
Abstract
The importance of hypoxia inducible factor (HIF) as the master regulator of hypoxic responses is well established. Oxygen-dependent prolyl hydroxylase domain enzymes (PHDs) negatively regulate HIF directing it to the path of degradation under normoxia and are, consequently, attractive therapeutic targets. Inhibition of PHDs might upregulate beneficial HIF-mediated processes. In this study, we have examined the efficacy of PHD inhibitor ethyl 3,4-dihydroxy benzoate (EDHB) in affording protection against hypoxia-induced oxidative damage in L6 myoblast cells. L6 cells were exposed to hypoxia (0.5 % O2) after preconditioning with EDHB for different times. Levels of HIF-1α, oxidative stress and antioxidant status were measured after hypoxia exposure. Preconditioning with EDHB significantly improved cellular viability, and the diminished levels of protein oxidation and malondialdehyde indicated a decrease in oxidative stress when exposed to hypoxia. EDHB treatment also conferred enhanced anti-oxidant status, as there was an increase in the levels of glutathione and antioxidant enzymes like superoxide dismutase and glutathione peroxidase. Further, augmentation of the levels of HIF-1α boosted protein expression of antioxidative enzyme heme-oxygenase I. There was enhanced expression of metallothioneins which also have antioxidant, anti-inflammatory properties. These results thus accentuate the potential cytoprotective efficacy of EDHB against hypoxia-induced oxidative damage.
Similar content being viewed by others


Introduction
Oxygen is the regulator of various metabolic processes in the body. Oxygen homeostasis thus needs to be tightly regulated to meet the requirement for critical oxygen-dependent processes. The condition of hypoxia occurs when availability or delivery of oxygen decreases below the required level for sustaining various processes at cellular level or at tissue level. Hypoxia enhances the generation of reactive oxygen species (ROS), which regulate the cellular response to low oxygen tension. As a part of various metabolic processes, cells continuously produce ROS which are neutralized by the antioxidant defense system which includes both enzymatic and non-enzymatic antioxidants. During high altitude exposure, hypoxic stress induces an imbalance between the oxidant and antioxidant systems due to high ROS levels [24]. The antioxidant system, including enzymatic antioxidants like superoxide dismutase (SOD), glutathione peroxidase (GPx) and glutathione s-transferase (GST) and non-enzymatics like reduced glutathione (GSH), tries to maintain the balance between the production and elimination of ROS levels, but, under severe cellular oxidative stress, these cannot provide complete protection, thus resulting in damage to lipids, proteins and DNA [1, 4, 25, 29].
The cellular responses to hypoxia are complex and involve the regulation of various signaling pathways and transcription factors. There are various mechanisms to sense, adapt and respond at varying oxygen levels [31]. Regulation by a family of transcription factors called hypoxia inducible factor-1 (HIF-1) is one of the important cellular responses to low oxygen concentration. HIF-1 is a heterodimer consisting of two subunits, one of which is a constitutively expressed β subunit and the other is α subunit. HIF-1α is continuously synthesized and rapidly degraded by the ubiquitosomal pathway under normoxia and stabilized under hypoxic conditions. The alpha subunit of HIF-1 then combines with the β subunit and binds to hypoxia-responsive elements in target genes, along with its co-activators, and elicits various cellular responses. HIF-1 is regulated by certain oxygenases which have been identified as key oxygen sensors, a family of O2-dependent prolyl hydroxylase domain enzymes (PHD). Under normoxia, in the presence of oxygen, ferrous ion and 2-oxoglutatrate, these enzymes hydroxylate specific proline residues of HIF-1 leading to its degradation by the ubiquitosomal pathway [41]. During hypoxia, due to decreased oxygen availability, the process of proline hydroxylation is constrained because of inhibition of PHD enzymes resulting in stabilization of HIF-1, thus activating its target genes involved in compensation of hypoxia, including EPO (erythropoetin), VEGF (vascular endothelial growth factor), and HO-1 (heme-oxygenase-1). Recent studies by Yang et al. [38, 39] have shown the protective role of hypoxia preconditioning with PHD inhibitors by significantly reducing the cell death in C6 glioma cells, suggesting involvement of HIF-1 in the development of tolerance to brain injury. Hypoxic preconditioning is the process which makes a tissue more tolerant to subsequent lethal conditions like hypoxia and ischemia [9, 12]. Preconditioning with PHD inhibitor ethyl 3,4-dihydroxy benzoate (EDHB) has been shown to have a cytoprotective effect against oxidative stress in primary astrocytes, suggesting a role of HIF in maintaining oxidative status [6]. EDHB is an analogue of 2-oxoglutarate and thus a competitive inhibitor of PHDs. It is known as protocatechuic acid and is present in plant foods such as olives, roselle, du-zhong, and white grape wine [21]. It has been reported to have antioxidant [40], cardioprotective [33], neuroprotective [22], antimicrobial [28], anti-inflammatory and myoprotective activity [32], as well as anti-ulcer activity. Therefore, we proposed to explore the modulatory role of EDHB as a hypoxic preconditioning agent in the amelioration of hypoxia-mediated damage in the myoblast cell line.
