
Abstract
Coastal marshes are composed of distinct vegetation assemblages that contribute to ecosystem functions and services over time and space. In recent decades, natural and anthropogenic changes have led to large-scale marsh loss along the Chenier Plain of the Gulf of Mexico. An understanding of the composition and dominance of the vegetation assemblages that form these coastal marshes is necessary to effectively assess and monitor their stability under changing conditions. To investigate marsh vegetation assemblages along the Texas Chenier Plain, we analyzed the composition and dominance of 135 marsh plots distributed across two study areas bordering the East Bay of Galveston Bay in Chambers County, Texas using cluster analysis. Seven general vegetation assemblages were identified. The assemblages were evaluated using indicator species analysis and Multi-Response Permutation Procedures and compared to marsh vegetation assemblages found in the Louisiana Chenier Plain. Our data provide a foundation for better understanding of vegetation patterns found along the East Texas Chenier Plain allowing managers and conservationists the ability to better evaluate the effects of marsh change due to climatic and human driven pressures.
Similar content being viewed by others

Introduction
Coastal marshes provide important ecosystem functions that contribute many ecosystem goods and services to humanity. For example, coastal marshes provide storm buffering and flood control, carbon sequestration, water filtration and detoxification, and provide fisheries production to coastal communities (Mitsch and Gosselink 2000; Koch et al. 2009; Barbier 2011; Deegan et al. 2012). These important goods and services, however, are at risk because of the vulnerability of coastal marshes to global change. The Intergovernmental Panel on Climate Change (IPCC) has predicted that global warming will result in eustatic sea-level rise of approximately 20–60 cm over the next century (Nicholls et al. 2007). Within the western Gulf of Mexico, sea-level is estimated to rise 5–10 mm yr−1 (Parris et al. 2012), and if coastal marsh accretion is unable to keep pace with sea-level rise marshes will be damaged or lost (Reed 1995). Coastal marsh ecosystem goods and services are intimately linked to marsh vegetation through their composition and dominance. For instance, the effects of seasonality on standing biomass production influence the ability of vegetation to attenuate storm waves providing a service of coastal protection (Koch et al. 2009). Abiotic and biotic interactions control the composition and dominance of marsh vegetation (Mitsch and Gosselink 2000). Knowing how the composition and dominance of characteristic marsh vegetation respond to environmental changes (e.g. accelerated sea-level rise, increased storm events, alterations of climate regimes and anthropogenic modification) is critical for effectively managing and conserving these systems. Our study focuses on the variation in composition and dominance of different marsh species as indicators of vegetation assemblages along the Gulf of Mexico Chenier Plain of Texas.
Vegetation classification is used to find floristic similarities among assemblages of vegetation that results from underlying processes; these processes lead to variations in species composition and dominance that define the groups. Classification can be applied to identify characteristic vegetation assemblages, and these assemblages can be used to assess past conditions and monitor future shifts in vegetation.
Coastal marshes are affected by external forces over a wide range of spatial and temporal scales, and unique processes act to control vegetation assemblages of the Texas Chenier Plain. The system is classified as micro-tidal, defined as a system where the difference between high and low tide is less than 2 m (Walker 2005). The occurrence of strong winter cold fronts and frequent tropical storm events leads to irregular flooding of the coastal marshes via wind generated tides (Li et al. 2011), which generally have a greater impact on the marsh vegetation than astronomical tides (cf. Kim et al. 2011). Factors related to tidal inundation are not the only variables influencing marsh function, an increase in human population of 35 % from 2000 to 2010 (Census Bureau 2010) in the Houston metropolitan area has led to land use change. Moreover, land use change has resulted in a 10–25 % increase in parcelization of coastal prairie along Galveston Bay (Wilkins et al. 2000) which has led to reductions in connectivity of marsh hydrology. The ability to identify characteristic marsh vegetation assemblages along the Texas Chenier Plain is desirable to help assess ecosystem services and monitor marsh loss that has been increasing over the last century (Kennish 2001; Cahoon 2006; Cahoon et al. 2006; FitzGerald et al. 2008; Gedan et al. 2009; Ravens et al. 2009). The marshes along the Texas Gulf Coast, specifically those found in the Galveston Bay system, provide fisheries, recreation, agriculture land, and provide a protective buffer from storm surges. The population of the greater Houston metropolitan area is approximately 6.8 million people and the coastal system is essential to their society and economy (Yoskowitz et al. 2012).
Past studies conducted along the Chenier Plain of Louisiana have investigated the plant communities, their structure, and their dominant species (Chabreck 1972; Visser et al. 1999, 2000). Visser et al. (1999, 2000) used TWINSPAN to group major vegetation assemblages along the Louisiana Chenier Plain. Their study used helicopter transects at 3 km intervals to identify species abundance at a 0.8 km spacing along each transect. Visser et al. (2000) identified seven vegetation assemblages at a regional scale that reflected the dominant vegetation of the Louisiana Chenier Plain. While it is reasonable to expect that the coastal marshes of the Louisiana Chenier Plain are similar to those found along the east Texas Chenier Plain, no comparable studies in Texas exist. White et al. (1985), using the National Wetlands Inventory (NWI), classified the coastal Texas marshes as part of a study characterizing the submerged lands of Texas; broadly their study addressed the spatial distribution of the dominant vegetation zones of the Galveston Bay system. However, their study did not identify specific vegetation assemblages within the Galveston Bay system and was primarily conducted to delineate broad wetland boundaries along the Gulf of Mexico.
