
Abstract
We investigated the effects of several tree species on dehydrogenase and urease activities in soils derived from two different parent materials (glaciofluvial sand and loess) in forested areas in southern Poland. We hypothesized that coniferous forests (pine, spruce) alter the soil cation exchange capacity (CEC) and decrease soil pH and, therefore, might decrease soil enzyme activities compared with broadleaf species growing on similar soils. Eight paired plots (12 × 12 m) were established on glaciofluvial sand in pine (Pinus sylvestris) + oak (Quercus robur) and spruce (Picea abies) + pine stands, as well as on loess-derived soils: beech (Fagus sylvatica) + pine and hornbeam (Carpinus betulus) + pine stands. Each plot was a 4 × 4 m grid with 16 sampling points. In soil samples pH, soil texture, and organic carbon, nitrogen, base cation contents, dehydrogenase and urease activities were determined. On both parent materials, the soil pH was lower under coniferous species than under broadleaf species. The acidifying effect of tree species on sandy soil was in the order of spruce = pine > oak, while that on loess was pine > beech > hornbeam. Hornbeam and oak increased the soil pH and stimulated enzyme activity in the soil. The content of fine fraction enhanced potential enzyme activities in soils, thus the loess soils had greater dehydrogenase and urease activity. The results suggest that pine stores more soil organic C in association with silt increasing the pool of stabilized soil organic C.
Similar content being viewed by others


Introduction
Changes in the species composition of forest stands may lead to modifications of soil properties. Tree species affect soil organic matter (SOM) accumulation (Augusto et al. 2002; Hobbie et al. 2007; Mueller et al. 2012), pH (Nihlgård 1971; Paluch and Gruba 2012; Gałka et al. 2014; Łabaz et al. 2014), and the cation-exchange capacity (CEC) of the soil (Hobbie et al. 2007; Gruba and Mulder 2015). The soil microbial biomass and soil enzyme activity could be affected by tree species in a variety of ways (Grayston and Prescott 2005; Adamczyk et al. 2014). Tree species may affect soil enzyme activities by influencing the intrinsic properties of tree-derived SOM, pH modification, or nutrient availability (Kotroczó et al. 2014). Forest litter and root exudates deliver organic matter that contains various constituents, such as soluble sugars, organic acids, amino acids, or starch (Baldrian and Šnajdr 2011). The specific composition of organic matter may lead to the modification of soil acidity and nutrient pools (Prescott 2005; Mueller et al. 2012; Prescott and Grayston 2013). Generally, coniferous forests have more acidifying effects on soil than deciduous or mixed forests. Augusto et al. (2002) ranked tree species in order of decreasing acidifying ability: conifers ≥ beech, oak, and birch ≥ maple, hornbeam, ash, and lime. pH is an important variable that influences SOM decomposition dynamics. Soil pH governs the ionization of functional groups of organic molecules, as well as the conformations of substrates and enzymes (Frankenberger and Johanson 1982). Several recent investigation showed that in acid forest soils soil pH is related to the saturation of CEC with aluminium (Al) and hydrogen (H), rather than base cations (BC = Ca + Mg + K + Na) (Skyllberg 1999; Gruba and Mulder 2015). Thus, tree species by modification of relationships between Al, H and BC influence soil pH, and other soil properties, such as microbial activity. Enzyme activities are unequally distributed among soil particle-size fractions (Saviozzi et al. 2007). Similar to the enzyme activity, soil organic matter is also heterogeneously distributed among particle size fractions (Lagomarsino et al. 2009). Sorption by soil minerals can protect organic material from microbial attack. Sand has only weak bonding affinities to soil organic matter, while clay particles provide a large surface and numerous reactive sites to adsorb soil organic matter (Allison and Jastrow 2006).
Only scattered information exists concerning the complexity of the relationships between trees species, soil properties, and enzyme activities. The measurement of the activity of enzymes involved in the cycling of nutrients in the soil may be used to assess the quality and health of soil. Soils with greater microbial diversity and biochemical activity are characterized by higher resistance to changes in the environment (Allison and Martiny 2008). The soil quality reflects the condition of the forest stand. Dehydrogenases (DHs) and ureases (URs) are the most important soil enzymes involved in the carbon (C) and nitrogen (N) cycles (Sardans et al. 2008). DH activity plays a role in the biological oxidation of SOM (Makoi and Ndakidemi 2008). URs regulate the release of N–NH\( _{4}^{ + } \) through urea hydrolysis, which is essential in the chain of hydrolysis of amino compounds (Aragón et al. 2014). Wolińska and Stępniewska (2012), Veres et al. (2013) showed that soil enzyme activities are “sensors” of soil organic matter (SOM) decomposition. Decomposition of organic matter in soil depends on substance properties and accessibility of microorganisms and their enzymes.
