
Abstract
The Late Paleocene locality of Louvois is located about 20 km south of Reims, in the department of Marne (France). These marly sediments have yielded numerous vertebrate remains. The Louvois fauna is coeval with those of the localities of Cernay-lès-Reims and Berru and is dated as reference-level MP6, late Thanetian. Here we provide a detailed description of the remains of giant flightless gastornithids that were preliminarily reported in a study of the vertebrate fauna from Louvois. These fragmentary gastornithid remains mainly include a carpometacarpus, several tarsometatarsi, and numerous pedal phalanges. These new avian fossils add to the fossil record of Gastornis, which has been reported from various Early Paleogene localities in the Northern Hemisphere. Tarsometatarsi and pedal phalanges show large differences in size, which may be interpreted as sexual size dimorphism. The specimens from Louvois are morphologically different from those of Gastornis parisiensis and G. russelli, from Cernay and Berru. The Louvois carpometacarpus is also different from that of the North American G. giganteus, and the Louvois pedal phalanges are shorter and stouter than in other species of Gastornis. Moreover, the Louvois tarsometatarsi are more similar to that of the much younger G. sarasini, from Monthelon, reference-level MP 10, late Ypresian. We refrain from assigning the Louvois specimens to a new species of Gastornis and we designate it as Gastornis sp., owing to the fragmentary nature of the material. However, the morphological features of the Louvois material are sufficiently distinct for us to propose that three different forms of Gastornis were present in the Late Paleocene of North-eastern France.
Similar content being viewed by others

Introduction
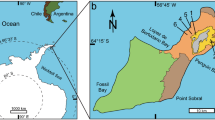
The fossiliferous locality of Louvois was discovered by M. Laurain during the digging of a ditch for a gas pipeline, and was reported by Laurain et al. (1985) and Laurain and Meyer (1986). Louvois is located about 20 km south of Reims, in the Parc Naturel Régional de la Montagne de Reims, in the department of Marne (Fig. 1). Fossils were found in marly sediments, which lie directly above the Late Cretaceous Chalk, and which are covered by colluviations of flint clays and siliceous limestones. The Louvois locality has yielded a rich vertebrate fauna, which was collected by Pierre Louis and given to the Muséum national d’Histoire naturelle (Paris). This fauna includes a small number of mammals and many aquatic forms such as crocodiles, turtles, amphibians and fishes. Remains of the giant flightless gastornithids are abundant, albeit very fragmentary (Louis 1996). The Louvois fauna is coeval with those of the localities of Cernay-lès-Reims and Berru and is dated as Late Paleocene, Thanetian, reference level MP6 (Buffetaut and Angst 2014). It is very similar to the fauna from Cernay, but the relative abundance of various vertebrate groups is different (Louis 1996). Louvois is located only 30 km south of the well-known localities of Cernay and Berru (Fig. 1), which have provided abundant material of Gastornis (Lemoine 1878, 1881; Martin 1992; Buffetaut 1997; Angst and Buffetaut 2013; Buffetaut and Angst 2014).

Map of France showing the localities where remains of Gastornis were found. Thanetian: 1 Cernay-lès-Reims, MP 6a; 2 Berru, MP 6a; 3 Louvois, MP 6; 4 Rivecourt, MP 6b. Ypresian: 5 Meudon, MP 7; 6 Passy, MP 7; 7 Mutigny, MP 7; 8 La Borie, Saint-Papoul, MP 8–9; 9 Monthelon, MP 10. These localities were reported by Buffetaut and Angst (2014), except that of Mutigny, which was reported by Russell et al. (1982), Martin (1992) and Louis (1996)
Remains of Gastornis were first discovered in the Early Eocene of Meudon near Paris (Hébert 1855). Since then, many specimens of these flightless birds have been reported from various Early Paleogene localities in the Northern Hemisphere (see Buffetaut 1997, 2013; Buffetaut and Angst 2013, 2014; Bourdon et al. 2014). Gastornis is currently known from the Middle Paleocene (Selandian) of Germany (Weigelt 1939; Mayr 2007); Late Paleocene (Thanetian) of France (Lemoine 1878, 1881; Martin 1992; Louis 1996; Buffetaut 1997; Angst and Buffetaut 2013; Smith et al. 2014) and Belgium (Dollo 1883); Early Eocene (Ypresian) of France (Hébert 1855; Milne-Edwards 1867–1868; Gaudry 1882; Schaub 1929a, b; Buffetaut 2008; Bourdon et al. 2014), England (Newton 1885, 1886), North America (Cope 1876; Matthew and Granger 1917; Andors 1988, 1992; Eberle and Greenwood 2012) and China (Hou 1980; Buffetaut 2013); and Middle Eocene (Lutetian) of Germany (Fischer 1962, 1978; Berg 1965; Hellmund 2013). Here we provide a detailed description of the remains of Gastornis from Louvois, which were mentioned by Louis (1996).
Materials and methods
The anatomical terminology follows Baumel and Witmer (1993), with English equivalents of the Latin nomenclature. The material is deposited in the collections of the Muséum national d’Histoire naturelle, Paris.
Some homologous elements, for example the distal tarsometatarsi, exhibit large differences in size. The material was therefore subdivided into two groups according to size. Most of the material belongs to the larger form. The smaller form is represented by a distal part of left tarsometatarsus (28 L), a phalanx 1 digit IV (15 L), a distal part of phalanx 1 digit IV (21 L), and a phalanx 2 digit IV (5 L).
