Abstract
Currently, there is no effective treatment for neurological complications of infection with the human immunodeficiency virus that persists despite the use of combination antiretroviral therapy. A medium throughput assay was developed for screening neuroprotective compounds using primary mixed neuronal cells and mitochondrial toxin 3-nitropropionic acid. Using this assay, a library of 2,000 compounds was screened. Out of 256 compounds that showed variable degrees of neuroprotection, nine were related to epicatechin, a monomeric flavonoid found in cocoa and green tea leaves that readily crosses the blood–brain barrier. Hence, catechin, epicatechin, and the related compound, epigallocatechin gallate (EGCG) were further screened for their neuroprotective properties against HIV proteins Tat and gp120, and compared to those of resveratrol. Epicatechin and EGCG targets the brain-derived neurotrophic factor (BDNF) and its precursor proBDNF signaling pathways, normalizing both Tat-mediated increases in proapoptotic proBDNF and concomitant Tat-mediated decreases in the mature BDNF protein in hippocampal neurons. Epicatechin and epigallocatechin gallate were more potent than catechin or resveratrol as neuroprotectants. Due to its simpler structure and more efficient blood–brain barrier penetration properties, epicatechin might be the best therapeutic candidate for neurodegenerative diseases including HIV-associated neurocognitive disorders where oxidative stress is an important pathophysiological mechanism.
Similar content being viewed by others


Introduction
Infection with the human immunodeficiency virus (HIV) can result in severe complications affecting the nervous system, such as HIV sensory distal neuropathy (HIV-SN) and HIV-associated neurocognitive disorders (HAND). These complications occur and persist despite the advent and implementation of combined active antiretroviral therapy (CART) (McArthur et al. 2010). As effective CART allows HIV-infected patients to live longer, a growing proportion of patients experience some form of neuropathy and cognitive deficits. These complications are a result of oxidative stress-induced neurotoxicity that results directly from HIV infection in tissue reservoirs, where effective levels of antiretroviral drugs may be hard to achieve and low level HIV replication may continue. The coat protein of HIV, gp120, can cause neurotoxicity by multiple mechanisms. Exposure of neurons and glial cells to toxic viral proteins such as Tat, which can be produced by HIV-infected cells despite the use of CART, can set up positive feedback loops resulting in widespread neuronal dysfunction. Oxidative stress is the final common pathway for the neurotoxicity induced by both viral proteins (Mattson et al. 2005). Development of therapeutic strategies to protect against this neuronal damage and the ensuing clinical deficits remains a major unmet medical need. Neuroprotective compounds, if they can be identified and developed preclinically, may provide a treatment for HIV-SN and HAND.
One approach to identify potential neuroprotective agents would be to develop a medium throughput screening assay to assess the protective efficacy of a wide array of diverse chemical compounds. We have developed assays that utilize mixed primary cultures of neurons and glial cells to replicate the cellular conditions in situ. These cultures have been adapted to microtiter plates and exposed to oxidative stressors, such as 3-nitropropionic acid (3-NP), to mimic oxidative damage which results from HIV infection of these tissues. Confirmatory assays are then performed using HIV toxic proteins gp120 and Tat. Thus, we screened a library of 2,000 compounds comprised of natural substances, FDA-approved drugs, and compounds with prior human exposure (Spectrum Collection, Microsource Discovery). We identified epicatechin, epigallocatechin gallate (EGCG) and other catechin flavonoids as potential neuroprotective agents. These compounds are found in high concentrations in green tea leaves, purple grapes (Dani et al. 2009), and in seeds of the cacao tree which is used to make chocolate. While these compounds have reported antioxidant activities, these compounds have not been evaluated against mitochondrial toxins or HIV toxic proteins.
Materials and methods
Cell culture
Neuronal cultures from rodent cerebral cortex and from hippocampus were prepared from embryonic day 18 Sprague–Dawley rats using methods similar to those described previously (Haughey et al. 2004). Tissues were dissociated by gentle trituration with a firepolished glass pipette in calcium-free Hank’s balanced salt solution. The single cell suspension was centrifuged at 1,000×g and re-suspended in minimal essential medium containing 10 % heat-inactivated fetal bovine serum and 1 % (v/v) antibiotic and antimycotic solution (penicillin G 104 U/mL, streptomycin 10 mg/mL, and amphotericin B 25 mg/mL; Sigma, St Louis, MO, USA). Cells were allowed to attach for 3 h before the media was replaced with serum-free neurobasal medium containing 2 % (v/v) B-27 supplement (Gibco, Rockville, MD, USA) and 1 % (v/v) antibiotic and antimycotic mix (Sigma). Rat mixed hippocampal neurons were generated from freshly cultured rat hippocampi in neurobasal media containing 5 % (v/v) fetal bovine serum and 2 % (v/v) B27 supplement. Hippocampal neurons were plated in 96-well plates at a density of 4 × 105 cells/mL for neurotoxicity studies.