There are many approaches to identify tidal marsh vegetation assemblages; one such approach relies strictly on vegetation composition and dominance without considering controlling environmental processes (Lausi and Feoli 1979; Visser et al. 2000; Little et al. 2010). Another approach relies on abiotic factors, such as salinity, inundation, topographic features and elevation, in concert with floristic composition to identify vegetation assemblages (Hickey and Bruce 2010; Kim et al. 2010). Both approaches are needed and, in fact, work well when used together, classifying dominant vegetation in tandem with measurements of the abiotic factors (salinity for example) that enable these species to become dominant. An example of such an integration of both classification and ordination on wetland vegetation can be found in Kim et al. (2010), who used surface elevation and distance from shoreline or tidal creeks to examine how topographic factors influenced marsh vegetation composition and distribution using nonmetric multidimensional scaling (NMDS). Salinity regimes, however, can be highly variable, and in our study, we use a classification based on dominant vegetation only.
In this study, we objectively classified the dominant vegetation assemblages of the East Bay of Galveston Bay, a portion of the Texas Chenier Plain, using quantitative techniques and then compared our findings to Visser et al. (2000) and their classification of the coastal marshes along the Chenier Plain of Louisiana.
Methods
Study Site
The area of focus for our study is along the East Bay of Galveston Bay in Chambers County, TX, the seventh largest bay-estuary-lagoon system in the United States (White et al. 1985). Galveston Bay is composed of four individual bays: West Bay, Galveston Bay, Trinity Bay, and East Bay. Galveston Bay is protected from the rest of the Gulf of Mexico by a peninsula and barrier-island complex (White et al. 1985; White and Tremblay 1995) consisting of the Bolivar peninsula, Galveston Island and Follets Island, which provides protection from the open Gulf by reducing energy from wave action. Galveston Bay contains roughly 105,500 hectares of wetlands including coastal marshes.
The East Bay marks the westernmost terminus of the Gulf Coast Chenier Plains, a series of stranded beach ridges composed of sediment transported from the Mississippi River (Gosselink 1978; Augustinus 1989; Chabreck et al. 1989; Visser et al. 2000). The East Bay is largely protected by the Bolivar peninsula and its waters vary in depth from about 1.2 to 2.4 m (Lester 2002) and the bottom is formed of fine mud and shell (White et al. 1985). Tidal inflow occurs in East Bay via Rollover Pass and Bolivar Roads. Freshwater inputs come from the Trinity River in Trinity Bay and through bayou sources and precipitation. The salinity levels of the East Bay vary based on freshwater input but range from 5 ppt to 15 ppt (White et al. 1985).
The marshes along East Bay consist of intermediate to brackish marshes with some transitional and tidal freshwater marshes intermixed throughout the study site. The soil structure along East Bay is of alkaline and saline, clayey and loamy soils of the Harris-Veston-Ijam association (Crout 1976) which are a complex of mollisols (1–25 %) and entisols (60–76 %). Vegetation most common to the East Bay includes Spartina patens (Ait.) Muhl., Spartina spartinae (Trin.) Merr. Ex Hitchc., Distichlis spicata (L.) Greene, Paspalum vaginatum Sw., Schoenoplectus americanus (Pers.) Volkart ex Schinz & R. Keller, and Bolboschoenus robustus (Pursh) Soják (Johnson 2011). Marsh Hay Cord Grass (S. patens), is the dominant marsh grass found along the East Bay and the Texas coast in general (Stutzenbaker 1999). S. patens is typical in brackish marshes where it grows in dense mat like stands (Brewer and Grace 1990) owing to its ability to tolerate a wide range of conditions (Broome et al. 1995).
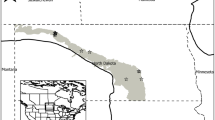
Our study area includes both brackish and intermediate marsh types of the Texas Chenier Plain. Vegetation surveys were conducted at two locations along the East Bay: Canada Ranch and Anahuac National Wildlife Refuge (NWR) (Fig. 1). The study areas along the Texas Chenier Plain are approximately 50 to 200 km from the Louisiana Chenier Plain and the study sites of Visser et al. (2000).

Study locations in Chambers County Texas along the East Bay of Galveston Bay in black. Inset map shows the Gulf Coast Chenier Plain. Visser et al.’s. (2000) study area included much of the Louisiana Chenier Plain, from the Louisiana/Texas border to Vermillion Bay Louisiana. The Gulf Coast Chenier Plain is shaded in gray
Canada Ranch
Canada Ranch is a commercial cow-calf ranch in Chambers County, Texas. The ranch lies approximately 6 km from the shores of East Bay and 4 km from Trinity Bay. Although the ranch is appreciably inland from the 2 bays, it is influenced by tidal waters from both. Gordy Marsh and Lone Oak Bayou contribute tidal water out of Trinity Bay. As a result of damage incurred during Hurricane Ike (September, 2008), the Robinson Lake channel allows tidal water to flow north out of the East Bay onto the ranch. The Canada Ranch marsh vegetation is managed for cattle grazing, primarily through burning and herbicidal application.