Institutional abbreviations: AMNH, American Museum of Natural History, New York, USA; LACM, Natural History Museum of Los Angeles County, Los Angeles, California, USA; MNHN, Muséum national d’Histoire naturelle, Paris, France; NHMUK, Natural History Museum, London, United Kingdom; NMNH, National Museum of Natural History, Washington D.C., USA; NYPM-PU, Princeton University Collection, Peabody Museum of Natural History, Yale University, New Haven, Connecticut, USA.
Systematic palaeontology
Gastornithiformes STEJNEGER, 1885
Gastornithidae FÜRBRINGER, 1888
Gastornis HÉBERT, 1855
Gastornis sp.
Description
Tarsometatarsi (Fig. 2a–e)
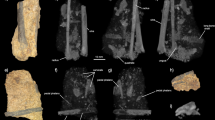
The material includes: a left tarsometatarsus (27 L + 24 L) with the distal part of the shaft and the trochlea metatarsi (tr. met.) III, the other trochleae being broken; a left tarsometatarsus (29 L) with the distal part of the shaft, the three trochleae being broken; a left tarsometatarsus (28 L) with the distal part of the shaft and the tr. met. III; an immature tr. met. III (13 L); an immature left tr. met. II (19 L). Their common characteristics are as follows: tr. met. III strongly projecting dorsally, with two condyles clearly separated by a longitudinal groove (27 L + 24 L); distal vascular foramen wide open and situated very close to the lateral intertrochlear groove; absence of distal interosseous canal; presence of slightly concave fossa for metatarsal I located on the plantar surface, proximal to tr. met. II. On the immature tr. met. III 13 L, both condyles are well preserved on the plantar surface, and the lateral condyle extends proximally farther than the medial one. On the dorsal surface of this trochlea the condyles are incompletely preserved, but it is obvious that the medial condyle is wider than the lateral one. On the lateral side of the plantar surface of tarsometatarsus 27 L, there is a lateral plantar crest that follows the medial side of the fossa for metatarsal I and that ends at the proximal part of the fracture of tr. met. II. The tr. met. II was plantarly displaced, according to the position of the fracture located at its base. The three adult tarsometatarsi strongly differ in size, specimen 28 L being much smaller than the other two ones (Table 1).

a–f Gastornis sp., Louvois. a, b 24+27 L, left tarsometatarsus, distal part, large form, dorsal (a) and plantar (b) views. c 29 L, left tarsometatarsus, distal part, large form, dorsal view. d, e 28 L, left tarsometatarsus, distal part, small form, dorsal (d) and plantar (e) views. f 31 L, right humerus, fragment of proximal part, caudal view
Pedal phalanges (Figs. 3, 4k–w)
The pedal phalanges of Gastornis giganteus from North America have been described in detail by Andors (1988), and these descriptions made it possible for us to identify the phalanges of the Gastornis from Louvois. Measurements of phalanges are provided in Tables 2 and 3.

a–v Gastornis sp., Louvois, pedal phalanges. a, b 17 L, right Ph. 1 D I, large form, dorsal (a) and plantar (b) views. c–f 7 L, Ph. 2 D I, juvenile, large form, dorsal (c), plantar (d), proximal (e) and lateral (f) views. g, h 12 L, right Ph. 1 D II, large form, dorsal (g) and plantar (h) views. i–j 11 L, right Ph. 1 D IV, juvenile, large form, dorsal (i) and plantar (j) views. k, l 15 L, left Ph. 1 D IV, juvenile, small form, dorsal (k) and plantar (l) views. m, n 14 L, right Ph. 2 D III, large form, dorsal (m) and plantar (n) views. o, p 4 L, right Ph. 3 D III, large form, dorsal (o) and plantar (p) views. q, r 10 L, left Ph. 3 D III, large form, dorsal (q) and plantar (r) views. s–v 3 L, right Ph. 4 D III, large form, dorsal (s), lateral (t), plantar (u) and proximal (v) views

a–d Gastornis sp., Louvois, 22 L, right carpometacarpus, ventral (a), dorsal (b), cranial (c) and caudal (d) views. e, f Gastornis parisiensis, Cernay, MNHN CR 10537, right carpometacarpus, ventral (e) and dorsal (f) views. g, h Gastornis giganteus, Bighorn Basin, Wyoming, USA, LACM 6882/31732, right carpometacarpus, ventral view (g), left carpometacarpus, ventral view (h). i Gastornis sp., Louvois, 23 L, fragment of synsacrum, ventral view. j Gastornis sp. Louvois, 33 L, fragment of cervical vertebra, dorsal view. k–w Gastornis sp., Louvois, pedal phalanges. k, l 2 L, right Ph. 2 D IV, large form, immature, dorsal (k) and plantar (l) views. m–o 8 L, right Ph. 2 D IV, large form, dorsal (m), plantar (n) and proximal (o) views. p–r 5 L, right Ph. 2 D IV, small form, dorsal (p), plantar (q) and proximal (r) views. s–u 18 L, left Ph. 3 D IV, large form, dorsal (s), plantar (t) and proximal (u) views. v–w 9 L, right Ph. 4 D IV, large form, dorsal (v) and plantar (w) views
Digit I
17 L distal part of the right phalanx 1 D I (Fig. 3a, b)
The distal part is dorsoventrally flattened and slightly wider than the shaft. The distal articular surface (articular trochlea) shows a faintly discernible median groove. The medial portion of the trochlea projects medially and dorsally. The foveae for collateral ligaments are shallow. The shaft section is taller than it is wide at the level of the broken proximal part. The dorsal surface is rounded and the plantar surface is flattened. This phalanx is slightly larger than the corresponding phalanges of Gastornis giganteus (Andors 1988, p. 492, Table 13).