Human neuronal cultures were prepared as described previously (Magnuson et al. 1995). Briefly, the cells were mechanically dissociated, suspended in Opti-MEM with 5 % (v/v) heat-inactivated fetal bovine serum, 0.2 % (v/v) N2 supplement (Gibco), and 1 % (v/v) antibiotic and antimycotic solution and plated in flat-bottomed 96-well plates. The cells were maintained in culture for at least 1 month before use. Mixed neuronal cultures were grown in low serum conditions with N-2 supplement to encourage neuronal growth and inhibit glial proliferation. These cultures contain 70 % neurons, 30 % astrocytes, and <1 % microglia. These cultures can be maintained for several months. Previous experiments in our laboratory and those of others have shown that prolonged cultures of neurons ensures the expression of excitatory amino acid receptors and growth of processes (Magnuson et al. 1995; Nath et al. 1996). The original screening assays were done with three to four replicates per plate, including an internal positive control of 10 μM resveratrol as a neuroprotectant.
Neurons were plated on polyethyleneimine-coated 15-mm diameter glass coverslips at a density of 2 × 105 cells/mL for calcium imaging and in 35-mm diameter plastic plates at a density of 4 × 105 cells/mL for neurotoxicity studies.
Cell survival assay
Neuronal cell viability was assessed with 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. This assay (Mosmann 1983) is based on the ability of a mitochondrial dehydrogenase enzyme from viable cells to cleave the tetrazolium rings of the pale yellow MTT and form dark blue formazan crystals. These crystals are largely impermeable to cell membranes, and thus, accumulate within healthy cells. The resultant formazan precipitates are solubilized with dimethyl sulfoxide (DMSO) and read on a multiwell scanning spectrophotometer (SpectraMAX M5e, Molecular Devices). The number of surviving cells is directly proportional to the level of the formazan product created.
Compound screening
The Spectrum Collection of compounds by Microsource Discovery contains 2,000 compounds, of which about half are FDA-approved drugs and the remaining compounds are natural products or other compounds with some prior human exposure and safety testing data. The compound collection is dispensed and maintained on 96-well plates at a concentration of 10 mM in 100 % DMSO and stored at −80 °C. The compound mother plates were thawed one time in order to make four sets of daughter plates, and one set of daughter plates was used for these screening assays.
Toxicity assays
Rat mixed hippocampal cultures were treated with test compounds for 1 h at 37 °C prior to addition of 3-NP, Tat, or gp120. Cells were then exposed to these toxins for 18 h, after which time cell viability was assessed by MTT assay (Mosmann 1983). For Tat toxicity assays, purified recombinant Tat1-72 was utilized at 500 nM final concentration. After 18 h of exposure to Tat, the MTT assay was utilized to assess cytotoxicity. Hydrogen peroxide neurotoxicity was achieved with 2 h exposure to 100 μM H2O2, which is applied to the cultures following a 1-h preincubation with the neuroprotectant. Cell viability was quantified by MTT assay. Moderate throughput functional screening assays were done in triplicate or quadruplicate wells, while confirmatory dose–response assays were done with N = 8 per treatment group and replicated three times.
Fluorescence axodendritic degeneration assays
Rat mixed hippocampal or cortical neurons were transfected with a plasmid containing the neuron-specific CaMKII promoter, with a fusion construct of tau protein and td-Tomato genes or βIII tubulin–tdTomato genes (Nucleofector 2S, Lonza, MD), according to the manufacturer’s instructions. Transfection efficiency was about 60 %. After 7–10 days in culture, cells were utilized in neurotoxicity assays to assess the effects of catechins on Tat-mediated axodendritic degeneration. Following catechin treatment and Tat exposure, images were captured via live imaging on a Zeiss AxioObserver Z1 inverted microscope and the relative fluorescence of Tau-tdTomato was determined with a SpectraMax M5e multimode plate reader (Molecular Devices, CA).
HIV-1 proteins and reagents
The preparation of recombinant Tat1-72 protein has been described previously (Ma and Nath 1997). Tat protein varies in length from 86 to 101 amino acids and is made from two exons. The first exon form the first 72 amino acids and the neurotoxic epitope resides in this region (Nath et al. 1996), hence we have utilized recombinant Tat1-72 for these studies. The toxic effects of Tat1-72 can be blocked by anti-Tat Ig and likewise Tat∆31-61 has been used as a control (Turchan et al. 2001). Gp120 derived from HIVSF was acquired from the NIH-AIDS repository (Bethesda, MD, USA).
Statistical analysis
All data are represented as mean ± SEM and analyzed by one-way analysis of variance (ANOVA). Group-wise post hoc comparisons were assessed by Newman–Keuls multiple comparison tests, with statistical significance at p < 0.05.
