
Summary
Four new species of Maesa are described and illustrated from the Malesian region: M. inculticola Utteridge and M. pilosa Utteridge are described from Borneo; M. fraseriana Utteridge and M. malayana Utteridge are described from Peninsular Malaysia. Distribution maps and notes on the conservation status and taxonomic affinities of each species are provided.
Similar content being viewed by others


Introduction
The Old World genus Maesa Forssk. comprises approximately 150 species and was last monographed by Mez (1902). The South East Asian members of the genus are being revised for Flora Malesiana with revisions and new taxa published to date in the Philippines (Utteridge & Saunders 2000, 2004) and New Guinea (Sleumer 1987; Utteridge 2000, 2001, 2003). As part of this revisionary process, this paper describes four new species of Maesa from the Sunda shelf area of western Malesia; a key to the genus is in preparation for Flora Malesiana and Flora of Peninsular Malaysia.
Whilst the phylogenetic position of the genus Maesa is now clear (as sister to the genera traditionally placed in the Theophrastaceae, Myrsinaceae and Primulaceae), the family in which Maesa has been placed has changed due to different interpretations of family limits within this group (see Stevens 2001; Angiosperm Phylogeny Group 2009: 116). Traditionally the genus was placed in Myrsinaceae (as a monogeneric subfamily Maesoideae; see Mez 1902), then it was split from the Myrsinaceae into its own family Maesaceae (see Anderberg et al. 2000), and most recently placed in an enlarged Primulaceae (see Angiosperm Phylogeny Group 2009); the last is followed here.
Materials and Methods
Gross morphological observations were made from herbarium material held at A, AAU, K, KEP, L, SAN, and SING (herbarium acronyms follow Thiers 2010). Floral measurements were made from rehydrated material. The description of leaf morphology, particularly venation follows Hickey (1979); two dimensional shapes follow the Systematics Association Committee (1962) terminology. In the descriptions, reproductive shoot leaves, bracts and bracteoles are used when referring to structures on the plagiotropic reproductive shoots; it could be argued that the reproductive shoot leaves are bracts but for descriptive purposes this term refers to the leaves subtending the inflorescences, with the term bracts used for the structures that subtend the pedicels, whilst the bracteoles are on the pedicel and precede the hypanthium (see Prenner et al. 2009 for further discussion). The bracteoles are referred to as pseudo-opposite because they can appear to be opposite each other at the base of the ovary, however species of Maesa never exhibit opposite phyllotaxy, branching etc., and so the bracteole position is one derived from an alternate position.
Two types of indumentum are found in the genus: single-celled hairs of variable length, and irregularly shaped peltate scales (see Caris et al. 2000: 89). The new species described here all lack scales but have combinations of different hair lengths. Descriptive terms of indumentum have a varied, and often extremely ambiguous, usage (for example, see Hewson 1988), and most Maesa species would be described as having pubescent, pilose, hispid or hirsutellous parts (sensu Hewson 1988); note that glabrous is used here to refer to lack of any indumentum (hairs and scales). These descriptive terms are useful to portray the general appearance of hairs on a plant, but are not used here in the descriptions as they do not adequately represent the most informative attribute of the hairs — the length, which is given in the species descriptions, together with the density. This is best observed using a binocular dissecting microscope with a magnification of ×(20 –) 40, but where a species has two different ‘cohorts’ of hair length, e.g., M. pilosa Utteridge, this is easily observed with a loupe.
Specimens were mapped and a conservation assessment of the species undertaken using GIS methods (see Willis et al. 2003; Moat 2007), with the categories of threat following the IUCN guidelines (IUCN 2001). When missing from the label, longitude and latitude were gathered from the GEOnet Names Server: http://earth-info.nga.mil/gns/html/index.html, and from maps including Google Earth; these are shown in square brackets in the specimen list below.
Taxonomic Treatment
Maesa fraseriana Utteridge sp. nov. Maesa sumatranae Scheff. atque M. macrothyrsae Miq. proxima sed ab ambabus speciebus foliis ellipticis ad basin rotundatis vel cuneatis differt. A M. sumatrana etiam pedunculis atque pedicellis indumentum ferentibus differt. A M. macrothyrsa etiam fructibus globosis (nec ellipsoideis) differt. Typus: Malaysia, Pahang, Fraser’s Hill, 3°43'N, 101°45'E, 11 Oct. 1979, fl., Kochummen FRI 11484 (holotypus K!; isotypus KEP!).