Anahuac National Wildlife Refuge
Anahuac NWR consists of 13,759 hectares of coastal marshes and prairies that are managed for the benefit of neotropical migratory birds and waterfowl by the United States Fish and Wildlife Service (USFWS). The refuge is located in the southeast portion of Chambers County, Texas, approximately 2 km from the eastern boundary of the Canada Ranch. Anahuac NWR is bordered on the south by the East Bay and is bisected through the middle by Oyster Bayou which empties into the bay. USFWS uses a variety of methods to manage its marshes including a controlled burning rotation, water level adjustment, and grazing. The refuge manages salinity targets through the use of structural marsh management and aims to keep the water-level within 5 cm of the marsh elevation (Walther personal communication). The brackish and intermediate marshes found in the southern portion of the refuge were chosen for this study.
Though we did not measure salinity during our floristic sampling, the USFWS, at Anahuac NWR, randomly sampled surface salinity during the same time that we collected our data, with measurements ranging between 8 and 14 ppt during the two sampling days.
Data Collection
Sample plots were established to capture the variability of the marsh vegetation typical along the Texas Chenier Plain. Preliminary site visits provided geolocated photographs that were used to aid in the classification of a 2008 Landsat 5 TM scene (Path 25 Row 40) into five marsh zones which were used to stratify the sampling. We employed the program ENVI (ITT 2009) to perform unsupervised classification using an ISODATA classifier. The output spectral classes were assigned to each of the five marsh zones (high marsh, mid marsh, low marsh, forested and unvegetated). 135 coordinates were selected randomly based on the proportion of the area that occurred in each marsh zone to stratify the samples (high marsh 45, mid marsh 75, low marsh 11, forested 3, and unvegetated 1). Forested and unvegetated plots were included to account for marsh species found in Chinese Tallow (Triadica sebifera) stands and potentially unvegetated salt pans respectively.
Each plot was sampled to record plants in a 1 m × 1 m quadrat sampling frame sub-divided into 25, 20 cm × 20 cm subunits. If a species was present in a subunit, it was noted and the total of a given species in a sample ranged from 0 to 25 representing relative dominance of vegetation at a given plot. All plots were sampled between May 21st, 2010 and July 16th, 2010 to minimize variation in seasonal marsh phenology. Sixty two plots were sampled at the Canada Ranch and 73 plots were sampled at Anahuac NWR. Nomenclature follows Stutzenbaker (1999) with additional nomenclature from Godfrey and Wooten (1981a, b).
Elevation measurements were extracted from a Light Detecting and Ranging (LiDAR) dataset (1.4 m ground sample distance and +/− 18.5 cm vertical accuracy in open terrain) collected across Chambers County in 2006 by Sanborn Mapping Company, Inc. for the Texas Water Development Board (TWDB) and the Federal Emergency Management Agency (FEMA). Each sample plot was assigned the elevation of the LiDAR point that fell closest to it.
Statistical Analysis
We used hierarchical agglomerative cluster analysis, a technique for defining groups based on underlying or latent structure (Gauch and Whittaker 1981), to describe vegetation groups. Analyses were conducted using PC-ORD software (McCune and Mefford 1999) with Euclidian distance paired with Ward’s linkage to construct a similarity matrix (McCune and Grace 2002). Indicator species analysis (ISA) was conducted using PC-ORD (McCune and Mefford 1999) to determine the degree of association between a species and groups identified in the cluster analysis. Relative dominance of indicator species was also calculated for each quadrat and the mean relative dominance was determined for each assemblage. We included an additional “ecologically informed” (EI) grouping level, consisting of seven groups, in the analysis which was based on conditions observed qualitatively during data collection and a review of relevant literature; this grouping level improved explanatory power of the assemblages and was the product of a non-linear trimming of the cluster dendrogram. The group mean p values were used to evaluate the appropriateness of the grouping levels (McCune and Grace 2002; Abella et al. 2012). The naming convention used to identify the assemblages was based on the species in the group with the highest occurrence. Finally, we used Multi-Response Permutation Procedures (MRPP) using PC-ORD software (McCune and Mefford 1999) to test if the groups identified in the classification procedure were statistically different from each other. All quadrats were analyzed and all species were included in the analysis.
Results
Thirty Five species were identified at the East Bay marshes (Table 1). The cluster analysis, aided by ISA (Table 2) and MRPP, identified seven statistically different groups (p < 0.01) retaining approximately 70 % of the original information creating ecologically meaningful vegetation assemblages. Group mean p values from 5000 runs of a Monte Carlo test were used to investigate the best grouping levels, where the smallest mean p value indicated the most representative grouping level (Table 3).