The aim of this study was to investigate the effects of different tree species (Scots pine (Pinus sylvestris), pedunculate oak (Quercus robur), Norway spruce (Picea abies), European beech (Fagus sylvatica), and common hornbeam (Carpinus betulus)) on the dehydrogenases and urease activities in soils derived from two different parent materials (glaciofluvial sand and loess). The study was conducted in temperate forests in southern Poland. We hypothesized that coniferous forests (pine, spruce) alter the soil CEC and decrease soil pH and, therefore, might decrease soil enzyme activities compared with broadleaf species growing on similar soils. We expect that in soils on both parent materials the pine forests cause a high organic matter accumulation and decrease decomposition rates.
Materials and methods
Sampling sites
The study was conducted in forested areas of southern Poland. Eight plots (12 × 12 m) were established in pine (P. sylvestris), oak (Q. robur), spruce (P. abies), beech (F. sylvatica), and hornbeam (C. betulus) forest stands. The plots were paired (one of them always being pine), and the parent material was similar for each pair. On glaciofluvial sands, we located: pair 1, pine 1-oak, in the Dąbrowa Tarnowska forest district (50 km east of Krakow); and pair 2, pine 2-spruce, in the Niepołomice forest district (40 km east of Krakow). On soil developed from loess, we set up pair 3, pine 3-beech, and pair 4, pine 4-hornbeam, which were located in the Krzeszowice forest district (20 km west of Krakow) (Table 1). Arenosol and Podzol (see Table 1) were derived from sandy glacial deposits. In loess soils (Luvisols) the silt is dominant fraction. Each plot experiment was designed over a regular 4 × 4 m grid, in which 16 sampling points were designated. Soil sampling at a depth of 0–10 cm was performed after removal of the residual organic horizon from the sampling points.
Soil analysis
Soil samples obtained in the field were dried and sieved through 2.0-mm mesh. The pH of the samples was analyzed in H2O using the potentiometric method. The soil texture was determined by laser diffraction (Analysette 22, Fritsch, Idar-Oberstein, Germany). The content of carbon (C) and nitrogen (N) were measured by an elemental analyzer (LECO CNS TrueMac Analyzer (Leco, St. Joseph, MI, USA). As all the samples were carbonate-free, C was assumed to be organic carbon (C = Corg). Base cations (BC = Ca2+, Mg2+, K+, Na+) concentrations were determined after extracting 10 g of soil with 100-mL 1 mol dm−3 of a NH4Cl solution, followed by inductively coupled plasma-optical emission spectrometry (ICP-OES) (iCAP 6500 DUO, Thermo Fisher Scientific, Cambridge, UK). Total acidity (Ht) of the soil was measured after extracting 5 g of soil with 30 mL 1 mol L−1 (CH3COO)2Ca (shaking time 1 h), followed by filtration. Soil on the filter was washed several times by extractant solution up to volume 200 mL (Buurman et al. 1996). 25 mL of the obtained solution was titrated by potentiometric titration (automatic titrator Metler Toledo) to pH 8.2 with 0.1 mol L−1 NaOH. The CuCl2–extractable aluminium (Al)—assumed to be exchangeably and organically bound Al was determined by ICP in the supernatant of a single extraction of 3 g of soil with 30 mL of 0.5 mol L−1 CuCl2 (Juo and Kamprath 1979) after 2-h extraction time, followed by centrifugation at 1500×g the concentration of Al was determined by means of ICP. All extraction procedures described above were followed by filtration through a 0.45-μm Millipore membrane filter.
Enzyme analysis
Dehydrogenase activity (DH) was determined by the reduction of 2,3,5-triphenyltetrazolium chloride (TTC) to triphenyl formazan (TPF) using Lenhard’s method according to the Casida procedure (Alef and Nannipieri 1995). Briefly, 6 g of soil was incubated with 1 mL of 3 % TTC for 24 h at 37°C. TPF was extracted with ethyl alcohol and measured spectrophotometrically.