Materials and methods
Chemicals
All chemicals were purchased from Sigma (St. Louis, USA) and SRL. Antibodies were purchased from Santa Cruz Biotech (Santa Cruz, CA, USA).
Cell line
L6 (rat skeletal muscle myoblasts) cell line were obtained from NCCS, Pune (India), grown and maintained as myoblasts in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10 % fetal bovine serum (FBS), 0.1 g/l streptomycin, gentamycin and ampicillin, 2.2 g/l sodium bicarbonate and kept in an incubator with 5 % CO2 at 37 °C.
Hypoxia exposure studies and cell viability assay
MTT [3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide tetrazolium] assay was used to determine the cytotoxic effect of hypoxic exposures. It measures the activity of enzymes that reduce MTT or related dyes to formazan dyes, giving a purple color and thus allowing the assessing the viability (cell counting) and the proliferation of cells (cell culture assays). In brief, after washing with PBS, cells were trypsinized, centrifuged at 2,500 rpm for 10 min, and seeded in 96-well plates. After 70–80 % confluence, the cells were exposed to different hypoxic environments by varying O2 concentration, i.e., 0.5, 1, 3 % in incubator at 37 °C for different time intervals. After each exposure, 50 µl MTT (2 mg/ml) was added to each well and incubated for 3 h at 37 °C. Afterwards, the media were decanted off from plates and formazan crystals were dissolved in 200 µl dimethyl sulfoxide. Absorbance was measured at 595 nm. The optimum concentration of O2 was found to be 0.5 %. Further experiments were done at this O2 concentration.
Drug response studies with EDHB
EDHB at the concentrations of 500 and 1,000 µM was added, when the myoblast culture became 70–80 % confluent and exposed to hypoxia for different time durations, i.e., 12, 24 and 48 h at 0.5 % O2 concentration. EDHB was prepared in DMSO and diluted to the required concentration in incomplete media. Morphometric analysis of cells was carried out using a Phase contrast microscope.
HIF estimation by ELISA
HIF-1α protein concentration in cell lysate was determined using sandwich enzyme-linked immunoassay (Quantikine R&D systems, Minneapolis, MN, USA) according to the manufacturer’s protocol, and measured on a microplate reader (Fluostar OMEGA; BMG Labtech).
Biochemical analysis
For biochemical analysis, cells were trypsinized after washing with PBS and centrifuged at 2,500 rpm for 10 min. The supernatant was discarded, the pellet was resuspended in 1× lysis buffer (50 mM Tris pH 7.4, 300 mM sodium chloride, 10 %v/V Glycerol, 3 mM EDTA, 1 mM MgCl2, 20 mM sodium orthovanadate, 25 mM sodium fluoride, 1 %v/v Triton X-100) and incubated on ice for 20 min. This was followed by centrifugation at 10,000 rpm for 15 min at 4 °C. The supernatant was stored at −80 °C for further analysis. Protein concentration was determined by Lowry’s method [23].
Reactive oxygen species estimation
Reactive oxygen species (ROS) such as superoxide and hydrogen peroxide are continually produced during metabolic processes. ROS generation is normally counter-balanced by the action of antioxidant enzymes and other redox molecules. However, excess ROS can lead to cellular injury in the form of damaged DNA, lipids and proteins. ROS generation by cells can be detected by using 2′,7′-dichlorfluorescein-diacetate (DCFH-DA) [3]. The detection is based on the conversion of nonfluorescent DCFH-DA to the highly fluoresecent compound 2′,7′-dichlorfluorescein (DCF) [18]. Briefly, cells were plated in 96-well black tissue culture plates. After exposure for different durations, 10 µl DCFH-DA (50 µM) was added to each well and incubated at 37 °C. 2′,7′-dichlorofluoroscein formation was determined fluorimetrically after 30 min using Fluostar OMEGA (BMG Labtech) at an excitation of 488 nm and emission at 525 nm. The fluorescent intensity parallels the amount of ROS formed.
Malondialdehyde (MDA)
Reaction of thiobarbituric acid with MDA results in formation of thiobarbituric acid reactive substances (TBARS), which are used to measure the MDA level, which is used as an index of lipid peroxidation [30]. Briefly, 100 μl cell lysate was mixed with 300 μl tricarboxylic acid (TCA 20 %) and then reacted with 750 µl thiobarbituric acid (0.67 %) for 1 h at 80 °C. Thiobarbiturate was used as standard and OD was measured at 531 nm. The levels of lipid peroxides were expressed as MDA nmol/mg protein.
