
Abstract
Background
Efficacy of artemisinin (ART) agents, a critical element of current malaria control efforts is threatened by emergence and spread of resistance. Mutations in pfkelch13 gene associated with ART-resistance evolved in Southeast Asia (SEA). k13 mutations whose role in ART-resistance remains unknown, have subsequently emerged independently across all malaria-affected regions. The aim of this systematic review was to determine the prevalence and identify risk factors of Plasmodium falciparum k13 mutations in malaria-endemic countries.
Methods
An electronic search of studies from 2014 to date was done in MEDLINE via PubMED, SCOPUS, EMBASE and LILACS/VHL databases. Mesh terms and Boolean operators (AND, OR) were used. Two librarians independently conducted this search (RS and AK). The articles were screened for inclusion using a priori criteria set following PRISMA-P and STREGA guidelines. Three independent reviewers (NL, BB, and OM) extracted the data. Data analysis was performed in Open Meta Analyst software. Random effects analysis (DL) was used and heterogeneity established using I2-statistic.
Results
A total of 482 articles were retrieved from Pubmed = 302, Lilacs/Vhl = 50, Embase = 80, and Scopus = 37; Bibliography/other searches = 13, of which 374 did not meet the inclusion criteria. The aggregate prevalence of single nucleotide polymorphisms (SNPs) in pfkelch13 gene was 27.6% (3694/14,827) (95% CI 22.9%, 32.3%). Sub-group analysis showed that aggregate prevalence of non-synonymous SNPs in pfkelch13 gene was higher, 45.4% (95% CI 35.4%, 55.3%) in Southeast Asia as opposed to 7.6% (95% CI 5.6%, 9.5%) in the African region. A total of 165 independent k13 mutations were identified across malaria-affected regions globally. A total of 16 non-validated k13 mutations were associated with increased ART parasite clearance half-life (t1/2 > 5 h). The majority, 45.5% (75/165), of the mutations were reported in single P. falciparum parasite infections. Of the 165 k13-mutations, over half were reported as new alleles. Twenty (20) non-propeller mutations in the pfkelch13 gene were identified.
Conclusion
This review identified emergence of potential ART-resistance mediating k13 mutations in the African region. Diversity of mutations in pfkelch13 gene is highest in African region compared to SEA. Mutations outside the pfkelch13 propeller region associated with increased ART parasite clearance half-life occur in malaria-affected regions.
Similar content being viewed by others


Background
The useful life of artemisinin (ART) agents in malaria treatment is at risk due to emergence of several resistance alleles in pfkelch13 gene (k13) [1]. This is further worsened by the current lack of effective vaccine and alternative medicines to ART agents in malaria management. Decreased Plasmodium falciparum sensitivity to ART agents is rapidly evolving and expanding globally since being first reported in southeast Asia (SEA) [2, 3]. Widespread prevalence of parasite ART-resistance threatens current gains in malaria control and eradication efforts. This is especially the case as ART agents remain the only effective malaria treatment to date [4]. Diversity in pfkelch13 gene polymorphisms hinders efforts to track and contain the threat of parasite ART-resistance especially in high malaria transmission regions. In addition, the role of a majority of reported African k13 mutations in causing ART-resistance remains unknown [5].
An increase of 2 million malaria cases was observed in 2017 when 219 million cases were reported compared to 217 million in 2016 [4]. With malaria being one of the leading causes of morbidity and mortality especially in tropical countries, a further increase of an already high burden of malaria is a cause for concern. It is an indication of the need to rethink current malaria control efforts. With no widely effective vaccine, ART agents and their derivatives remain the only effective agents currently used in malaria control efforts. However, decreased P. falciparum parasite sensitivity that has emerged in SEA presents a threat to future efficacy of these agents.
Due to occurrence of multiple polymorphisms in pfkelch13 gene, there is a need to regularly keep track of various emerging alleles in different malaria-affected regions globally. In this systematic review, the aim was to describe potential emergence and/or spread of ART-resistance alleles in pfkelch13 gene across malaria-affected regions. The findings of this review are useful to malaria policy makers and research scientists as it provides a guide on potential k13 mutations whose role in ART-resistance needs to be urgently established, especially in malaria-endemic regions outside SEA.
Methods
Protocol development
In 2017, a review protocol (#CRD 42018084624) was developed and registered in International Prospective Register of Systematic Reviews (PROSPERO: http://www.crd.york.ac.uk/prospero) and thereafter published it in peer review journal [6]. The recommendations of PRISMA-P [7] and STREGA guidelines [8] were observed during protocol development and review conduct.
Review question
The review intended to answer the question: What is the aggregate prevalence of pfkelch13 gene mutations in malaria-affected countries since change in malaria treatment policy to ART agents? [6].
Search strategy
Two librarians (AK and RS) conducted independent search of MEDLINE via PubMED, SCOPUS, EMBASE, and LILACS/VHL databases. The search covered a period from 2014 to date. Database search was performed to identify studies that investigated prevalence of k13 gene polymorphisms among P. falciparum parasites in malaria-affected countries. The following search terms were combined using Boolean operators “OR” for synonymous terms and “AND” across the elements of PECOS (Population, Exposure, Comparator, Outcomes and Study design). The terms used include, “K13-propeller gene polymorphisms”, “K13-polymorphisms”, “K13-gene polymorphisms”, “K13-propeller gene mutations”, “K13-mutation”, “Artemisinin resistance genes”, “Artemisinin resistance alleles” “Artemisinin resistance mutations”, “Artemisinin resistance polymorphisms”, “Plasmodium falciparum”, “Plasmodium parasite”, “Artemisinin”, “Artemether”, “Artesunate”, “Dihydroartemisinin”, “Artemisinin agents”, ACT, ART, “malaria-affected countries”.
Searching for articles in selected databases using the stated search terms was restricted to title or abstract. To maximize outcomes of the search, specific terms were not included such as those that focus on study design. The review found articles in French and Spanish, in which case Google translator (https://translate.google.com/) was used for translation to English prior to screening of the articles for inclusion.
Additional searches
Screening reference lists of included studies was a key targeted article search approach. Further, a researcher in malaria (PR) was contacted for recent publications on k13 gene polymorphisms in P. falciparum in different malaria-affected countries. There was no language restriction in article search and no search of grey literature.