7 L ungual phalanx 2 D I, immature (Fig. 3c–f)
This phalanx is very short and slightly curved plantarly. The proximal articular surface (articular cotyla) is circular, the dorsal surface is rounded and the plantar surface is flattened. The neurovascular grooves are well developed and converge dorsally at the apex.
Digit II
12 L Right phalanx 1 D II (Fig. 3g, h)
The shaft is cylindrical, slightly flattened in its distal part. The partially preserved articular cotyla is shallow, subtriangular, flattened on the lateral and plantar surfaces and rounded on the dorsal and medial surfaces. The proximal part of the plantar surface exhibits a shallow median groove that is bounded on either side by plantar ridges. The lateral plantar ridge is incompletely preserved. The articular trochlea is partly preserved, both on its dorsal and plantar surfaces. The medial condyle of the trochlea is expanded medially and plantarly. The intercondylar groove is shallow. The medial ligamental fovea is shallow and the lateral one is deeper.
Digit III
14 L Right phalanx 2 D III (Fig. 3m, n)
This phalanx is short and wide. Part of the articular cotyla and articular trochlea is missing. The articular cotyla is semi-circular, slightly concave mediolaterally, with dorsal and plantar rims projected proximally in the midline. The articular trochlea is wide, with a deep and massive medial condyle and a broad and shallow intercondylar groove. The partly preserved lateral ligamental fovea is very deep, much deeper than the medial one. The plantar surface is concave.
4 L Right phalanx 3 D III and 10 L Left phalanx 3 D III (Fig. 3o–r)
These two phalanges are slightly different and do not seem to come from the same individual. Phalanx 10 L is short and massive. Its articular cotyla is mediolaterally concave with extensor and flexor tubercles both projecting in the midline. The articular trochlea is mediolaterally elongate and shows two barely distinct condyles, which are separated by a shallow groove. The lateral ligamental fovea is deep and proximodistally elongate. The medial ligamental fovea is barely distinct. The plantar surface is pierced by a large quantity of small openings. Phalanx 4 L is very similar to phalanx 10 L. The extensor tubercle and the lateroplantar corner of the articular cotyla are broken. The deep lateral ligamental fovea is circular in shape and extends proximally on most the lateral surface of the bone, whereas the medial ligamental fovea is very slightly developed.
3 L Right ungual phalanx 4 D III (Fig. 3s–v)
This phalanx is a markedly flattened and slightly plantarly curved claw. Its proportions are very different from those of the ungual phalanx of digit II. The apex and a part of the lateral side are broken. The articular cotyla is mediolaterally elongate and concave in both dorsoplantar and mediolateral directions. Bony excrescences mask the neurovascular grooves on each side of the articular surface (Fig. 3v). The plantar rim of the articular cotyla protrudes in the midline as a very low flexor tubercle. The plantar surface is flattened. The outgrowth on the lateral side forms a ridge that is only partly preserved (Fig. 3u), and the outgrowth on the medial side forms a right angle with the plantar surface (Fig. 3t).
Digit IV
11 L Right immature phalanx 1 D IV (Fig. 3i, j)
The proximal articular surface is flattened with numerous holes and grooves giving a vermiculated appearance. A multitude of small orifices, visible with a binocular microscope, are located on both proximal and distal articular surfaces. The surface of the shaft is fibrous at the level of its junction with the articular surfaces, and with numerous little holes. This phalanx is wider in the proximal part than in the distal one, and the shaft progressively narrows in distal direction. The lateral part of the articular cotyla is dorsoplantarly narrower than the medial part, and is prolonged by a smooth ridge that extends to mid-length of shaft. The lateral condyle of the articular trochlea projects laterally, plantarly and proximally, forming a distinct tubercle. The medial ligamental fovea is shallow, whereas the lateral ligamental fovea is well developed and circular in shape. The plantar surface is flat and does not show longitudinal plantar ridges, especially the prominent medial plantar ridge described by Andors (1988, p. 261). However, Andors also indicates that this medial plantar ridge is “merely incipient” in the specimen previously named Omorhamphus storchii, which is an immature specimen of Gastornis giganteus (Andors 1992, p. 112). Despite being very immature, the phalanx 11L is 75.9 mm in total length, which is close to the dimensions of the largest individuals of G. giganteus (68.6 to 80.1 mm, in Andors 1988, Table 13).
15 L Left immature phalanx 1 D IV, small form (Fig. 3k, l)
This phalanx shows the same characteristics as phalanx 11 L, albeit more immature. On the articular trochlea, the lateral condyle is less prominent proximolaterally than in phalanx 11 L. This specimen most probably belongs to the small form.
21 L distal part of right immature phalanx 1 D IV, small form?
This phalanx shows a proximolaterally projecting lateral condyle.
3 Phalanges 2 D IV: 2 L right phalanx, large form, slightly immature (Fig. 4k, l); 8 L right phalanx, large form (Fig. 4m–o); 5 L right phalanx, small form (Fig. 4p–r)
These phalanges are short, stout and dorsoplantarly flattened, but not as stout as phalanx 2 D III. The articular cotyla is bluntly triangular in shape. It is slightly dorsoplantarly concave with a small ridge in the middle. The extensor tubercles are well developed. The articular trochlea exhibits two condyles separated by a shallow intercondylar groove. The lateral condyle projects proximolaterally. The plantar surface is concave and pierced by numerous small foramina. The medial ligamental fovea is shallow, whereas the lateral ligamental fovea is deep and curved proximally. Phalanx 5 L shows the same morphological characteristics as phalanx 8 L, but its size is on average 28 % smaller.