Multi-Response Permutation Procedures produce effect and test statistics for each grouping level (Table 3). The effect statistic shows how much homogeneity occurs within an assemblage, and McCune and Grace (2002) suggest that an effect value greater than 0.30 is good for ecological data. The EI grouping level had an effect statistic of 0.33 while grouping level 4 had an effect statistic of 0.27. The test statistic indicates how much separation between groups occurs in an assemblage; the more negative the value the more separation there is between groups. Our results demonstrated that groups 5–10 had strong within group homogeneity and high between group separation. We choose to continue our analysis with the EI grouping level due to enhanced explanatory power and strong statistical support of the identified vegetation assemblages. We discuss the seven vegetation assemblages below in relationship to environmental conditions that were observed qualitatively during data collection. The environmental variables were not tested statistically in this study.
Group one consisted of 22 plots dominated by the species Paspalum vaginatum. Other species were present and the P. vaginatum group was distributed equally between the Canada Ranch (12 plots) and Anahuac NWR (ten plots). At the Canada Ranch, P. vaginatum plots occurred at elevations above 2.00 m and were clustered near the northwest part of the ranch, while at Anahuac NWR the P. vaginatum plots were located at elevations less than 0.50 m and were spread across the refuge.
Group two consisted of six plots dominated by Schoenoplectus americanus. Anahuac NWR contained five of the plots. Canada Ranch contained one plot, this plot occurred east of the Robinson lake channel where little management activity has occurred since the bridge was damaged during Hurricane Ike. The plots consisted of nearly homogenous stands of Sch. americanus but there were some occurrence of S. patens and Bolboschoenus robustus. All plots occurred at elevations less than 1.00 m.
Group three, mixed vegetation sites, consisted of 20 plots with no single dominant species present and with no strong indicator species present (sees Table 2). The distribution of plots shows considerable variability with plots occurring in all marsh types, ranging in elevation from 0.20 m to 2.90 m, and observed conditions ranging from dry salt pans to inundated low marsh. Grazed and burned conditions were also noted at mixed vegetation sites.
Group four consisted of nine plots dominated by Spartina spartinae. S. spartinae, Gulf Cordgrass, is a marsh grass occurring at higher elevations in intermediate to saline marshes. S. spartinae dominated all plots in this group, nearly exclusively, with very few other species present. Three S. spartinae plots occurred at Anahuac NWR, all three along an elevated ridge, at elevations of approximately 1.00 m and classified as high marsh sites. At the Canada Ranch, six S. spartinae plots were sampled all occurring as high marsh sites except for one plot which occurred on an elevated ridge near the Gordy Marsh tidally influenced area of Canada Ranch. Canada Ranch S. spartinae plots range in elevation from 0.50 m to 1.10 m.
Group five consisted of five high marsh plots all clustered near the northwest portion of the Canada Ranch. The group contained low growing herbs such as Diodia virginiana, and shrub species including Croton sp., Borrichia frutescens as well as the grasses S. spartinae and P. vaginatum. The elevation ranged from 1.00 m to 2.20 m at this group level.
Group six consisted of 17 plots dominated by Distichlis spicata. In many cases, D. spicata was accompanied by S. patens and B. robustus but the D. spicata group contained twelve different species occurring in at least one plot. Elevation ranged from −0.03 m on the edge of a flooded pond, to 1.00 m on an elevated ridge.
Group seven consisted of 56 plots dominated by S. patens. The S. patens group was distributed across the entire study area of both the Canada Ranch and Anahuac NWR and was the dominant marsh species observed during field sampling occurring under a wide range of conditions.
Discussion
Gulf Coast Chenier Plain Vegetation Assemblages
Most investigations of Chenier Plain marshes of the Gulf of Mexico Coast have been conducted in Louisiana (Chabreck 1972; Johnson et al. 1985; Visser et al. 1999, 2000; Gabrey and Afton 2001). To our knowledge, there has been no previous research identifying and detailing the vegetation assemblages of the Texas Chenier Plain. Our study is the first to use quantitative methods to identify marsh vegetation assemblages of the western Chenier Plain of east Texas. Specifically, we used hierarchical cluster analysis to identify seven statistically significant vegetation assemblages common along the Texas Chenier Plain.
Results of our study suggest that the Chenier Plain of Texas is qualitatively similar to the Chenier Plain assemblages of Louisiana described by Visser et al. (2000) (Table 4). The seven assemblages represent different marsh vegetation types. When comparing the vegetation assemblages to the Louisiana assemblages described by Visser et al. (2000), the S. patens, and S. americanus groups corresponded with the Louisiana Mesohaline Wiregrass group, D. spicata corresponded to the Louisiana Mesohaline Mixture group, S. spartinae corresponded to the Louisiana Oligohaline Wiregrass group, and P. vaginatum corresponded with the Louisiana Oligohaline Paspalum group. Both our Mixed group and D. virginiana were not comparable to any of the Louisiana assemblages identified in Visser et al. (2000) because they are fine scale nested groups that were likely undetectable in Visser’s helicopter transects.