http://www.ipni.org/urn:lsid:ipni.org:names:77118904-1
Scrambling shrub, described as woody climber to 20 ft, or small shrub to 6 ft; hairs 0.025 – 0.05 mm long, scales absent. Vegetative shoots hairy. Vegetative shoot leaves laminas elliptic, 6.3 – 7.5 × 23.3 – 30.6 cm, coriaceous, glabrous ad- and abaxially, apex acute-attenuate, base cuneate, margins entire; midrib glabrous ad- and abaxially; secondary veins brochidodromus, 6 – 8 pairs per leaf, glabrous ad- and abaxially; petioles 10.8 – 13.8 mm long, glabrous. Reproductive shoots 8.5 – 18.6 cm long, glabrous; inflorescences produced along entire length of shoot, entire length of shoot with leaves subtending inflorescences. Reproductive shoot leaves not appearing ± clustered; lamina elliptic to ovate, (5 –) 7 – 8.5 (− 13.4) × 2.5 – 3.5 (− 4.4) cm, attaining full size along the entire length of the shoot, coriaceous, glabrous ad- and abaxially, apex acute to attenuate, base cuneate or rounded, margins entire, midrib glabrous ad- and abaxially; secondary veins brochidodromus, 5 – 11 pairs per leaf, glabrous; petiole (6 –) 7.5 – 11.5 (− 17.5) mm long, glabrous or rarely sparsely hairy. Staminate inflorescences lateral (axillary) [only a single inflorescence observed], subtended by a reproductive shoot leaf, compound racemose, branched to 2 orders with c. 1 first order branch; primary axis c. 7 cm long, sparsely hairy; pedicels c. 1.5 mm long, sparsely hairy; bracts triangular, c. 65 mm long, glabrous, margins densely ciliate, apex acute; bracteoles pseudo-opposite at the base of the hypanthium, sometimes connate, triangular, c. 0.6 mm long, glabrous, margins densely ciliate, apex acute. Staminate flowers pentamerous; calyx-lobes broadly ovate-triangular, c. 0.5 × c. 0.8 mm, glabrous, glandular canals not evident, margins densely ciliate, apex acute; corolla c. 1.25 mm long, white; corolla lobes broadly ovate, c. 0.7 × 0.85 mm, glandular canals not evident; anthers c. 0.25 mm long; filaments c. 25 mm long; pistillode (including hypanthium) ellipsoid, c. 0.9 mm long; hypanthium glabrous; vestigial style c. 0.25 mm long; vestigial stigma unlobed. Pistillate inflorescences as staminate inflorescences except racemose or compound racemose, with 1 – 2 first order branches only; primary axis (2.5 –) 7 – 20 cm long, hairy; pedicels 0.9 – 2 mm long hairy to sparsely hairy; bracts 0.6 – 1 mm long; bracteoles 0.6 – 1 mm long. Pistillate flowers as staminate flowers except calyx-lobes (0.6 –) 1 – 1.2 × 0.75 – 1 mm; corolla 1.3 – 2.2 mm long; corolla lobes 0.8 – 1.2 × 1 – 1.3 mm; staminodes 0.3 – 0.7 mm from base of corolla; vestigial anthers 0.2 – 0.3 mm long; vestigial filaments 0.3 – 0.5 mm long; ovary (including hypanthium) 0.9 – 1.4 mm long; style and stigma 0.25 – 0.5 mm long, stigma unlobed. Fruits globose, 2.6 – 4 × (1.9 –) 2.8 – 3.8 mm, green when immature, creamy‐white when mature, glabrous, rarely sparsely hairy at the base; fruit wall 0.2 – 0.4 mm thick. Pedicels 1.2 – 2 mm long; bracteoles pseudo-opposite at the base of the fruit; persistent calyx-lobes not overlapping, appressed. Seeds 5 – 9 per fruit. Figs 1, 2.

Maesa fraseriana. A habit; B corolla detail; C detail of flower, corolla removed (note fused bracteole margins); D detail of flower, corolla removed (note free bracteole margins); E fruit. A, E from Latiff 4105; B – D from Kochummen FRI 11484. drawn by lucy t. smith.

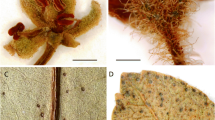
Maesa fraseriana. A detail of infructescence from Julius et al. FRI 73602; B habit. photos: timothy utteridge.

Maesa inculticola. A habit; B detail flowering shoot; C detail of inflorescence; D fruit. A from Yii et al. S 43119; B from Ilias Paie & Yeo Eng Teck S 38353; C, D from George S 40448. drawn by lucy t. smith.

Maesa malayana. A habit; B detail of abaxial leaf surface; C detail of stem; D inflorescence; E flower bud; F flower, corolla removed; G corolla with fully developed stamens; H corolla with staminodes. All from J. F. Maxwell 78 – 203. drawn by lucy t. smith.

Maesa pilosa. A habit; B detail of adaxial leaf surface; C detail of abaxial leaf surface; D detail of stem; E fruit on raceme; F fruit. All from Rantai Jawa et al. S 82868. drawn by lucy t. smith.
distribution. Malaysia: Peninsular Malaysia. Currently known from several collections from the Fraser’s Hill area and a single specimen collected from Ulu Klang. Kiew (1998: 10) notes that several Fraser’s Hill endemics have later been found from the Genting Highlands (between Fraser’s Hill and Ulu Klang), and it is possible that this species is found throughout this part of the central range, but more collections are needed to fill the gap. Map 1.

