
Abstract
Intercellular transport in C4 plant, including maize, is particularly important due to the necessity of efficient exchange of photoassimilates between different types of cells, where specific stages of C4 photosynthesis occur. The transport on the trail: mesophyll—bundle sheath—vascular parenchyma occurs by symplast, through plasmodesmata. The next stage, phloem loading (the transport of photosynthetic products to the companion cell/sieve element complex) belongs to the apoplasmic type, without the participation of plasmodesmata. This work concerns verification of the hypothesis about the impact of low temperature on changes in expression of genes, strictly related to phloem loading process as well as intercellular (symplasmic) transport. For that purpose, in situ hybridization assay for several mRNAs and the determination of changes in the expression level of these transcripts using RT-qPCR technique were conducted. Transcripts related to phloem loading, e.g., sucrose transporters, interactor of proton pump, aquaporins and plasmodesmata-associated genes, coding: TMV-MP30 binding protein and, remorins were selected. Studies were performed on two lines of maize differing in chilling-sensitivity level. Plants were treated with low temperature (day/night): 14/12 and 8/6 °C in two periods: 4 and 28 h. Cold-treatment resulted in a diverse response of tested maize plants, mainly changes in the gene expression under exposition at shock temperature level (6–8 °C) and after prolonged chilling-treatment (28 h). The localization of sucrose transporter and aquaporin transcripts in the thick-walled sieve tubes indicates the potential role of these cells in the phloem loading process. The pronounced changes in expression of plasmodesmata-related transcripts indicate their evident participation in response to cold stress.
Similar content being viewed by others

Introduction
Maize, thermophilic plant originating in subtropical regions, in moderate climate conditions as in Poland is often exposed to cold temperatures, especially in the spring at the beginning of the growing season. It is assumed that the minimum temperatures of the soil during germination of maize seedlings should be above 8 °C. In the next stage, after the germination, temperature should be ca. 15 °C, and the optimum temperature for photosynthesis is about 30 °C (Bennett et al. 1982). The inhibition of maize growth and development is observed at cold in the range of 10–15 °C (moderate chilling). In contrast, cold (2–8 °C) cause irreversible changes which often leads to plant’s death.
Reactions of maize to cold conditions were studied mainly in the aspect of the efficiency of photosynthesis, damages of the photosynthetic apparatus (Miedema 1982; Baker et al. 1983; Fryer et al. 1995; Savitch et al. 2011) and antioxidant enzymes (Kingston-Smith and Foyer 2000). The process of photosynthesis in maize belonging to C4 plants, is characterized by spatial distribution of the individual stages—in the mesophyll cells the primary fixation of CO2 occurs, while in bundle sheath cells decarboxylation of organic acids and restoration of acceptor molecules takes place. This spatial distribution requires efficient transport of photosynthetic metabolites between these cell types to maintain appropriate performance of photosynthesis. The movement of photosynthesis products on the trail mesophyll—bundle sheath—vascular parenchyma occurs through cytoplasmic channel of plasmodesmata (Evert et al. 1996) and is called symplasmic transport. The transport to the complex: companion cell/sieve tube, called phloem loading, occurs by apoplasmic way with the participation of sucrose transporters and proton pump. Argument for the involvement of these proteins in apoplasmic phloem loading is their localization—for instance mRNA of sucrose transporter 1 (SUT1), was predominantly found within sieve tubes in tobacco, potato, and tomato in various tissues (leaves, stems, petioles, roots) (Kühn et al. 1997).
In addition, it was found that transport of water from xylem to sieve tubes generating a high hydrostatic pressure in the phloem may involve aquaporins (Sakurai et al. 2005, 2008). Aquaporins are localized in different tissues, cells and structures (Robinson et al. 1996; Barkla et al. 1999; Sakurai et al. 2008), mainly in areas closely associated with intensive transport of water (more in review: Kjellbom et al. 1999). Additionally, these proteins may be involved in transport of gases and ions, and also in cell signaling or responses to different stresses (Maurel 2007 and references therein).