Protein oxidation
Protein oxidation was measured by determining the carbonyl groups after derivitization of proteins with dinitrophenyl hydrazine (DNPH) [19]. Briefly, 100 μl cell lysate was incubated with 100 μl 10 mM DNPH in 2 M HCl for 60 min at 50 °C. Protein was then precipitated using 20 % TCA and the untreated DNPH was removed by centrifugation at 14,000g for 10 min. The pellet was resuspended in 1 M NaOH and the absorbance was measured at 450 nm.
Enzymatic and non-enzymatic antioxidants
Reduced glutathione was measured by the method of Kum Talt et al. [17]. GSH is oxidized by 5, 5′dithiobis (2-nitrobenzoic acid) (DTNB) to give GSSG with the formation of 5-thio 2-nitrobenzoic acid (TNB). Briefly, cell lysate was precipitated with diluted (1:2) metaphosphoric precipitating the reagent (3.34 gm metaphosphoric acid, 0.4 gm EDTA, 60 gm NaCl in 200 ml distilled water). Then, after centrifuging at 8,000 rpm for 15 min. 250 µl supernatant was mixed with 1 ml of 0.3 M sodium phosphate solution and 125 μl DTNB (80 mg in 1 % sodium citrate soln). The rate of formation of TNB was followed at 412 nm. Activity of glutathione peroxidase (GPx, EC 1.11.1.9) was determined using commercially available kits (Randox, UK) as per the manufacturer’s instructions. Superoxide dismutase (SOD, EC 1.15.1.1) was determined using the protocol described by Kakkar et al. [16]. Briefly, 100 µl sample was mixed with 1.5 ml SOD buffer (50 mM Tris–cacodylate buffer, pH 8.2), 300 µl nitroblue tetrazolium chloride (NBT) (1 mM in distilled water), 300 µl Triton X-100 (0.01 %), 800 µl distilled water and 10 µl pyrogallol (60 mM). Absorbance was measured at 540 nm at an interval of 15 s for 180 s. A single unit of enzyme was expressed as 50 % inhibition of NBT reduction/min/mg protein.
Western blotting
Protein expression studies for heme oxygenase-1 [HO-1(H-105), rabbit polyclonal, cat no. SC10789] and metallothioneins [MT (FL-61), rabbit polyclonal, cat no. SC11377] was done by western blotting. Briefly, 50 μg protein of each cell lysate was subjected to SDS-PAGE and electroblotted on to a nitrocellulose membrane (Millipore, USA). The membranes were blocked with 3 % BSA for 2 h, washed with Tris buffer saline with Tween 20 (0.1 %) (TBST) and probed with the respective mouse/rabbit monoclonal/polyclonal antibodies (Santa Cruz) for 3 h. The membranes were washed with TBST and incubated with antimouse/antirabbit-IgG-HRP conjugate (1:40,000) for 2 h. The membranes were then incubated with chemiluminescent substrate (Sigma) and the bands were developed using X-ray films (Kodak, USA). The densities of bands were quantified using LabWorks software (UVP; Bioimaging Systems, UK).
Statistical analysis
All the experiments were performed twice and in triplicates. Data are presented as mean ± SD. Data were analyzed using one–way analysis of variance with post hoc Bonferroni analysis using SPSS, and p < 0.05 was considered as statistically significant.
Results
Effect of hypoxic exposure on cell viability
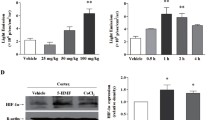
L6 (rat muscle cells) cells were exposed to different O2 concentrations, i.e., 3, 1 and 0.5 % for 0, 12, 24, 48 and 72 h. Cell viability was studied by MTT assay. No significant fall in cell viability was observed at 3 % O2 after 72 h, while there was a little decrease in viability at 1 % O2 after 48 and 72 h. Exposure to 0.5 % O2 led to a reduction in cell viability from 81 % at 24 h to 60 % after 72 h exposure. Since very little effect was observed on cell viability at 1 and 3 %, 0.5 % O2 concentration was taken as the optimum hypoxia exposure condition for further experiments (Fig. 1).

Effect of hypoxia on cell viability for optimization of exposure conditions. L6 muscle cells were exposed to varying O2 concentrations for different times and cell viability was determined by MTT assay. Values are mean ± SD. a = 0.5 % O2 vs. 3 % O2, b = 0.5 % hyp vs. 1 % hyp, c = 1 % hyp vs. 3 % hyp, *Significantly different (p < 0.01)
Effect of EDHB on cell viability
L6 cells were treated with different EDHB concentration, i.e., 500 and 1,000 μM for different durations, i.e., 12, 24 and 48 h. There was a significant increase (p < 0.01) in absorbance of cells treated with 1,000 μM EDHB after 12 h (6 %), 24 h (5 %) and 48 h (20 %) of hypoxic exposure as compared to hypoxia control, indicating enhanced cell viability. Cells treated with 500 μM EDHB also showed an increase in OD after 24 h (5 %), and 48 h (6 %) respectively. Morphometric analysis of cells by phase contrast microscope also showed similar results, as there were no detrimental changes in morphology or cell count in EDHB-treated cells (Figs. 2, 3).