Distribution of Maesa fraseriana [▲] and M. malayana [●].
specimens examined. malaysia. Pahang: Fraser’s Hill, [3°43'N 101°45'E], 13 Dec. 1930, fl., Deris 22561 (K!); Fraser’s Hill, along road, [3°43'N 101°45'E], 11 Nov. 1976, fl. & fr., Keng et al. 70 (L!); Fraser’s Hill, hill slope opposite morgue close to Glen bungalow, [3°43'N 101°44'30"E], 12 Nov. 1981, fr., Keng et al. T3/72 (SING!); Fraser’s Hill, 4,000 ft, 11 Oct. 1979, fl., Kochummen FRI 11484 (K!); Bukit Frasers, Jalan Taman Bunga, 28 Nov. 1993, fr., Latiff 4105 (K!); Fraser’s Hill, Selangor Residency Bungalow, [3°42'30"N 101°44'20"E], 27 Sept. 1959, fr., M. Shah & Nur 684 (K!); Gap road, Fraser’s Hill, 3,800 ft, 17 April 1955, fl., Purseglove 4130 (K!); Fraser’s Hill, 50 m before Rompin Trail start, 3°43.04'N 101°44.01'E, 1218 m, 26 Oct. 2010 (fr.), Julius et al. FRI 73602 (K!, KEP!, L!, SAN!, SING!); Selangor: Ulu Klang, [3°14'N 101°45'E], 24 Jan. 1966, fr., Hardial & Sidek 502 (K!); gated road below Fraser’s Hill, [3°41'10"N 101°44'50"E], 31 March 1971, fl., Whitmore FRI 15974 (K!, L!).
habitat. Lower montane forest, roadside and forest edge; alt. c. 1,200 m.
conservation status. Endangered (EN B1 ab(iii)). Maesa fraseriana is known from five localities based on ten collections with all the collections from Fraser’s Hill except for a single outlier from Ulu Klang. Fraser’s Hill is a popular resort town in a central position in the Main Range of Peninsular Malaysia, and the area’s flora, and its conservation status, has been recently documented in detail (Kiew 1998). Threats to habitats in the Fraser’s Hill area include road building, resort development and agriculture, and Kiew (1998: 23) notes that within Fraser’s Hill “no area has legal or permanent protection” and “in the long-term, the flora of Fraser’s Hill is under threat”. The extent of occurrence (EOO) of the collections of Maesa fraseriana is 1,000 km2 (including the Ulu Klang specimen) — within the IUCN threshold for Endangered (EN). To date, this species has yet to be collected from the Genting Highlands, an area that is one of the most developed hill resorts in Malaysia and faces similar threats to Fraser’s Hill (Hamilton 1995). Therefore, with the restricted EOO, known from no more than five locations, and the evidence for habitat decline in the Fraser’s Hill area, a rating of EN B1 ab(iii) is assigned here to M. fraseriana (following IUCN 2001).
etymology. This species is named for Fraser’s Hill, the type locality.
notes. Maesa fraseriana is distinct amongst the scrambling species of Maesa in South East Asia in possessing the following combination of character states: small, elliptic leaves (laminas 5 – 13.4 × 2.5 – 4.4 cm), which attain full size along the length of the reproductive shoot; short hairs found only on the inflorescence axis and pedicels; and glabrous, globose fruits.
In the Malay Peninsula there are two scrambling species with which this new species could be confused: Maesa macrothyrsa Miq. and M. sumatrana Scheff, but these species have reproductive shoot leaves that reduce in size toward the distal portion of the shoot, resulting in the reproductive shoots appearing to be a massive terminal inflorescence. In addition, M. fraseriana differs in the sparsely hairy inflorescence axis (all floral parts are densely hairy in M. macrothyrsa), the bracteoles which may be fused (alternate and free in M. macrothyrsa), and, most conspicuously, the globose fruits (ovoid or ellipsoid in M. macrothyrsa). M. fraseriana differs from M. sumatrana in the coriaceous leaves with cuneate to rounded bases (chartaceous with cordate to rounded bases in M. sumatrana), the sparsely hairy inflorescence axis (inflorescence glabrous in M. sumatrana), and the pseudo-opposite bracteoles at the apex of the pedicel which are often fused together (see Fig. 1C; bracteoles alternate or pseudo-opposite and never fused in M. sumatrana).
Kiew (1998) recorded only Maesa ramentacea (Roxb.) A. DC. from Fraser’s Hill. That probably refers to this species (no specimens are cited in Kiew 1998), as all collections of this species were previously determined as M. ramentacea, a name widely used throughout South East Asia for ± glabrous species of Maesa with elliptic leaves that, when strictly applied, refers to a tree species found throughout India, Burma and Thailand and reaching Malaysia in Langkawi.