Data management
EndNote software version X7 (Thomson Reuters, 2015) was used in data management for the review articles. All identified titles of articles were imported into the software and duplicates removed. Article titles were then screened and grouped into different eligibility categories (included or excluded) following a priori criteria. Data extraction form was developed in Excel 2007, pre-tested on 5 articles and adjusted using findings of the pilot test. Data were extracted in triplicate and kept using the adjusted tool.
Eligibility criteria for article selection
The review included articles from studies that investigated prevalence of K13-mutations among P. falciparum parasites in all malaria-affected regions/countries. Studies included were those that assessed for molecular markers of ART-resistance in pfkelch13 gene. Studies that reported both molecular and phenotypic ART-resistance were included. Additionally, studies that assessed for k13 mutations both before and after official introduction of ART combination therapy (ACT) in malaria treatment were included. Studies that analyzed for k13 gene mutations using sequencing techniques were included. All studies that used cross-sectional and clinical trial designs in data collection were included.
Exclusion criteria for ineligible articles
Studies that were excluded focused on dhfr, dhps, pfAP2, PfATPase6/SERCA, and PvK12 resistance alleles, malaria treatment, malaria epidemiology and/or transmission, methods/drug targets, non-Plasmodium malaria parasites, pharmacogenetics, review/opinion papers, non-human participants, mathematical modelling of resistance, studies primarily done prior to official introduction of ART agents in malaria treatment, articles which reported only synonymous k13 mutations, and citations whose full text articles could not be retrieved.
Data extraction
Data extraction form was developed in Microsoft Excel 2007 (Microsoft Corp, Washington, USA) and extracted the following information from included articles: author, year of study, country/region, study design, sample collection method, laboratory where analysis was performed, allele calling algorithm, source of DNA, year when ACT was initiated, period/years covered by study, number of parasite DNA where genotyping was attempted, number of parasite DNA samples where genotyping was successful, k13 mutations confirmed to cause delayed ART parasite clearance (validated SNPs in k13 propeller mutations), non-validated SNPs in k13 gene (k13 mutations that have not been confirmed to cause delayed ART parasite clearance), non-propeller k13 mutations, non-validated k13 mutations associated with delayed ART parasite clearance, nucleotide changes in k13 mutations, genotyping method used, duration of ART use, nature of ART use (monotherapy/combination) and source of k13 mutation.
Minimizing bias in article identification, selection and data extraction
A second librarian (RS) validated electronic search in PubMed by performing an independent and duplicate search. A second reviewer (EAO) screened all full text articles excluded by the first reviewer (MO). Any discrepancies among the reviewers were resolved by discussion and consensus. Two reviewers (NL and BB) performed duplicate and independent data extraction. Any disagreement between the reviewers was resolved by discussion and consensus. Any further disagreement between the two reviewers was referred to the third reviewer for a final decision (MO).
Data synthesis
Extracted data were transferred to Open Meta Analyst software for analysis [9]. Both structured narrative and quantitative syntheses of extracted data were applied as appropriate. In the structured data synthesis descriptive summaries of outcomes of interest were generated. These included: non-synonymous SNPs in pfkelch13 gene, duration of ART use prior to data collection, source of mutations (indigenous/introduced), nature of ART use (combination or monotherapy), allele calling algorithms, nucleotide changes in k13 mutations, spread of k13 mutaions, pfkelch13 mutations not confirmed to cause delayed ART parasite clearance (non-validated) but associated with delayed P. falciparum parasite ART clearance, genotype success rate, frequency of k13 mutations.
DerSimonian–Laird (DL) random effects analysis was used in establishing summary estimate of prevalence of non-synonymous SNPs in pfkelch13 gene in malaria-affected regions using Open Meta Analyst software. Sub-group analysis was performed, region (SEA, Africa, India), and study design (cross-sectional, clinical trial designs). Heterogeneity in included articles was inferred from the summary estimates of I2-statistic. The I2 statistic was used to indicate percentage (%) heterogeneity that could be attributed to between-study variance. Interpretation: I2 = 25% (small heterogeneity), I2 = 50% (moderate heterogeneity), I2 = 75% (large heterogeneity) [10].
Missing data and risk of bias assessment
The variables that were missing from included articles were recorded as not reported. Statistical tests were not used in handling missing data. However, authors whose articles had some missing variables were contacted but did not get any feedback.
The risk of publication bias was assessed using indirect assessment of rank correlation between effect size and sample size (Kendall’s tau) method. In this method correlation of the articles is interpreted from the analysis output in Open Meta (Analyst) software where 1, represent perfect correlation and 0 no correlation.
Results
Study identification and selection
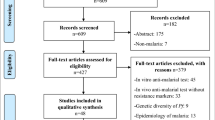
The search of Medline via PubMed, Embase, Lilacs/Vhl and Scopus databases yielded a total of 469 citations on P. falciparum k13 gene single nucleotide polymorphisms (SNPs). Additionally, 12 citations were obtained from bibliography search and one article was obtained through contact with a malaria researcher (PR). After adjusting for duplicates, 409 citations remained. Of these, 296 articles were discarded after reviewing the titles and abstracts as they did not meet a priori inclusion criteria. Five citations were discarded as the full text articles could not be obtained. A total of 113 full text articles were screened in detail using pre-determined inclusion criteria. A total of 50 articles were included in the final full text review (Fig. 1).

Prisma flow diagram showing article selection for the review
Characteristics of included studies
Twenty-five articles were from studies in SEA, 2 from India, 21 from Africa and one from studies both in Africa and China. Two studies, by Ashley et al. [11] and Menard et al. [12] were performed in multiple centers both in Africa and SEA. The reported study designs included: randomized control trials (3), cohort (2), non-randomized trials (2), and cross-sectional studies (43).
Over half, 52% (26/50) of included articles did not report the year of official initiation of ART use in malaria treatment in the region/country where data were collected. While in 24 studies that reported years of official introduction of ART use, the mean duration of time from official initiation of ACT use to data collection was 9.3 ± 7.2 years. In addition, countries where studies were conducted, ART agents had been in use in malaria treatment for on average 15.4 ± 7.7 years from official introduction to date (2018). The majority of studies on P. falciparum k13 mutation were done after official introduction of ART agents in malaria treatment.