Results
Development of a neurotoxicity assay for screening neuroprotectants
To develop a reliable and reproducible neurotoxicity assay, we used primary neuronal cultures derived from rat hippocampi. We kept the species and age of the animal, the anatomical region from which neurons were obtained, and the culture conditions including days in culture constant for all experiments. 3-nitropropionic acid was chosen as a toxin since it is stable and is a well-characterized mitochondrial toxin that causes oxidative stress. We found that the compound also has a large dynamic range and its toxic properties in neurons were highly reproducible. For these reasons, we were able to adapt the neurotoxicity assay to microtiter plates, which allowed the manual screening of a large number of compounds. Based on the dose response, we chose 3 mM 3-NP for all subsequent experiments since it provided 20–35 % toxicity (Fig. 1). Our intra-assay variability was 8–10 %.

Titration of 3-NP toxicity in rat mixed hippocampal neurons and assay validation. Rat hippocampal cultures were exposed to various concentrations of 3-nitropropionic acid (3-NP) for 18 h and cell viability was determined with an MTT endpoint (see Materials and methods for assay details). An average toxicity of between 20 and 35 % was observed with 3 mM 3-NP exposure; hence this concentration was used for subsequent assays
Neuroprotection by the catechin class of compounds
The spectrum library of compounds contains 2,000 FDA-approved drugs and natural substances. We screened the entire library at a single concentration of 10 μM of each compound. We chose 10 μM as a reasonable in vitro screening dose because it may be a maximal level attained in the CNS following systemic administration. We observed that 256 compounds showed >50 % protection. Of these, 146 showed >100 % protection and 53 showed >200 % protection (see Fig. 2). The library contained 13 compounds that were derived from or related to epicatechin. Of these, one was toxic, three had no effect, and the remaining nine showed variable amounts of neuroprotection (see Table 1). Of these compounds we chose epicatechin, (+) catechin, and epigallocatechin gallate for further experiments since they showed the most neuroprotection and had the simplest structure. Their structure also shows similarity to that of resveratrol, in that all of them have aromatic phenolic substituents. They have the basic structure of two phenolic rings linked by three carbons that form an oxygenated heterocycle (Fig. 3). The catechins may exist in cis ((−)epicatechin; Fig. 3a) or trans ((+)catechin; Fig. 3b) configuration. The subfamilies of catechins (Table 1) are formed by linkage of various groups to the heterocycle, such as gallic acid (Fig. 3d). Resveratrol is a known neuroprotective agent (Richard et al. 2011); hence, it was used as a positive control in subsequent experiments. In addition to its use as a positive control for our experiments, resveratrol was also part of the spectrum library of compounds and showed about 40 % neuroprotection in our screening assay.

Screening of the Spectrum collection with the 3-NP neurotoxicity assay: a total of 2,000 compounds were screened using the 3-NP neurotoxicity assay in rat mixed hippocampal cultures, with screening at 10 μM drug concentration against the toxic effects of 3 mM 3-NP. From these screening assays, 256 compounds showed >50 % protection. Additionally, of these “hits”, 146 compounds displayed complete (100 % protection) and 53 demonstrated >200 % protection

Chemical structures of catechins compounds used in these studies. a (−) Epicatechin (molecular mass 290) which is the cis form of b (+)-catechin. c Resveratrol (molecular mass 228) and d epigallocatechin gallate (molecular mass 458)
Dose–response curves of neuroprotection by epicatechin and catechin
Using the 3-NP neurotoxicity assay, we found that epicatechin and (+) catechin protected in a dose-responsive manner. Each of these compounds had a linear dose response. They showed nearly complete neuroprotection at 20 μM concentration. Resveratrol was used as a positive control which showed similar neuroprotective properties, and required about 20 μM concentrations to demonstrate greater than 50 % protection. It is interesting that the EGCG compound was more active than the other three, with near complete protection at about 10 μM (Fig. 4).

Catechins protect against 3-NP-mediated oxidative stress in hippocampal cultures. a Epicatechin and (+) catechin (5–20 μM) were preincubated with rat mixed hippocampal cultures for 1 h, followed by 18 h exposure to 3-NP. Neuronal cell viability was assessed by MTT endpoint. The data from eight replicates were utilized for each treatment group, and were evaluated by ANOVA for significance. Group-wise post hoc comparisons were assessed by Newman–Keuls multiple comparison test, with statistical significance at p < 0.05 versus 3-NP. Epicatechin was more effective than catechin at all concentrations. Full dose response with b epicatechin, c EGCG, and d resveratrol demonstrate concentration-dependent neuroprotective effects of these compounds
Epicatechin and catechin protect against HIV proteins
To determine if epicatechin can protect against the neurotoxic effect of HIV proteins, we exposed neurons to HIV proteins Tat and gp120 in the presence of epicatechin. We found that epicatechin protected in a concentration-dependent manner against Tat and gp120 with significant effects at a dosage of 10 μM or above (Fig. 5a and b). In comparison, resveratrol showed protection against Tat protein at a dosage of 2.5 μM or above (Fig. 5c). EGCG was the most potent catechin compound, which exhibited nearly complete protection from Tat neurotoxicity at concentrations of 2 μM or greater (Fig. 5c).