In C4 plants, low temperature causes inhibition of assimilates translocation, including phloem loading (Wardlaw and Bagnall 1981; Potvin et al. 1984, 1985). For instance, in southern population of grass Echinochloa crus-galli (L.), low night temperature reduced drastically translocation of carbon and increased the turn-over times of the photosynthetic poll (Potvin et al. 1984, 1985). Changes in transport efficiency were associated with decrease of photosynthesis and chlorosis of leaves in this species (Potvin et al. 1985). Similarly, in Sorghum bicolor plants, translocation of photosynthesis products is inhibited by temperatures below 5 °C (Wardlaw and Bagnall 1981).
In maize, as previously demonstrated in our group, the intercellular transport of photoassimilates, as well as phloem loading are inhibited at low temperature (Sowiński et al. 2001, 2003; Bilska and Sowiński 2010). Further, it has been proven using microarray technique (Trzcinska-Danielewicz et al. 2009; Sobkowiak et al. 2014), that among the hundreds of genes whose expression is altered by cold, many were potentially involved in the transport of photosynthetic products in the leaf including genes likely involved in the regulation of the operation of plasmodesmata: TMV-MP30 binding protein and remorins (Ding et al. 1992; Raffaele et al. 2009), as well as genes important in the phloem loading process: sucrose transporters and aquaporins as potential controllers of this process by the regulation of water flow from xylem to phloem (Patrick et al. 2001). However, for majority of maize genes databases provide only ‘provisional’ or ‘putative’ annotations without confirmed function through, for instance, localization of transcripts. The aim of this work is to verify the hypothesis that low temperature may cause changes in expression of genes which function is precisely (with confirmed localization) related to phloem loading process as well as intercellular (symplasmic) transport. For this purpose, in situ hybridization technique was used with confirmation of gene expression changes by RT-qPCR.
Materials and methods
Plant material, growth condition and chilling-treatment periods
For studies chilling-tolerant (CT) S68911 and chilling-sensitive (CS) S160 maize lines (Z. mays spp. indentata, dent) were used. Both lines represent dent kernel type and come from the same gene pool (Stiff Stalk Synthetic). Seeds were germinated in wet sand in darkness at 25 °C. Seedlings were transferred to hydroponic growth system with Knop’s solution supplemented with Hoagland’s micronutrients. Seedlings were grown in a growth chamber using parameters: 14 h/10 h (light/darkness period), irradiance 250 µmol quanta m−2 s−1, temperature: 24 °C/22 °C (day/night), 60–80 % humidity. When the third leaf was fully developed, at the beginning of the light period, plants were transferred to low temperature 14/12 or 8/6 °C (day/night) for either 4 or 28 h (for RT-qPCR) and 4 h (for in situ hybridization). Non-chilled plants were used as control. Each analysis was repeated three times in three independent experiments.
In situ hybridization
Studies were performed at the transmission electron microscopic level to identify the vascular vein cells of a relatively small size (thin-, thick-walled sieve tubes). The material for electron microscopy was fixed according to a standard procedure (PFA + GA/PIPES) and was embedded in acrylic resin LR-White (Sigma). The ultrathin sections (80–100 nm) were obtained using a Leica Ultracut UTC ultramicrotome.
For labeling of probes DIG Oligonucleotide Tailing Kit, 2nd Generation (Roche Applied Science) was chosen, where using terminal deoxynucleotidyl transferase (TdT) and a mixture of nucleotides Dig-11-dUTP/dATP (1/10), digoxigenin at the 3′ end was attached. For detection DIG-probes under electron microscope F(ab ‘) fragment of Sheep-anti-digoxigenin antibody conjugated to 10-nm colloidal gold (Aurion) was used. From 11 antisense DNA oligonucleotide probes with a length of 22–30 nucleotides six probes with confirmed localization were selected for further analysis (Table 1). Probes corresponded to a part of 70-nt microarray probes from Maize Oligonucleotide Array Project, Arizona (ftp.maizegdb.org/MaizeGDB/FTP/arizona_maize_arrays/; Trzcinska-Danielewicz et al. 2009). Two negative controls were applied, in the first one, probe in the sense orientation was used, and in the second one, hybridization without antisense probe was performed. As a positive control β-actin sequence as a probe was used.
