
Abstract
We studied the effect of ditch blocking on vegetation composition and water-table depths in a blanket peatland. Measurements were made for a period of four years (water tables) and five years (vegetation) in the inter-ditch areas of three experimental treatments: (i) open ditches, (ii) ditches blocked with closely-spaced dams and (iii) ditches partially infilled with peat and blocked with dams. It is often assumed that ditch blocking will lead to an increase in the abundance of Sphagnum and, potentially, a reduction in the abundance of sedges, particularly the cotton grasses. However, our data show no treatment effects on the abundance of either group. We did find an effect of time, with the abundance of both sedges and Sphagnum spp. varying significantly between some years. For the sedges there was no systematic change over time, while for the Sphagnum spp. abundance tended to increase through the study period. This systematic change was not related to a measure of the vigour of the sedges, although vigour was lower towards the end of the study compared to the beginning. Our vegetation data are consistent with our water-table data. As with plant type abundance, we did not find any statistically significant differences in water-table depths between treatments, both for annual averages and summer averages. We comment on why ditch blocking does not seem to have affected water tables and vegetation composition at our study site.
Similar content being viewed by others

Introduction
In this paper we consider how restoration of drained blanket peatlands affects the composition of their vegetation. Although globally rare (Gallego-Sala and Prentice 2012), blanket peatlands are the most important peatland type in the UK, both by area (15,736 km2—c. 87% of the total UK peatland area—Baird et al. (2009)) and carbon (C) stored (conservatively, at least 1000 Mt C—see end note). Large areas of these peatlands have been ditch drained, with drainage carried out to improve the grazing potential of the land (Holden et al. 2004; Parry et al. 2014). Because of concerns that drainage has damaged the conservation value and C storage potential of these peatlands, there have been calls to undertake large-scale damming or infilling of drainage ditches as a restoration measure (Bain et al. 2011). Despite such calls, there is a general lack of information on how peatland water tables respond to ditch blocking and how blocking affects the composition of blanket peatland vegetation (Parry et al. 2014).
Stewart and Lance (1991) investigated the effect of open ditches, but not blocked ditches, on both water tables and vegetation on blanket bog at Moor House in the N Pennines, N England. The ditches they investigated were roughly contour-parallel. They took a long-term perspective (>20 years) on how drainage affects vegetation and also undertook a short-term (<2 month) study on water-table behaviour between the ditches. They found that, within 20 years of drains being cut, Sphagnum capillifolium (Ehrh.) Hedw. had disappeared from ditch edges and virtually disappeared from inter-ditch areas away from the edges. For other blanket peatland plant species the effects of ditches appeared to be more local to the drain or to change over time since drainage. For example, in an apparent response to drainage, the cover of Calluna vulgaris (L.) Hull increased and peaked at eight years but later declined to become statistically insignificant. In their hydrological study (summer 1979), Stewart and Lance (1991) found that average water-table depths varied spatially by as much as 15 cm, with the deepest water tables close to the ditches and shallowest towards the mid-points between the ditches. The hydrological data collected by Stewart and Lance (1991) confirmed that at least 20% of the area between drains was affected by the drain (i.e., was drier than would be the case in the absence of the drain). Their S. capillifolium data accord with the hydrological data in that the species disappeared from drain edges where drainage effects were greatest; however, that it also declined substantially in the inter-ditch areas away from drain edges may suggest that drains caused all of the inter-drain area to be drier than would be the case naturally.
In a study of drainage and sheep grazing effects on the abundance of different plant species within a Welsh blanket peatland, Wilson et al. (2010) found some evidence that blanket bog species such as Eriophorum angustifolium Honck were less abundant in a zone extending up to 15 m from drainage ditches but found that drainage effects were partly confounded by the effects of sheep grazing. Wilson et al. (2011) note that their vegetation results are consistent with other work at their study site (Wilson et al. (2011)—see below) that shows that ditches lower water tables in their vicinity.