Maesa inculticola Utteridge sp. nov. M. ramentaceae (Roxb.) A. DC. similis sed planta omnino glabra (nec ramulis inflorescentiae indumentum ferentibus), foliis ellipticis (nec ovatis) inflorescentia non ramosa differt. Maesa sumatranae Scheff. etiam aliquantum similis sed habitu arborescenti nec scandenti et inflorescentiis omnibus axillaribus (numquam terminalibus) differt. Typus: Malaysia, Sarawak, 4th Division, Path to Lambir Hill NP, Ulu Sg. Lebau, 20th mile, Miri/Bin rd, [4°2'N 113°57'E], 2 Nov. 1976, fl., Ilias Paie & Yeo Eng Teck S 38353 (holotypus K!; isotypi KEP, L!, MO, SAN, SAR).
http://www.ipni.org/urn:lsid:ipni.org:names:77118905-1
Shrubs or small trees, to 5 m; hairs absent, scales absent. Vegetative shoots and leaves not recorded. Reproductive shoots length not recorded, glabrous; inflorescences produced along entire length of shoot, with leaves subtending inflorescences. Reproductive shoot leaves lamina elliptic or ± oblong, (4.2 –) 8.3 – 8.9 × (1.4 –) 2.2 – 3.4 cm, chartaceous, glabrous ad- and abaxially, apex acute to attenuate (rarely acuminate), base cuneate, margins entire; midrib ad- and abaxially glabrous; secondary veins brochidodromus, 5 – 7 pairs per leaf, glabrous; petiole 9.5 – 12.5 mm long, glabrous. Staminate inflorescences and flowers not seen. Pistillate inflorescences lateral (axillary), always subtended by a reproductive shoot leaf, racemose, unbranched; primary axis (1.75 –) 4 – 5.75 cm long, glabrous; pedicels 1 – 2 mm long, glabrous; bracts triangular, 0.6 – 0.7 mm long, glabrous, margins entire, apex acute; bracteoles pseudo-opposite at the base of the hypanthium, triangular, 0.5 – 0.75 mm long, glabrous, margins entire, apex acute. Pistillate flowers tetramerous; calyx-lobes broadly triangular, 0.65 – 0.7 × 0.7 – 0.75 mm, glabrous, glandular canals indistinct, margins glabrous, apex rounded; corolla 0.8 – 1 mm long, white; corolla lobes broadly ovate, 0.5 – 0.65 × 0.65 – 0.75 mm, apex rounded, glandular canals 2 per petal; vestigial anthers 0.1 – 0.2 mm long; vestigial filaments 0.25 – 0.35 mm long; pistil (including hypanthium) obconical, 0.65 – 0.75 mm long; hypanthium glabrous, style 0.2 – 0.25 mm long; stigma unlobed. Fruits globose, 2.8 – 3.3 × 3 – 3.25 mm, green to white, glabrous; pedicels 2.2 – 3 mm long; bracteoles pseudo-opposite at base of fruit. Seeds c. 15 per fruit. Fig. 3.
distribution. Malaysia: endemic to the Lambir Hills area of Sarawak. Map 2.

Distribution of Maesa inculticola [▲] and M. pilosa [●].
specimens examined. malaysia. Sarawak: 4th Division [Bintulu Division], Lambir National Park, [4°12'N 113°59'E], 550 m, 27 Sept. 1978, fl. & fr., George S 40448 (E, K!, KEP, L!, SAN, SAR); ibid., Path to Lambir Hills NP, Ulu Sg. Lebau, 20th mile, Miri/Bin rd, [4°2'N 113°57'E], 2 Nov. 1976, fl., Ilias Paie & Yeo Eng Teck S 38353 (K!, KEP, L!, MO, SAN, SAR); ibid., summit of Lambir Hill, Lambir National Park, [4°11'55"N 113°59'50"E], 500 m, 21 April 1981, fr., Yii et al. S 43119 (K!, KEP, L!, SAN, SAR).
habitat. Restricted to heath-kerangas-forest; alt. from c. 500 m.
conservation status. Near Threatened (NT). Maesa inculticola, which is known from three collections from three locations, all in Lambir Hills National Park, has a very small EOO of 3.7 km2, and thus falls into the EOO range of Critically Endangered (CR). However, because the species is found within the protected area of the Lambir Hills NP (which appears to be covered with undisturbed vegetation on the most recent Google Earth imagery), is known from relatively recent collections and is not known to be collected or harvested, a rating of Near Threatened (NT) is more appropriate. This rating may have to be revised to a threat rating if there are threats to the park (such as a change in land use or a development), or if further visits to Lambir Hills NP reveal the population to be declining; an expansion of the NP and an increase in the population may lead to a rating of Least Concern (LC).
etymology. The epithet refers to the restricted habitat requirements of the species. Heath forest in northern Borneo is termed kerangas, a word from the Iban language meaning ‘land that will not grow rice’ and this, interestingly, echoes the use of the phrase ‘loca inculta’ as a term for heathland in Latin (for example, see Simpson 1968); thus, the epithet is derived from the Latin phrase and refers to the species ‘dwelling in’ an area of land unsuitable for cultivation.