To increase membrane permeability to the probe penetration, the ultrathin sections were incubated for 1 min with proteinase K (1 µg/ml). After the prehybridization step, the sections were incubated overnight at 37 °C in the hybridization buffer with DIG-probes at concentration of 100 pmol/ml. Posthybridization washing, detection using sheep-anti-digoxigenin antibody and staining were conducted according to Smoliński et al. (2007). The observations were done using transmission electron microscope (model JEM 1400; JEOL Co., Japan).
Quantitative real-time PCR
For RT-qPCR RNA isolation and purification RNeasy Plant Mini Kit (Qiagen) was used according to the manufacturer’s protocol. A single sample consisted of three pieces of leaf randomly taken. Leaves were homogenized by grinding in liquid nitrogen using sterile mortar and pestle. After isolation and purification of total RNA, measurements of its quantity and quality were performed using Nanodrop ND-1000 spectrophotometer (Nanodrop, USA). Additional quality control was made by means of agarose gel (2 %) electrophoresis with formaldehyde and ethidium bromide. Reverse transcription reaction was conducted using RevertAid first Strand Synthesis Kit (Fermentas) according to manufacturer’s protocol. Primers (Table 2) were designed using open access software from PREMIER Biosoft International (NetPrimer: http://www.premierbiosoft.com/netprimer) and Invitrogen (OligoPerfect Designer: http://tools.lifetechnologies.com/content.cfm?pageid=9716). One of the primers from each pair had a sequence corresponding to microarray probe sequence from Maize Oligonucleotide Array Project, Arizona (ftp.maizegdb.org/MaizeGDB/FTP/arizona_maize_arrays/; Trzcinska-Danielewicz et al. 2009). On the basis of test of three sequences: GAPDH, β-actin, ubiquitin and literature data (Gomez-Anduro et al. 2011; Sekhon et al. 2011), as reference sequence, ubiquitin (UBQ) was used. This sequence was characterized by the constant expression in all experimental variants. Real-time PCR was carried out in a MyiQ2 (Bio-Rad) thermocycler using SYBR Green JumpStart Taq Ready Mix (Sigma). On every plate three replicates were performed for each sample and three for a template-free control. The standard amplification protocol for RT-qPCR has been described elsewhere (Trzcinska-Danielewicz et al. 2009). The data was analyzed with iQ5 Optical System Software (Bio-Rad). The Ct value (threshold cycle, the moment when a threshold fluorescence level is reached by the amplification product) was determined for all the samples, and the relative expression values were calculated using the method of Dussault and Pouliot (2006), where the normalized difference between Ct values is represented by the formula: ΔΔCt = [(CtGOI K − CtHKG K) − (CtGOI C − CtHKG C)], where: CtG.O.I. and CtHK.G are the threshold cycles of the gene of interest and the reference sequence (house-keeping gene), respectively (K—control sample from non-chilled plants, C—sample from chilled plants).
Results
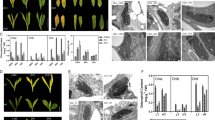
For six of the eleven transcripts localization suggesting connection with transport processes (phloem loading and transport via plasmodesmata) was observed (Table 1). As shown by in situ hybridization, transcripts related to phloem loading process, i.e.: SUT2, SUT4, PPI1 and aquaporin were localized in the companion cells and sieve elements (Table 1; Fig. 1). Transcripts of sucrose transporters SUT2 and SUT4 were localized mainly in the cytoplasm of companion cells and thin-walled sieve tubes (Table 1; Fig. 1 a-d). In addition, SUT2 transcript was localized in thick-walled sieve tubes (Fig. 1a, b). The transcripts of PPI1 and aquaporin showed a similar localization as transcripts of sucrose transporters—the labeling was mainly observed in the companion cells, thin-walled sieve tubes and in vascular parenchyma cells (Table 1; Fig. 1 e–h). Transcripts of plasmodesmata-related genes: remorin and TMV-MP30, were mainly localized in the cytoplasm and the cortical ER of Kranz mesophyll and bundle sheath cells (Table 1; Fig. 2a–d). In these cells, the accumulation of gold particles often occurred in the vicinity of pit-fields or near individual plasmodesmata (Fig. 2a, c). In the case of TMV-MP30, clear accumulation of gold particles in thin-walled sieve tubes was also noted (data not shown).