On a blanket peatland in NE Scotland, Bellamy et al. (2012) looked at drainage and re-wetting effects on vegetation composition using two sites where ditches had been blocked and two where the ditches remained open. Unlike the ditches investigated by Stewart and Lance (1991), the ditches studied by Bellamy et al. (2012) mostly ran downslope. Vegetation composition and cover were measured in transects perpendicular to the long-axes of the ditches. However, while one blocked site showed the patterns expected—more wet indicator species and fewer dry indicators than the open-ditch sites—the other blocked site did not. Bellamy et al. (2012) did not look at water tables at their sites, so it is unclear if the apparent inconsistency between the two blocked sites was caused by hydrological differences.
Most recently, Williamson et al. (2017) investigated vegetation changes in response to drain blocking in a blanket peatland in N Wales (in an area adjacent to the one used for the current study—see “Materials and methods” section below). They found no effects of blocking on vegetation composition and suggested that peat subsidence may explain their findings (see “Discussion and conclusion” section below). They undertook a one-off vegetation survey of the peatland next to open ditches and next to ditches that had been blocked for one, four and five years. Because they did not measure vegetation changes over time they were unable to determine whether factors related to the year in which they did their vegetation survey (e.g., an unusually wet year prior to the survey) masked any effects of drainage.
In all of the above studies, the inter- or between- ditch areas were investigated. In a study of greenhouse gas fluxes from a Welsh blanket peatland, Cooper et al. (2014) noted a distinct change in vegetation over a 27 month period within blocked (infilled) ditches. Over this time Eriophorum vaginatum L. colonised and came to dominate the within-ditch area. However, changes in E. vaginatum abundance were not monitored quantitatively; i.e., numerical estimates of abundance were not made. Peacock et al. (2013) measured changes in the vegetation cover of pools formed by dams on another blanket peatland site in N Wales (the one used for the current study—see "Materials and methods" section below). Eighteen months after their formation, pools had been colonised by a mixture of E. vaginatum (average of 37% cover) and Sphagnum cuspidatum Ehrh. ex Hoffm. (38% cover) (n = 60 pools). Like Cooper et al. (2014), Peacock et al. (2013) did not look at the vegetation in inter-ditch areas.
Work has also been done on drain-blocking effects on the vegetation of both raised bogs and fens, with examples including Komulainen et al. (1999) and Haapalehto et al. (2011). However, it is difficult to gauge how relevant the findings from these studies are to blanket peatlands. Key peat properties may differ between peatland types, and raised bogs, for example, appear to have a higher permeability (hydraulic conductivity) than blanket bogs which will affect the degree to which drains and drain blocking influence water tables (Evans et al. 2014) and, therefore, vegetation.
A number of studies have been conducted on how drain blocking affects blanket peatland water tables. Examples include Holden et al. (2011) and Wilson et al. (2010). Both considered drains that run cross slope and both found the effect of ditch blocking on water tables was statistically significant but relatively modest—of the order of 2–3 cm. Whether such changes are sufficient to cause changes in vegetation composition are unclear. As noted above, for the same sites reported in Wilson et al. (2010, 2011) found some effect of drains on the composition of blanket peatland vegetation, although this finding was partly confounded by sheep grazing effects. This finding suggests that small changes in water-table depth can cause changes in vegetation composition. However, studies on the hydrological regimes of raised and blanket bogs by, respectively, Belyea (1999) and Laine et al. (2007) suggest that water-table depths in different microhabitats or vegetation types vary by at least 5–10 cm. For example, Laine et al. (2007) found that average water-table depths consistently differed by 5–10 cm between lawns and hollows (for microhabitat nomenclature see Belyea and Clymo (2001)) and by 10–15 cm between lawns and hummocks. Therefore, changes in mean water-table depths of a few cm from drain blocking may be insufficient to cause substantial changes in vegetation type or of the relative abundance of a particular vegetation type’s component species. Despite such uncertainties on drain blocking effects, some prominent reviews such as Lindsay (2010) suggest that ditch blocking on blanket bogs will lead to an increase in the abundance of Sphagnum mosses at the expense of cotton grasses. However, there is a lack of data on ditch blocking effects; in particular, there is a lack of fully replicated experimental studies where both vegetation and water tables are studied concurrently and changes over time measured. To help address this research gap we conducted field trials on a UK blanket peatland. We studied the response of Sphagnum spp. and Eriophorum spp. over a five-year period after ditch blocking, and monitored water tables for the first four years. We focused on the inter- or between- ditch zone because, by area, this is the largest part of any ditch-blocked peatland.