Urease activity (UR) was determined according to Tabatabai and Bremner (1972) using a water-urea solution as a substrate. This activity was determined by the NH4 + released after a 2-h incubation at 37°C. The concentration of NH4 + was measured at 410 nm by the colorimetric method (Alef and Nannipieri 1995).
Statistical analysis
Principal Components Analysis (PCA) method was used to evaluate the relationships between soil properties, forest species and parent material. In PCA analysis the parent material was expressed by the content of sand, silt and clay. Properties of paired plots were compared using a parametric honestly significant difference (HSD) test. Pearson correlation coefficients between enzyme activities and soil characteristics were also calculated. General linear model (GLM) was used for the investigated the effect of the species composition, parent material and chemical properties on the enzyme activity. All the statistical analyses were performed with Statistica 10 software (2010).
Results
The highest dehydrogenase activity was found in the glaciofluvial sand soils under the oak stand, the lowest dehydrogenase activity was found in the glaciofluvial sand soils under the pine stand. The highest urease activity was found in the loess soils under the hornbeam stand, the lowest urease activity was found in the glaciofluvial sand soils under the pine stand. The significant differences in soil enzyme activity were noted between all the tree stands except paired beech and pine stands. Ranges of pHs obtained at the investigated plots (irrespectively of tree species) showed that sandy soils had lower pHs than soils derived from loess (3.73–4.09 and 4.06–5.03, respectively), however, the differences were relatively small. On glaciofluvial sand, the soil pHs under pine stands were significantly lower than that under oak stands (3.73 and 4.09, respectively). No significant differences in soil pHs were noted between paired spruce and pine stands (Table 2). On loess, the soil pH under pine stands was significantly lower than that under broadleaf stands (Table 2). Differences in the pHs of the studied species pairs varied. The smallest difference in pHs (0.2) was observed in soil developed on loess in the pine 3-beech pair. A greater difference (0.4) was recorded in the soil formed on glaciofluvial sands in the pine 1-oak pair, whereas the greatest difference (0.8) was found on loess soils in the pine 4-hornbeam pair. Soils of the loess stands had higher base cation contents than sandy soils with similar C contents (Tables 2, 3). In soils derived from both sands and loess, the base cation contents decreased in the following order: Ca2+ > Mg2+ ≥ K+ > Na+, irrespectively of the forest type. The C contents of glaciofluvial sand soils were lower than those of loess soils. Moreover, in sandy soils, the C content did not differ between species, whereas in loess soils, the C content was significantly higher in soil under pine forests than under beech or hornbeam forests (Table 2).
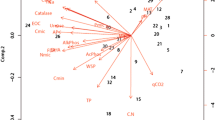
A projection of the variables on the factor-plane clearly demonstrated correlation between the variables and tree species (supplementary variables). Two main factors had a significant total impact (69.19 %) on the variance of the variables in soil. Factor 1 explained 28.81 % of the variance of the examined properties, and Factor 2 explained 39.38 % of the variance (Fig. 1). The soil of hornbeam stands had higher UR and DH activity. In contrast, the soils of coniferous stands were characterized by a high C/N ratio. The soil of hornbeam stands had the highest pH, and Ca2+ and Mg2+ contents. The soils developed from fluvioglacial sands contained a lot of weakly decomposed organic matter (high C/N ratio). Soils developed from loess, with a large content of dust and clay were characterized by higher pH, a higher content of base cations and a better rate of organic matter decomposition. Urease and dehydrogenase activities were associated with the pH (Tables 4, 5). The activity of the studied soil increased with the increase in pH. Enzyme activity positively correlated with the nitrogen content and negatively with C/N ratio. Urease activity was positively correlated with silt and clay content and negatively correlated with sand content (Table 4). In addition, GLM analysis confirmed the simultaneous impact of species composition and parent material on the enzymatic activity (Table 5).

Projection of the variables on the factor-plane in soils on both parent material (O oak, S spruce, P pine, H hornbeam, B beech, P pine, DH dehydrogenase activity, UR urease activity, C carbon content, N nitrogen content, Ht total acidity)
For further analysis, enzyme activities were expressed in relation to Ct. Such relative values of enzyme activities were always lower in pine forest soils (Fig. 2). For DHs, the highest values were noted in oak and hornbeam forest soils, and similar results were obtained for URs (Fig. 3). Figures 4 and 5 show the relationships between DH activity, pH, and the C/N ratio. The increase in DH activity in soils on both parent materials correlated with high pH and low C/N ratio. The highest DH activity occurred in an oak stand on soil formed on glaciofluvial sands and under a hornbeam stand on loess.