In situ localization of transcripts related to phloem loading process in control (non-chilled) and chilled leaves of chilling-tolerant (CT) and chilling-sensitive (CS) maize lines. Localization of transcripts of sucrose transporters: SUT2 in control (a) and chilled (14 °C) plants of CS maize line (b); SUT4 transcript in control (c) and chilled (8 °C) plants of CT maize line (d). Localization of transcripts of proton pump interactor (PPI1) in control (e), and chilled (8 °C) plants (f) of CT maize line and aquaporin transcript in control (g) and chilled (8 °C) (h) plants of CS maize line. Arrows indicate a single gold particles or clusters of gold grains. VP vascular parenchyma, CC companion cell, SE thin-walled sieve tube, SET thick-walled sieve tube. Scale bar 200 nm

In situ localization of transcripts of plasmodesmata-related proteins in control (non-chilled) and chilled leaves of chilling-tolerant (CT) and chilling-sensitive (CS) maize lines. Localization of remorin transcript near PD at KMS/BS interface in control (a) and chilled (14 °C) plants (b) of CS maize line; TMV-MP30 transcript near PD at KMS/BS interface and ER in control plants of CS maize line (c) and in chilled (8 °C) plants of CT maize line (d). Negative control, where the hybridization with specific DIG-probe was omitted (e); negative control with DIG-probe sense for TMV-MP30 transcript (f); positive control with localization of β-actin transcript (g). Arrows indicate a single gold particles or clusters of gold grains. KMS Kranz mesophyll, BS bundle sheath, VP vascular parenchyma, CC companion cell, SE thin-walled sieve tube, Pd plasmodesmata. Scale bar 200 nm
Images obtained for the two types of negative controls—in the variant where the hybridization with specific DIG-probe was omitted and in the variant where the sense DIG-probe was used confirmed the reliability of in situ hybridization method—single grains of colloidal gold, randomly arranged in maize cells were observed (Fig. 2e, f). Similarly, the positive control where β-actin transcript was used, successfully verified this method. The intensive labeling of β-actin transcript in most types of maize cells was observed (Fig. 2g).
The results of transcriptomic analysis by RT-qPCR showed that the character of gene expression changes in cold-treated plants of both, CT and CS maize lines was similar (Fig. 3). However, in CS maize line, these changes were more distinct (Fig. 3e–h). Generally, for genes related to phloem loading process, sucrose transporters SUT2 and SUT4 as well as, putative proton pump interactor (PPI1) but not aquaporin, weak expression was observed (Fig. 3). For aquaporin transcript, the repression was noted in the case of both types of stress, with clear effect in CS maize line (Fig. 3). Transcripts of plasmodesmata-related proteins showed clear repression, especially in the leaves of CS maize line, after prolonged period of chilling-treatment (Fig. 3f, h).

Difference in the gene expression shown as ΔΔCt (±SD) between control and cold-treated plants of two maize lines: chilling-tolerant (CT) and chilling-sensitive (CS) stressed for 4 and 28 h at 14 and 8 °C temperature. Gene expression differences were estimated by RT-qPCR with ubiquitin as a reference gene
Generally, the cold did not cause an alternation of labeling intensity, what was partly confirmed by RT-qPCR results. In the case of both sucrose transporters transcripts, SUT2 and SUT4, no marked changes in the density of labeling were observed (Fig. 1a–d). Similarly, the results of PPI1 and aquaporin transcripts labeling showed no difference in density of colloidal gold particles, both within CS maize line and between CS and CT maize lines (Fig. 1e–h). However, for transcripts of plasmodesmata-related proteins (remorin and TMV-MP30), the substantial reduction in the density of labeling in cells of CS maize line, particularly in the cytoplasm near the plasmodesmata was noted (Fig. 2a–d).