Effect of EDHB on cell viability by MTT assay for different times at 0.5 % exposure. Values are mean ± SD. #Significantly different with respect to hypoxia control, *significantly different (p < 0.01)

Morphometric analysis of cell viability of L6 cell after 0.5 % hypoxic exposure at different times by phase contrast microscopy. There were no detrimental changes in morphology or cell count in EDHB-treated cells
Effect of EDHB on HIF-1α
HIF-1 functions as a master regulator of hypoxia responsive genes. Under normoxia, PHDs hydroxylate HIF resulting in degradation of HIF. In contrast, under hypoxic conditions, prolyl hydroxylation is suppressed resulting in the stabilization of HIF-1α. Exposure to hypoxia (0.5 % O2) resulted in an increase in HIF-1α level at all time points (12, 24 and 48 h) as compared to normoxia control. Preconditioning with EDHB further elevated HIF-1α protein with/without hypoxia. EDHB at 500 µM concentration resulted in maximum rise in levels of HIF-1 after 24 h (1.7 fold) even under normoxia, which was further augmented under hypoxic exposure (Fig. 4).

Effect of EDHB on HIF levels estimated by ELISA. After 0.5 % hypoxic exposure for different times, HIF levels were estimated in cell lysate. A marked increase in the level of HIF was observed in the cells treated with EDHB after hypoxia exposure. Values are mean ± SD. #Significantly different with respect to normoxia control (p < 0.01), ##significantly different with respect to normoxia control (p < 0.05), *significantly different with respect to hypoxia control (p < 0.01), **significantly different with respect to hypoxia control (p < 0.05). NC normoxia control, HC hypoxia control, ND1 normoxia EDHB 500 μM, HD1 hypoxia EDHB 500 μM, ND2 normoxia EDHB 1,000 μM, HD2 hypoxia EDHB 1,000 μM
Effect of EDHB on oxidative status under hypoxic condition
ROS levels
ROS was measured by determining the fluorescein DCF moiety released in the media of exposed and control cells. Higher ROS levels were observed in EDHB-treated cells at all time points as compared to the respective controls (Fig. 5). ROS generation is normally counter-balanced by the action of antioxidant enzymes and other redox molecules, and there is evidence which suggests that ROS may act as intracellular signaling molecules participating in the processes of growth, differentiation, cellular apoptosis and gene expression.

EDHB supplementation reduce oxidative stress. a Levels of ROS was determined by measuring the fluorescein DCF. An increase in ROS levels was observed in drug groups under hypoxic conditions. b MDA is used as an indicator of lipid peroxidation. MDA levels were estimated by measuring thiobarbituric acid reactive substances. There was decrease in the levels of MDA after hypoxia exposure in EDHB-treated cells. c PO was measured as the carbonyl content after protein derivitization by DNPH. Reduction in PO levels was observed in EDHB-treated cells as compared to controls. Values are mean ± SD. ##Significantly different with respect to normoxia control (p < 0.05), #significantly different with respect to normoxia control (p < 0.01), *significantly different with respect to hypoxia control (p < 0.01), **significantly different with respect to hypoxia control (p < 0.05). NC normoxia control, HC hypoxia control, ND1 normoxia EDHB 500 μM, HD1 hypoxia EDHB 500 μM, ND2 normoxia EDHB 1,000 μM, HD2 hypoxia EDHB 1,000 μM
Malondialdehyde (MDA)
ROS degrades polyunsaturated lipids by forming malondialdehyde. This is a reactive aldehyde and most frequently used as an indicator of lipid peroxidation, which is measured by the extent of peroxide generation from free radical mechanisms and removal by the antioxidant system. Products of lipid peroxidation like thiobarbituric acid are thus used to measure the extent of peroxidation. At 0.5 % hypoxic exposure, a significant (p < 0.01) rise in MDA levels was seen in the control hypoxia group at different time points and the maximum was observed after 48 h. In the cells treated with 500 µM EDHB, there was a fall in the levels of MDA under hypoxic conditions as compared to hypoxia control at all time points (Fig. 5). Treatment with 1,000 µM EDHB further reduced MDA levels (1.5 times) on hypoxic exposure. Despite the increase in ROS, there was a significant reduction in MDA by EDHB under hypoxia.
Protein oxidation
After derivitization of proteins with DNPH, carbonyl groups were estimated as the extent of protein oxidation (PO). Exposure of proteins to reactive oxygen leads to modification in the protein structure which then impinges on cellular metabolism. In our study, there was a decrease in protein carbonyl content in drug-treated groups, indicating protection of cells against oxidative damage. There was a significant increase (p < 0.01) in protein oxidation in untreated hypoxia-exposed cells after 12 (11.6 %), 24 (15 %), and 48 (40 %) h as compared to normoxia control, while an appreciable (p < 0.01) decrease was observed by EDHB treatment at all time durations. Treatment with 500 µM EDHB resulted in 12.45 and 50 % decreases in PO after 24 and 48 h, respectively, as compared to hypoxia control. Treatment with 1,000 µM EDHB further reduced the PO by 19.33, 30.2 and 69 % after 12, 24 and 48 h, respectively (Fig. 5).