Protection by catechins and resveratrol against HIV-protein-induced neurotoxicity. Using rat hippocampal cultures, we assessed the neuroprotective efficacy of epicatechin against HIV proteins, a Tat and b gp120; and the efficacy of c EGCG as well as d resveratrol against the neurotoxicity elicited by HIV Tat. Various concentrations of these compounds were pre-incubated with hippocampal cultures for 1 h, followed by 18 h exposure to HIV Tat1-72 or gp120. Neuronal cell viability was quantified by MTT endpoint. The data from eight replicates were utilized for each treatment group, and were evaluated by ANOVA for significance. Group-wise post hoc comparisons were assessed by Newman–Keuls multiple comparison test. *p < 0.05, **p < 0.01, ***p < 0.005 versus Tat or gp120. With each of these compounds, nearly complete protection is seen with concentrations of ≥10 μM. However, of all the catechin compounds, EGCG showed the most potent neuroprotection
Neuroprotective properties against hydrogen-peroxide-induced toxicity
Minimal protection was seen with each of the compounds against H2O2-induced neurotoxicity with significant protection only at 20 μM concentration for epicatechin, catechin, and resveratrol. EGCG was protective at lower concentrations (Fig. 6). We also discovered that the neuroprotective properties of epicatechin were light sensitive, and that exposure to ambient light at atmospheric oxygen for more than 1–2 h resulted in loss of its neuroprotective properties (data not shown). Hence all assays were performed with freshly prepared solutions of these catechins compounds and additional precautions were taken using light barriers. Collectively, it suggests that likely these compounds are easily oxidized resulting in loss of activity.

Catechins protect against hydrogen peroxide-mediated neurotoxicity. a Epicatechin, b (+) catechin, c EGCG, and d resveratrol were preincubated with hippocampal neurons for 1 h prior to 2 h exposure to 100 μM hydrogen peroxide (H2O2). Cell viability was quantitated by MTT analysis. The data from eight replicates were utilized for each treatment group, and were evaluated by ANOVA for significance. Group-wise post hoc comparisons were assessed by Newman–Keuls multiple comparison test. *p < 0.05; **p < 0.01 versus hydrogen peroxide. Minimal protection is seen with each of these compounds
Protection from Tat-mediated neuritic damage by catechins
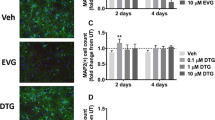
In order to determine whether catechins can protect against the neurotoxic effect of HIV proteins on neuronal processes as well as cell bodies, we exposed fluorescently labeled neurons to HIV proteins Tat in the presence of epicatechin or EGCG (Fig. 7). We found that epicatechin protected in a concentration-dependent manner against Tat with significant effects at a dosage about 10 μM (Fig. 7a). In comparison, EGCG was the more potent catechin compound, which exhibited nearly complete protection from Tat neurotoxicity at concentrations of 1 μM or greater (Fig. 7b). The exposure of cortical neurons to HIV Tat resulted in fewer labeled neurons per field, as well as a simplified neuronal axodendritic profile (Fig. 7c). Similarly, mixed hippocampal neurons labeled by transfection with βIII tubulin–td Tomato displayed significant losses of neuritic processes following exposure to Tat (Fig. 8). The catechins protected the neurites from Tat-mediated damage at 1–10 μM. The effect of epicatechin and EGCG treatment was observed to be protection of neurons, as well as the neuritic processes from the neurotoxic effects of Tat.