Visser et al. (2000) noted that over 51 % of the Louisiana classified marshes were oligohaline marsh groups dominated by S. patens; our results are qualitatively similar (Table 4). The most common vegetation assemblage in our study was S. patens, with 42 % of the plots classified as this assemblage. Not only was S. patens the dominant assemblage, but the species also occurred at least once in 68 % of the total sampled plots. S. patens, found in brackish marshes, is generally located above the mean high water line because it is less able to efficiently oxygenate its rhizomes under frequently waterlogged and inundated conditions (Bertness 1991b). Because this study found such large homogeneous stands of S. patens, we postulate that the marshes of the East Bay are generally above mean high water and that inundation depth is low at tidally influenced locations, occurring usually during spring high tides or during storm events. Broome et al. (1995) proposed that S. patens is able to tolerate a wide range of environmental conditions, but Bertness (1991a) found that despite its tolerance of variable conditions, its distribution limits in the low marshes were constrained by physical conditions (e.g. frequent inundation by saline water and waterlogged soil). At higher elevations S. patens is a weaker competitor and is displaced by competitive dominants such as Juncus species (Emery et al. 2001) which we found along the East Bay.
D. spicata is a perennial grass of brackish to saline marshes that is capable of rapidly colonizing disturbed patches (Bertness 1991a). D. spicata is a competitively inferior species and although it can rapidly dominate a disturbed site, it is usually displaced by S. patens as conditions improve restricting D. spicata to generally inhospitable locations (Emery et al. 2001). In our study, D. spicata was found in more frequently inundated locations, implying a more stressful environment, though we acknowledge that while we measured surface elevation we did not measure inundation depth and that inundation is based on qualitative observations at our sampled plots. The Louisiana Mesohaline Mixture group of Visser et al. (2000) was the only category containing D. spicata as a dominant with S. patens and matched up well with our D. spicata group.
P. vaginatum is a grass found in brackish to fresh marshes usually in wet soils with low salinity levels, generally less than 3.5 ppt (Stutzenbaker 1999). The species was identified as falling into the Louisiana Oligohaline Paspalum group of Visser et al. (2000) with the lowest occurrence in their study. We identified the P. vaginatum group occurring in 16 % of the sampled marsh plots. The P. vaginatum assemblage was co-dominated by S. patens. P. vaginatum is a desired grass for grazing purposes and is managed for, by burning, at the Canada Ranch due to its high protein content for cattle likely resulting in the higher occurrence in our study.
The hierarchical cluster analysis captured the floristic similarity of vegetation assemblages that are a result of underlying physical processes occurring in the coastal marshes and expressed through changes in species dominance and composition. In other studies, along with vegetation competition, elevation and inundation frequency are generally hypothesized to be the primary environmental gradients responsible for the variability in marsh structure (Redfield 1972; Adam 1990; Casanova and Brock 2000; Mitsch and Gosselink 2000; Emery et al. 2001). It is very likely that these same conditions are responsible for the composition and dominance of the marsh in our study, although this relationship was not explicitly tested.
An understanding of vegetation composition and dominance can be used to enhance long-term coastal marsh management and conservation efforts. For example, the 2012 Louisiana Comprehensive Master Plan for a Sustainable Coast integrates a number of modules into their coastal model including a vegetation module that requires vegetation composition as an input parameter (CPRA 2012). Vegetation composition is a critical component of surface hydrology models, as well as habitat suitability models, and can be used in protection, planning, and restoration projects (Visser et al. 2012).
Texas Chenier Plain Vegetation Dynamics
Our study of coastal marshes and their related vegetation zones have focused nearly exclusively on marshes designated as salt marshes, where the limits of marsh vegetation are delimited by the mean high water line. Few salt marshes exist along the Chenier Plain because salinity levels rarely exceed 18 ppt (Visser et al. 2000). In our study, the marshes were typically classified as brackish to intermediate, as opposed to true salt marsh. Salinity regime is controlled through complex coastal dynamics that vary through space and time, including freshwater input and elevation (Silvestri et al. 2005), and is likely only one component involved in organization of coastal marsh vegetation assemblages. Brewer and Grace (1990) compared the diversity of marsh species between Texas and Georgia salt marshes and showed that complex processes, such as irregular flooding, may be contributing to the difference in diversity and structure of the marshes and ultimately the factors determining the patterns of marsh zonation. Brackish marshes along the Gulf Coast are less influenced by mean high water but rather are affected by a combination of irregular tidal inundation, precipitation (Kunza and Pennings 2008), and the influence of wind driven flooding of the marshes (Johnson 2011). This phenomenon has received less attention in the literature and may be important to understand the differences between marshes as recently shown for salt marshes in Denmark (Kim et al. 2011). Kim et al. (2011) employed similar methods as our study while investigating the vegetation floristics of the Danish salt marshes.
Most coastal marshes along the Gulf of Mexico and the Atlantic coast contain Spartina alterniflora as the dominant vegetation species (Kirwan et al. 2009) due to its ability to oxygenate the soil environment through oxygen transport by aboveground tissues to the rhizomes (Teal and Kanwisher 1966). S. alterniflora, however, is conspicuously absent from all plots sampled along the East Bay. This may be due in part to the irregular inundation regime experienced at the study site. Kunza and Pennings (2008) note that the lower marsh zones along the coast of Texas commonly consist of S. alterniflora and one or two other species. Bertness (1991b) shows through experimental plot manipulation that under less frequent inundation, S. alterniflora is competitively displaced by S. patens, and we suggest this may explain its absence at our study sites.