Discussion
This paper attempts to determine the cellular localization and changes in gene expression in response to low temperature in two maize lines of different cold sensitivity. The hypothesis about the effect of low temperature on expression of genes connected with transport processes was verified. In a first step, to find out which of the selected 11 transcripts are localized in cells/areas related to transport processes (plasmodesmata, sieve elements) in situ hybridization was performed in leaves of control (non-chilled) plants. For six transcripts specific localization was successfully confirmed (Table 1). Generally, transcripts related to phloem loading process, sucrose transporters, PPI1 and aquaporins were localized in companion cells and thin-walled sieve tubes (Table 1; Fig. 1), while transcripts of plasmodesmata-related proteins in Kranz mesophyll and bundle sheath cells were mainly observed (Table 1; Fig. 2a–d). The specific localization of transcripts (mainly sucrose transporters) in sieve elements (Fig. 1) confirms earlier assumption about putative transport of mRNA via plasmodesmata into the phloem (Kühn et al. 1997; Xoconostle-Càzares 1999) and in case of maize, there is the extensive plasmodesmal connection between parenchymatic cells and sieve elements (Evert et al. 1978). For instance, CmPP16 protein from Cucurbita maxima determines the symplasmic transport of mRNA into sieve elements (Xoconostle-Càzares 1999). In turn, the localization of transcript (SUT2) within thick-walled sieve elements (Fig. 1a, b) may indicate a more important role of these cells than previously assumed. Evert et al. (1978) supposed that these cells may participate in retrieval of substances, which enter to apoplast in the transpiration stream.
Among previously selected four transcripts of different aquaporins, for one localization in phloem elements (GRMZM2G178693) was successfully confirmed (Table 1; Fig. 1g, h). This observation may prove their participation in the phloem loading, which for most aquaporin forms are not completely clear, although in maize, various types of aquaporins are localized in different cells of plant tissues and indirect part of these proteins in phloem loading, by transporting water from xylem to sieve tubes has been suggested (Chaumont et al. 2001).
Results of RT-qPCR showed that changes in the expression of genes related to phloem loading were relatively small and in most cases upregulation was observed (Fig. 3). Only, in the case of prolonged period of chilling-treatment, in CS maize line the weak repression of SUT2 was observed (Fig. 3f, h). It could be explained by the mechanism, in which one transporter is not directly involved in the sugar transport, but it works as a modulator of the expression and activity of another carrier (Lalonde et al. 1999). For instance, AtSUT2 transporter localized in companion cells and sieve tubes in Arabidopsis thaliana may be involved in the regulation of SUT1 and SUT4 transporters (Barker et al. 2000). It should be noted that although SUT2 and SUT4 genes in maize are orthologues of the Arabidopsis genes, they may perform other functions in this species (Lalonde et al. 2004). However, the repression of SUT2 transcription in plants of CS maize line chilled for 28 h may explain the inhibition of phloem loading (Sowiński 1995; Sowiński et al. 2001; Bilska and Sowiński 2010), while the induction of SUT4 expression at 8 °C is difficult to interpret due to the fact that phloem loading process in maize is completely inhibited at temperature less than 10 °C (Sowiński et al. unpublished). It was also possible that expression of sucrose transporters depends on the current concentration of sucrose in cells (Lalonde et al. 1999). In this scenario, low level of sucrose in the cell is detected by intracellular receptor that activates signaling leading to the induction of the expression of genes associated with high affinity/low capacity carriers type, which include, i.a., SUT1. Conversely, a high level of sugar in the cell may cause repression of this type of transporter. Also, at high concentration of sugars in the apoplast, the induction of the low affinity/high capacity transporters type, which include i.a. SUT4 (Weise et al. 2000), can increase the efficiency of sucrose transport. In turn, the weak induction of expression of PPI1 transcript in variants with short (4 h) chilling-treatment in CS maize line (Fig. 3e, g) can be interpreted in two ways. First, due to the fact that proton pump interactor may be involved in the regulation of proton pomp functioning (Turgeon 2006) it can show changes in the expression depending on the activity of proton pump, independently of the experimental conditions. On the other hand, the clear induction of PPI1 expression in CS maize line after short chilling-treatment may indicate a type of ‘first’ reaction of plants to the relatively short-term stress.
Clear repression of aquaporin was observed in CS line, after prolonged period of chilling-treatment at 14 °C and in both variants at 8 °C (Fig. 3 f–h). Indeed, in this maize line, phloem loading is inhibited after just one hour of exposure to low temperature (14 °C), but after a longer period of cold-treatment these changes are more pronounced (Bilska and Sowiński 2010). Thus, the possibility that the inhibition of phloem loading is connected, at least partially, with limitation of the water flow from xylem to phloem cannot be excluded.