Effect of tree species on the variation of dehydrogenase (DH) activity per unit of carbon

Effect of tree species on the variation of urease (UR) activity per unit of carbon

Three-dimensional plot of the relationship between dehydrogenase activity, pH in H2O, and the C/N ratio in soils on glaciofluvial sands in different tree stands (circles with O—group of oak plots, circles with S—group of spruce plots, circles with P—group of pine plots); regression plane marked by black lines

Three-dimensional plot of the relationship between dehydrogenase activity, pH in H2O, and the C/N ratio in soils on loess in different tree stands (circles with H group of hornbeam plots, circles with B group of beech; circles with P group of pine plots); regression plane marked by black lines
Discussion
When we take into account only pine sites, the C content in soils derived from glaciofluvial sand was found to be lower than in the loess soils. Our results suggest that pine stores more soil organic C in association with clay and silt increasing the pool of stabilized organic C in soil. According to Six et al. (2002), Dobarco and Van Miegroet (2014), Gruba et al. (2015) in the mineral soils, the persistence of soil organic C is further enhanced by the mineral matrix through additional protection mechanisms, such as the isolation of organic matter inside aggregates and surface interactions between organic compounds and mineral particles, mainly from the silt and clay fraction. The loess soils under the pine forest revealed higher enzyme activity than the soils developed on glaciofluvial sand. The loess soils contain more silt and clay fractions, which promotes enzyme activity. Allison (2006) and Ling et al. (2014) found that clay minerals, enhanced potential enzyme activities in soils. Gianfreda et al. (2005) reported a correlation between the activity of enzymes (arylsulfatase, β-glucosidase, phosphatase, urease and dehydrogenase) and soil texture. They found a positive correlation between the enzymatic activity and clay and silt contents, and a negative correlation with the content of sand. Soil containing more clay fraction may maintain larger microbial communities and lower fluctuation of water availability (Chodak and Niklińska 2010). Urease activity is mainly located in the clay-sized fraction and depends on mineral sorption processed (Kandeler et al. 1999).
We also noted a significant correlation between enzyme activity and C/N ratio. A lower C/N ratio was found in broadleaf stands compared with coniferous stands. The C/N ratio is often used to describe litter quality, and broadleaf species have a lower C/N ratio than pine and spruce species (Handsson et al. 2011). The slow rate of SOM decomposition in coniferous forests was linked to lower litter quality, as evidenced by higher C/N ratios (Zhang et al. 2008). According to van Nevel et al. (2014), the C/N ratio of topsoil in oak stands was lower than that in pine stands. According to Fontaine et al. (2003) not only the amount of organic matter in the soils is important but most of all its quality, as organic matter affects the supply of energy for microbial growth and enzyme production. The differences in enzyme activities and other soil characteristics among deciduous trees is the effect of differing contents of lignin and nitrogen. Hobbie et al. (2007) and Vesterdal et al. (2012) suggest that tree species influence the microbial decomposition mainly by differing contents of lignin and nitrogen; species with high lignin content and low nitrogen content decompose more slowly, which has a long term impact on the C/N ratio in soils. According to Jacob et al. (2010) the lignin content in the leaves of Fagus sylvatica and Carpinus betulus amounts to 88.7 and 80.7 mg g−1. Conifers typically have higher lignin content (25–33 % of dry weight) than broadleaf trees (20–25 %) (Sjöström and Westermark 1999). Differences in enzyme activity between pine and spruce stands can be associated with nitrogen content. In soils of spruce stands the higher nitrogen content were recorded. The increase in the availability of nitrogen is the cause of stimulation of microbial activity (Qian et al. 2014; Ye et al. 2014). The results confirmed a strong relationship of the enzymatic activity with the organic matter quality and pH. The increase in pH caused by the influence of the species results in improving the quality and rate of organic matter decomposition and in the increase in enzyme activity regardless of parent material type and particle size. Acidifying species (pine and spruce) affect the decrease in the rate of organic matter decomposition and at the same time reduce the enzyme activity. Among the species the hornbeam on loess and oak on fluvioglacial sand influenced most beneficially the organic matter quality, pH and the stimulation of microbial activity.