Implications and Future Research
Coastal marshes are some of the most productive ecosystems worldwide and provide a large number of ecosystem services (Costanza et al. 1997). According to the Millennium Ecosystem Assessment (MEA 2005), greater than one third of the world’s population now lives in the coastal zone and depend on these ecosystem services. Wetland loss along the Gulf of Mexico has been documented as a result of many factors. Subsidence has been extensive near the Houston metropolitan area because of oil, gas, and fluid extraction (White et al. 2002; Morton et al. 2006) in addition to compactional subsidence and geosynclinal downwarping (White and Tremblay 1995; Ravens et al. 2009). Urbanization has led to direct marsh loss through dredging, filling, and construction activities (Kennish 2001). Hydrological change has also occurred owing to levee and canal construction which restricts sediment inflow and increases salinity levels leading to physiological stress and often death of marsh vegetation (Kennish 2001). All of the aforementioned changes and marsh loss in the northern Gulf of Mexico have been occurring at rates faster than anywhere else in the United States (White and Tremblay 1995). To understand the complex vegetation assemblages and the ecosystem services that they provide, observations of the system must be made at appropriate scales (Day et al. 2008), especially in the context of climate change. The concept of ecosystem services, a term coined by Ehrlich and Ehrlich (1981), is, as Peterson et al. (2010) noted, a concept that was developed to teach us that ecosystems provide service to us, and that the function of the ecosystem, whether understood or not, is valuable to humanity. Our study provides a landscape-level analysis of vegetation patterns along the Texas Chenier Plain that may be used as a floristic benchmark to assess changes in vegetation dominance and composition. In many cases, it is hypothesized that coastal marsh vegetation are expected to migrate towards higher elevation as sea-level rises (Donnelly and Bertness 2001) but due to rapid urbanization and fragmentation of coastal areas their path may be limited and restricted to available space (Feagin et al. 2010), which has been decreasing. Due to the high population levels of the Houston metropolitan area near the study sites and their dependence on the ecosystem services provided, it is critical to gain a better understanding of the vegetation patterns of Texas Chenier Plain marshes so that landowners and land managers may better manage and conserve this system and its resources through science based decision making.
We acknowledge that our study is limited by the lack of quantitative measurements of environmental gradients and assume that the observed marsh composition is the result of these unmeasured environmental variables. Future research in the region should be directed towards identifying specific processes, such as hydrologic regimes, that impact the observed marsh patterns. Specifically, future research should focus on how vegetation composition and dominance vary as a result of inundation caused by wind driven versus astronomical tide. Despite these limitations our results begin to characterize the vegetation assemblages of the east Texas Chenier Plain, which is information that can be used to help evaluate marsh change in response to climatic and human driven pressures to the system.
References
Abella SR, Prengaman KA, Embrey TM, Schmid SM, Newton AC, Merkler DJ (2012) A hierarchical analysis of vegetation on a Mojave Desert landscape, USA. J Arid Environ 78:135–143
Adam P (1990) Saltmarsh ecology. Cambridge University Press, Cambridge
Augustinus PGEF (1989) Cheniers and chenier plains: a general introduction. Mar Geol 90:219–229
Barbier EB (2011) Progress and challenges in valuing coastal and marine ecosystem services. Rev Environ Econ Policy 6:1–19
Bertness MD (1991a) Interspecific interactions among high marsh perennials in a New England salt marsh. Ecology 72:125–137
Bertness MD (1991b) Zonation of Spartina patens and Spartina alterniflora in New England salt marsh. Ecology 72:138–148
Brewer JS, Grace JB (1990) Plant community structure in an oligohaline tidal marsh. Vegetatio 90:93–107
Broome SW, Mendelssohn IA, Mckee KL (1995) Relative growth of Spartina patens (Ait.) Muhl. and Scirpus olneyi gray occuring in a mixed stand as affected by salinity and flooding depth. Wetlands 15:20–30
Cahoon DR (2006) A review of major storm impacts on coastal wetland elevations. Estuar Coasts 29:889–898
Cahoon DR, Hensel PF, Spencer T, Reed DJ, McKee KL, Saintilan N (2006) Coastal wetland vulnerability to relative sea-level rise: wetland elevation trends and process controls. In: Verhoeven JTA, Beltman B, Bobbink R, Whigham DF (eds) Wetlands and natural resource management. Springer, Berlin, pp 271–292