Materials and methods
Study site and meteorological conditions
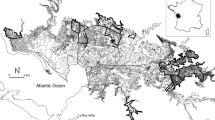
The field trial was carried out on part of the Migneint blanket bog in N Wales, UK (52.97°N, 3.84°W). The field site was located near the top of a north-facing hillslope at c. 500 m above sea level, with a total area of c. 2 ha drained by a set of 12 parallel ditches running approximately downslope, which are believed to have been dug in the 1980s (Fig. 1). The ditches had a mean spacing of 16 m (range 11–26 m), mean slope of 4.5° (range 3.9–5.1°), and a mean monitored length of 99 m (range 84–107 m). They were mostly shallow (50–70 cm deep) and in some cases overgrown by vegetation, although all were found to be hydrologically functional at the start of the experiment. Measured peat depth across the study area ranges from around 0.5 m to 2 m. The peat overlies Cambrian mudstones and siltstones (Lynas 1973) but these appear to be largely hydrologically isolated from the overlying peat. The pre-trial vegetation comprised a typical blanket bog assemblage including Calluna vulgaris, E. vaginatum, and various species of Sphagnum, although the abundance of the latter group was variable and sometimes low (see "Results" section).

Map of the experimental area. Meteorological data were collected using an on-site automatic weather station (AWS), the location of which is also indicated
In February 2011, eight of the 12 ditches were blocked using two different methods, with four open ditches retained as controls. Ditches were assigned to treatments by first placing them into three groups of four based on their measured pre-blocking discharge rate, and then randomly assigning one ditch from each subset into one of the three treatment/control categories. The two ditch-blocking methods used were damming, and reprofiling plus damming (henceforth referred to as ‘reprofiling’). For the dammed treatment, peat dams were constructed at regular intervals along the ditch using peat extracted from ‘borrow pits’ adjacent to the ditch, creating a sequence of pools behind the dams. For ditches in the reprofiling treatment, ditch vegetation was removed, the peat base compressed to destroy any natural pipes (sensu Jones 1981) that may have been present, the ditch partially infilled with peat scraped from inter-ditch areas, and the vegetation replaced. This treatment also involved the construction of peat dams, but pools tended to be shallower or absent due to the partial infilling of the ditch.
Water tables and meteorological conditions were measured up to March 2015. The weather at the site (as measured using an on-site automatic weather station) was highly variable over this period and is summarised in Fig. 2. The annual mean air temperature was 7.8 °C (March 2011–March 2012–Year 1), 6.5 °C (2012/13–Year 2), 6.9 °C (2013/14–Year 3) and 7.6 °C (2014/15–Year 4). In the same period, the rainfall for each project year was 2255, 2409, 1786 and 1936 mm, respectively. Year 3 had a substantially drier summer (Jun, Jul, Aug) than the previous two years. The cold winter of 2012–2013 and very cold early spring of 2013 also stand out, as does the late winter and early spring drought and high temperatures of February–March 2012 (Fig. 2).

Meteorological conditions at the experimental site between 1st March 2011 and 1st March 2015. The black bars indicate daily rainfall. Average daily air temperature is denoted by the green–brown line, daily high temperature by the red dashes and daily low by the blue dashes
Vegetation sampling
The vegetation at the site was monitored using permanent 1 m × 1 m quadrats set up in 2010 before the ditch blocking took place (Fig. 1). Twenty-four permanent quadrats were established 2 m to the east and west of each ditch. These were placed away from those inter-ditch areas where peat was used for damming or infilling of the ditches (see above). Measurements took place once per year at the end of the growing season (late August to early October), and comprised the following:
-
The abundance of all sedges (including the cotton grasses (Eriophora) and Carex nigra (L.) Reichard) and all Sphagnum spp. was estimated using a nested frequency measure. These species were chosen because they are believed to be important controls on methane (CH4) emissions from peatlands (e.g., Lindsay (2010), Green and Baird (2012))—the wider study, of which the vegetation study was a part, was an investigation of how ditch blocking affects blanket peatland CH4 budgets. The measurement of nested frequency involved noting the presence or absence of each target sedge and Sphagnum species in each of the 100 10 cm × 10 cm squares comprising each quadrat. A species was marked as present even if it was not rooted in a 10 cm × 10 cm square. Using this method, a species present in, for example, 46 of the 100 squares, would have a nested frequency of 46% or 0.46. On a species by species basis, this method will give a value that is correlated to cover (and which becomes closer to cover as square size decreases). For any single species, or group of related species, it provides a simple and objective measure that allows abundance to be compared over space (between locations or management treatments) or over time (across successive years after management change). It is, however, unreliable for comparing abundance between different species. For example, a species like Cranberry (Vaccinium oxycoccos L.) has thread-like branches which often form a ‘net’ underneath taller shrubs. Depending on square size, a species such as this could produce a very high nested frequency even though its real cover was low. In contrast, tussocks of sedges such as E. vaginatum might have a relatively high true cover but occupy relatively few 10 cm × 10 cm squares. As such, it makes little sense to compare these species using nested frequency.