The chemical composition of SOM largely results from microbial transformation of tree litter, whose composition, in turn, influences the activity and diversity of soil microorganisms (Ushio et al. 2008; Pérez-Bejarano et al. 2010). Soil organic matter is an organic carrier of soil enzymes. It is always difficult to determine whether the observed modifications in the enzymatic activities are due to the organic matter content in the soils or due to actual differences in enzymatic activity (Allison and Jastrow 2006; Trasar-Cepeda et al. 2008). One of the approaches to solving this issue is to use the values of enzyme activity per unit of organic carbon. Expressing activity on an organic matter basis shows a microbial property, as it expresses the nutritional status of organic matter present from the perspective of the microorganisms (Wang et al. 2012). Trasar-Cepeda et al. (2008) expressed the values of activity per unit of C to compare soils under different types of use. Here, DH and UR activities were expressed per unit of C to compare the activities of enzymes in coniferous and broadleaf forest soils. The use of this relationship revealed that enzyme activities in the soils affected by broadleaf forests were higher than these in pine soils. Conifer litter contains more components that are recalcitrant to decomposition than broadleaf litter, resulting in litter accumulation in the forest floor and the formation of acidic compounds (Berg 2000). Moreover, in acidic soils, soil fauna is less active, thereby decreasing the amount of organic matter mixing with the mineral soil (Thuille and Schulze 2006), and more material remains on the forest floor.
In the soils of pine stands on both parent materials, the smallest DH and UR activities, as well as the lowest pH, were noted. pH affects soil enzymes activity by changing in the ionic form of the active sites of the enzymes and by affecting the affinity of the substrate to the enzyme (Wolińska and Stępniewska 2012). We conclude that the decrease in pH was probably caused by tree species-related modification of the relationship between H+ and Al3+, which affected the composition of CEC, thereby leading to shifts in microbial community composition and size, as well as enzyme dynamics. The pHs of soil controls enzyme activities via its influences on enzyme conformations and on the adsorption of soil colloids (Frankenberger and Johanson 1982; Turner 2010). According to Šnajdr et al. (2013), tree species is the most important factor affecting the activity of extracellular enzymes. As extracellular enzymes are produced by soil microorganisms, differences in enzyme activities may be partly caused by differences in microbial community composition (Urbanová et al. 2015).
On both parent materials, the soil pHs was lower under coniferous species than broadleaf species. Large differences in soil pHs confirmed the stronger acidifying effect of pine and spruce compared with that of the deciduous species. Many previous studies demonstrated a strong acidifying effect of coniferous species on soil. Kulhavy et al. (2010) found a decrease in pH and the soil CEC under spruce stands compared with those under beech stands. Gałka et al. (2014) found a lower pH and increased content of exchangeable aluminum under spruce stands compared with a broadleaf forest. We ranked the investigated tree species according to their acidifying effect on soil in the order of spruce = pine > oak on sandy soils, and pine > beech > hornbeam on loess soils, which is consistent with the results of Augusto et al. (2002), who ordered the acidifying effect of common European tree species as follows: P. abies, P. sylvestris > Abies alba > Betula pendula, F. sylvatica, Quercus petraea, Q. robur > Acer platanoides, Carpinus betulus, Fraxinus excelsior, and Tilia cordata. According to Gruba and Mulder (2015), tree species influence soil pH by modifying the composition of CEC. Soil under conifers has a higher percentage of hydrogen in CEC, while soils under oak stands have higher pHs due to a greater saturation with aluminum ions, i.e. Al ions behave like a base cation. The same authors confirmed the previous findings of Skyllberg et al. (2001), who demonstrated that in acidic forest soils (with pH < 4.5), base cations, such as Ca2+ or Mg2+, do not influence soil pH due to small share in CEC, moreover, pH is related to the relative ratios between cations (data not shown in Figs. 1 and 2), rather than to the their absolute contents. pH value is the strongest driver of Al concentrations in soil solution, therefore, in acid soils the highest concentrations of Al are expected. However, potential toxicity of Al is usually expressed by Ca to Al ratio. The Ca:Al molar ratio thresholds suggested by Cronan and Grigal (1995), indicating higher risk of Al stress, is 1. So far, the toxicity of Al to trees was confirmed only in case of laboratory experiments and young trees (seedlings). There is no confirmed evidence for Al toxicity in field conditions (e.g. De Wit et al. 2010), also in acid Polish forest soils (Gruba et al. 2013).