Changes in expression of plasmodesmata-related genes were evident (Fig. 3), and are in line with earlier observation concerning closure of plasmodesmata at cold (Bilska and Sowiński 2010). The pronounced repression of remorin gene in CS line after 28-h of chilling-treatment (Fig. 3f, h) was confirmed by decrease of the labeling density (Fig. 2a, b). Previously, remorins have been localized within the plasmodesmata in Solanum lycopersicum (Raffaele et al. 2009). The authors indicated the potential role of these proteins in the response to biotic stress, more specifically—regulation of the transport efficiency of potato virus X (PVX) via plasmodesmata. Orthologues of maize remorin (GRMZM2G081949) in Arabidopsis thaliana belong to remorin group 4 (AtREM4.1 and AtREM4.2) and they are highly regulated by different abiotic and biotic stresses (Son et al. 2014). In above studies, however, the induction of AtREMs was observed under osmotic stress, abscisic acid (ABA) and in senescing leaves.
In the case of second examined plasmodesmata-related gene: TMV-MP30, the repression was noted in most variants for CS maize lines (Fig. 3f–h), which was confirmed by in situ hybridization (Fig. 2c, d). Protein coded by this gene is probably involved in the regulation of transport of TMV virus by plasmodesmata (Ding et al. 1992; Kragler et al. 2003; Wright et al. 2007; Sambade et al. 2008). The observed changes in the gene expression and transcript localization in maize plants under low temperature may be indicative of the nonspecific reaction during activation of defense mechanism typical for different types of stresses.
Conclusion
In this work, we investigated changes in maize gene (related to intercellular transport) expression in cold condition with the confirmation of their subcellular localization. Marked changes in the expression level of genes coding plasmodesmata-related proteins: remorin and TMV-MP30, which are involved in the regulation of virus transport through plasmodesmata, may suggest non-specific reaction to stress in the chilled maize plants. The localization of sucrose transporter and aquaporin transcripts in companion cells and thin-walled sieve tubes indicates the involvement of these proteins in the phloem loading process. In addition, the localization of SUT2 transcripts in thick-walled sieve tubes may indicate that both types of sieve tubes (thin- and thick-walled) are involved in the transport of photoassimilates. The assignment of the biological function of thick-walled sieve tubes opens up new directions in this research area.
Author contribution statement
A. Bilska-Kos and P. Sowiński contributed to the idea of this work. Experiments and data analyses were carried out by A. Bilska-Kos, M. Grzybowski, M. Jończyk and P. Sowiński. The manuscript was written by A. Bilska-Kos, M. Jończyk and P. Sowiński. All the authors approved the manuscript in its final form.
Abbreviations
- DIG:
-
Digoxygenin
- GA:
-
Glutaraldehyde
- PFA:
-
Paraformaldehyde
- PIPES:
-
1,4-Piperazinediethanesulfonic acid
- PPI1:
-
Proton pomp interactor
- SUT:
-
Sucrose transporter
- TMV-MP30:
-
Tobacco mosaic virus movement protein
References
Baker NR, East TM, Long SP (1983) Chilling damage to photosynthesis in young Zea mays: II. Photochemical function of thylakoids in vivo. J Exp Bot 34:189–197
Barker L, Kühn C, Weise A, Schulz A, Gebhardt C, Hirner B, Hellmann H, Schulze W, Ward JM, Frommer WB (2000) SUT2, a putative sucrose sensor in sieve elements. Plant Cell 12:1153–1164
Barkla BJ, Vera-Estrella R, Pantoja O, Kirch H-H, Bohnert HJ (1999) Aquaporin localization—how valid are the TIP and PIP labels? Trends Plant Sci 4:86–88
Bennett K, McPherson H, Warrington I (1982) Effect of pretreatment temperature on response of photosynthesis rate in maize to current temperature. Aust J Plant Physiol 9:773–781