18 L Left phalanx 3 D IV (Fig. 4s–u)
This phalanx is short and trapezoidal, the medial side being shorter than the lateral side. The slightly concave articular cotyla is bluntly triangular, nearly semi-circular in shape. The extensor tubercle is moderately projecting. The articular trochlea exhibits two condyles, and the intercondylar groove is almost absent. The plantar surface is concave and shows numerous small holes. The medial ligamental fovea is almost absent, whereas the lateral ligamental fovea is circular and deep.
9 L Right phalanx 4 D IV (Fig. 4v, w)
This phalanx is very short and trapezoidal. It is shorter laterally than medially, unlike phalanx 3 D IV. The articular trochlea joins the articular cotyla on the lateral side. The articular cotyla and the plantar surface are slightly concave. The extensor tubercle is rounded. The medial condyle of the distal trochlea is broken. The medial ligamental fovea is indistinct, and there is no space on the lateral side for a fovea.
Humerus (Fig. 2f)
Specimen 31 L consists of a fragment of proximal part of right humerus. It includes the base of the ventral tubercle and a part of the humeral head. On the caudal surface, these two parts are separated by a shallow depression, which can be interpreted as the capital groove. A series of deep foramina are situated beneath the distal edge of the humeral head. The cranial surface is flat. The depth of the humeral head is 22.1 mm (as preserved). A few foramina are also present beneath the edge of the humeral head in a humerus of Gastornis parisiensis from Berru (MNHN BR12137), albeit less developed than in the humerus from Louvois.
Carpometacarpus (Fig. 4a–d)
The material from Louvois includes an almost complete right carpometacarpus (22 L). This bone is slightly concave ventrally. The distal and caudal parts of the carpal trochlea and the distal part of the alular process are missing. The distal part of the symphysis of the major and minor metacarpal bones is incompletely preserved. The carpal trochlea is very wide dorsoventrally. The cranial carpal fovea is wide and pierced by multiple small foramina. The extensor process is blunt. The stout major metacarpal bone is almost circular in cross section, and it tapers and flattens distally. The minor metacarpal bone is very narrow, flattened craniocaudally, and oriented somewhat obliquely. It is fused with the major metacarpal bone over half of the total length of the bone. The intermetacarpal space is short but well developed. There is neither a pisiform process nor an infratrochlear fossa on the proximal part of the ventral surface. It is not possible to tell whether articular facets for the wing phalanges were present, since these parts of the bone are not well preserved. The morphological characteristics of this carpometacarpus agree with those given by Andors for G. giganteus. Another similarity between the two carpometacarpi is the presence of very numerous little foramina in the cranial carpal fossa. This feature also occurs in one of the carpometacarpi of specimen LACM 6882/31732 (Fig. 4g, h), in which this fossa “forms a deep cribriform groove” (Andors 1988, p. 210). In the Louvois carpometacarpus, small foramina are also present in the proximal part, in the region of fusion between the major, minor and alular metacarpal bones. Measurements of the carpometacarpus 22 L are given in Table 4.
Cervical vertebra (Fig. 4j)
Specimen 33 L is a fragment of a cervical vertebra, including the vertebral arch, vertebral foramen and part of the left cranial zygapophysis. The vertebral body and the other zygapophyses are not preserved. A medial and two transverse-oblique crests are found on the dorsal surface. A crescent-shaped area ligamenti elastici is located in the caudal part of the vertebral arch and shows roughened tuberosities. This vertebra is characterized by the small craniocaudal length of the vertebral arch. In Gastornis parisiensis from La Borie, the cervical vertebrae are also greatly abbreviated craniocaudally (Bourdon et al. 2014). The medial longitudinal length of the vertebral arch of the Louvois vertebra (30 mm) is comparable to that of the cervical vertebrae from La Borie. Other measurements (in mm) are as follows: minimal width of vertebral arch, 45.0; transverse diameter of vertebral foramen, 12.2; dorsoventral diameter of vertebral foramen, 11.3.
Synsacrum (Fig. 4i)
Specimen 23 L corresponds to the small part of synsacrum situated at the level of the acetabular foramina. It includes the ventral surface of the synsacrum and the synsacral canal. On the left side, the synsacrum exhibits the costal process of the last lumbar vertebra, which is inserted in a socket. The tip of this process is damaged, and originally rested against the preacetabular wing of the ilium. On the right side, the socket is also present but the costal process is broken. The ventral surface is flat at the level of the costal processes and does not show any ventral sulcus. More caudally the surface shows a blunt median ridge bounded on each side by a depressed area. The synsacral canal is wide at the cranial end (13.3 mm), but its width strongly decreases towards the caudal end (5.7 mm). The width of the cranial part between the tips of the costal processes is estimated at 60 mm, whereas it measures 73.1 mm in G. giganteus AMNH 6169 (Andors 1988).
In addition to the specimens described above, the material of Gastornis from Louvois also includes the following elements (not figured): 1 L, corpus atlantis; 25 L, 30 L, 34 L, fragments of vertebrae; 37 L, 38 L, 41 L, fragments of pelvis?; 20 L, femoral head; 26 L, fragment of distal part of immature tibiotarsus.