Conclusions
Lower pHs and slow rate of soil organic matter is presumably a major factor affecting microbial community composition and size and, therefore, pH affects enzyme dynamics. The lowest enzyme activities were found in acidic soils in coniferous forest stands. We ranked the investigated tree species with respect to their stimulating effect on enzyme activities. For sandy soils, the order was oak > spruce > pine, while on loess soils, it was hornbeam > beech > pine. A relationship between enzyme activity and tree species-related SOM quality was noted. Lower C/N ratios confirm the fast rate of SOM decomposition and the increase in DH activity. The results suggest that pine stores more soil organic C in association with silt and clay increasing the pool of longer residence time soil organic C. The silt fraction, enhanced potential enzyme activities in soils. The loess soils contained more fine fractions which promoted enzyme activity.
References
Adamczyk B, Kilpeläinen P, Kitunen V, Smolander A (2014) Potential activities of enzymes involved in N, C, P and S cycling in boreal forest soil under different tree species. Pedobiologia 57:97–102
Alef K, Nannipieri P (1995) Enzyme activities. In: Alef K, Nannipieri P (eds) Methods in applied soil microbiology and biochemistry. Academic Press, London
Allison SD (2006) Soil minerals and humic acids alter enzyme stability: implications for ecosystem processes. Biogeochemistry 81:361–373
Allison SD, Jastrow JD (2006) Activities of extracellular enzymes in physically isolated fractions of restored grassland soils. Soil Biol Biochem 38:3245–3256
Allison SD, Martiny BH (2008) Resistance, resilience, and redundancy in microbial communities. Proc Nat Acad Sci, USA 105:11512–11519
Aragón R, Sardans J, Peñuelas J (2014) Soil enzymes associated with carbon and nitrogen cycling in invaded and native secondary forests of northwestern Argentina. Plant Soil 384:169–183
Augusto L, Ranger J, Binkley D, Rothe A (2002) Impact of several common tree species of European temperate forests on soil fertility. Ann For Sci 59:233–253
Baldrian P, Šnajdr J (2011) Lignocellulose-degrading enzymes in soil. In: Shukla G, Varma A (eds) Soil enzymology. Springer, Berlin, pp 167–186
Berg B (2000) Litter decomposition and organic matter turnover in northern forest soils. Forest Ecol Manag 133:13–22
Buurman P, van Lagen B, Velthorst EJ (1996) Manual for soil and water analysis. Backhuys Publishers, Leiden
Chodak M, Niklińska M (2010) Effect of texture and tree species on microbial properties of mine soils. Appl Soil Ecol 46:268–275
Cronan CS, Grigal DF (1995) Use of calcium/aluminum ratios as indicators of stress in forest ecosystems. J Environ Qual 24:209–226
De Wit HA, Eldhuset TD, Mulder J (2010) Dissolved Al reduces Mg uptake 758 in Norway spruce forest: results from a long-term field manipulation experiment in Norway. For Ecol Manag 259:2072–2082
Dobarco MR, Van Miegroet H (2014) Soil organic carbon storage and stability in the aspen-conifer ecotone in montane forests in Utah, USA. Forests 5:666–688
Fontaine S, Marotti A, Abbadie L (2003) The priming effect of organic matter: a question of microbial competition. Soil Biol Biochem 35:837–843
Frankenberger WTJ, Johanson JB (1982) Effect of pH on enzyme stability in soils. Soil Biol Biochem 14:433–437
Gałka B, Kabała C, Łabaz B, Bogacz A (2014) Influence of stands with diversed share of Norway spruce in species structure on soils of various forest habitats in the Stołowe Mountains. Sylwan 158:684–694
Gianfreda L, Rao AM, Piotrowska A, Palumbo G, Colombo C (2005) Soil enzyme activities as affected by anthropogenic alterations: intensive agricultural practices and organic pollution. Sci Total Environ 341:265–279
Grayston SJ, Prescott CE (2005) Microbial communities in forest floors under four tree species in 766 coastal British Columbia. Soil Biol Biochem 37:1157–1167
Gruba P, Mulder J (2015) Tree species affect cation exchange capacity (CEC) and cation binding properties of organic matter in acid forest soils. Sci Total Environ 511:655–662