GSH status
Glutathione directly scavenges ROS and forms GSSG with the help of GPx and thus protects cells from oxidative damage. Therefore, an increase in GSH is indicative of enhanced antioxidant status that helps in protection. An increase in GSH levels was observed on hypoxia exposure as compared to normoxia control, which was further boosted by EDHB under normoxia as well as hypoxia. Cells treated with 1,000 and 500 µM EDHB showed much higher GSH levels (61 and 51 %, respectively, p < 0.01) even after 48 h exposure as compared to hypoxia control groups, indicating enhanced antioxidative status, thus providing sufficient protection to the cells against oxidative damage (Fig. 6).

Augmentation of antioxidant status by EDHB. a Increased levels of GSH in EDHB-treated groups, indicative of enhanced anti-oxidant status thus helps in protection against hypoxia-induced damage. b GPx inhibits the formation of free radicals by reducing peroxides to water. Elevated levels of GPx in EDHB-treated groups signifies reduced oxidative stress. c A higher level of SOD in EDHB-treated groups indicates a potential defense system against ROS, as SOD provides the first line of defence against oxidative stress. Values are mean ± SD. #Significantly different with respect to normoxia control (p < 0.01), ##significantly different with respect to normoxia control (p < 0.05), *significantly different with respect to hypoxia control (p < 0.01). NC normoxia control, HC hypoxia control, ND1 normoxia EDHB 500 μM, HD1 hypoxia EDHB 500 μM, ND2 normoxia EDHB 1,000 μM, HD2 hypoxia EDHB 1,000 μM
GPx activity
GPx enzyme catalyses the reduction of H2O2 and organic peroxides (R–O–O–H) to water and the corresponding stable alcohol by using glutathione as reducing agent, thus inhibiting the formation of free radicals. Therefore, increase in activity of GPx is indicative of enhanced anti oxidative activity. Significant increase in levels of GPx was observed in EDHB-treated cells under normoxia as compared to control. After 0.5 % hypoxic exposure there was appreciable (p < 0.01) rise in GPx levels by 500 µM EDHB at all-time points which was boosted further by 1,000 µM EDHB as compared to hypoxia control (Fig. 6).
Sod
Superoxide dismutase (SOD), is one of the most important antioxidative enzymes, catalyzes the dismutation of the superoxide anion into hydrogen peroxide and molecular oxygen. We observed a significant decrease in SOD in the control group after 24 and 48 h hypoxia (p < 0.01). EDHB treatment resulted in augmentation of SOD levels as compared to controls after hypoxia exposure (Fig. 6). This rise in SOD levels suggests the role of SOD in providing protection against oxidative stress under hypoxic condition in EDHB-treated cells.
Protein expression studies
HO-1 is induced by diverse problems including heme oxidants, hyperoxia and has potent antioxidant and anti-inflammatory properties. Preconditioning with EDHB (500 and 1,000 µM) resulted in increased expression of HO-1 after hypoxic exposure as compared to control. At different time durations there was a difference in the expression of HO-1. Maximum expression was seen in cells treated with 1,000 µM EDHB under hypoxic condition, which was further enhanced after 48 h exposure to hypoxia as compared to hypoxia control. It also confers protection against reactive oxygen intermediates. Thus, increased expression of HO-1 after hypoxic preconditioning with EDHB suggests the promotion of antioxidant and anti-inflammatory status (Fig. 7). Similarly, a progressive increase in expression of MT-1 was observed with time and concentration of EDHB, suggesting the promotion of antioxidant activity by EDHB (Fig. 7). Both doses of EDHB accorded almost similar protection against hypoxia, therefore the lower dose of 500 µM can be used for further studies.