notes. Maesa inculticola Utteridge is distinguished by its tree habit, completely glabrous throughout, the relatively small elliptic or ± oblong leaves with entire margins, the relatively short unbranched inflorescences, and, especially, the tetramerous flowers. This new species is unlikely to be confused with other Maesa species in Borneo being the only completely glabrous tree species with elliptic, entire leaves and tetramerous flowers on the island. Specimens of M. inculticola were initially determined as M. ramentacea but, as previously discussed under M. fraseriana, M. ramentacea s.s. is restricted to mainland Asia as far south as Langkawi. Whilst M. ramentacea is absent from Borneo, the name has been widely used to refer to at least two distinct taxa: the new species described here and a commonly collected scrambling/climbing species of Maesa best referred to as M. sumatrana Scheff.
Morphologically, Maesa inculticola differs from M. ramentacea s.s. in the following characters: small tree to 5 m (M. ramentacea can be tree up to 15 m high); completely glabrous throughout (M. ramentacea with a shortly hairy inflorescence axis); leaves elliptic or ± oblong with an acute to attenuate apex (leaves ovate with a long attenuate apex in M. ramentacea); the unbranched inflorescence axis (inflorescence with up to 15 first order branches branched to two orders in M. ramentacea); and the tetramerous flowers (pentamerous in M. ramentacea).
Maesa sumatrana is the earliest available name for the scrambling/climbing species of Maesa found from southern Thailand through Peninsular Malaysia to the northern plains of Sumatra, and throughout Borneo. M. inculticola differs from M. sumatrana in the tree habit (M. sumatrana is a scrambler or climber over forest edges and gaps); the elliptic or ± oblong leaves (leaves ovate in M. sumatrana); the architecture of the reproductive shoots with unbranched inflorescences having subtending leaves along the entire length of the shoot (inflorescences branched to three orders with subtending leaves becoming reduced toward the distal end of reproductive shoots in M. sumatrana to form a massive terminal ‘inflorescence’); and the tetramerous flowers (pentamerous in M. sumatrana).
Maesa malayana Utteridge sp. nov. M. ramentaceae (Roxb.) A. DC. affinis sed pilis brevibus atque longis mixtis (nec brevibus tantum) in omnibus partibus dispositis, pilis in ramulis dense dispositis (nec ramulis glabris) atque in pagina adaxiali laminae etiam praesentibus (nec lamina omnino glabra), et lamina ad marginem serrulata nec integra differt. Typus: Malaysia, Pahang, Tanah Rata, Cameron Highlands, c. 1,600 m, 16 April 1978, fl., J. F. Maxwell 78 – 203 (holotypus AAU!; isotypi L!, SINU).
http://www.ipni.org/urn:lsid:ipni.org:names:77118906-1
Maesa macrophylla sensu Stone (1989a: 283).
Trees to c. 4 m, dbh 20 cm; densely hairy throughout with hairs of two distinct lengths: 0.05 – 0.15 mm long, and 0.4 – 8 mm long, scales absent. Vegetative shoots and leaves not recorded. Reproductive shoots length not recorded, densely hairy; inflorescences produced along entire length of shoot, entire length of shoot with leaves subtending inflorescences. Reproductive shoot leaves not appearing ± clustered; lamina ovate, 9.8 – 11.7 × 3.6 – 4.4 cm, attaining full size along the entire length of the shoot, chartaceous, sparsely hairy ad- and abaxially, apex attenuate, base cuneate-obtuse to rounded, margins seemingly entire but minutely serrulate with 6 – 8 teeth per side; midrib ad- and abaxially densely hairy; secondary veins semicraspedodromus, 7 – 8 pairs per leaf, indumentum as midrib; petiole 13 – 14 mm long, densely hairy. Staminate inflorescences lateral (axillary), always subtended by a reproductive shoot leaf, compound racemose, branched to 2 orders with 14 – 21 first order branches; primary axis 15 – 16.5 cm long, densely hairy, pedicels 1 – 1.6 mm long, hairy with short hairs; bracts triangular, 0.5 mm long, glabrous, margins densely ciliate, apex acute; bracteoles alternate or pseudo-opposite at base of hypanthium, triangular, 0.5 mm long, glabrous, margins densely ciliate, apex acute. Staminate flowers pentamerous; calyx-lobes triangular, 0.5 – 0.65 × 0.7 – 0.8 mm, glabrous, glandular canals 4 – 5 per sepal, margins ciliate, apex acute; corolla 2 – 2.2 mm long; corolla lobes ovate, 1 – 1.25 × 1 – 1.25 mm, apex rounded; glandular canals 5 – 6 per petal; anthers 0.3 – 0.4 mm long; filaments 0.5 – 0.7 mm long; pistillode (including hypanthium) obconical, 0.7 – 0.9 mm long; hypanthium glabrous; vestigial style and stigma 0.3 – 0.45 mm long, vestigial stigma unlobed. Pistillate inflorescences not seen. Pistillate flowers as staminate flowers except corolla 1.5 – 1.65 mm long; corolla lobes ovate, 0.75 – 1 × 1 mm, apex rounded; glandular canals 5 – 6 per petal, staminodes 0.2 mm from base of corolla; vestigial anthers c. 0.2 mm long; vestigial filaments 0.3 – 0.4 mm long. Fruits not seen. Fig. 4.