One study by Ariey et al. [1] used laboratory-adapted P. falciparum parasites from both SEA and sub-Saharan Africa while all other studies collected P. falciparum parasite isolates from symptomatic patients. ART agents were reportedly used as combination therapy in malaria treatment in all malaria-affected countries from where data were collected.
Genotyping errors
Genotyping was conducted in total of 31,655 P. falciparum DNA isolates of which 27,667 (87.4%) were successfully genotyped (Table 1). The genotypes were assigned to successfully sequenced parasite DNA in small batches in all studies. Twenty-five studies collected P. falciparum-infected blood samples using filter papers (Whatman 903), EDTA vacutainer tubes (14 studies) while 11 studies did not report how samples were collected. The allele calling algorithms used included: Mega software ver 6.06, Bioedit ver 7.0.1, Sequencer software ver 5.4.5, Jalview software ver 5.1, Lasergene Genomic Suite, DnaSP software ver 5.50, Genome Assembly Program GAP4, Genescan, PROVEAN software tool, R software, Genotyper software and CEQ 2000 genetic Analysis System Software.
Heterogeneity in the included studies
Heterogeneity was assessed in different sub-groups based on study design. The sub-groups included: cross-sectional studies (43), randomized control trials (RCT) (3: [11, 13, 14]), cohort (2: [15, 16]), and non-randomized clinical trials (2: [17, 18]). There was high heterogeneity across all sub-groups, cross-sectional studies (I2 = 99.38%, P < 0.001), RCTs (I2 = 74.4%, P = 0.02) and cohort studies (I2 = 99.81%, P < 0.001). Heterogeneity was still high when assessed across regions, SEA (I2 = 99.68%, P < 0.001), and Africa (I2 = 95.81%, P < 0.001).
Codon positions where SNPs occur in pfkelch13 gene
The majority, 86.7% (143/165) of observed non-synonymous SNPs occur within propeller region of pfkelch13 gene. Of 84 non-synonymous SNPs in K13-gene in SEA, 66 (78.6%) are in propeller region (≥ 441) while 18 (21.4%) are out-side the propeller region (≤ 440) of pfkelch13 gene. The non-synonymous SNPs in k13 gene that occurred out-side propeller region in SEA included N87K (1 infected patient), G112E (1 infected patient), E130G (1 infected patient), L143P (1 infection), T149S (1 infection), K189T (31 infections), K189N (2 infections), R223K (1 infection), R255K (3 infections), D281V (1 infection), R254L (1 infection), I376V (1 infection) and E252Q (4 infections), K438N (1 infection), N11Y (1 infection), I352T (2 infections), E294G (1 infection), I205T (1 infection).
While of 76 non-synonymous SNPs in k13 gene reported among sub-Saharan African parasites, 71 (93.4%) were in propeller region of pfkelch13 gene. The k13 mutations that occurred outside propeller region of pfkelch13 gene included K123R (1 infected patient), N137S (1 infected patient), T149S (1 infected patient), K189N (1 infected patient), K189T (18 infected patients). The review observed that a total of 22 non-synonymous SNPs, 13.3% (22/165) in the pfkelch13 gene occur outside the propeller region. In India, of the 07 k13 mutations only one mutation K189T (1 infection) occurred outside the propeller region.
Double mutations in the same codon was reported at codon positions K189T/N [11, 19, 20]); N554L/H/S [21,22,23,24]); S522C/A [11]; [12, 25, 26]); P637I/D [20, 22, 27]); D648N/Y/H [21,22,23,24]); M476I/K/V [11, 13, 28,29,30,31] (Table 1). The nucleotide changes in codon positions with multiple mutations are, P637 → I, D (GTT → ATT or GTT → AAT), K189 → T, N (AAA → ACA or AAA → AAT), S522 → C, A (AGT → TGT or AGT → AGG), N554 → L, H or S (AAT → AGT, AAT → CAT or AAT → AGT) and D648 → N, Y or H (GAT → AAT, GAT → TAT or GAT → CAT).
Overall prevalence of non-synonymous SNPs in pfkelch13 gene
The overall prevalence of non-synonymous SNPs in pfkelch13 gene (validated and non-validated) in reviewed articles from malaria-endemic countries is 27.6% (3694/14,827) (95% CI 22.9–32.3%). In SEA, the observed prevalence of validated k13 mutations is 31.7% (1268/5328) (95% CI 19.3–44%). While the overall prevalence of non-synonymous SNPs (validated and non-validated) in SEA is 45.4% (4246/9352) (95% CI 35.4–55.3%). In African region the overall prevalence of non-synonymous SNPs (non-validated) in the k13 gene is 7.6% (415/5450) (95% CI 5.6–9.5%). In India, overall prevalence of non-synonymous k13 mutations is 2.2% (14/638).
Epidemiology of non-synonymous SNPs in pfkelch13 gene
Of 165 non-synonymous SNPs in pfkelch13 gene reported in this review, 75 (45.5%) were found in single P. falciparum parasite infections. In SEA, 24 of 84 (28.6%) reported SNPs were found in single P. falciparum parasite infections. The validated k13 mutation (confirmed to cause delayed ART parasite clearance) found in most malaria parasite infections (884) in SEA was C580Y. The non-validated k13 gene mutations occurring in a high number of P. falciparum infections in SEA include F446I (242), G449A (20), N458Y (22), 479 (30), 537 (31), P574L (100), A675V (28), G538V (36), and R561H (39). In Africa, 47 of 76 (61.8%) reported non-synonymous k13 mutations were found in single P. falciparum parasite infections. The most common k13 mutation, 40.8% (31/76) in Africa is A578S. The other non-synonymous SNPs in pfkelch13 gene frequently reported among African parasite infections include 189T (18), D464H (09), D584V (05), V637I/D (05), N554L (04) and D648Y/H (03).
The k13 mutation P574L that commonly occurs in SEA was reported in Rwanda [32] and in a study on immigrants from Africa to China [31]. The other mutations reported in low frequency in both Africa and SEA include A675V, P584L, R575K, 522C 189T, R561H, M476I, F446I, N554S, and A578S. Two studies by Feng et al. [33] and Yang et al. [31] in both Africa and China, reported a total of 21 different k13 gene mutations of which 20 were found in only one parasite infection. Twenty of the 74 (27%) non-synonymous k13 mutations in Africa were also reported in SEA. In India, of 5 reported non-synonymous k13 mutations, 4 were found in only one P. falciparum parasite infection while a mutation, A578S was found in 2 P. falciparum infections in India. All the mutations found in India were also reported in other malaria-endemic regions. In China, the most commonly reported k13 propeller gene mutations are F446I and P574L. The other SNPs reported in low frequency in China include H719N, M579I, F495L, L492S, C469F/Y, and N458Y (Table 1).