Comparisons with other European forms of Gastornis
The tarsometatarsi from Louvois can be compared with a tarsometatarsus from Berru, which was formerly preserved in Dr. Levé’s collection, and which is now in a private collection (Fig. 5a, c). This tarsometatarsus is figured in Martin (1992: fig. 5, A-F), Angst (2014: fig. 27) and Buffetaut and Angst (2014: fig. 3, B). A cast of this specimen (no. L3092) and a cast of another tarsometatarsus from Dr. Levé’s collection (no. L3093) are housed in the MNHN collection. These two tarsometatarsi were assigned to Gastornis parisiensis Hébert, 1855 by Martin (1992). In these specimens, the tr. met. III is only slightly projecting dorsally and the tr. met. IV is only slightly plantarly displaced. In the Louvois tarsometatarsi, the tr. met. III is strongly projecting dorsally. The tr. met. IV is broken in the specimens from Louvois, but it seems that this trochlea was plantarly displaced according to the position of the fracture surfaces. In specimen no. L3093, there is a foramen in the lateral intertrochlear notch that indicates the presence of a dorsally closed distal interosseous canal, whereas the distal interosseous canal is dorsally open in no. L3092. In the tarsometatarsi from Louvois, the distal vascular foramen is located very close to the intertrochlear notch and there is no indication of distal interosseous canal. Lastly, there is no distinct fossa for metatarsal I on the plantar surface of the two tarsometatarsi from Berru, while this fossa is present in the tarsometatarsi from Louvois.

Comparison of the distal part of left tarsometatarsi, in plantar views: a Gastornis parisiensis, cast no. L3092, MNHN Paris, from Berru; b Gastornis sarasini, from Monthelon (after Schaub 1929b); c Gastornis sp. no. 24 + 27 L, from Louvois
A trochlea met. III of G. parisiensis, from the “Conglomérat de Passy”, has been described and figured by Milne-Edwards (1867–1868: pl. XXIX, figs. 6–7). Its age is Early Eocene, reference-level MP 7 (Buffetaut and Angst 2014). Its dimensions are similar to those of the larger form from Louvois. The distal part of the tarsometatarsus is known in G. sarasini from Monthelon (Fig. 5b), and is figured in Schaub (1929b: fig. 1–3), Mayr (2009: fig. 6.5, C), Hellmund (2013: fig. 7, b), and Buffetaut and Angst (2014: fig. 3, D). The tarsometatarsi from Louvois are more similar to that of G. sarasini than to those from Berru. In G. sarasini the tr. met. III projects dorsally and extends proximally as a flattened ridge, there is no closed distal interosseous canal, and the fossa for metatarsal I is present. The tarsometatarsus 27 L + 24 L from Louvois is similar to that of Monthelon because, as in the latter, the width of the shaft strongly decreases proximally above the distal vascular foramen. In contrast, in the specimens from Berru, the shaft remains wide above the distal vascular foramen, and the medial and lateral edges of the shaft are almost parallel. The tarsometatarsus of the small species G. russelli from Berru (R3560) is more similar to those of G. parisiensis from the same locality than to the Louvois tarsometatarsi, because the shaft has parallel edges above the distal vascular foramen, and because the fossa for metatarsal I is indistinct (Martin 1992: fig. 6). It is not possible to compare the Louvois tarsometatarsi with those of G. geiselensis Fischer, 1962, from Geiseltal, because the trochleae are very poorly preserved in the latter species (Fischer 1962: figs. 9–10; Fischer 1978: pl. XI, fig. 7, a–b; Hellmund 2013: figs. 3 and 7, 8).
Phalanges 1 of posterior digit II are known from the localities of Berru (MNHN no. R3567) and from the collection of the University of Reims (no. LR BR A5), Cernay and Meudon (Gaudry 1882) for Gastornis parisiensis, Monthelon for G. sarasini (Schaub 1929a, b), and Geiseltal for G. geiselensis (Hellmund 2013). Their dimensions are given in Table 5. In the phalanx 1 D II from Louvois (12 L), the total length is close to that of G. parisiensis from Berru (R3567) but the phalanx has wider proximal and distal extremities. The size of phalanx 1 D II is close to that of G. geiselensis, and larger than those of G. parisiensis from Cernay, Berru (LR BR A5) and Meudon, and G. sarasini from Monthelon. A phalanx 2 of posterior digit III is known from the locality of Berru (MNHN no. R3564; see Table 6). The phalanx 2 D III from Louvois (14 L) is shorter and wider than that of Berru. As a whole, the Louvois phalanges are shorter and stockier than the phalanges of G. parisiensis figured by Lemoine (1881: pl. 6, figs. 6 and 8) or Buffetaut (1997: fig. 1, d, e).
A right carpometacarpus of G. parisiensis, CR 10537 (Fig. 4e, f), from Cernay, is figured by Lemoine (1893, pl. XI, figs. 3–3a), and Lambrecht (1933, fig. 166, A, B). This carpometacarpus differs markedly from that of Louvois. It is more flattened dorsoventrally, the carpal trochlea is globular, and the alular metacarpal is almost semi-circular in shape and only slightly projected cranially. The minor metacarpal is parallel to and fused throughout almost its entire length with the major metacarpal, and the intermetacarpal space consists of three small foramina on the ventral surface (Fig. 4e) and an elongate groove on the dorsal surface (Fig. 4f). There are no visible articular surfaces for the wing phalanges. The Cernay major metacarpal is rectilinear, while that of Louvois tapers distally and is ventrally concave. Finally, the Cernay carpometacarpus is smaller than the Louvois one; its total length is only 67.3 mm while that of Louvois is 81.8 mm.
Comparison with Gastornis giganteus
The dimensions of the tr. met. III of the larger form from Louvois are within the range of tr. met. III of G. giganteus, whereas the dimensions of the smaller form are outside this range. The phalanges of the large form from Louvois are shorter and proportionally stockier than in G. giganteus, for example LACM 6882/31732 (Andors 1988, table 13). The ratio between the total length and the proximal width of the phalanges is always higher in G. giganteus than in the Gastornis from Louvois.