distribution. Known only from Tanah Rata in the Cameron Highlands. Map 1.
specimen examined. malaysia . Pahang, Tanah Rata, Cameron Highlands, c. 1,600 m, 16 April 1978, fl., J. F. Maxwell 78 – 203 (holotype AAU!; isotypes L!, SINU).
habitat. Collected from ‘partly open margins of evergreen forest along the road’ fide Maxwell 78 – 203.
conservation status. Data Deficient (DD). Maesa malayana is currently only known from a single collection from Tanah Rata in the Cameron Highlands — a tourist resort and agricultural centre with extensive tea plantations and areas of vegetable production (Aiken & Leigh 1992). The Cameron Highlands Wildlife Reserve protected area was created in 1962 and has areas of primary and secondary forest, but urban and agricultural expansion has impacted montane forests in the area (Soh et al. 2006). Until more collections of this species are made, and its habitat preferences understood, a preliminary conservation assessment of data deficient, following IUCN (2001), is given.
etymology. The name refers to the plant’s distribution from Peninsular Malaysia.
notes. A highly distinctive species, Maesa malayana is instantly recognisable on account of the dense covering of hairs of two distinct non-overlapping different length ‘classes’ that cover all parts of the plant, including the adaxial lamina surface, a character which is not commonly found in the genus.
This species is similar to the Peninsular Malaysian species Maesa ramentacea, and to M. pilosa from Borneo (see below). It differs from M. ramentacea in having two different hair length classes throughout (M. ramentacea with very short hairs only); densely hairy branches (M. ramentacea with glabrous branches); and serrulate leaf margins (M. ramentacea with entire margins). M. pilosa is similar to M. malayana in possessing two different hair length classes, but the two species differ in leaf morphology: M. malayana has ovate leaves (M. pilosa: the leaves are ovate-elliptic to narrowly elliptic), which are comparatively smaller in size being only 9.8 – 11.7 × 3.6 – 4.4 cm (M. pilosa: 13 – 22 × 3.5 – 7.8 cm), and having only 7 – 8 pairs of secondary veins per leaf (M. pilosa: 11 – 22).
This species is only known from a single collection, which shows predominately male characteristics, especially the long inflorescence with no fruit set, and staminate flowers with fully formed stamens. Although some flowers have staminodes with vestigial anthers (see Fig. 4H), there is no fruit set, nor flowers remaining on the inflorescence after the corollas have fallen that would indicate the presence of fertile female parts. Utteridge & Saunders (2001) established that the Hong Kong species M. japonica (Thunb.) Moritzi ex Zoll. and M. perlarius (Lour.) Merr. were functionally dioecious with different degrees of sexual dimorphism, but staminate flowers with reduced stamens, as seen in the type collection of M. malayana, was not a morphological combination encountered in that study. Further collections, as well as field observations, are needed to establish the exact nature of the sexuality of M. malayana.
Maesa pilosa Utteridge sp. nov. Maesa procerae B. C. Stone similis sed pilis longis in omnibus partibus plantae dispositis (nec planta omnino glabra) et venis lateralibus foliorum utrinque 11 – 22 (nec 8 – 12 tantum) numero differt. Typus: Malaysia, Sarawak, Genting Mawang, trail to summit of Bukit Entimau, [1°38'N 112°8'E], 600 m, 30 March 2002, fr., Rantai Jawa et al. S 82868 (holotypus K!; isotypi KEP, L, SAR).
http://www.ipni.org/urn:lsid:ipni.org:names:77118907-1
Shrub 3 – 6.1 m, 15 – 20.3 cm girth (collecting notes); densely hairy throughout with hairs of two lengths: hairs 0.1 – 0.25 mm long and 0.4 – 1.2 mm long, scales absent. Vegetative shoots and leaves not recorded. Reproductive shoots length not recorded, densely hairy; inflorescences produced along entire length of shoot, with leaves subtending inflorescences. Reproductive shoot leaves not appearing ± clustered; lamina ovate-elliptic to narrowly elliptic, (13 –) 18.5 – 22 × (3.5 –) 5 – 7.8 cm, attaining full size along the entire length of the shoot, coriaceous, densely hairy, becoming sparsely hairy ad- and abaxially with short and long hairs, apex attenuate to acuminate, acumen 2 – 3 cm long, base rounded to acute, margins serrulate, with 7 – 14 teeth per side; midrib ad- and abaxially hairy, with long hairs; secondary veins semicraspedodromus, 11 – 18 (− 22) pairs per leaf, indumentum as midrib, appearing ± bullate above due to the impressed venation; petiole (11 –) 16 – 22 mm long, densely hairy. Staminate inflorescences and flowers not seen. Pistillate inflorescences at anthesis and flowers not seen. Pistillate inflorescences in fruit lateral (axillary), always subtended by a reproductive shoot leaf, compound racemose, with (0 –) 4 – 7 first order branches only; primary axis (3 –) 5 – 10.5 cm long, densely hairy. Fruits globose, 2 – 3 × 2 – 2.8 mm, brown (distinctly lineate when dry); glabrous; pedicels at fruiting stage 1 – 2.75 mm long; bracteoles pseudo-opposite at base of fruit; seeds 9 – 15 per fruit. Fig. 5.