Effect of EDHB on expression of HO-1 and MT-1.was studied by western blot. L6 cells were exposed to 0.5 % hypoxia and protein expression studies were done in cell lysate. Blots were normalized with β-actin as loading control. ##Significantly different with respect to normoxia control (p < 0.05), *significantly different with respect to hypoxia control (p < 0.01), **significantly different with respect to hypoxia control (p < 0.05). NC normoxia control, HC hypoxia control, ND1 normoxia EDHB 500 μM, HD1 hypoxia EDHB 500 μM, ND2 normoxia EDHB 1,000 μM, HD2 hypoxia EDHB 1,000 μM
Discussion
The present study was designed to investigate the efficacy of PHD inhibitor EDHB in alleviating the deleterious effect of hypoxia-mediated oxidative injury in L6 myoblast cell line. There are various enzymatic and non-enzymatic defense mechanisms against reactive oxygen species. Balance between oxidant and antioxidant levels determines the extent of protection of cells from oxidative stress [20, 35]. Increases in the concentration of free radicals can cause damage at various levels of cellular organisation, thus leading to conditions of oxidative stress. We observed appreciable increases in ROS level after hypoxic exposure and also in EDHB-treated groups with/without hypoxia with time. Parallel to this, there was a time-dependent rise in MDA level subsequent to hypoxia in the control as well as in EDHB-treated cells, though the increase was less pronounced in treated groups as compared to untreated hypoxia control. Hypoxia resulted in increases in protein oxidation with time whereas the opposite was observed after EDHB treatment. Notwithstanding the elevation in ROS, the diminished levels of MDA and protein carbonyls in EDHB-treated groups as compared to the control hypoxic group indicated protective efficacy of EDHB against oxidative damage. These results concurred with the improved cell viability as was observed by pre-treatment with EDHB, further confirming the protection accorded by EDHB against hypoxia-induced oxidative injury. The increased ROS might have significance as signaling molecules as also reported by other researchers [7]. MDA and PO, the indicators of oxidative stress in cells, damage membranes, enzymes, etc. [8]. Due to their high abundance and rapid reaction rates, proteins are the major target for oxidative damage [13]. Earlier studies have shown that activated oxygen has the potential to react with all the amino acids [11]. Carbonyl groups can also be generated by the reaction of amino acids with the product of the reaction between ROS and lipids or carbohydrates leading to loss of protein function [2, 34].
To explore the efficacy of EDHB against oxidative stress, we examined the levels of various antioxidant markers. GSH, which is one of the most important endogenous antioxidants, is a tripeptide thiol and acts as an intracellular antioxidant to reduce peroxides by supplying its sulfhydryl group. Glutathione with the help of GPx directly scavenges ROS and forms GSSG and thus protects cells from oxidative damage. In the present study, a significant increase in GSH levels was observed with time in the groups treated with different concentrations of EDHB indicating improved antioxidant status. SOD is known to provide the first line of defence against oxidative stress. There was a decrease in SOD levels in cells exposed to hypoxia whereas the increase in SOD with time in EDHB-treated cells on hypoxia exposure indicates an increase in the inhibition of the formation of hydroxyl ions from hydrogen peroxide, thus providing protection against oxidative stress [14, 26]. Glutathione peroxidase is the enzyme involved in the reduction of H2O2 to H2O by oxidizing glutathione (GSH). Glutathione peroxidase prevents the formation of hydroxyl radicals by the reduction of hydrogen peroxide and hydroperoxides (formed from fatty acids that can initiate lipid peroxidation) into water and diatomic oxygen, thereby effectively removing the toxic peroxides from living cells [27]. In support of this, in our study, elevated levels of GPx with EDHB revealed greater protection against oxidative stress.
Heme oxygenase plays an important role in the free heme-mediated oxidative process. Heme oxygenase is the rate limiting enzyme in the degradation of heme and plays an important role in defense mechanisms. HO-1 is induced by its substrate heme, and its induction occurs as an adaptive and beneficial response against various stimuli like oxidative stress resulting from hypoxia, ischemia or I/R injury. HO-1 has anti-oxidant, anti-inflammatory and cytoprotective functions [10]. An EDHB-mediated increase in HO-1 expression as observed in the present study can be attributed to enhanced levels of HIF, which is the central regulator of various responses under hypoxia [36]. Studies have reported the induction of HIF1 expression by PHD inhibitors and their role in tolerance to oxidative injury in primary astrocytes [5, 6, 37]. Metallothioneins (MT) are small cysteine-rich proteins that play a cytoprotective role in stress situations such as oxidative stress and are potent hydroxyl radical scavengers. The primary determinant of MT protection against oxidative stress is the release of zinc sequestered by MT and its subsequent uptake by plasma membranes, since zinc protects against lipid peroxidation and thereby stabilizes membranes [15]. Enhanced expression of MT by EDHB also indicates an antioxidant property of EDHB. Thus, the improved antioxidant status and HIF-1-mediated induction of HO-1 might be responsible for according protection against oxidative damage, thereby improving the adaptive responses under hypoxic conditions, thus emphasizing the significant cytoprotective role of EDHB in protecting against oxidative stress under hypoxia. These findings provide the basis for the possible use of EDHB for preconditioning against hypoxia-induced cell damage and also as a potent antioxidant.