Occurrence of k13 gene mutations and ART use
In some studies in Africa, k13 gene mutations were reported among P. falciparum parasites prior to official introduction of ACT in malaria treatment. These mutations include: E602D (Djaman et al. [34]: Ivory Coast); Y588C, E556K (Guerra et al. [27]: Equatorial Guinea). A study in Thailand by Putaporntip et al. [35] also reported existence of E605G and N609S mutations prior to introduction of ACT in malaria treatment. Of these mutations only one SNP at codon position E556K of pfkelch13 gene persisted even after introduction of ART use in malaria treatment [27, 31]. A different allele at the same codon position 556 (E556D) was reported in SEA [29, 35,36,37].
Of 50 reviewed articles, 83% (34) indicated that the k13 mutations reported were indigenous to the region/country where the study was conducted. New SNPs in pfkelch13 gene were reported each time in over half, 54% (27/50) of the studies. In SEA, ART agents have been in use in malaria treatment for close to five decades [38]. While in Africa, ACT has been used in malaria treatment for over a decade (~ 15 years).
Non-validated, non-synonymous SNPs in pfkelch13 gene associated with delayed ART parasite clearance
A total of 16 non-validated, non-synonymous k13 mutations associated with delayed ART parasite clearance were reported by different studies, of which F446I (98 infections), P574L (61 infections), N458Y (22 infections), M476I (30 infections), A675V (28 infections), N537D (31 infections), and K189T (49 infections) were the most reported. A mutation at codon position A675V was reported in SEA, Uganda and Rwanda (East Africa) while Q613E and K189T were reported in different studies done in Senegal (West Africa) and SEA.
Non-synonymous SNPs in pfkelch13 gene associated with delayed ART clearance among African P. falciparum parasites include: Q613H (Senegal [22], P574L (Rwanda [32]; China–Africa [31], A675V (Rwanda: [32]; Uganda: [39], P553L (Kenya: [26], and K189T (Senegal: [22]; Uganda: [20]) (Table 2).
Nucleotide changes of most reported non-validated k13 gene SNPs
Single nucleotide polymorphism at codon position 574 (CCT → CTT) of the k13 gene was reported by up to 5 different studies. The majority of frequently reported non-validated non-synonymous SNPs in k13 gene occurred among African P. falciparum parasites (Table 3).
Non-synonymous SNPs in non-propeller region (< 440) of pfkelch13 gene
The most frequently reported non-propeller k13 mutation occurred at codon position K189T. The other common non-propeller k13 mutations are located at codon positions 255, 252 and 352 of the pfkelch13 gene. Non-propeller SNP at codon position K189T has been associated with delayed ART P. falciparum parasite clearance (t1/2 > 5 h) [11]. Mutation at codon position K189T was reported in both Africa and SEA (Table 4).
Discussion
Malaria incidence rate reduced by 18% globally, from 76 to 63 cases per 1000 population at risk between 2010 and 2016 [4]. However, potential widespread emergence of P. falciparum parasite, mosquito resistance to ART agents and insecticides, respectively, is a threat to these gains in malaria control and elimination [4]. The review observed that overall prevalence of non-synonymous SNPs in propeller region of pfkelch13 gene (both validated and non-validated) in malaria-affected regions is 27.6%. k13 mutations confirmed to cause delayed ART parasite clearance (validated k13 mutations) were observed in only SEA with aggregate prevalence of 45.4%. The observed prevalence of non-synonymous k13 mutations varied in different regions with SEA, having the highest prevalence of k13 mutations, while African region had higher diversity and low frequency of reported polymorphisms.
Since the first discovery of k13 mutations as independent markers of ART-resistance in Western Cambodia [2] and later in Thai-Myanmar [40], there has been an overall upward trend in allele frequency globally [14]. In this review up to 165 k13 mutations are reported across malaria-affected regions with 110 being reported in only one geographical location (SEA). Only 20% of the mutations reported in Africa were also detected in SEA. In addition, new mutations were reported each time in over 50% of the studies. This review highlights presence of diverse but low frequency occurrence of non-synonymous SNPs in pfkelch13 gene across malaria-affected regions globally. In addition, the role of a majority of these alleles in parasite ART-resistance remains unknown.
Mutations R539T, C580Y (validated) and P574L (candidate) were reported in China-Africa studies. A study by Yang et al. [31] reported R539T and P574L among parasites from Angola and Equatorial Guinea, while Huang et al. [36] found C580Y and R539T among P. falciparum parasites isolated among immigrants to China from Ghana. These mutations could have been imported to China from Africa and is not clear whether they spread to Africa from SEA or emerged independently. A majority of the k13 mutations arise independently as indicated by studies done in SEA [14, 41]. The current reported SEA alleles associated with ART-resistance among P. falciparum parasites observed in Africa and China could be due to independent emergence. Independent emergence of resistance alleles in pfkelch13 gene potentially responsible for the observed variations in k13 mutations across malaria-affected regions is influenced by several factors. These include widespread uncontrolled use of ART agents in addition to occurrence of anti-malarial medicines with sub-therapeutic amounts of active agents, especially in low and middle income countries. The extent to which different regions/countries are affected by these factors differs and could contribute to observed variations in k13 mutations. Furthermore, existence of different parasite founder populations (genetic background) [42] in addition to variations in environmental conditions influences independent emergence of k13 mutations.