-
The ‘vigour’ of the sedges within each quadrat was also recorded by measuring, for each sedge species, the height and maximum width of the ten leaves nearest the centre of the quadrat. Where there were fewer than ten leaves, measurements were still made (and the number of leaves noted).
We grouped the quadrat data on abundance into a sedge group and a Sphagnum group. The sedge group was, in practice, almost entirely made up of E. vaginatum, while the Sphagnum group comprised, in order of abundance: S. capillifolium, Sphagnum fallax (Klinggr.) Klinggr., and Sphagnum papillosum Lindb.. These three species have a tendency to occupy different microhabitats (Clymo 1973), so a case could be made for separating them in any analysis. However, we chose to group them into a single category for two reasons. First, all were relatively sparse at the beginning of the project, and overall Sphagnum abundance can be taken as an indicator of site wetness (with wetter sites often having a higher overall Sphagnum abundance than drier sites). Secondly, although they can occupy different microhabitats in undisturbed sites, the conditions under which they grow well can be quite variable and will depend on facilitation by, and competition with, other species. Thus, while a species may be associated with a particular water-table regime in a natural site, it is by no means clear that the same species will reliably indicate that water-table regime when it colonises a restored site. Wheeler et al. (2002), for example, make note of a study that showed that S. fallax, which is usually associated with hollows and wet lawns, was capable of colonising ‘dried-out’ bogs, this being facilitated by a cover of Polytrichum alpestre Hoppe and E. vaginatum.
Water table measurements
Water tables were measured using dipwells every three to six weeks, with the higher frequency during the growing season. The dipwells were placed 2 m west and east of the ditches (n = 24 wells) and c. 5 m downslope of the permanent quadrats. They were made from 1 m lengths of polyvinyl chloride (PVC) pipe with an outside diameter of 32 mm and a 3.5 mm wall thickness. The pipes were perforated with 8 mm diameter holes drilled at 100 mm intervals along four lines running lengthwise along the pipe, with the holes in each line offset from those in neighbouring lines by 50 mm. The dipwells were tested for their response times and all showed a 90 percent recovery within 120 min during typical water levels, with most recoveries substantially quicker than this.
Data analysis
For the vegetation data we investigated whether blocking treatment (control or open ditches, dammed, or reprofiled), position (east or west of the ditch) and time (Years 0–5; 2010–2015) had any effect on the nested frequency of the sedges and the Sphagna. For each plant group, a factorial, repeated-measures ANOVA was used (in IBM SPSS Statistics version 23). The sedge data were transformed (to the power of four) because they failed the test for normality. Where Mauchly’s test indicated a violation of the sphericity assumption, changes were made to the degrees of freedom using a Greenhouse Geisser correction to reduce the chances of a Type I error. Where p ≤ 0.05, Tukey’s HSD (honest significant difference) test was used to identity which blocking treatments, sides (east or west) or times (year of survey) were significantly different from each other.
The sedge vigour data from the 24 quadrats were collated and graphed according to treatment, position, and year. As noted above, by far the most common sedge at the site was E. vaginatum, with the other recorded species only appearing in a few quadrats and then not all of the time. Therefore, we restricted our attention to the leaf statistics from E. vaginatum. In addition, leaf width in E. vaginatum was found not to vary (it was almost always 0.5 mm or less) so we considered leaf length as the main indicator of vigour. As well as graphing the leaf length data, we investigated the effects of treatment, quadrat position (east or west) and year using the same ANOVA as applied to the nested frequency data.