Gruba P, Mulder J, Brożek S (2013) Modelling the pH dependency of dissolved calcium and aluminium in O, A and B horizons of acid forest soils. Geoderma 206:85–91
Gruba P, Socha J, Błońska E, Lasota J (2015) Effect of variable soil texture, metal saturation of soil organic matter (SOM) and tree species composition on spatial distribution of SOM in forest soils in Poland. Sci Total Environ 521–522:90–100
Handsson K, Olsson BA, Olsson M, Johansson U, Kleja DB (2011) Differences in soil properties in adiacent stands of Scots pine, Norway spruce and Siver birch in SW Sweden. Forest Ecol Manag 262:522–530
Hobbie SE, Ogdahl M, Chorover J, Chadwick OA, Oleksyn J, Zytkowiak R, Reich PB (2007) Tree species effects on soil organic matter dynamics: the role of soil cation composition. Ecosystems 10:999–1018
Jacob M, Viednz K, Polle A, Thomas FM (2010) Leaf litter decomposition in temperate deciduous forest stands with a decreasing fraction of beech (Fagus sylvatica). Oecologia 164:1083–1094
Juo ASR, Kamprath EJ (1979) Copper chloride as an extractant for estimating the potentially reactive aluminium pool in acid soils. Soil Sci Soc Am J 43:35–38
Kandeler E, Stemmer M, Klimanek EM (1999) Response of soil microbial biomass, urease and xylanase within particle size fraction to long-term soil management. Soil Biol Biochem 31:261–273
Kotroczó Z, Veres Z, Fekete J, Krakomperger Z, Tóth JA, Lajtha K, Tóthmérisz B (2014) Soil enzyme activity in response to long-term organic matter manipulation. Soil Biol Biochem 70:237–243
Kulhavy J, Mensik L, Fabianek T, Drapelova I, Remes M (2010) How the different tree species composition can alter throughfall, chemical properties of subsurface runoff and soil chemistry. 19th World Congress of Soil Science. Soil Solutions for a Changing World, 1–6 August 2010. Brisbane, Australia. Published on DVD, 28–31
Łabaz B, Gałka B, Bogacz A, Waroszewski J, Kabała C (2014) Factors influencing humus forms and forest litter properties in the mid-mountains under temperate climate of southwestern Poland. Geoderma 230–231:265–273
Lagomarsino A, Grego S, Marhan S, Moscatelli MC, Kandeler E (2009) Soil management modifies micro-scale abundance and function of soil microorganisms in a Mediterranean ecosystem. Eur J Soil Sci 60:2–12
Ling N, Sun Y, Ma J, Guo J, Zhu P, Peng C, Yu G, Rau W, Guo S, Shen Q (2014) Response of the bacterial diversity and soil enzyme activity in particle-size fraction of Mollisol after different fertilization in a long-term experiment. Biol Fert soil 50:901–911
Makoi JHJR, Ndakidemi PA (2008) Selected soil enzymes: examples of their potential roles in the ekosystem. Afr J Biotechnol 7:181–191
Mueller KE, Eissenstat DM, Hobbie SE, Oleksyn J, Jagodziński AM, Reich PB, Chadwick OA, Chorover J (2012) Tree species effects on coupled cycles of carbon, nitrogen, and acidity in mineral soils at a common garden experiment. Biogeochemistry 111:601–614
Nihlgård B (1971) Pedological influence of spruce planted on former beech forest soils in Scania, South Sweden. Oikos 22:302–314
Paluch J, Gruba P (2012) Effect of local species composition on topsoil properties in mixed stands with silver fir (Abies alba Mill.). Forestry 85:413–426
Pérez-Bejarano A, Mataix-Solera J, Zornoza R, Guerrero C, Arcenegui V, Mataix-Beneyto J, Cano-Amat S (2010) Influence of plant species on physical, chemical and biological soil properties in a Mediterranean forest soil. Eur J Forest Res 129:15–24
Prescott CE (2005) Do rates of litter decomposition tell us anything we really need to know? Forest Ecol Manag 220:66–74
Prescott PC, Grayston SJ (2013) Tree species influence on microbial communities in litter and soil: current knowledge and research needs. Forest Ecol Manag 309:19–27
Qian X, Gu J, Sun W, Li Yd, Qx Fu, Xj Wang, Gao H (2014) Changes in the soil nutrient levels, enzyme activities, microbial community function, and structure during apple orchard maturation. Appl Soil Ecol 77:18–25
Sardans J, Peñuelas J, Ogaya R (2008) Experimental drought reduced acid and alkaline phosphatase activity and increased organic extractable P in soil in a Quercus ilex Mediterranean forest. Eur J Soil Biol 44:509–520