Over half of the observed k13 mutations were each reported in a single P. falciparum parasite infection. A study by Straimer et al. [43] showed that each single nucleotide change in pfkelch13 gene is independently associated with an increase in parasite survival during ART exposure. Different k13 propeller mutations mediate different levels of ART-resistance [16, 40]. Currently 6 ART-resistance alleles (N458Y, Y493H, R539T, I543T, R561H, C580Y) are confirmed (validated) as causes of ART parasite resistance [1]. Overtime a mutation at codon position C580Y has emerged as the most dominant in SEA and is associated with highest parasite clearance half-life [14]. The variation in extent to which these mutations affect ART parasite clearance half-life may be attributed to involvement of other loci in the genome [41, 42]. The numbers of k13 mutations that are associated with increased clearance half-life of P. falciparum parasites by ART agents are rising over time. This has presented a challenge to policy makers and scientists who need to keep up with frequently validating different mutations. The review reports 16 k13 mutations that are not validated (not confirmed) despite being associated with an increase in ART parasite clearance half-life. Some of the 16 potential ART-resistance alleles occur among African P. falciparum parasites (K189T, P553L, P574L, A675V, M476K, and Q613H). Despite the current association of some of these alleles with increased parasite clearance half-life, there is a need to validate all frequently reported SNPs in the k13 gene among P. falciparum parasite populations in different regions, especially in Africa, which bears over 90% of malaria burden. A recent study by Mukherjee et al. showed that parasites carrying k13 mutation D584V had increased in vitro RSA0–3 h survival. The mutation D584 V was reported in SEA [17, 44] and among African migrants to China [36].
In the current review, 20 of 165 observed k13 mutations occur outside propeller region of pfkelch13 gene. Two of these mutations (A189T and E252Q) have been associated with increased ART parasite clearance half-life [42]. A mutation in codon position K189T was reported in both SEA and Africa. Molecular screening for ART-resistance currently is based on sequencing propeller region of pfkelch13 gene (1, 725,980–1726, 940 bp; amino acid positions 419–707) [1]. However, this review shows that studies have recently established existence of non-synonymous mutations outside the propeller region of pfkelch13 gene that are associated with increased ART parasite clearance half-life. In Cambodia, a mutation E252Q was reported by Wang et al. [19], Tun et al. [30], Ashley et al. [11], while D281V and K189T were in a study by Ashley et al. [11]. In Africa, mutation K189T was reported by Boussaroque et al. [22] (Senegal) and Conrad et al. [20] (Uganda). These mutations may provide valuable clues for the existence of gene polymorphisms potentially associated with ART-resistance outside the propeller region of pfkelch13 gene [42].
On the Thai-Myanmar border, ACT was first introduced in malaria treatment in 1994 with resistance suspected just over one decade later, in 2008, and subsequently confirmed in a study by Ariey et al. [1]. The review observed non-synonymous k13 mutations among P. falciparum parasites that occurred prior to official introduction of ART agents in malaria treatment. These included: E602D, I588C, E252Q, and E556K and were all reported in African P. falciparum parasites. Only one of these mutations, E556K, persisted in parasite population even after official introduction of ART agents in malaria treatment. A study by Hermission [45] indicated that ART-resistance alleles in k13 gene became established within a year of official ART deployment in malaria treatment in SEA. In malaria-affected countries outside SEA however, after over a decade of ART use in malaria treatment, there is still no confirmed k13 resistant alleles among parasite population. In Africa, which bears over 90% of malaria burden, this has been welcome news. However, current resistance surveillance reports have indicated potential emergence of parasites with increased ART clearance half-life in the region.
Although ART agents with effective partner drugs still cure patients harbouring slow ART clearing parasites [14, 46], resistance development to ART component of the combination will expose more parasites to partner drug alone, increasing likelihood of ACT failure. The current failure by dihydroartemisinin-piperaquine combination to cure patients in Western Cambodia is a demonstration of this phenomenon [14]. The review observed that ART is used as combination agents in malaria treatment across malaria-affected countries. The use of combination ART agents (ACT) in malaria treatment potentially reduces resistance development [47]. However, eradication of ART-resistant malaria parasites necessitates complete elimination of all malaria since, as malaria incidence reduces, the few infections which remain to be cleared are almost all resistant [47]. Using long-lasting insecticide-treated mosquito nets alongside ACT could go a long way in eradicating malaria disease due to the synergistic effect on malaria incidence [47].
Use of ART agents in malaria treatment influences development of parasite resistance. The review observed emergence of multiple k13 mutations across malaria-affected countries following introduction of ART agents. A study by Amaratunga et al. [48] showed that ART agents select for resistance among P. falciparum parasites. In addition, it is probable that use of partner drugs such as mefloquine [49], piperaquine [14], and lumefantrine [50] to which the parasites are resistant may be influencing ART-resistance development. The magnitude of ACT selective pressure on development of parasite ART-resistance is thus likely to be greater, especially in infections with concomitant resistance to partner drugs [47]. The current malaria combination therapy may benefit from having more than two active pharmaceutical ingredients. The use of combination therapy having more than two active ingredients has proved effective in restraining HIV resistance development [51]. However, treatment of infectious diseases in most low and middle income countries is faced with the challenge of limited therapeutic options affecting potential use of more than two active ingredients in malaria combination therapy.
The review had some limitations, few studies were retrieved and reviewed from India and none from Central Asia and South America. This was mainly due to poorly designed studies, which were excluded. However, the application of extensive literature search and a priori selection criteria reduces the potential effect of this limitation on the overall review outcome. Most of the articles poorly reported their findings with some omitting a few review variables. To reduce the effect of missing variables, outcomes were recalculated for the review variables using data reported in the articles. In addition, corresponding authors of the articles were contacted for missing information, although no response was received.
Conclusion
The number of potential ART resistance alleles in the pfkelch13 gene is rising across malaria-affected regions. The k13 mutations with potential role in ART-resistance have been reported to occur outside the propeller region of pfkelch13 gene. Among African P. falciparum parasites although not validated, k13 mutations associated with delayed parasite clearance have been reported. Overall prevalence of non-synonymous SNPs in the k13 gene is higher in SEA with higher diversity observed among African parasites. k13 mutations occurring in more than one geographical region affected by malaria is increasing, however, polymorphisms associated with ART resistance have not yet been observed outside SEA.
Abbreviations
- PRISMA:
-
preferred reporting items for systematic reviews and meta-analysis
- STROBE:
-
strengthening reporting of observational studies in epidemiology
- PROSPERO:
-
International Prospective Register of Systematic Reviews
- STREGA:
-
Strengthening the Reporting of Genetic Association Studies
References
Ariey F, Witkowski B, Amaratunga C, Beghain J, Langlois AC, Khim N, et al. A molecular marker of artemisinin-resistant Plasmodium falciparum malaria. Nature. 2014;505:50–5.