Protection by catechins against HIV-Tat-induced neurotoxicity. Using rat cortical cultures which were expressing the Tau-tdTomato gene under control of the CamKII promoter, we assessed the neuroprotective efficacy of epicatechin (left panel, a) and EGCG (right panel, b) against the neurotoxicity elicited by HIV Tat. Various concentrations of these compounds were pre-incubated with cortical cultures for 1 h, followed by 24 h exposure to 500 nM HIV Tat1-72. Images were acquired by live imaging in an unbiased manner from a Zeiss AxioObserver inverted microscope. Fluorescence intensity was measured with a SpectraMax M5e plate reader. The data from six replicates were utilized for each treatment group, and were evaluated by ANOVA for significance. Group-wise post hoc comparisons were assessed by Newman–Keuls multiple comparison test. *p < 0.05, **p < 0.01, ***p < 0.005 versus 500 nM Tat. Nearly complete protection is seen with each compound, with epicatechin requiring concentrations of 10 μM, while EGCG at 1 μM was maximally protective. Representative fluorescence images of these treatments are depicted in (c), where media (top row, a), 500 nM Tat (top row, b), epicatechin + Tat (middle row, 0.1 μM (c), 1 μM (d), and 10 μM (e)), and EGCG + Tat (bottom row, 0.1 μM (f), 1 μM (g), and 10 μM (h)). Scale bar = 100 μm

Protection by catechins against HIV-Tat-induced neurotoxicity. Using rat hippocampal cultures which were expressing the βIII Tubulin–tdTomato gene under control of the CamKII promoter, we assessed the neuroprotective efficacy of epicatechin and EGCG against the neurotoxicity elicited by HIV Tat. Various concentrations (0.1, 1, or 10 μM) of these compounds were pre-incubated with rat mixed hippocampal cultures for 1 h, followed by 24 h exposure to 500 nM HIV Tat1-72. Images were acquired by live imaging in an unbiased manner from a Zeiss AxioObserver inverted microscope. The data from six replicates were utilized for each treatment group. Nearly complete protection is seen with each compound at 1–10 μM, with epicatechin requiring concentrations of 10 μM, while EGCG at 1 μM was maximally protective. Representative fluorescence images of these treatments are depicted, where media (top row, a), 500 nM Tat (top row, b), epicatechin + Tat (middle row, 0.1 μM (c), 1 μM (d), and 10 μM (e)), and EGCG + Tat (bottom row, 0.1 μM (f), 1 μM (g), and 10 μM (h)). Scale bar = 50 μm
Potential neuroprotective mechanism(s) of catechin-mediated protection from HIV
In order to gain some insight into the neuroprotective mechanism of the catechins, neuronal cultures were treated with vehicle, Tat, Tat plus epicatechin (1 μM), or Tat plus EGCG (1 μM) for 24 h (Fig. 9). Cell lysates of the treated neurons were probed with antibodies to numerous proteins, such as inducible nitric oxide synthase (iNOS), IL-6 as well as potential neurotrophic proteins, such as NGF, BDNF, and GDNF. HIV Tat treatment resulted in nearly a twofold increases in iNOS and IL-6, but only the Tat-mediated increase in iNOS was attenuated by the catechins (Fig. 9). Tat exposure resulted in a decrease in the neurotrophic BDNF, but concomitantly stimulated an increase in the pro-apoptotic proBDNF. The catechins, utilized at neuroprotective 1 μM concentration, increased neurotrophic BDNF levels to nearly 50 % above control media/DMSO-vehicle-treated levels, while decreasing the proapoptotic proBDNF to more than 50 % less than control levels. The effects of catechins on the BDNF proteins were specific, since no effect of these compounds was evident on NGF or GDNF proteins (data not shown). These data suggest that the neuroprotective effects of catechins may result by modulating the BDNF proteins and their signaling pathways.

Tat-mediated expression of signaling proteins in mixed rat neuronal cultures. Rat mixed cortical cultures (106 cells per well on six-well plates) were treated with culture media plus 0.1 % DMSO vehicle, 500 nM Tat, or Tat plus 1 μM epicatechin or 1 μM EGCG for 24 h. After 24 h, cell lysates were generated, proteins separated by SDS–polyacrylamide gel electrophoresis, transferred to PVDF membranes and probed with antibodies to iNOS and IL-6 (left panel) and BDNF (right panel). The BDNF antibody recognized the precursor of BDNF (proBDNF) at 32 kDa as well as the mature BDNF protein at 13 kDa. Gel loading was normalized by BCA protein determinations, and blots were normalized by GAPDH immunoreactivity. The iNOS antibody recognized both neuronal NOS, whose expression remained constant with Tat treatment, along with the slightly smaller iNOS protein. Protein expression levels relative to control media (+0.1 % DMSO vehicle) treatment were made by densitometry with NIH ImageJ software. The data from three independent replicates were utilized for each treatment group, and were evaluated by ANOVA for significance. Group-wise post hoc comparisons were assessed by Newman–Keuls multiple comparison test. *p < 0.05, **p < 0.01, ***p < 0.005 versus 500 nM Tat. i-NOS from BD Biosciences, San Jose, CA; IL6 from ABCAM, Cambridge, MA; GAPDH from Sigma-Aldrich, St. Louis, MO; BDNF – Santa Cruz Biotechnology, Santa Cruz, CA
Discussion
High-throughput assays have been the mainstay of pharmaceutical companies for drug discoveries. Recently, the National Institutes of Health and other academic institutions have put in major effort towards the development of such assays for drug discovery that would particularly target rare disorders or other diseases that pharmaceutical companies have traditionally ignored. The neurological disorders associated with HIV infection fall into this category. Most throughput assays are either enzyme based or binding assays. However, cellular functions are very complex and involve multiple pathways, hence we developed a moderate throughput assay using primary mixed neuronal cultures and used rigorous experimental conditions to minimize variability and maximize reproducibility. We used 3-NP for causing neurotoxicity since its acts on the mitochondria and mitochondrial dysfunction have been implicated in most neurodegenerative diseases. Further, 3-NP is a stable compound with a wide and reproducible dose curve. Using this assay, we have identified several groups of compounds that have variable neuroprotective properties. We focused our initial efforts on catechins since nine related compounds had neuroprotective properties and have been shown to cross the blood–brain barrier (Faria et al. 2011). Further these compounds are natural substances present in green tea and other plants although the gallate group is predominant in green tea (Henning et al. 2003; Fraga and Oteiza 2011) Dark chocolate from the plant Theobroma cocao is also a rich source of epicatechin (Engler et al. 2004). Hence, humans have been regularly exposed to these compounds in food products for hundreds or thousands of years.