Bilska A, Sowiński P (2010) Closure of plasmodesmata in maize (Zea mays) at low temperature: a new mechanism for inhibition of photosynthesis. Ann Bot 106:675–686
Chaumont F, Barrieu F, Wojcik E, Chrispeels MJ, Jung R (2001) Aquaporins constitute a large and highly divergent protein family in maize. Plant Physiol 125:1206–1215
Ding B, Haudenshield JS, Hull RJ, Wolf S, Beachy RN, Lucas WJ (1992) Secondary plasmodesmata are specific sites of localization of the tobacco mosaic virus movement protein in transgenic tobacco plants. Plant Cell Online 4:915–928
Dussault AA, Pouliot M (2006) Rapid and simple comparison of messenger RNA levels using real-time PCR. Biol Proced Online 8:1–10
Evert RF, Eschrich W, Heyser W (1978) Leaf structure in relation to solute transport and phloem loading in Zea mays L. Planta 138:279–294
Evert RF, Russin WA, Bosabalidis M (1996) Anatomical and ultrastructural changes associated with sink-to-source transition in developing maize leaves. Int J Plant Sci 157:247–261
Fryer MJ, Oxborough K, Martin B, Ort DR, Baker NR (1995) Factors associated with depression of photosynthetic quantum efficiency in maize at low growth temperature. Plant Physiol 108:761–767
Gomez-Anduro G, Ceniceros-Ojeda EA, Casados-Vazquez LE, Bencivenni C, Sierra-Beltran A, Murillo-Amador B, Tiessen A (2011) Genome-wide analysis of the beta-glucosidase gene family in maize (Zea mays L. var B73). Plant Mol Biol 77:159–183
Kingston-Smith AH, Foyer CH (2000) Bundle sheath proteins are more sensitive to oxidative damage than those of the mesophyll in maize leaves exposed to paraquat or low temperatures. J Exp Bot 51:123–130
Kjellbom P, Larsson C, Johansson I, Karlsson M, Johanson U (1999) Aquaporins and water homeostasis in plants. Trends Plant Sci 4:308–314
Kragler F, Curin M, Trutnyeva K, Gansch A, Waigmann E (2003) MPB2C, a microtubule-associated plant protein binds to and interferes with cell-to-cell transport of tobacco mosaic virus movement protein. Plant Physiol 132:1870–1883
Kühn C, Franceschi VR, Schulz A, Lemoine R, Frommer WB (1997) Macromolecular trafficking indicated by localization and turnover of sucrose transporters in enucleate sieve elements. Science 275:1298–1300
Lalonde S, Boles E, Hellmann H, Barker L, Patrick JW, Frommer WB, Ward JM (1999) The dual function of sugar carriers: transport and sugar sensing. Plant Cell 11:707–726
Lalonde S, Wipf D, Frommer WB (2004) Transport mechanisms for organic forms of carbon and nitrogen between source and sink. Annu Rev Plant Biol
Maurel C (2007) Plant aquaporins: Novel functions and regulation properties. FEBS Lett 581:2227–2236
Miedema P (1982) The effects of low temperature on Zea mays. Adv Agron 35:93–128
Patrick JW, Zhang W, Tyerman SD, Offler CE, Walker NA (2001) Role of membrane transport in phloem translocation of assimilates and water. Funct Plant Biol 28:697–709
Potvin C, Goeschl JD, Strain BR (1984) Effects of Temperature and CO2 enrichment on carbon translocation of plants of the C4 grass species Echinochloa crus-galli (L.) Beauv. from cool and warm environments. Plant Physiol 75:1054–1057
Potvin C, Strain BR, Goeschl JD (1985) Low night temperature effect on photosynthate translocation of two C4 grasses. Oecologia 67:305–309
Raffaele S, Bayer E, Lafarge D, Cluzet S, German Retana S, Boubekeur T, Leborgne-Castel N, Carde J-P, Lherminier J, Noirot E, Satiat-Jeunemaître B, Laroche-Traineau J, Moreau P, Ott T, Maule AJ, Reymond P, Simon-Plas F, Farmer EE, Bessoule J-J, Mongrand S (2009) Remorin, a solanaceae protein resident in membrane rafts and plasmodesmata, impairs potato virus X movement. Plant Cell Online 21:1541–1555
Robinson DG, Sieber H, Kammerloher W, Anton RS (1996) PIP1 aquaporins are concentrated in plasmalemmasomes of Arabidopsis thaliana mesophyll. Plant Physiol 111:645–649
Sakurai J, Ishikawa F, Yamaguchi T, Uemura M, Maeshima M (2005) Identification of 33 rice aquaporin genes and analysis of their expression and function. Plant Cell Physiol 46:1568–1577