The carpometacarpus 22 L (Fig. 4a–d) is much larger than the carpometacarpi of G. giganteus (Fig. 4g, h). The dorsoventral depth of the carpal trochlea, the craniocaudal width of the proximal metacarpal symphysis, and the length from the proximal articular surface of the carpal trochlea to the distal extremity of the major metacarpal bone are on average 50 % larger in the Gastornis from Louvois (Table 5). The main difference between the carpometacarpus from Louvois and those of G. giganteus is the presence of an intermetacarpal space in the specimen from Louvois, while in the North American form “all that remains of the primary intermetacarpal space are dorsal and ventral grooves penetrated by at most one or two minute foramina” (Andors 1988, p. 209). In contrast to the Louvois specimen, the ventral surface of the synsacrum of G. giganteus exhibits a shallow ventral sulcus bordered on either side by a smooth ridge. Moreover, in G. giganteus, the costal processes of the last lumbar vertebra are not inserted in sockets.
Discussion
Taxonomic considerations
Hellmund (2013) recognized five valid species of Gastornis, including G. parisiensis, G. russelli, G. sarasini, G. giganteus, and G. geiselensis. A sixth species, G. xichuanensis, is known from the Early Eocene of China (Buffetaut 2013). However, the number of species of Gastornis is still uncertain, because an in-depth systematic revision of this taxon has not been undertaken yet.
The Gastornis from Louvois differs from the coeval G. parisiensis from Cernay and Berru in several morphological characteristics: shaft of tarsometatarsus narrower above trochleae; well-developed fossa for metatarsal I delimited by a ridge; posterior phalanges proportionally shorter and more robust; carpometacarpus longer, stouter, less flattened, distally tapered, showing well-developed intermetacarpal space and more prominent alular metacarpal process.
In G. parisiensis from La Borie (Saint-Papoul, Southern France), Bourdon et al. (2014) also noticed some morphological differences in the quadrate compared to the material of Cernay and Berru. La Borie locality corresponds to the reference-level MP 8–9 and its age is between 55 and 51 Ma (Buffetaut and Angst 2014).
According to the morphology of the tarsometatarsus, the Gastornis from Louvois is more similar to G. sarasini from Monthelon (Fig. 5), which, unfortunately, is only known from a distal part of tarsometatarsus and a pedal phalanx. The Gastornis from Monthelon was found in the “Sables à Unio et à Térédines” (Duprat 1997) that correspond to reference-level MP 10, which is between 50.5 and 47.8 Ma in age (Buffetaut and Angst 2014). Therefore, it is much younger than the Gastornis from Louvois, the difference in age being about 8 Ma. It is possible that the Gastornis from Louvois and G. parisiensis belong to two different lineages, and that the Louvois lineage is represented by G. sarasini in the Eocene. However, we hesitate to assign the Louvois specimens to a new species of Gastornis due to the fragmentary nature of the material. Hence, we assign the Louvois specimens to Gastornis sp.
The Gastornis from Louvois occurs in the same levels, and in the same area, as the species G. parisiensis from Cernay and Berru, and G. russelli from Cernay. This means that three different forms of Gastornithidae occurred in the Reims area during the late Thanetian. However, it is possible that the three forms of Gastornis were not present at the same time and in the same place. The Gastornis from Louvois may have had a slightly different habitat and/or diet from those of the other two species.
Sexual size dimorphism
Differences in size are noticeable in the tarsometatarsi and some phalanges. Tarsometatarsi 24 L + 27 L and 29 L are between 30 and 65 % larger than the tarsometatarsus 28 L (Table 1). This small-sized tarsometatarsus cannot be attributed to the smaller species Gastornis russelli described by Martin (1992). The latter species is known from an almost complete tarsometatarsus (holotype no. R3560) and a bill fragment. The measurements given by Martin (1992, p. 102) for the tarsometatarsus are inaccurate. The length (129 mm) is actually 164 mm, the proximal width (31 mm) is actually 48.0 mm, and the distal width (32 mm) is actually 52.6 mm. However, the small form from Louvois is larger than G. russelli. The width of tr. met III on the distal surface is 19.4 mm for G. russelli versus 24 mm (estimated) for the Louvois small form 28 L. Likewise, the depth of medial condyle is 23.2 mm for G. russelli versus 34 mm (estimated) for Louvois small form 28 L. In the pedal phalanges, the phalanx 1 of digit II of the large form (12 L) is on average 29 % larger than the phalanx of the small form (21 L). For the phalanges 1 of digit IV, the large form is on average 50 % larger than the small form, but these two phalanges are juvenile and it is possible that the small form did not reach the same degree of development as the large form. Lastly, for the two phalanges 2 of digit IV, which are adult specimens, the large form is on average about 40 % larger than the small form.