Several studies have investigated the neuroprotective properties of epigallocatechin gallate (reviewed in (Weinreb et al. 2009; Mandel et al. 2008)), the main catechin polyphenol constituent in green tea, and found that it protects neurons against 3-NP (Kumar and Kumar 2009) amyloid beta peptide (He et al. 2011), traumatic injury (Wei et al. 2011), cerebral ischemia (Park et al. 2009), cisplatin-induced neurotoxicity (Lee et al. 2010), and rotenone-induced neurotoxicity (Moldzio et al. 2010). However, one study failed to show any neuroprotection against 6-hydroxydopamine (Leaver et al. 2009). Although the exact mechanism of action of EGCG remains unknown, prevention of generation of reactive oxygen species, and nitric oxide synthesis has been implicated (Kim et al. 2009). These compounds have also been shown to inhibit ERK and NF-κB pathways (Fraga and Oteiza 2011; Lee et al. 2010). It can also cause proliferation of neural progenitor cells (Yoo et al. 2010). Our data in this study implicates the BDNF pathway in the neuroprotective actions of the catechins. The proapoptotic proBDNF (Teng et al. 2005) induces neuronal apoptosis via activation of a receptor complex of p75NTR and sortilin. proBDNF is converted to the mature and neuroprotective BDNF by extracellular proteases (Lu et al. 2005). Perhaps exposure of neurons to Tat caused increases in proBDNF (along with decreases in BDNF) by blocking the proteolytic processing of the proBDNF, which was reversible by catechin treatment. Interestingly, gp120 treatment also caused decreased BDNF levels in vitro and in vivo after 24 h of exposure (Nosheny et al. 2004), preceding caspase-3 activation and neuronal cell death. It is possible that catechin-mediated protection from gp120 (Fig. 5) may also utilize this mechanism. Recent studies in the CSF of HIV-infected patients demonstrate a strong correlation with increasing severity of HIV-mediated neurologic disease and a decline in growth factors, particularly BDNF and NT-3 (Meeker et al. 2011). Thus, a potential mechanism of catechin-mediated neuroprotection which promoted BDNF availability and signaling may be an effective strategy against HIV-mediated neurotoxicity and HAND. While the potential role of BDNF as a neurotherapeutic agent has long been considered (Rothman et al. 2012), its use has been limited due to the need to directly administer it to the brain as a recombinant protein or via viral vectors (Kells et al. 2008). Catechins thus represent a novel set of compounds by their ability to act via the BDNF pathway and may thus open a new area in neurotheraputics research.
Importantly, epigallocatechin has been shown to be neuroprotective following oral administration in an animal model of acute neurotoxicity (Kang et al. 2010) and aging-related oxidative injury in the brain (Li et al. 2010). Other compounds related to the catechins have not been well studied. We found that epigallocatechin gallate was only moderately neuroprotective against 3-NP neurotoxicity. Catechin, epicatechin, and epicatechin monogallate were much more potent than epigallocatechin in our screening assay. Of these compounds epicatechin had the best potency against 3-NP-induced neurotoxicity although they seemed to be similarly protective against HIV proteins with apparent EC50 of 1 μM compared to an EC50 of 10 μM for 3-NP neurotoxicity. Epicatechin gallate has been shown to have cytoprotective effects in a glial cell line at much lower concentrations of 0.1 to 1 μM (Abib et al. 2010). However, catechin and epicatechin have a smaller molecular mass and are efficient in crossing the blood–brain barrier (Faria et al. 2011). Catechin has also been shown to improve blood flow by causing cerebral vasodilatation (Drouin et al. 2011). The bioavailability of the compounds improves with repeated dosing. In one study, catechin and epicatechin were undetectable in the brain of rodents after administration of a single dose; however, after daily administration for 10 days, it reached 576.7 ± 227.7 and 290.7 ± 45.9 pg/g of brain tissue, respectively (Ferruzzi et al. 2009). Defining the therapeutic window of these compounds will be critical since at very high dosages, they may have proapoptotic properties and can cause neurotoxicity (Yin et al. 2009). Our observations also suggested a similar trend. We found that resveratrol was also neuroprotective against HIV-Tat protein-induced neurotoxicity; however, due to its hydrophilic properties, it does not cross the blood–brain barrier as efficiently.