Casanova MT, Brock MA (2000) How do depth, duration and frequency of flooding influence the establishment of wetland plant communities? Plant Ecol 147:237–250
Census Bureau (2010) Census of population, public law 94–171. US Census Bureau, Washington
Chabreck RH (1972) Vegetation, water, and soil characteristics of the Louisiana coastal region. Baton Rouge, LA
Chabreck RH, Joanen T, Paulus SL (1989) Southern coastal marshes and lakes. In: Smith LM, Pederson RL, Kaminski RM (eds) Habitat management for migrating and wintering waterfowl in North America. Texas Tech University Press, Lubbock
Costanza R, D’Agrosa C, de Groot R, Farber S, Grasso M, Hannon B, Limburg K, Naeem S, O’Neill RV, Paruelo J, Raskin RG, Sutton P, van den Belt M (1997) The value of the world’s ecosystem services and natural capital. Nature 387:253–260
CPRA (2012) Louisiana’s comprehensive master plan for a sustainable coast. CRPA Baton Rouge, LA
Crout JD (1976) Soil survey of Chambers County, Texas
Day JW, Christian RR, Boesch DF, Yanez-Arancibia A, Morris JT, Twilley RR, Naylor LA, Schaffner L, Stevenson C (2008) Consequences of climate change on the ecogeomorphology of coastal wetlands. Estuar Coasts 31:477–491
Deegan LA, Johnson DS, Warren RS, Peterson BJ, Fleeger JW, Fagherazzi S, Wolheim WM (2012) Coastal eutrophication as a driver of salt marsh loss. Nature 490:388–392
Donnelly JP, Bertness MD (2001) Rapid shoreward encroachment of salt marsh cordgrass in response to accelerated sea-level rise. Proc Natl Acad Sci U S A 98:14218–14223
Ehrlich PR, Ehrlich A (1981) Extinction: the causes and consequences of the disappearance of species. Random House, New York
Emery NC, Ewanchuk PJ, Bertness MD (2001) Competition and salt-marsh plant zonation: stress tolerators may be dominant competitors. Ecology 82:2471–2485
Feagin RA, Martinez ML, Mendoza-Gonzalez G, Costanza R (2010) Salt marsh zonal migration and ecosystem service change in response to global sea level rise: a case study from an urban region. Ecol Soc 15:14
FitzGerald DM, Fenster MS, Argow BA, Buynevich IV (2008) Coastal impacts due to sea-level rise. Ann Rev Earth Planet Sci 36:601–647
Gabrey SW, Afton AD (2001) Plant community composition and biomass in Gulf Coast Chenier Plain marshes: responses to winter burning and structural marsh management. Environ Manag 27:281–293
Gauch HG, Whittaker RH (1981) Hierarchical classification of community data. J Ecol 69:537–557
Gedan KB, Silliman BR, Bertness MD (2009) Centuries of human-driven change in salt marsh ecosystems. Ann Rev Mar Sci 1:117–141
Godfrey RK, Wooten JW (1981a) Aquatic and wetland plants of southeastern united states dicotyledons. University of Georgia Press, Athens
Godfrey RK, Wooten JW (1981b) Aquatic and wetland plants of southeastern united states monocotyledons. University of Georgia Press, Athens
Gosselink JG (1978) An ecological characterization study of the Chenier Plain coastal ecosystem of Louisiana and Texas. Slidell FWS/OBS-78/9–78/11
Hickey D, Bruce E (2010) Examining tidal inundation and salt marsh vegetation distribution patterns using spatial analysis (Botany Bay, Australia). J Coast Res 26:94–102
ITT (2009) ITT visual information solutions; environment for visualizing images. Boulder, CO
Johnson JS (2011) Changing vegetation dynamics of a coastal salt marsh: Galveston Bay, Texas. Thesis Texas A&M University, College Station
Johnson WB, Sasser CE, Gosselink JG (1985) Succession of vegetation in an evolving river delta, Atchafalaya Bay, Louisiana. J Ecol 73:973
Kennish MJ (2001) Coastal salt marsh systems in the U.S.: a review of anthropogenic impacts. J Coast Res 17:731–748
Kim D, Cairns DM, Bartholdy J (2010) Environmental controls on multiscale spatial patterns of salt marsh vegetation. Phys Geogr 31:58–78
Kim D, Cairns DM, Bartholdy J (2011) Wind-driven sea-level variation influences dynamics of salt marsh vegetation. Ann Assoc Am Geogr 101:231–248
Kirwan ML, Guntenspergen GR, Morris JT (2009) Latitudinal trends in Spartina alterniflora productivity and the response of coastal marshes to global change. Glob Chang Biol 15:1982–1989
Koch EW, Barbier EB, Silliman BR, Reed DJ, Perillo GME, Hacker SD, Granek EF, Primavera JH, Muthiga N, Polasky S, Halpern BS, Kennedy CJ, Kappel CV, Wolanski E (2009) Non-linearity in ecosystem services: temporal and spatial variability in coastal protection. Front Ecol Environ 7:29–37
Kunza AE, Pennings SC (2008) Patterns of plant diversity in Georgia and Texas salt marshes. Estuar Coasts 31:673–681
Lausi D, Feoli E (1979) Hierarchical classification of European salt marsh vegetation based on numerical methods. Vegetatio 39:171–184
Lester J (2002) The state of the bay: a characterization of the Glaveston Bay ecosystem. 2nd edition. Galveston Bay National Estuary Program, Webster