Saviozzi A, Cardelli R, Labbaci L, Levi-Minzi R, Riffaldi R (2007) Selected enzyme activities in particle-size fractions from an organically and conventionally managed soil. Fres Environ Bull 16:1195–1200
Six J, Conant RT, Paul EA, Paustian K (2002) Stabilization mechanisms of soil organic matter: implications for C-saturation of soils. Plant Soil 241:155–176
Sjöström E, Westermark U (1999) Chemical composition of wood and pulps: basic constituents and their distribution. In: Sjöström E, Alén R (eds) Analytical methods in wood chemistry, pulping, and papermaking. Springer Series in Wood Science, pp 1–19
Skyllberg U (1999) pH and solubility of aluminium in acidic forest soils: a consequence of reactions between organic acidity and aluminium alkalinity. Eur J Soil Sci 50:95–106
Skyllberg U, Raulund-Rasmussen K, Borggaard OK (2001) pH buffering in acidic soils developed under Picea abies and Quercus robur—affects of soil organic matter, adsorbed cations and soil solution ionic strength. Biogeochemistry 56:51–74
Šnajdr J, Dobiášová P, Urbanová M, Petránková M, Cajthaml T, Frouz J, Baldrian P (2013) Dominant trees affect microbial community composition and activity in post-mining afforested soils. Soil Biol Biochem 56:105–115
Tabatabai MA, Bremner JM (1972) Assay of urease activity in soils. Soill Biol Biochem 4:479–487
Thuille A, Schulze ED (2006) Carbon dynamics in successional and afforested spruce stands in Thuringia and the Alps. Global Change Biol 12:325–342
Trasar-Cepeda C, Leirós MC, Gil-Sotres F (2008) Hydrolytic enzyme activities in agricultural and forest soils. Some implications for their use as indicators of soil quality. Soil Biol Biochem 40:2146–2155
Turner BL (2010) Variation in pH optima of hydrolytic enzyme activities in tropical rain forest soils. App Environ Micro 76:6485–6493
Urbanová M, Šnajdr, Baldrian P (2015) Composition of fungal and bacterial communities in forest litter and soil is largely determinant by dominant trees. Soil Biol Biochem 84:53–64
Ushio M, Wagai R, Balser TC, Kitayama K (2008) Variations in the soil microbial community composition of a tropical montane forest ecosystem: does tree species matter? Soil Biol Biochem 40:2699–2702
Van Nevel L, Mertens J, De Schrijver A, De Neve S, Verheyen K (2014) Can schrub species with higher litter quality mitigate soil acidification in pine and oak forests on pour sandy soils? Forest Ecol Manag 330:38–45
Veres Z, Kotroczó Z, Magyaros K, Tóth JA, Tóthmérész B (2013) Dehydrogenase activity in a litter manipulation experiment in temperate forest soil. Acta Silvatica and Lignaria Hungaric 9:25–33
Vesterdal L, Elberling B, Christiansen JR, Callesen I, Schmidt IK (2012) Soil respiration and rates of soil carbon turnover differ among six common European tree species. Forest Ecol Manag 264:185–196
Wang B, Xue S, Lin GB, Zheng GH, Li G, Ren ZP (2012) Changes in soil nutrient and enzyme activities under different vegetations in the Loess Plateau area, Northwest China. Catena 92:186–195
Wolińska A, Stępniewska Z (2012) Dehydrogenase activity in the soil environment. In: Canuto RA (eds) Dehydrogenases. InTech. doi:10.5772/48294
Ye X, Liu H, Li Z, Wang Y, Wang H, Liu G (2014) Effects of green manure continuous application on soil microbial biomass and enzyme activity. J Plant Nutr 37:498–508
Zhang D, Hui D, Luo Y, Zhou G (2008) Rates of litter decomposition in terrestrial ecosystems: global patterns and controlling factors. J Plant Ecol 1:85–93
Acknowledgments
This study was supported by the Polish-Norwegian Research Fund (PNRF-68-AI-1/07).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article

Cite this article
Błońska, E., Lasota, J. & Gruba, P. Effect of temperate forest tree species on soil dehydrogenase and urease activities in relation to other properties of soil derived from loess and glaciofluvial sand. Ecol Res 31, 655–664 (2016). https://doi.org/10.1007/s11284-016-1375-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11284-016-1375-6