Scholz T, González-Solís D, Mendoza-Franco EF. Atlas of the helminth parasites of cichlid fish of Mexico. Praha: Academia; 2001. p. 1–165.
Díaz-Pardo E, Pineda-López R, Pineda-López R, Hernández Camacho N. Las comunidades acuáticas de la zona metropolitana de la ciudad de Querétaro. Nuevos Tiempos. 2009;14:12–22.
Hernández-Martínez M. Estudio helmintológico de tres especies de peces cultivados en dos centros acuícolas del estado de Sonora, México. Univers Ciencia. 1992;9:111–5.
Flores-Crespo J, Flores-Crespo R. Monogéneos, parásitos de peces en México: estudio recapitulativo. Técn Pec Méx. 2003;41:175–92.
Aguilar-Aguilar R, Rosas-Valdez R, Martínez-Aquino A, Pérez-Rodríguez R, Domínguez-Domínguez O, Pérez-Ponce de León G. Helminth fauna of two cyprinid fish (Campostoma ornatum and Codoma ornata) from the upper Piaxtla River, Northwestern Mexico. Helminthologia. 2010;47(4):251–6.
Flores-Crespo J, Flores CR. Principales tremátodos y céstodos de importancia económica en acuicultura. In: Tópicos de parasitología animal; céstodos y tremátodos. Cuernavaca: Universidad Autónoma del Estado de Morelos; 1993. p. 13–36.
Caspeta-Mandujano JM, Cabañas-Carranza G, Mendoza-Franco EF. Helmintos parásitos de peces dulceacuícolas mexicanos. Caso Morelos. Cuernavaca/México DF: UAEM/AGT editorial; 2009. p. 1–129
Aguilar-Aguilar R, Martínez-Aquino A, Espinosa-Pérez H, Pérez-Ponce de León G. Helminth parasites of freshwater fishes from Cuatro Cienegas, Coahuila, in the Chihuahuan Desert of Mexico: inventory and biogeographical implications. Integr Zool. 2014;9(3):328–39.
Armijo OA. Algunas enfermedades que se presentan en los centros acuícolas de México. In: II Simposio Latinoamericano de Acuicultura 1980; México, D.F.; 1980. p. 1607–18
Flores-Sotelo MT. Comparación de los helmintos parásitos de Hybopsis boucardi (Günther, 1968) (Pisces: Cyprinidae) en dos localidades del municipio de Coatlán del Río en el estado de Morelos, México. B.Sc. thesis, Univers Autón Estado Morelos; 1998. p. 1–45
Price CE, Henderson A. Monogenean parasites of Mexican freshwater fishes I. Introductory remarks, with an account of the parasite genus Dactylogyrus Diesing, 1850. An Inst Biol UNAM. 1969;40:195–204.
Monks S, Pulido-Flores G, Bautista-Hernández CE, Alemán-García B, Falcón Ordaz J, Gaytán-Oyarzún JC. El uso de helmintos parásitos como bioindicadores en la evaluación de la calidad del agua: Lago de Tecocomulco vs. Laguna de Metztitlán, Hidalgo, México. In: Pulido-Flores G, Monks S, editors. Estudios científicos en el estado de Hidalgo y zonas aledañas Volumen II. Lincoln: Zea Books; 2013. p. 25–34.
Scholz T, Vargas-Vázquez J, Moravec F, Vivas-Rodríguez C, Mendoza-Franco E. Cenotes (sinkholes) of the Yucatan Peninsula, Mexico as a habitat of adult trematodes of fish. Folia Parasitol. 1995;42(1):37–47.
Pineda-López R, Salgado-Maldonado G, Soto-Galera E, Hernández-Camacho N, Orozco-Zamorano A, Contreras-Robledo S, et al. Helminth parasites of viviparous fishes in México. In: Uribe MC, Grier HJ, editors. Viviparous fishes. Homestead: New Life Publications; 2005. p. 455–74.
Pinacho-Pinacho CD, García-Varela M, Hernández-Orts JS, Mendoza-Palmero CA, Sereno-Uribe AL, Martínez-Ramírez E, et al. Checklist of the helminth parasites of the genus Profundulus Hubbs, 1924 (Cyprinodontiformes, Profundulidae), an endemic family of freshwater fishes in Middle-America. ZooKeys. 2015;523:1–30.
Pinacho-Pinacho CD, Pérez-Ruiz MA, Sereno-Uribe AL, García-Varela M, Martínez-Ramírez E. Richness and similarity of helminth communities of the freshwater fish Profundulus punctatus (Pisces: Cyprinodontidae) from Oaxaca, Mexico. Rev Mex Biodiv. 2014;85(4):1129–38.
Acknowledgments
The authors are indebted to Norman Mercado-Silva (Universidad Autónoma del Estado de Morelos) and to the late Edmundo Díaz-Pardo (Universidad Autónoma de Querétaro) for their help in the collection of fishes in Querétaro; to Eliot Camacho (Centro Acuícola El Geranio, N. Chupícuaro, Gto.) for help in collecting fishes in Guanajuato and Michoacán; to Ismael Guzmán (INECOL) for support in the lab and in the analyses of molecular data; to Octavio Rojas-Soto (INECOL) for assistance in preparing the map; and to Gina Gallo (INECOL) for help in preparing figures. A preliminary version of this study was presented at the XIII ICOPA 2014 conference.
Funding
This research was supported by grants to MRG from Consejo Nacional de Ciencia y Tecnología (CONACyT no. CB168306), Fundación Produce Veracruz/Red Nacional de Información e Investigación en Pesca y Acuicultura (FUNPROVER/RNIIPA), and INECOL. Part of this study was supported by The Royal Society research grant project no. RG150640 awarded to GP. AGV thanks CONACyT for her postdoctoral fellowship (Becario 20340). Funding bodies had no role in the design of the study, and collection, analysis, and interpretation of data, nor in writing the manuscript.
Availability of data and material
The datasets supporting the conclusions of this article are included within the article. Sequences of G. tomahuac n. sp. are submitted in the GenBank database under accession numbers KJ621983 (type-locality) and KR815847–KR815852 (Río Lerma basin paratypes/voucher specimens), while those of G. lamothei are submitted under accession numbers KX555666–KX555668. The type-specimens of G. tomahuac n. sp. are submitted in Colección Nacional de Helmintos, Mexico City.
Authors’ contributions
MRG conceived, designed and coordinated the study. MRG, URM, AGV, MAF, APS, GP collected and analysed the data. MRG drafted and corrected the manuscript. All authors read and approved the final manuscript.
Competing interests
The authors declare that they have no competing interests.
Consent for publication
Not applicable.
Ethics approval and consent to participate
To conduct this work, permits were obtained from the Mexican Ministries of Agriculture, Livestock, Rural Development, Fisheries and Food (SAGARPA) and of the Environment and Natural Resources (SEMARNAT) under scientific collection permits DGOPA/01173/120208.0107 and SGPA/DGVS/02967/14, respectively. Field sampling in Mexico did not require additional approval by an Institutional Animal Care and Use Committee (IACUC) or equivalent animal ethics committee; nor consent to participate. Fish were killed humanely in the field by pithing and then preserved in 96% ethanol.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Rubio-Godoy, M., Razo-Mendivil, U., García-Vásquez, A. et al. To each his own: no evidence of gyrodactylid parasite host switches from invasive poeciliid fishes to Goodea atripinnis Jordan (Cyprinodontiformes: Goodeidae), the most dominant endemic freshwater goodeid fish in the Mexican Highlands. Parasites Vectors 9, 604 (2016). https://doi.org/10.1186/s13071-016-1861-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-016-1861-2