We separated position (east or west of the ditch) in the vegetation analysis because of uncertainty about which side might show the greater drainage effect in two of the control ditches (Holden et al. 2017). After ditch blocking, water could flow across some blocked ditches towards the control (open ditches). Because the orientation of the ditches was somewhat oblique to the steepest gradient (see Fig. 1), this flow of water meant that there was a possibility the eastern sides of two of the open ditches could become wetter after blocking took place and not act as reliable controls.
To account for differences in the intervals between manual measurements of water-table depth, we used time-weighted annual means in our statistical analysis. This removes biases that may be caused by a higher frequency of readings at one time of year compared to another. We analysed separately how the weighted average annual water tables 2 m to the west and 2 m east of the ditches varied between the treatments and between years (1, 2, 3 and 4: 2011–2014) using a repeated-measures ANOVA (in IBM SPSS Statistics version 23). In a similar fashion we considered the effect of treatment and year on average summer water-table depths. We did so because summer water tables may have a greater effect on vegetation composition than water tables at other times of year. We defined summer water table as the time-weighted average water-table depth for the period spanning June, July and August.
Results
Permanent quadrats: abundance of sedges and Sphagnum spp.
Summary data from the 24 quadrats have been separated according to plant grouping (sedge and Sphagnum) and whether the quadrat was to the east or west of the ditch and are shown in Tables 1, 2, 3 and 4. The data in the tables suggest the following:
-
For the quadrats to the east of the ditches:
-
Sedge: sedges were very abundant across all of the treatments during the period of the experiment. There is the suggestion of an increase in abundance in Year 4 (2014), with differences in abundance between the other years being less obvious.
-
Sphagnum: in general, there were year-on-year increases in nested frequency of Sphagnum spp. in the control and dammed treatments, but no obvious pattern over time for the reprofiled treatment. The initial difference in average Sphagnum abundance between the control and dammed remained over time as abundance in both increased broadly in parallel.
-
-
For the quadrats to the west of the ditches:
-
Sedge: the nested frequency of sedges was consistently high across all years in the control and dammed treatments. The reprofiled treatment had somewhat lower nested frequencies between Year 1 (2011) and Year 3 (2013) than the other treatments.
-
Sphagnum: the nested frequency of Sphagnum spp. increased in all treatments between Year 0 (2010) and Year 5 (2015). Sphagnum abundance was consistently higher in the control (open ditch) treatment than in the two re-wetting treatments. Both re-wetting treatments had a similar level of Sphagnum abundance.
-
The ANOVA test on the sedge abundance data revealed that there was no treatment effect (p = 0.903) or position (east, west) effect (p = 0.983). Time (year of measurement) has a significant influence (p = 0.037) with Year 4 = Year 5 > Year 1 = Year 2 (Year 0 and Year 3 were not significantly different from any other year). There is no interaction effect between time, treatment and position (p = 0.867). The analysis of the Sphagnum nested frequency data revealed no effect of treatment (p = 0.088) or position (p = 0.298). Time has a significant influence (p < 0.0001), with Year 5 = Year 4 > Year 3 > Year 2 = Year 1 > Year 1 = Year 0. There is no interaction effect between time, position and treatment (p = 0.952). In short, these results suggest (a) that drainage/blocking treatment has had no effect on sedge and Sphagnum abundance and (b) that significant inter-annual changes in sedge and Sphagnum abundance can occur.
Vigour (leaf length) of E. vaginatum in the quadrats
The leaf length (vigour) data from the 24 quadrats 2 m either side of each ditch are shown in Figs. 3 and 4. The graphs suggest that leaf length did not vary with treatment or position, but that there is a possible trend of decreasing leaf length over time. The statistical analysis confirms these visual impressions. The repeated-measures ANOVA revealed no treatment (p = 0.945) or position (p = 0.585) effects. Time (year of survey) did have a significant influence (p < 0.0001), with Year 0 = Year 1 = Year 2 > Year 5 = Year 3 = Year 4. There was no interaction effect between time, position and treatment (p = 0.353).