The degree of sexual size dimorphism is variable in large flightless birds. An extreme reversed sexual size dimorphism was found in the Moa (Dinornithidae), extinct giant ratites endemic to New Zealand (Bunce et al. 2003; Olson and Turvey 2013). In these New Zealand Moa, the largest females reached about 150 % the height and 280 % the weight of the largest males. In extant large flightless palaeognaths, the male is larger in Struthio camelus and Rhea americana, the sexes are similar in size in Pterocnemia pennata, and the female is larger in Casuarius casuarius and Dromaius noveahollandiae (Blake 1977). However, no clear evidence for extreme sexual size dimorphism was found in Genyornis newtoni (Dromornithidae), an extinct giant anseriform endemic to Australia (Chan 2014). A possible sexual size dimorphism in the Gastornithidae has been suggested by Hellmund (2013) for G. geiselensis, and by Bourdon et al. (2014) for G. parisiensis. Hellmund (2013) proposed an increase in size through time, between the levels corresponding to reference-level MP 11 and those corresponding to reference-level MP 13, as an alternative hypothesis. This latter hypothesis is not well supported, however, since the largest femur (total length ca. 33 mm) and the smallest femur (total length ca. 30 mm) are found in sites corresponding to the same reference-level (MP 11). The hypothesis of a sexual size dimorphism in Gastornis is likely, but it is not possible to determine whether the largest form from Louvois corresponds to males or to females.
References
Andors, A. V. (1988). Giant groundbirds of North America (Aves, Diatrymidae). Unpublished Ph.D. Thesis, Columbia University, New York.
Andors, A. V. (1992). Reappraisal of the Eocene groundbird Diatryma (Aves: Anserimorphae). Natural History Museum of Los Angeles County, Science Series, 36, 109–125.
Angst, D. (2014). Successeurs des dinosaures ? Paléobiologie et paléoécologie d’un Oiseau géant terrestre du Paléogène. Unpublished Ph.D. Thesis, Université Claude Bernard-Lyon 1, Lyon.
Angst, D., & Buffetaut, E. (2013). The first mandible of Gastornis Hébert, 1855 (Aves, Gastornithidae) from the Thanetian (Paleocene) of Mont-de-Berru (France). Revue de Paléobiologie, 32, 423–432.
Baumel, J. J., & Witmer, L. M. (1993). Osteologia. In J. J. Baumel (Ed.) Handbook of Avian Anatomy. Nomina Anatomica Avium, 2nd edition. Nuttall Ornithological Club, 23, 45–132.
Berg, D. E. (1965). Nachweis des Riesenlaufvogels Diatryma im Eozän von Messel bei Darmstadt/Hessen. Notizblatt des hessischen Landesamtes für Bodenforschung, 93, 68–72.
Blake, E. R. (1977). Manual of Neotropical Birds, vol. 1. Chicago: University of Chicago Press.
Bourdon, E., Mourer-Chauviré, C., & Laurent, Y. (2014). The birds (Aves) from the Early Eocene of La Borie, southern France. Acta Palaeontologica Polonica. doi: 10.4202/app.00083.2014.
Buffetaut, E. (1997). New remains of the giant bird Gastornis from the Upper Paleocene of the eastern Paris Basin, and the relationships between Gastornis and Diatryma. Neues Jahrbuch für Geologie und Paläontologie Monatshefte, 3, 179–190.
Buffetaut, E. (2008). First evidence of the giant bird Gastornis from southern Europe: a tibiotarsus from the Lower Eocene of Saint-Papoul (Aude, southern France). Oryctos, 7, 75–82.
Buffetaut, E. (2013). The giant bird Gastornis in Asia: a revision of Zhongyuanus xichuanensis Hou, 1980, from the Early Eocene of China. Paleontological Journal, 47(11), 1302–1307.
Buffetaut, E., & Angst, D. (2013). “Terror cranes” or peaceful plant-eaters: changing interpretations of the palaeobiology of gastornithid birds. Revue de Paléobiologie, 32(2), 413–422.
Buffetaut, E., & Angst, D. (2014). Stratigraphic distribution of large flightless birds in the Paleogene of Europe and its palaeobiological and palaeogeographical implications. Earth-Science Reviews, 138, 394–408.
Bunce, M., Worthy, T. H., Ford, T., Hoppit, W., Willerslev, E., Drummond, A., & Cooper, A. (2003). Extreme reversed sexual size dimorphism in the extinct New Zealand moa Dinornis. Nature, 425, 172–175.
Chan, N. R. (2014). Does size variation in Genyornis newtoni (Aves, Dromornithidae) encompass eggshell safety limits? Journal of Vertebrate Paleontology, 34(4), 976–979.
Cope, E. D. (1876). On a gigantic bird from the Eocene of New Mexico. Proceedings of the Academy of Natural Sciences of Philadelphia, 28(2), 10–12.
Dollo, L. (1883). Note sur la présence du Gastornis edwardsii Lemoine dans l’assise inférieure de l’étage landénien à Mesvin, près Mons. Bulletin du Musée Royal d’Histoire Naturelle de Belgique, 2, 297–305.
Duprat, M. (1997). Les faciès à mammifères (MP 6 à MP 16) dans le Nord-Est du Bassin de Paris (France): Argumentation du modèle tectono-sédimentaire des dépôts paléogènes. In J.-P. Aguilar, S. Legendre, & J. Michaux (Eds.), Actes du Congrès BiochroM’97 (pp. 315–336). Institut de Montpellier: Mémoires et Travaux E. P. H. E.
Eberle, J. J., & Greenwood, D. R. (2012). Life at the top of the greenhouse Eocene world—a review of the Eocene flora and vertebrate fauna from Canada’s High Arctic. GSA Bulletin, 124, 3–23.
Fischer, K. (1962). Der Riesenlaufvogel Diatryma aus der eozänen Braunkohle des Geiseltales. Hallesches Jahrbuch für Mitteldeutsche Erdgeschichte, 4, 26–33.
Fischer, K. (1978). Neue Reste des Riesenlaufvogels Diatryma aus dem Eozän des Geisteltales bei Halle (DDR). Annalen für Ornithologie, 2, 133–144.