Oxidative and nitrosative stress play an important role in mediating the neuropathogenesis of HIV infection and protein carbonyls, nitrosylated and hydroxynonenol-modified proteins can be found in brain and CSF of these patients, where they correlate with the severity of neurocognitive impairment (Turchan et al. 2003; Li et al. 2008). HIV proteins Tat and gp120 have been implicated in causing excitotoxic neurotoxicity by directly acting on neurons or via indirect mechanisms leading to activation of macrophages and glial cells. Oxidative stress is the final common pathway in all these mechanisms (Mattson et al. 2005). Currently available antiretroviral medications are unable to control complete viral replication in the brain and they do not have any effect of Tat production once proviral DNA has been formed. In this study, we found that the catechins were able to block the neurotoxic effects of the HIV proteins and thus may be of therapeutic potential in HAND. However, conducting clinical trials in this patient population carries many challenges and to date all trials with potential neurotherapeutic agents have failed in phase 2 studies. The availability of good surrogate markers is critical for monitoring such clinical trials so that early decisions on the continuation of these trials can be made. Thus, measurement of oxidized or nitrosylated proteins in CSF represents an excellent marker for monitoring clinical trials (Schifitto et al. 2009). It is important to note that even though catechins have been shown to be protective in a variety of models of neurodegenerative diseases, to date no clinical trials have been conducted with these compounds. This is in part because monitoring clinical end points in these diseases requires large sample sizes and monitoring of patients for long periods of time. CSF is not routinely obtained from these patients for either diagnostic purposes or for monitoring clinical progression. In patients with HAND, CSF monitoring of viral load and inflammatory markers is common practice, hence this population might represent an excellent opportunity for monitoring the therapeutic efficacy of these compounds.
In conclusion, we have developed a novel screening assay for identification of neuroprotective compounds and identified the family of catechins as a class of compounds with potent neuroprotective properties. Of these compounds, epicatechin is the most promising therapeutic candidate because of its high potency, simple structure, excellent bioavailability following oral administration, and efficient ability to cross the blood–brain barrier.
References
Abib RT et al (2010) Genoprotective effects of the green tea-derived polyphenol/epicatechin gallate in C6 astroglial cells. J Med Food 13(5):1111–1115
Dani C et al (2009) Antioxidant activity and phenolic and mineral content of rose grape juice. J Med Food 12(1):188–192
Drouin A et al (2011) Catechin treatment improves cerebrovascular flow-mediated dilation and learning abilities in atherosclerotic mice. Am J Physiol Heart Circ Physiol 300(3):H1032–H1043
Engler MB et al (2004) Flavonoid-rich dark chocolate improves endothelial function and increases plasma epicatechin concentrations in healthy adults. J Am Coll Nutr 23(3):197–204
Faria A et al (2011) Insights into the putative catechin and epicatechin transport across blood–brain barrier. Food Funct 2(1):39–44
Ferruzzi MG et al (2009) Bioavailability of gallic acid and catechins from grape seed polyphenol extract is improved by repeated dosing in rats: implications for treatment in Alzheimer’s disease. J Alzheimers Dis 18(1):113–124
Fraga CG, Oteiza PI (2011) Dietary flavonoids: role of (−)-epicatechin and related procyanidins in cell signaling. Free Radic Biol Med 51(4):813–823
Haughey NJ et al (2004) Perturbation of sphingolipid metabolism and ceramide production in HIV-dementia. Ann Neurol 55(2):257–267
He Y et al (2011) Prolonged exposure of cortical neurons to oligomeric amyloid-beta impairs NMDA receptor function via NADPH oxidase-mediated ROS production: protective effect of green tea (−)-epigallocatechin-3-gallate. ASN Neuro 3(1):e00050
Henning SM et al (2003) Catechin content of 18 teas and a green tea extract supplement correlates with the antioxidant capacity. Nutr Cancer 45(2):226–235
Kang KS et al (2010) Dual beneficial effects of (−)-epigallocatechin-3-gallate on levodopa methylation and hippocampal neurodegeneration: in vitro and in vivo studies. PLoS One 5(8):e11951
Kells AP, Henry RA, Connor B (2008) AAV-BDNF mediated attenuation of quinolinic acid-induced neuropathology and motor function impairment. Gene Ther 15(13):966–977
Kim CY et al (2009) Neuroprotective effect of epigallocatechin-3-gallate against beta-amyloid-induced oxidative and nitrosative cell death via augmentation of antioxidant defense capacity. Arch Pharm Res 32(6):869–881