References
Bailey DM, Davies B, Young IS (2001) Intermittent hypoxic training: implications for lipid peroxidation induced by acute normoxic exercise in active men. Clin Sci (Lond) 101(5):465–475
Berlett BS, Stadtman ER (1997) Protein oxidation in aging, disease, and oxidative stress. J Biol Chem 272(33):20313–20316
Cathcart R, Schwiers E, Ames BN (1983) Detection of picomole levels of hydroperoxides using a fluorescent dichlorofluorescein assay. Anal Biochem 134(1):111–116
Cesaratto L, Vascotto C, Calligaris S, Tell G (2004) The importance of redox state in liver damage. Ann Hepatol 3(3):86–92
Chan DA, Sutphin PD, Denko NC, Giaccia AJ (2002) Role of prolyl hydroxylation in oncogenically stabilized hypoxia-inducible factor-1alpha. J Biol Chem 277(42):40112–40117. doi:10.1074/jbc.M206922200
Chu PW, Beart PM, Jones NM (2010) Preconditioning protects against oxidative injury involving hypoxia-inducible factor-1 and vascular endothelial growth factor in cultured astrocytes. Eur J Pharmacol 633(1):24–32
D’Autreaux B, Toledano MB (2007) ROS as signaling molecules: mechanisms that generate specificity in ROS homeostasis. Nat Rev Mol Cell Biol 8(10):813–824. doi:10.1038/nrm2256
Del Rio D, Stewart AJ, Pellegrini N (2005) A review of recent studies on malondialdehyde as toxic molecule and biological marker of oxidative stress. Nutr Metab Cardiovasc Dis 15(4):316–328. doi:10.1016/j.numecd.2005.05.003
Dirnagl U, Simon RP, Hallenbeck JM (2003) Ischemic tolerance and endogenous neuroprotection. Trends Neurosci 26(5):248–254. doi:10.1016/s0166-2236(03)00071-7
Farombi EO, Surh YJ (2006) Heme oxygenase-1 as a potential therapeutic target for hepatoprotection. J Biochem Mol Biol 39(5):479–491
Garrison WM (1987) Reaction mechanisms in the radiolysis of peptides, polypeptides, and proteins. Chem Rev 87(2):381–398
Gidday JM (2006) Cerebral preconditioning and ischaemic tolerance. Nat Rev Neurosci 7(6):437–448. doi:10.1038/nrn1927
Gracanin M, Hawkins CL, Pattison DI, Davies MJ (2009) Singlet-oxygen-mediated amino acid and protein oxidation: formation of tryptophan peroxides and decomposition products. Free Radic Biol Med 47(1):92–102
Huang CC, Lin TJ, Lu YF, Chen CC, Huang CY, Lin WT (2009) Protective effects of l-arginine supplementation against exhaustive exercise-induced oxidative stress in young rat tissues. Chin J Physiol 52(5):306–315
Jiang L-J, Maret W, Vallee BL (1998) The ATP—metallothionein complex. Proc Natl Acad Sci USA 95(16):9146–9149
Kakkar P, Das B, Viswanathan PN (1984) A modified spectrophotometric assay of superoxide dismutase. Indian J Biochem Biophys 21(2):130–132
Kum-Tatt L, Tan I-K (1974) A new colorimetric method for the determination of glutathione in erythrocytes. Clin Chim Acta 53(2):153–161. doi:10.1016/0009-8981(74)90093-X
LeBel CP, Ischiropoulos H, Bondy SC (1992) Evaluation of the probe 2′,7′-dichlorofluorescin as an indicator of reactive oxygen species formation and oxidative stress. Chem Res Toxicol 5(2):227–231
Levine RL, Garland D, Oliver CN, Amici A, Climent I, Lenz A-G, Ahn B-W, Shaltiel S, Stadtman ER (1990) Determination of carbonyl content in oxidatively modified proteins. Methods Enzymol 186:464–478
Limón-Pacheco J, Gonsebatt ME (2009) The role of antioxidants and antioxidant-related enzymes in protective responses to environmentally induced oxidative stress. Mutat Res Genet Toxicol Environ Mutagen 674(1):137–147
Lin C-Y, Huang C-S, Huang C-Y, Yin M-C (2009) Anticoagulatory, antiinflammatory, and antioxidative effects of protocatechuic acid in diabetic mice. J Agric Food Chem 57(15):6661–6667
Lomb DJ, Desouza LA, Franklin JL, Freeman RS (2009) Prolyl hydroxylase inhibitors depend on extracellular glucose and hypoxia-inducible factor (HIF)-2alpha to inhibit cell death caused by nerve growth factor (NGF) deprivation: evidence that HIF-2alpha has a role in NGF-promoted survival of sympathetic neurons. Mol Pharmacol 75(5):1198–1209. doi:10.1124/mol.108.053157
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ (1951) Protein measurement with the Folin phenol reagent. J Biol Chem 193(1):265–275
Magalhães J, Ascensão A, Soares JM, Ferreira R, Neuparth MJ, Marques F, Duarte JA (2005) Acute and severe hypobaric hypoxia increases oxidative stress and impairs mitochondrial function in mouse skeletal muscle. J Appl Physiol 99(4):1247–1253