Noedl H, Se Y, Schaecher K, Smith BL, Socheat D, Fukuda MM. Evidence of artemisinin-resistant malaria in western Cambodia. N Engl J Med. 2008;359:2619–20.
Dondorp AM, Nosten F, Yi P, Das D, Phyo AP, Tarning J, et al. Artemisinin resistance in Plasmodium falciparum malaria. N Engl J Med. 2009;361:455–67.
WHO. World malaria report 2018. Geneva: World Health Organization; 2018.
Kamau E, Campino S, Amenga-Etego L, Drury E, Ishengoma D, Johnson K, et al. K13-propeller polymorphisms in Plasmodium falciparum parasites from sub-Saharan Africa. J Infect Dis. 2015;211:1352–5.
Ocan M, Akena D, Nsobya S, Kamya MR, Senono R, Kinengyere AA, et al. K13-propeller gene polymorphisms in Plasmodium falciparum parasite population: a systematic review protocol of burden and associated factors. Syst Rev. 2018;7:199.
Moher D, Stewart L, Shekelle P. Implementing PRISMA-P: recommendations for prospective authors. Syst Rev. 2016;5:15.
Little J, Higgins JPT, Ioannidis JPA, Moher D, Gagnon F, von Elm E, et al. STrengthening the REporting of Genetic Association Studies (STREGA)—an extension of the STROBE statement. PLoS Med. 2009;6:e1000022.
Wallace BC, Schmid CH, Lau J, Trikalinos TA. Meta-analyst: software for meta-analysis of binary, continuous and diagnostic data. BMC Med Res Methodol. 2009;9:80.
Kontopantelis E, Springate DA, Reeves D. A re-analysis of the Cochrane Library Data: the dangers of unobserved heterogeneity in meta-analyses. PLoS ONE. 2013;8:e69930.
Ashley EA, Dhorda M, Fairhurst RM, Amaratunga C, Lim P, Suon S, et al. Spread of artemisinin resistance in Plasmodium falciparum malaria. N Engl J Med. 2014;371:411–23.
Ménard D, Khim N, Beghain J, Adegnika AA, Shafiul-Alam M, Amodu O, et al. A worldwide map of Plasmodium falciparum K13-propeller polymorphisms. N Engl J Med. 2016;374:2453–64.
Bonnington CA, Phyo AP, Ashley EA, Imwong M, Sriprawat K, Parker DM, et al. Plasmodium falciparum kelch 13 mutations and treatment response in patients in Hpa-Pun District, Northern Kayin State, Myanmar. Malar J. 2017;16:480.
Takala-Harrison S, Jacob CG, Arze C, Cummings MP, Silva JC, Dondorp AM, et al. Independent emergence of artemisinin resistance mutations among Plasmodium falciparum in southeast Asia. J Infect Dis. 2015;211:670–9.
Nyunt MH, Hlaing T, Oo HW, Tin-Oo LL, Phway HP, Wang B, et al. Molecular assessment of artemisinin resistance markers, polymorphisms in the k13 propeller, and a multidrug-resistance gene in the eastern and western border areas of Myanmar. Clin Infect Dis. 2015;60:1208–15.
Spring MD, Lin JT, Manning JE, Vanachayangkul P, Somethy S, Bun R, et al. Dihydroartemisinin–piperaquine failure associated with a triple mutant including kelch13 C580Y in Cambodia: an observational cohort study. Lancet Infect Dis. 2015;15:683–91.
Leang R, Taylor WR, Bouth DM, Song L, Tarning J, Char MC, et al. Evidence of Plasmodium falciparum malaria multidrug resistance to artemisinin and piperaquine in Western Cambodia: dihydroartemisinin-piperaquine open-label multicenter clinical assessment. Antimicrob Agents Chemother. 2015;59:4719–26.
Bayih AG, Getnet G, Alemu A, Getie S, Mohon AN, Pillai DR. A unique Plasmodium falciparum k13 gene mutation in Northwest Ethiopia. Am J Trop Med Hyg. 2016;94:132–5.
Wang Z, Shrestha S, Li X, Miao J, Yuan L, Cabrera M, et al. Prevalence of K13-propeller polymorphisms in Plasmodium falciparum from China-Myanmar border in 2007–2012. Malar J. 2015;14:168.
Conrad MD, Bigira V, Kapisi J, Muhindo M, Kamya MR, Havlir DV, et al. Polymorphisms in K13 and falcipain-2 associated with artemisinin resistance are not prevalent in Plasmodium falciparum isolated from Ugandan Children. PLoS ONE. 2014;9:e105690.
Win AA, Imwong M, Kyaw MP, Woodrow CJ, Chotivanich K, Hanboonkunupakarn B, et al. K13 mutations and pfmdr1 copy number variation in Plasmodium falciparum malaria in Myanmar. Malar J. 2016;15:110.
Boussaroque A, Fall B, Madamet M, Camara C, Benoit N, Fall M, et al. Emergence of mutations in the K13 propeller gene of Plasmodium falciparum solates from Dakar, Senegal, in 2013–2014. Antimicrob Agents Chemother. 2016;60:624–7.
Isozumi R, Uemura H, Kimata I, Ichinose Y, Logedi J, Omar AH, et al. Novel mutations in K13 propeller gene of artemisinin-resistant Plasmodium falciparum. Emerg Infect Dis. 2015;21:490–2.
Torrentino-Madamet M, Collet L, Lepere JF, Benoit N, Amalvict R, Menard D, et al. K13-propeller polymorphisms in Plasmodium falciparum isolates from patients in Mayotte in 2013 and 2014. Antimicrob Agents Chemother. 2015;59:7878–81.
Dorkenoo AM, Yehadji D, Agbo YM, Layibo Y, Agbeko F, Adjeloh P, et al. Therapeutic efficacy trial of artemisinin-based combination therapy for the treatment of uncomplicated malaria and investigation of mutations in k13 propeller domain in Togo, 2012–2013. Malar J. 2016;15:331.
Taylor SM, Parobek CM, DeConti DK, Kayentao K, Coulibaly SO, Greenwood BM, et al. Absence of putative artemisinin resistance mutations among Plasmodium falciparum in sub-Saharan Africa: a molecular epidemiologic study. J Infect Dis. 2015;211:680–8.