Mean leaf length of Eriophorum vaginatum in the quadrats 2 m to the west of the ditches. Error bars show ±1 SE

Mean leaf length of Eriophorum vaginatum in the quadrats 2 m to the east of the ditches. Error bars show ± 1 SE
Sphagnum abundance increases over time, while sedge vigour is lower in the latter part of the study period which may suggest a relationship or association between the two. We tested this possibility using repeated-measures mixed models in either direction (i.e., Sphagnum abundance and then sedge vigour as the dependent variable). In each case, the vegetation-related independent variable (Sphagnum abundance or sedge vigour) was removed from the model, with year (time) being the dominating driver of variance. Therefore, there does not appear to be an association between the two.
Water-table depths
Water tables at the study site tended to be shallow—mostly within 15–20 cm of the surface. Table 5 summarises the weighted annual average water table depths for the dipwells 2 m east and west of the ditches. The dammed and reprofiled treatments to the east of the ditches had a weighted annual average water-table depth of 8.1 and 8.4 cm, respectively, whilst for the drained treatment (control) the depth was 11.2 cm (Years 1–4). For the dipwells west of the ditches the dammed and reprofiled treatments had a weighted annual average water-table depth of 8.5 and 8.4 cm, respectively, whilst that for the drained treatment (control) was 13.4 cm (Years 1–4). Nevertheless, for both sets of dipwells there was no significant effect of drainage treatment (west p = 0.211; east p = 0.733). In both cases there was an effect of year (west p = 0.001; east p = 0.018). For the west-of-ditch dipwells, water tables in Year 3 were significantly deeper (by c. 1.7 cm) than in Year 2, while for the east-of-ditch dipwells water tables in Year 4 were significantly deeper (by c. 1.1 cm) than in Year 2. When the east and west dipwells are combined, the same trends are, unsurprisingly, apparent: there is no significant effect of drainage treatment (p = 0.197) on water tables, but a significant effect of year (p < 0.0001), with Years 3 and 4 having significantly deeper water tables (by c. 1.3 cm) than Year 2.
Table 6 summarises the weighted summer average water table depths for the dipwells 2 m east and west of the ditches. The dipwells east of the ditches had weighted average summer water-table depths of 12.4, 11.9, and 17.3 cm (dammed, reprofiled, and open (control) respectively; Years 1–4). The equivalent figures for the dipwells to the west were 12.1, 11.6, and 14.2 cm. There was no significant effect of drainage treatment for either east or west dipwells (west: p = 0.209; east p = 0.826). A significant effect of year on summer water tables was, however, apparent (west: p < 0.0001; east: p < 0.0001), with three distinct subsets: Year 3 (west 18.4 cm; east 16.5 cm) = Year 4 (west 16.9 cm; east 15.3 cm) > Year 1 (west 11.6 cm; east 10.8 cm) > Year 2 (west 8.6 cm; east 7.9 cm). When the east and west dipwells are combined there was no significant effect of drainage treatment (p = 0.294). A significant effect of year on the combined dataset was apparent (p < 0.0001), with three distinct subsets: Year 3 (17.4 cm) = Year 4 (16.1 cm) > Year 1 (11.2 cm) > Year 2 (8.3 cm).
Discussion and conclusion
Our data show no evidence that drain blocking leads to changes in sedge or Sphagnum abundance on blanket peatland. Similarly, there were no significant effects of drain blocking on annual average or summer average water tables. Given the water-table results it is not surprising that there is no vegetation effect. The lack of a treatment effect on water tables might seem somewhat surprising because the annual average and summer average water-table means for the dipwells next to the open ditches were consistently (i.e., across all four years) more than 2 cm deeper than for the wells next to the blocked ditches. However, the lack of a treatment effect may, in part, be a reflection of the relatively small sample size (12 wells in total and four per treatment) and high variability among the wells.