Magalhães J, Ascensao A, Viscor G, Soares J, Oliveira J, Marques F, Duarte J (2004) Oxidative stress in humans during and after 4 hours of hypoxia at a simulated altitude of 5500 m. Aviat Sp Environ Med 75(1):16–22
Manna I, Jana K, Samanta PK (2004) Intensive swimming exercise-induced oxidative stress and reproductive dysfunction in male wistar rats: protective role of alpha-tocopherol succinate. Can J Appl Physiol 29(2):172–185
Mazzetti I, Grigolo B, Pulsatelli L, Dolzani P, Silvestri T, Roseti L, Meliconi R, Facchini A (2001) Differential roles of nitric oxide and oxygen radicals in chondrocytes affected by osteoarthritis and rheumatoid arthritis. Clin Sci (Lond) 101(6):593–599
MeRkl R, HRádkoVá I, FIlIp V, ŠMIdRkal J (2010) Antimicrobial and antioxidant properties of phenolic acids alkyl esters. Czech J Food Sci 28(4):275–279
Møller P, Loft S, Lundby C, Olsen NV (2001) Acute hypoxia and hypoxic exercise induce DNA strand breaks and oxidative DNA damage in humans. FASEB J 15(7):1181–1186
Ohkawa H, Ohishi N, Yagi K (1979) Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal Biochem 95(2):351–358
Osorio-Fuentealba C, Valdes JA, Riquelme D, Hidalgo J, Hidalgo C, Carrasco MA (2009) Hypoxia stimulates via separate pathways ERK phosphorylation and NF-kappaB activation in skeletal muscle cells in primary culture. J Appl Physiol 106(4):1301–1310. doi:10.1152/japplphysiol.91224.2008
Philipp S, Cui L, Ludolph B, Kelm M, Schulz R, Cohen MV, Downey JM (2006) Desferoxamine and ethyl-3,4-dihydroxybenzoate protect myocardium by activating NOS and generating mitochondrial ROS. Am J Physiol Heart Circ Physiol 290(1):H450–H457. doi:10.1152/ajpheart.00472.2005
Raphael J, Rivo J, Beeri R, Abedat S, Gozal Y (2004) Mechanisms of Myocardial Ischemic Preconditioning: a potential protective role for HIF-1 in a rabbit model of regional myocardial ischemia. Anesthesiology 101:A717
Shulaev V, Oliver DJ (2006) Metabolic and proteomic markers for oxidative stress. New tools for reactive oxygen species research. Plant Physiol 141(2):367–372
Tirzitis G, Bartosz G (2010) Determination of antiradical and antioxidant activity: basic principles and new insights. Acta Biochim Pol 57:139–142
Wenger RH (2002) Cellular adaptation to hypoxia: O2-sensing protein hydroxylases, hypoxia-inducible transcription factors, and O2-regulated gene expression. FASEB J 16(10):1151–1162. doi:10.1096/fj.01-0944rev
Wright G, Higgin JJ, Raines RT, Steenbergen C, Murphy E (2003) Activation of the prolyl hydroxylase oxygen-sensor results in induction of GLUT1, heme oxygenase-1, and nitric-oxide synthase proteins and confers protection from metabolic inhibition to cardiomyocytes. J Biol Chem 278(22):20235–20239
Yang DI, Chen SD, Yang YT, Ju TC, Xu JM, Hsu CY (2004) Carbamoylating chemoresistance induced by cobalt pretreatment in C6 glioma cells: putative roles of hypoxia-inducible factor-1. Br J Pharmacol 141(6):988–996. doi:10.1038/sj.bjp.0705687
Yang YT, Ju TC, Yang DI (2005) Induction of hypoxia inducible factor-1 attenuates metabolic insults induced by 3-nitropropionic acid in rat C6 glioma cells. J Neurochem 93(3):513–525. doi:10.1111/j.1471-4159.2005.03032.x
Yen W-J, Chang L-W, Duh P-D (2005) Antioxidant activity of peanut seed testa and its antioxidative component, ethyl protocatechuate. Food Sci Technol 38(3):193–200
Yu F, White SB, Zhao Q, Lee FS (2001) HIF-1alpha binding to VHL is regulated by stimulus-sensitive proline hydroxylation. Proc Natl Acad Sci USA 98(17):9630–9635. doi:10.1073/pnas.181341498
Acknowledgments
The research is supported by financial assistance from the Defense Research and Development Organization (DRDO), Government of India. We are grateful to the Director, Dr S.B. Singh, DIPAS, DRDO, India, for providing all the support and facilities for conducting this research. Ms Charu and Ms Gurpreet Kaur are recipient of Senior Research fellowships from the Council for Scientific and Industrial Research (CSIR), Government of India.
Conflict of interest
The authors declare that they have no conflict of interests.
Author information
Authors and Affiliations
Corresponding author
About this article

Cite this article
Nimker, C., Kaur, G., Revo, A. et al. Ethyl 3,4-dihydroxy benzoate, a unique preconditioning agent for alleviating hypoxia-mediated oxidative damage in L6 myoblasts cells. J Physiol Sci 65, 77–87 (2015). https://doi.org/10.1007/s12576-014-0348-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12576-014-0348-1