distribution. Malaysia: Borneo. Endemic to Sarawak and currently only known from three collections. Map 2.
specimens examined. malaysia. Sarawak: Kapit Division, Genting Mawang, trail to summit of Bukit Entimau, Lanjak Entimau Wildlife Sanctuary, Mujok, [1°38'N 112°8'E], 600 m, 30 March 2002, fr., Rantai Jawa et al. S 82868 (K!, KEP, L, SAR); ibid.: Bukit Raya, [2°10'N 113°7'E], c. 400 m, 13 Nov. 1964, fr., Hj. Suib S 22263 (A, BO, K!, KEP, L, SAN, SAR, SING); Samarahan Division: Serian distr., Gunong Penrissen, [1°7'N 110°13'E], 4,000 ft [1,219 m], 29 April 1962 (fr.), Ilias Paie S 16394 (A, B, K!, L!, NB, S, SAR).
habitat. Lowland mixed dipterocarp and sub-montane forests: “open area” Rantai Jawa et al. S 82868, and “primary forest” Ilias Paie S 16394; alt. 400 – c. 1,220 m.
conservation status. Endangered (EN B1ab(iii)). Maesa pilosa is known from only three collections from three localities, the most recent from 2002. Two of the collections were made from protected areas (following IUCN & UNEP 2009): Ilias Paie S 16394 lies within the Gunong Penrissen Nature Reserve (an Indonesian reserve which straddles the Malaysian border), and Rantai Jawa et al. S 82868 from within the Lanjak Entimau Wildlife Sanctuary; however, Hj. Suib S 22263 from Bukit Raya is not in any protected area. The EOO of 3,150 km2 falls within the IUCN threshold of Endangered (EN), although these areas of Sarawak are poorly collected and with further exploration the EOO may increase and the conservation status will then be too high, but with the limited collections known to date, and following IUCN (2001) guidelines to use a precautionary rating in cases of uncertainty, the EN rating is appropriate. In addition, habitat degradation in SE Asia is well documented (e.g., Sodhi et al. 2004), even in protected areas in Borneo (Curran et al. 2004), and the low EOO, low number of locations, and documented habitat degradation give this species a rating of EN B1ab(iii) following IUCN (2001).
etymology. The name refers to the species having distinct hairs.
notes. Maesa pilosa is an elegant and highly distinctive species easily recognised by its ovate-elliptic to narrowly elliptic leaves that have long attenuate apices, the serrulate leaf margins, numerous arching secondary veins, and the hairs of two distinct non-overlapping lengths.
Maesa pilosa is morphologically similar to the Bornean species M. procera B. C. Stone (1989b) especially in the leaf morphology, but differs from that species in the following characters: hairy throughout (M. procera glabrous throughout); larger, ovate-elliptic to narrowly elliptic leaves with more secondary veins (M. procera 9 – 19 × 2 – 3.7 cm, narrowly oblong-lanceolate with 8 – 12 pairs of secondary veins); and the distinctly lineate fruits when dry (M. procera fruits sparsely lineate when dry). Differences between M. pilosa and M. malayana are discussed in the Notes to that species above.
References
Aiken, S. R. & Leigh, C. H. (1992). Vanishing Rain Forests: the Ecological Transition in Malaysia. (Oxford Biogeography series, 5). Clarendon Press, Oxford.
Anderberg, A. A., Ståhl, B. & Källersjö, M. (2000). Maesaceae, a new primuloid family in the order Ericales s.l. Taxon 49: 183 – 187.
Angiosperm Phylogeny Group (APG) (2009). An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. Bot. J. Linn. Soc. 161: 105 – 121.
Caris, P., Ronse de Craene, L. P., Smets, E. & Clinckemaillie, D. (2000). Floral development of three Maesa species, with special emphasis on the position of the genus within Primulales. Ann. Bot. 86: 87 – 97.
Curran, L. M., Trigg, S. N., McDonald, A. K., Astiani, D., Hardiono, Y. M., Siregar, P., Caniago, I. & Kasischke, F. (2004). Lowland forest loss in protected areas of Indonesian Borneo. Science 303: 1000 – 1003.
Hamilton, L. S. (1995). Mountain cloud forest conservation and research: a synopsis. Mountain Res. Developm. 15: 259 – 266.