Sakurai J, Ahamed A, Murai M, Maeshima M, Uemura M (2008) Tissue and cell-specific localization of rice aquaporins and their water transport activities. Plant Cell Physiol 49:30–39
Sambade A, Brandner K, Hofmann C, Seemanpillai M, Mutterer J, Heinlein M (2008) Transport of TMV movement protein particles associated with the targeting of RNA to plasmodesmata. Traffic 9:2073–2088
Savitch LV, Ivanov AG, Gudynaite-Savitch L, Huner NPA, Simmonds J (2011) Cold stress effects on PSI photochemistry in Zea mays: differential increase of FQR-dependent cyclic electron flow and functional implications. Plant Cell Physiol 52:1042–1054
Sekhon RS, Lin H, Childs KL, Hansey CN, Buell CR, de Leon N, Kaeppler SM (2011) Genome-wide atlas of transcription during maize development. Plant J 66:553–563
Smoliński DJ, Niedojadło J, Noble A, Górska-Brylass A (2007) Additional nucleoli and NOR activity during meiotic prophase I in larch (Larix decidua Mill.). Protoplasma 232:109–120
Sobkowiak A, Jończyk M, Jarochowska E, Biecek P, Trzcinska-Danielewicz J, Leipner J, Fronk J, Sowiński P (2014) Genome-wide transcriptomic analysis of response to low temperature reveals candidate genes determining divergent cold-sensitivity of maize inbred lines. Plant Mol Biol 85:317–331
Son S, Oh CJ, An CS (2014) Arabidopsis thaliana remorins interact with SnRK1 and play a role in susceptibility to Beet Curly Top Virus and Beet Severe Curly Top Virus. Plant Pathol J 30:269–278
Sowiński P (1995) Transport of assimilates from leaves to roots in cold-treated maize seedlings. Kinetics and assimilate distribution. Acta Physiol Plant 17:341–348
Sowiński P, Rudzińska-Langwald A, Dalbiak A, Sowińska A (2001) Assimilate export from leaves of chilling-treated seedlings of maize. The path to vein. Plant Physiol Biochem 39:881–889
Sowiński P, Rudzińska-Langwald A, Kobus P (2003) Changes in plasmodesmata frequency in vascular bundles of maize seedling leaf induced by growth at sub-optimal temperatures in relation to photosynthesis and assimilate export. Environ Exp Bot 50:183–196
Trzcinska-Danielewicz J, Bilska A, Fronk J, Zielenkiewicz P, Jarochowska E, Roszczyk M, Jończyk M, Axentowicz E, Skoneczny M, Sowiński P (2009) Global analysis of gene expression in maize leaves treated with low temperature: I. Moderate chilling (14 °C). Plant Sci 177:648–658
Turgeon R (2006) Phloem loading: how leaves gain their independence. Bioscience 56:15–24
Wardlaw IF, Bagnall D (1981) Phloem transport and the regulation of growth of Sorghum bicolor (Moench) at low temperature. Plant Physiol 68:411–414
Weise A, Barker L, Kühn C, Lalonde S, Buschmann H, Frommer WB, Ward JM (2000) A new subfamily of sucrose transporters, SUT4, with low affinity/high capacity localized in enucleate sieve elements of plants. Plant Cell 12:1345–1356
Wright KM, Chapman S, Roberts AG (2007) Plasmodesmal targeting and accumulation of TMV movement protein. Plant Signaling Behav 2:180–181
Xoconostle-Càzares B (1999) Plant paralog to viral movement protein that potentiates transport of mRNA into the phloem. Science 283:94–98
Acknowledgments
TEM observation was performed in the Laboratory of Electron Microscopy, Nencki Institute of Experimental Biology, PAS, Warsaw, Poland. This work was financially supported by Grant No 5496/B/P01/2010/39 from the National Science Centre, Poland.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by O Ferrarese-Filho.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Bilska-Kos, A., Grzybowski, M., Jończyk, M. et al. In situ localization and changes in the expression level of transcripts related to intercellular transport and phloem loading in leaves of maize (Zea mays L.) treated with low temperature. Acta Physiol Plant 38, 123 (2016). https://doi.org/10.1007/s11738-016-2151-5
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11738-016-2151-5