Guerra M, Neres R, Salgueiro P, Mendes C, Ndong-Mabale N, Berzosa P, et al. Plasmodium falciparum genetic diversity in continental Equatorial Guinea before and after introduction of artemisinin-based combination therapy. Antimicrob Agents Chemother. 2017;61:e02556.
Nyunt MH, Wang B, Aye KM, Aye KH, Han J-H, Lee S-K, et al. Molecular surveillance of artemisinin resistance falciparum malaria among migrant goldmine workers in Myanmar. Malar J. 2017;16:97.
Nyunt MH, Soe MT, Myint HW, Oo HW, Aye MM, Han SS, et al. Clinical and molecular surveillance of artemisinin resistant falciparum malaria in Myanmar (2009–2013). Malar J. 2017;16:333.
Tun KM, Imwong M, Lwin KM, Win AA, Hlaing TM, Hlaing T, et al. Spread of artemisinin-resistant Plasmodium falciparum in Myanmar: a cross-sectional survey of the K13 molecular markrer. Lancet Infect Dis. 2015;15:415–21.
Yang C, Zhang H, Zhou R, Qian D, Liu Y, Zhao Y, et al. Polymorphisms of Plasmodium falciparum k13-propeller gene among migrant workers returning to Henan Province, China from Africa. BMC Infect Dis. 2017;17:560.
Tacoli C, Gai PP, Bayingana C, Sifft K, Geus D, Ndoli J, et al. Artemisinin resistance-associated K13 polymorphisms of Plasmodium falciparum in Southern Rwanda, 2010–2015. Am J Trop Med Hyg. 2016;95:1090–3.
Feng J, Li J, Yan H, Feng X, Xia Z. Evaluation of antimalarial resistance marker polymorphism in returned migrant workers in China. Antimicrob Agents Chemother. 2015;59:326–30.
Djaman JA, Olefongo D, Ako AB, Roman J, Ngane VF, Basco LK, et al. Molecular epidemiology of malaria in Cameroon and Cote d’Ivoire. XXXI. Kelch 13 propeller sequences in Plasmodium falciparum isolates before and after implementation of artemisinin-based combination therapy. Am J Trop Med Hyg. 2017;97:222–4.
Putaporntip C, Kuamsab N, Kosuwin R, Tantiwattanasub W, Vejakama P, Sueblinvong T, et al. Natural selection of K13 mutants of Plasmodium falciparum in response to artemisinin combination therapies in Thailand. Clin Microbiol Infect. 2016;22:285.
Huang F, Takala-Harrison S, Jacob CG, Liu H, Sun X, Yang H, et al. A single mutation in K13 predominates in Southern China and is associated with delayed clearance of Plasmodium falciparum following artemisinin treatment. J Infect Dis. 2015;212:1629–35.
Talundzic E, Okoth SA, Congpuong K, Plucinski MM, Morton L, Goldman IF, et al. Selection and spread of artemisinin-resistant alleles in Thailand prior to the global artemisinin resistance containment campaign. PLoS Pathog. 2015;11:e1004789.
Wang Z, Wang Y, Cabrera M, Zhang Y, Gupta B, Wu Y, et al. Artemisinin resistance at the China-Myanmar border and association with mutations in the K13 propeller gene. Antimicrob Agents Chemother. 2015;59:6952–9.
Ikeda M, Kaneko M, Tachibana S-I, Balikagala B, Sakurai-Yatsushiro M, Yatsushiro S, et al. Artemisinin-resistant Plasmodium falciparum with high survival rates, Uganda, 2014–2016. Emerg Infect Dis. 2018;24:718–26.
Phyo AP, Nkhoma S, Stepniewska K, Ashley EA, Nair S, McGready R, et al. Emergence of artemisinin-resistant malaria on the western border of Thailand: a longitudinal study. Lancet. 2012;379:1960–6.
Anderson TJC, Nair S, McDew-White M, Cheeseman IH, Nkhoma S, Bilgic F, et al. Why are there so many independent origins of artemisinin resistance in malaria parasites? Biorxiv. 2016;1:1. https://doi.org/10.1101/056291.
Miotto O, Amato R, Ashley EA, MacInnis B, Almagro-Garcia J, Amaratunga C, et al. Genetic architecture of artemisinin-resistant Plasmodium falciparum. Nat Genet. 2015;47:226–34.
Straimer J, Lee MCS, Lee AH, Zeitler B, Williams AE, Pearl JR, et al. Site-specific editing of the Plasmodium falciparum genome using engineered zinc-finger nucleases. Nat Methods. 2012;9:993–8.
Imwong M, Jindakhad T, Kunasol C, Sutawong K, Vejakama P, Dondorp AM. An outbreak of artemisinin resistant falciparum malaria in Eastern Thailand. Sci Rep. 2015;5:17412.
Hermisson J, Pennings PS. Soft sweeps: molecular population genetics of adaptation from standing genetic variation. Genetics. 2005;169:2335–52.
Lopera-Mesa TM, Doumbia S, Chiang S, Zeituni AE, Konate DS, Doumbouya M, et al. Plasmodium falciparum clearance rates in response to artesunate in Malian children with malaria: effect of acquired immunity. J Infect Dis. 2013;207:1655–63.
Maude RJ, Pontavornpinyo W, Saralamba S, Aguas R, Yeung S, Dondorp AM, et al. The last man standing is the most resistant: eliminating artemisinin-resistant malaria in Cambodia. Malar J. 2009;8:31.
Amaratunga C, Witkowski B, Dek D, Try V, Khim N, Miotto O, et al. Plasmodium falciparum founder populations in Western Cambodia have reduced artemisinin sensitivity in vitro. Antimicrob Agents Chemother. 2014;58:4935–7.
Denis MB, Tsuyuoka R, Poravuth Y, Narann TS, Seila S, Lim C, et al. Surveillance of the efficacy of artesunate and mefloquine combination for the treatment of uncomplicated falciparum malaria in Cambodia. Trop Med Int Health. 2006;11:1360–6.
Denis MB, Tsuyuoka R, Lim P, Lindegardh N, Yi P, Top SN, et al. Efficacy of artemether-lumefantrine for the treatment of uncomplicated falciparum malaria in northwest Cambodia. Trop Med Int Health. 2006;11:1800–7.
Feder AF, Rhee SY, Holmes SP, Shafer RW, Petrov DA, Pennings PS. More effective drugs lead to harder selective sweeps in the evolution of drug resistance in HIV-1. eLife. 2016;5:e10670.