Hewson, H. J. (1988). Plant Indumentum. A Handbook of Terminology. Bureau of Flora and Fauna, Canberra.
Hickey, L. J. (1979). A revised classification of the architecture of dicotyledonous leaves. In: C. R. Metcalfe & L. Chalk (eds), Anatomy of the Dicotyledons, pp. 25 – 39. Clarendon Press, Oxford.
IUCN (2001). IUCN Red List Categories and Criteria: Version 3.1. IUCN Species Survival Commission. IUCN, Gland & Cambridge.
____ & UNEP (2009). The World Database on Protected Areas (WDPA). UNEP-WCMC, Cambridge.
Kiew, R. (1998). The Seed Plant Flora of Fraser’s Hill, Peninsular Malaysia: with Special Reference to its Conservation Status. Forest Research Institute Malaysia (FRIM), Kuala Lumpur.
Mez, C. (1902). Myrsinaceae. In: A. Engler (ed.), Das Pflanzenreich, 9 (IV. 236): 1 – 437. Wilhelm Engelmann, Berlin.
Moat, J. (2007). Conservation Assessment Tools Extension for ArcView 3.x, version 1.2. GIS Unit, Royal Botanic Gardens, Kew. Available at: http://www.rbgkew.org.uk/gis/cats
Prenner, G., Vergara-Silva, F. & Rudall, P. J. (2009). The key role of morphology in modelling inflorescence architecture. Trends Pl. Sci. 14: 302 – 309.
Simpson, D. P. (1968). Cassell’s New Latin-English English-Latin Dictionary; 5th edition. Cassell, London.
Sleumer, H. (1987). A revision of the genus Maesa Forssk. (Myrsinaceae) in New Guinea, and the Solomon islands. Blumea 32: 39 – 65.
Sodhi, N. S., Koh, L. P., Brook, B. W. & Ng, P. K. L. (2004). Southeast Asian biodiversity; an impending disaster. Trends Ecol. Evol. 19: 654 – 660.
Soh, M. C. K., Sodhi, N. S. & Lim, S. L. H. (2006). High sensitivity of montane bird communities to habitat disturbance in Peninsular Malaysia. Biol. Conserv. 129: 149 – 166.
Stevens, P. F. (2001 onwards). Angiosperm Phylogeny Website. Version 9, June 2008 [and more or less continuously updated since]. http://www.mobot.org/MOBOT/research/APweb/
Stone, B. C. (1989a). Myrsinaceae. In: F. S. P. Ng (ed.), Tree Flora of Malaya 4: 264 – 284. Forest Research Institute Malaysia, Kuala Lumpur.
____ (1989b). New and noteworthy Malesian Myrsinaceae, IV. Two new species in Embelia Burm. f. and Maesa Forssk. from Borneo. Proc. Acad. Nat. Sci. Philadelphia 141: 307 – 311.
Systematics Association Committee (1962). Terminology of simple symmetrical plane shapes (chart 1). Taxon 11: 145 – 156 & 245 – 247.
Thiers, B. (2010 and continuously updated). Index Herbariorum: A Global Directory of Public Herbaria and Associated Staff. New York Botanical Garden’s Virtual Herbarium. http://sweetgum.nybg.org/ih/
Utteridge, T. M. A. (2000). Two new species of Maesa (Myrsinaceae) from Puncak Jaya, New Guinea. Contributions to the flora of Mt Jaya, I. Kew Bull. 55: 443 – 449.
____ (2001). Two new species of Maesa (Maesaceae) from New Guinea. Kew Bull. 56: 677 – 683.
____ (2003). Maesa from New Guinea: a new species and revised description of Maesa beamanii. Contributions to the flora of Mt Jaya, X. Kew Bull. 58: 237 – 241.
____ & Saunders, R. M. K. (2000). Maesa reflexa (Maesaceae): a new species from Mindanao, Philippines. Kew Bull. 55: 981 – 984.
____ & ____ (2001). Sexual dimorphism and functional dioecy in Maesa perlarius and M. japonica (Maesaceae/Myrsinaceae). Biotropica 33 (2): 368 – 374.
____ & ____ (2004). The genus Maesa (Maesaceae) in the Philippines. Bot. J. Linn. Soc. 145: 17 – 43.
Willis, F., Moat, J. & Paton, A. (2003). Defining a role for herbarium data in Red List assessments: a case study of Plectranthus from eastern and southern tropical Africa. Biodivers. Conserv. 12: 1537 – 1552.
Acknowledgements
I thank Lucy T. Smith for the illustrations, Melanie Wilmot-Dear for translating the Latin diagnoses, and Melanie Wilmot-Dear, Rogier de Kok and two reviewers for very useful comments on the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Utteridge, T.M.A. Four new species of Maesa Forssk. (Primulaceae) from Malesia. Kew Bull 67, 367–378 (2012). https://doi.org/10.1007/s12225-012-9383-3
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12225-012-9383-3