Kumar P, Kumar A (2009) Protective effects of epigallocatechin gallate following 3-nitropropionic acid-induced brain damage: possible nitric oxide mechanisms. Psychopharmacology (Berl) 207(2):257–270
Leaver KR et al (2009) Oral pre-treatment with epigallocatechin gallate in 6-OHDA lesioned rats produces subtle symptomatic relief but not neuroprotection. Brain Res Bull 80(6):397–402
Lee JS et al (2010) Epicatechin protects the auditory organ by attenuating cisplatin-induced ototoxicity through inhibition of ERK. Toxicol Lett 199(3):308–316
Li W et al (2008) Nitrosative stress with HIV dementia causes decreased L-prostaglandin D synthase activity. Neurology 70(19 Pt 2):1753–1762
Li Q et al (2010) Chronic green tea catechins administration prevents oxidative stress-related brain aging in C57BL/6 J mice. Brain Res 1353:28–35
Lu B, Pang PT, Woo NH (2005) The yin and yang of neurotrophin action. Nat Rev Neurosci 6(8):603–614
Ma M, Nath A (1997) Molecular determinants for cellular uptake of Tat protein of human immunodeficiency virus type 1 in brain cells. J Virol 71(3):2495–2499
Magnuson DS et al (1995) Human immunodeficiency virus type 1 tat activates non-N-methyl-d-aspartate excitatory amino acid receptors and causes neurotoxicity. Ann Neurol 37(3):373–380
Mandel SA et al (2008) Simultaneous manipulation of multiple brain targets by green tea catechins: a potential neuroprotective strategy for Alzheimer and Parkinson diseases. CNS Neurosci Ther 14(4):352–365
Mattson MP, Haughey NJ, Nath A (2005) Cell death in HIV dementia. Cell Death Differ 12(Suppl 1):893–904
McArthur JC et al (2010) Human immunodeficiency virus-associated neurocognitive disorders: mind the gap. Ann Neurol 67(6):699–714
Meeker RB et al (2011) Protein changes in CSF of HIV-infected patients: evidence for loss of neuroprotection. J Neurovirol 17(3):258–273
Moldzio R et al (2010) Effects of epigallocatechin gallate on rotenone-injured murine brain cultures. J Neural Transm 117(1):5–12
Mosmann T (1983) Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays. J Immunol Methods 65(1–2):55–63
Nath A et al (1996) Identification of a human immunodeficiency virus type 1 Tat epitope that is neuroexcitatory and neurotoxic. J Virol 70(3):1475–1480
Nosheny RL et al (2004) Human immunodeficiency virus type 1 glycoprotein gp120 reduces the levels of brain-derived neurotrophic factor in vivo: potential implication for neuronal cell death. Eur J Neurosci 20(11):2857–2864
Park JW et al (2009) Green tea polyphenol (−)-epigallocatechin gallate reduces neuronal cell damage and up-regulation of MMP-9 activity in hippocampal CA1 and CA2 areas following transient global cerebral ischemia. J Neurosci Res 87(2):567–575
Richard T et al (2011) Neuroprotective properties of resveratrol and derivatives. Ann N Y Acad Sci 1215:103–108
Rothman SM, Griffioen KJ, Wan R, Mattson MP (2012) Brain-derived neurotrophic factor as a regulator of systemic and brain energy metabolism and cardiovascular health. Ann N Y Acad Sci. doi:10.1111/j.1749-6632.2012.06525.x
Schifitto G et al (2009) Selegiline and oxidative stress in HIV-associated cognitive impairment. Neurology 73(23):1975–1981
Teng HK et al (2005) ProBDNF induces neuronal apoptosis via activation of a receptor complex of p75NTR and sortilin. J Neurosci 25(22):5455–5463
Turchan J et al (2001) Estrogen protects against the synergistic toxicity by HIV proteins, methamphetamine and cocaine. BMC Neurosci 2:3
Turchan J et al (2003) Oxidative stress in HIV demented patients and protection ex vivo with novel antioxidants. Neurology 60(2):307–314
Wei IH et al (2011) (−)-Epigallocatechin gallate attenuates NADPH-d/nNOS expression in motor neurons of rats following peripheral nerve injury. BMC Neurosci 12:52
Weinreb O et al (2009) Neuroprotective molecular mechanisms of (−)-epigallocatechin-3-gallate: a reflective outcome of its antioxidant, iron chelating and neuritogenic properties. Genes Nutr 4:283–296
Yin ST et al (2009) Epigallocatechin-3-gallate induced primary cultures of rat hippocampal neurons death linked to calcium overload and oxidative stress. Naunyn Schmiedebergs Arch Pharmacol 379(6):551–564
Yoo KY et al (2010) (−)-Epigallocatechin-3-gallate increases cell proliferation and neuroblasts in the subgranular zone of the dentate gyrus in adult mice. Phytother Res 24(7):1065–1070
Acknowledgments
We thank Tanya Malpica-Llanos for technical assistance, Norman Haughey for providing rat neuronal cultures, and Avindra Nath at the National Institute of Neurological Diseases and Stroke, National Institutes of Health for carefully reading the manuscript and for helpful discussion/comments.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Nath, S., Bachani, M., Harshavardhana, D. et al. Catechins protect neurons against mitochondrial toxins and HIV proteins via activation of the BDNF pathway. J. Neurovirol. 18, 445–455 (2012). https://doi.org/10.1007/s13365-012-0122-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13365-012-0122-1