Gaudry, A. (1882). Sur de nouvelles pièces que M. Gaston Planté a recueillies dans les Argiles ligniteuses du Bas-Meudon. Bulletin de la Société Géologique de France, 10, 236–238.
Hébert, E. (1855). Note sur le tibia du Gastornis parisiensis. Comptes Rendus de l’Académie des Sciences, 40, 579–582.
Hellmund, H. (2013). Reapparaisal of the bone inventory of Gastornis geiselensis (Fischer, 1978) from the Eocene Geiseltal Fossillagerstätte (Saxony-Anhalt, Germany). Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 269(2), 203–220.
Hou, L. (1980). New form of the Gastornithidae from the Lower Eocene of the Xichuan, Honan. Vertebrata Palasiatica, 18, 111–115.
Lambrecht, K. (1933). Handbuch der Palaeornithologie. Berlin: Borntraeger Verlag. 1029 pp.
Laurain, M., Allouc, J., Leroux, J., Louis, P., Monciardini, C., & Morfaux, P. (1985). Notice explicative de la feuille Avize à 1/50 000. Orléans: BRGM édit. 37 pp.
Laurain, M., & Meyer, R. (1986). Stratigraphie et paléogéographie de la base du Paléogène champenois. Géologie de la France, 1986(1), 103–123.
Lemoine, V. (1878). Recherches sur les oiseaux fossiles des terrains tertiaires inférieurs des environs de Reims. Reims: Keller. 69 pp.
Lemoine, V. (1881). Recherches sur les oiseaux fossiles des terrains tertiaires inférieurs des environs de Reims. Deuxième partie (pp. 75–170). Reims: Matot-Braine.
Lemoine, V. (1893). Etude sur les os du pied des mammifères de la faune Cernaysienne et sur quelques pièces osseuses nouvelles de cet horizon paléontologique. Bulletin de la Société géologique de France, 21, 353–368.
Louis, P. (1996). Recherche de mammifères paléogènes dans les départements de l’Aisne et de la Marne pendant la deuxième moitié du vingtième siècle. In M. Godinot & P. D. Gingerich (Eds.), Paléobiologie et Evolution des mammifères paléogènes. Volume jubilaire en hommage à Donald E. Russell. Palaeovertebrata, 25(2–4), 83–113.
Martin, L. D. (1992). The status of the Late Paleocene birds Gastornis and Remiornis. Natural History Museum of Los Angeles County, Sciences Series, 36, 97–108.
Matthew, W. D., & Granger, W. (1917). The skeleton of Diatryma, a gigantic bird from the Lower Eocene of Wyoming. Bulletin of the American Museum of Natural History, 37, 307–326.
Mayr, G. (2007). The birds from the Paleocene fissure filling of Walbeck (Germany). Journal of Vertebrate Paleontology, 27(2), 394–408.
Mayr, G. (2009). Paleogene Fossil Birds. Berlin: Springer.
Milne-Edwards, A. (1867–1868). Recherches anatomiques et paléontologiques pour servir à l’histoire des oiseaux fossiles de la France. Tome premier. Victor Masson et Fils, Paris, 475 pp.
Newton, E. T. (1885). Gastornis klaasseni Newton, a gigantic bird from the Lower Eocene of Croydon. Geological Magazine, 3, 362–364.
Newton, E. T. (1886). On the remains of a gigantic bird (Gastornis klaasseni, n. sp.) from the Lower Eocene beds near Croydon. Transactions of the Zoological Society of London, 12, 143–160.
Olson, V. A., & Turvey, S. T. (2013). The evolution of sexual dimorphism in New Zealand giant moa (Dinornis) and other ratites. Proceedings of the Royal Society, B, 280, 20130401.
Russell, D. E., and coll. (1982). Tetrapods of the Northwest European Tertiary Basin. Geologisches Jahrbuch, A60, 5–74.
Schaub, S. (1929a). Über einen neuen Riesenvogel aus den Teredinasanden von Monthelon bei Epernay. Verhandlungen der Schweizerischen Naturforschenden Gesellschaft, 110, 195.
Schaub, S. (1929b). Über eocäne Ratitenreste in der osteologischen Sammlung des Basler Museums. Verhandlungen der Naturforschenden Gesellschaft in Basel, 40(2), 588–598.
Smith, T., Quesnel, F., De Ploëg, G., De Franceschi, D., Métais, G., et al. (2014). First Clarkforkian equivalent Land Mammal Age in the latest Palaeocene basal Sparnacian facies of Europe: fauna, flora, paleoenvironment and (bio)stratigraphy. PLoS One, 9(1), e86229. doi:10.1371/journal.pone.0086229.
Weigelt, J. (1939). Die Aufdeckung der bisher ältesten tertiären Säugetierfauna Deutschlands. Nova Acta Leopoldina, 7, 515–528.
Acknowledgments
We are very grateful to Pierre Louis for giving access to the fossil bird material from Louvois. We thank Carl Mehling (AMNH), Helen James and Marc Florence (NMNH), Sandra Chapman (NHMUK) and Ronan Allain (MNHN) for providing access to the fossil bird collections. Kenneth Campbell (LACM) kindly sent us some pictures of the G. giganteus material housed in his institution. This work was funded by a Carlsberg grant No. 2013_01_0480 (Carlsbergfondet) to EB. We also thank associate editor Loïc Costeur and two anonymous reviewers for useful comments on the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Mourer-Chauviré, C., Bourdon, E. The Gastornis (Aves, Gastornithidae) from the Late Paleocene of Louvois (Marne, France). Swiss J Palaeontol 135, 327–341 (2016). https://doi.org/10.1007/s13358-015-0097-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13358-015-0097-7