Li C, Roberts HH, Stone GW, Weeks E, Luo Y (2011) Wind surge and saltwater intrusion in Atchafalaya Bay during onshore winds prior to cold front passage. Hydrobiologia 658:27–39
Little AM, Guntenspergen GR, Allen FH (2010) Conceptual hierarchical modeling to describe wetland plant community organization. Wetlands 30:55–65
McCune B, Grace J (2002) Analysis of ecological communities. Mjm Software Design
McCune B, Mefford MJ (1999) PC-ORD for Windows, multivariate analysis of ecological data. MjM Software, Gleneden Beach
MEA (2005) Ecosystems and human well-being: current state and trends, volume 1. Island Press, Washington
Mitsch WJ, Gosselink JG (2000) Wetlands, 3rd edn. John Wiley and Sons, New York
Morton RA, Bernier JC, Barras JA (2006) Evidence of regional subsidence and associated interior wetland loss induced by hydrocarbon production, Gulf Coast region, USA. Environ Geol 50:261–274
Nicholls R, Wong PP, Burkett VR, Codignotto JO, Hay JE, McLean RF, Ragoonaden S, Woodroffe CD (2007) Coastal systems and low-lying areas. Climate change 2007: impacts, adaptations and vulnerability. In: Parry ML, Canziani OF, Palutikof JP, van der Linden PJ, Hanson CE (eds) Contirbution of working group II to the fourth assessment report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge
Parris A, Bromirski V, Burkett VR, Cayan D, Culver M, Hall JW, Horton R, Knuutri K, Moss R, Obeysekera J, Sallenger A, Weiss J (2012) Global sea level rise scenarios for the US National Climate Assessment. NOAA Tech Memo. OAR CPO-1
Peterson MJ, Hall DM, Feldpausch-Parker AM, Peterson TR (2010) Obscuring ecosystem function with application of the ecosystem services concept. Conserv Biol 24:113–119
Ravens TM, Thomas RC, Roberts KA, Santschi PH (2009) Causes of salt marsh erosion in Galveston Bay, Texas. J Coast Res 25:265–272
Redfield AC (1972) Development of a New England salt marsh. Ecol Monogr 42:201–237
Reed DJ (1995) The response of coastal marshes to sea-level rise: survival or submergence? Surf Process Landf 20:39–48
Silvestri S, Defina A, Marani M (2005) Tidal regime, salinity and salt marsh plant zonation. Estuarine Coastal Shelf Sci 62:119–130
Stutzenbaker CD (1999) Aquatic and wetland plants of the western Gulf Coast. Texas Parks and Wildlife, Austin
Teal JM, Kanwisher JW (1966) Gas transport in the marsh grass, Spartina alterniflora. J Exp Bot 17:355–361
Visser JM, Sasser CE, Chabreck RH, Linscombe R (1999) Long-term vegetation change in Louisiana tidal marshes, 1968–1992. Wetlands 19:168–175
Visser JM, Sasser CE, Linscombe RG, Chabreck RH (2000) Marsh vegetation types of the Chenier Plain, Louisiana, USA. Estuaries 23:318–327
Visser JM, Duke-Sylvester S, Broussard W, Carter J (2012) LAVegMod: Louisiana’s comprehensive master plan for a sustainable coast. Coastal Protection and Restoration Authority of Louisiana. Baton Rouge, LA
Walker HJ (2005) Ice-bordered coasts. In: Schwartz ML (ed) Encyclopedia of coastal science. Springer, Netherlands, pp 542–545
White WA, Tremblay TA (1995) Submergence of wetlands as a result of human-induced subsidence and faulting along the upper Texas Gulf-Coast. J Coast Res 11:788–807
White WA, Calnan TR, Morton RA, Kimble RS, Littleton TG, McGowen JH, Nance HS, Schmedes KE (1985) Submerged lands of Texas Galveston-Houston area: sediments, geochemistry, benthic macroinvertebrates, and associated wetlands. Bureau of Economic Geology, Austin
White WA, Morton RA, Holmes CW (2002) A comparison of factors controlling sedimentation rates and wetland loss in fluvial-deltaic systems, Texas gulf coast. Geomorphology 44:47–66
Wilkins N, Brown RD, Conner RJ, Engle J, Gilliland C, Hays A, Slack RD, Steinbach DW (2000) Fragmented lands: changing land ownership in Texas. Texas A&M Agriculture Program
Yoskowitz D, Carollo C, Beseres-Pollack J, Welder K, Santos C, Francis J (2012) Assessment of changing ecosystem services provided by marsh habitat in the Galveston Bay region. p. 75. Harte Research Institute
Acknowledgments
This research was funded through a Texas Sea Grant Minigrant to DC and CH. We would like to thank Eddie Gray and Bill Gammell for sharing their local knowledge and providing valuable insight into the East Bay marshes. We would like to thank Leroy Ezer for providing us access to the Canada Ranch. We are also grateful to Patrick Walther and the USFWS for providing us access and assistance at Anahuac NWR. We gratefully acknowledge two anonymous reviewers for providing constructive comments on an earlier version of this paper.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Johnson, J.S., Cairns, D.M. & Houser, C. Coastal Marsh Vegetation Assemblages of Galveston Bay: Insights for the East Texas Chenier Plain. Wetlands 33, 861–870 (2013). https://doi.org/10.1007/s13157-013-0443-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13157-013-0443-8