Bosman P, Stassijns J, Nackers F, Canier L, Kim N, Khim S, et al. Plasmodium prevalence and artemisinin-resistant falciparum malaria in Preah Vihear Province, Cambodia: a cross-sectional population-based study. Malar J. 2014;13:394.
Dwivedi A, Khim N, Reynes C, Ravel P, Ma L, Tichit M, et al. Plasmodium falciparum parasite population structure and gene flow associated to anti-malarial drugs resistance in Cambodia. Malar J. 2016;15:319.
Imwong M, Suwannasin K, Kunasol C, Sutawong K, Mayxay M, Rekol H, et al. The spread of artemisinin-resistant Plasmodium falciparum in the Greater Mekong subregion: a molecular epidemiology observational study. Lancet Infect Dis. 2017;17:491–7.
Kheang ST, Sovannaroth S, Ek S, Chy S, Chhun P, Mao S, et al. Correction to: prevalence of K13 mutation and Day-3 positive parasitaemia in artemisinin-resistant malaria endemic area of Cambodia: a cross-sectional study. Malar J. 2017;16:435.
Mohon AN, Alam MS, Bayih AG, Folefoc A, Shahinas D, Haque R, et al. Mutations in Plasmodium falciparum K13 propeller gene from Bangladesh (2009–2013). Malar J. 2014;13:431.
Thuy-Nhien N, Tuyen NK, Tong NT, Vy NT, Thanh NV, Van HT, et al. K13 Propeller mutations in Plasmodium falciparum populations in regions of malaria endemicity in Vietnam from 2009 to 2016. Antimicrob Agents Chemother. 2017;61:e01578.
Nyunt MH, Shein T, Zaw NN, Han SS, Muh F, Lee S-K, et al. Molecular evidence of drug resistance in asymptomatic malaria infections, Myanmar, 2015. Emerg Infect Dis. 2017;23:517–20.
Mishra N, Prajapati SK, Kaitholia K, Bharti RS, Srivastava B, Phookan S, et al. Surveillance of artemisinin resistance in Plasmodium falciparum in India using the kelch13 molecular marker. Antimicrob Agents Chemother. 2015;59:2548–53.
Mishra N, Bharti RS, Mallick P, Singh OP, Srivastava B, Rana R, et al. Emerging polymorphisms in falciparum Kelch 13 gene in Northeastern region of India. Malar J. 2016;15:583.
Balikagala B, Mita T, Ikeda M, Sakurai M, Yatsushiro S, Takahashi N, et al. Absence of in vivo selection for K13 mutations after artemether–lumefantrine treatment in Uganda. Malar J. 2017;16:23.
Escobar C, Pateira S, Lobo E, Lobo L, Teodosio R, Dias F, et al. Polymorphisms in Plasmodium falciparum K13-propeller in Angola and Mozambique after the introduction of the ACTs. PLoS ONE. 2015;10:e0119215.
Gupta H, Macete E, Bulo H, Salvador C, Warsame M, Carvalho E, et al. Drug-resistant polymorphisms and copy numbers in Plasmodium falciparum, Mozambique, 2015. Emerg Infect Dis. 2018;24:40–8.
Huang B, Deng C, Yang T, Xue L, Wang Q, Huang S, et al. Polymorphisms of the artemisinin resistant marker (K13) in Plasmodium falciparum parasite populations of Grande Comore Island 10 years after artemisinin combination therapy. Parasit Vectors. 2015;8:634.
Li J, Chen J, Xie D, Eyi UM, Matesa RA, Ondo Obono MM, et al. Limited artemisinin resistance-associated polymorphisms in Plasmodium falciparum K13-propeller and PfATPase6 gene isolated from Bioko Island, Equatorial Guinea. Int J Parasitol Drugs Drug Resist. 2016;6:54–9.
Muwanguzi J, Henriques G, Sawa P, Bousema T, Sutherland CJ, Beshir KB. Lack of K13 mutations in Plasmodium falciparum persisting after artemisinin combination therapy treatment of Kenyan children. Malar J. 2016;15:36.
Mvumbi DM, Bobanga TL, Kayembe JN, Mvumbi GL, Situakibanza HN, Benoit-Vical F, et al. Molecular surveillance of Plasmodium falciparum resistance to artemisinin-based combination therapies in the Democratic Republic of Congo. PLoS ONE. 2017;12:e0179142.
Ocan M, Bwanga F, Okeng A, Katabazi F, Kigozi E, Kyobe S, et al. Prevalence of K13-propeller gene polymorphisms among Plasmodium falciparum parasites isolated from adult symptomatic patients in northern Uganda. BMC Infect Dis. 2016;16:428.
Ouattara A, Kone A, Adams M, Fofana B, Maiga AW, Hampton S, et al. Polymorphisms in the K13-propeller gene in artemisinin-susceptible Plasmodium falciparum parasites from Bougoula-Hameau and Bandiagara, Mali. Am J Trop Med Hyg. 2015;92:1202–6.
Authors’ contributions
MO drafted the initial concept of the study. RS and AAK conducted database article search. MO, EAO, DA screened articles for inclusion in the review. MO, SN and KM developed data analysis plan and MO performed analysis of data regarding K13-mutations reported in different malaria affected regions. All authors read and approved final manuscript.
Acknowledgements
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
All data generated or analyzed during this study are included in this published article and its additional files.
Consent for publication
Not applicable.
Ethics approval and consent to participate
Not applicable.
Funding
The review has received funding from a Grant, Number D43TW010132, supported by Office of the Director, National Institutes of Health (OD), National Institute of Dental & Craniofacial Research (NIDCR), National Institute of Neurological Disorders and Stroke (NINDS), National Heart, Lung, and Blood Institute (NHLBI), Fogarty International Center (FIC), and National Institute on Minority Health and Health Disparities (NIMHD). Additional funding was obtained from Malaria training Grant Number D43TW010526. The funders have had no role in the design of the study and in writing of the review protocol.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Ocan, M., Akena, D., Nsobya, S. et al. K13-propeller gene polymorphisms in Plasmodium falciparum parasite population in malaria affected countries: a systematic review of prevalence and risk factors. Malar J 18, 60 (2019). https://doi.org/10.1186/s12936-019-2701-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12936-019-2701-6