Our water table results contrast with those of Holden et al. (2011) and Wilson et al. (2010). Both of those studies found a significant, albeit small (2–3 cm), difference between water tables next to open ditches and blocked ditches. Several explanations may be advanced for why our study differs from these previous investigations. One possibility is sample size as noted above. Another possibility relates to climate. Our site in the Migneint appears to be considerably wetter than the sites used by Holden et al. (2011) and Wilson et al. (2010). As we note above (Materials and methods, Study site and meteorological conditions) we recorded 2255, 2409, 1786 and 1936 mm in each year of the water-table part of the study (Years 1–4). Oughtershaw Moss which was studied by Holden et al. (2011) has an annual average rainfall of 1774 mm but was relatively dry during the period when water tables were monitored by the authors (2005 rainfall: 1567 mm, January to June, 2006 rainfall: 517 mm). Rainfall data for the individual sites studied by Wilson et al. (2010) are not available, although a lower-altitude gauge in the wider catchment has an annual average rainfall of 1635 mm (UK Met Office: goo.gl/QWQGeb; last accessed on 02.09.2016), and the years in which Wilson et al. (2010) took measurements appear to have been close to this average. The effect of a ditch on neighbouring water tables takes time to occur; water has to flow into the ditch to lower the adjacent water table. In wetter climates the soil between ditches will tend to be topped up more regularly with water so that the effect of a ditch on water tables is less clear-cut. Therefore, in wetter climates a drainage effect may be less obvious and more difficult to detect. A third explanation concerns the orientation of drains in the different studies. Our study drains more or less followed the gradient, while those investigated by Holden et al. (2011) and Wilson et al. (2010) were mostly cross-slope. Both types of drain configuration are common in the UK, but their effectiveness in controlling water tables may differ.
We have focused on water-table depths, because of their assumed ecological significance, rather than on how ditches affect the absolute position of the water table relative to a datum. However, changes (over time) to the peat adjacent to ditches may mask or complicate any effects ditches have on water-table depths, especially some years (>5–10 years) after ditch construction. Alterations to the peat alongside a ditch might be expected especially in the period immediately after ditch construction. As water tables fall next to the ditch they expose peat to rapid oxic decay and, in response, the peat may subside so that the peatland surface adjacent to ditches is lower than that midway between them. These possible changes have yet to be studied properly, but fine-scale topographic data from our study site in the Migneint are consistent with such processes occurring: we found the ground-surface midway between ditches was higher than the peat surface 2 m either side of the ditches which, in turn, was higher than the peat surface immediately next to the ditch (Holden et al. 2016, 2017); in other words there was a clear surface slope perpendicular to the ditch (towards the ditch channel). The outcome of such changes in the peatland surface is a reduction in the apparent drainage effect of a long-established ditch when considered in terms of water-table depths. Peat shrinkage brings the peat surface closer to the water level in the ditch, thus reducing water-table depths. Therefore, even though absolute water-table position (i.e. height above a datum) close to a ditch may be considerably lower (10 s of cm) than that midway between ditches, water-table depths may be similar between the two locations. Williamson et al. (2017) have termed this effect ‘self-rewetting’ and use it to explain a lack of effect of ditch blocking on blanket peatland vegetation. When ditches are blocked, the water level may be raised quite substantially in the ditch channel itself but this may have relatively little effect on water tables 2 m either side of the ditch where the sloping peatland surface is the greatest influence on the rate at which water is shed into the ditch and on water-table dynamics. Fine-scale topographic data for the areas between ditches are not reported by Holden et al. (2011) and Wilson et al. (2010) so it is not clear how much this effect may have influenced their data; the subsidence effect will depend on the initial size of the ditch and on the properties of the peat in which the ditch is constructed.
No clear temporal trends in sedge abundance were found, in contrast to the nested frequency of Sphagnum which increased fairly steadily over time. An apparent visual ‘correlation’ of this increase with a decrease in sedge vigour was not borne out in additional statistical analysis, and a change in sedge vigour cannot be used to help explain the Sphagnum trend (cf. Heijmans et al. (2002)). The hydrological analysis shows deeper water tables in Years 3 and 4, and these were the years during which Sphagnum abundance continued to increase, which is perhaps the opposite of what might be expected; it is usually assumed that wetter conditions will favour Sphagnum growth (e.g., Campeau and Rochefort 1996), although lags in plant response to water-table conditions may mean that it is not appropriate to compare plant abundance data directly with water-table data from the same year. Nevertheless, these results also suggest that a focus solely on water table as a control on peatland vegetation is unwise. A wide range of ecological and environmental factors can affect peatland plant growth. For example, Murray et al. (1993) investigated photo-inhibition in Sphagna in both field and laboratory experiments. Sphagnum plants exposed to high-light conditions, even for short periods of time (48 h), showed reduced photosynthetic capacity for periods of at least two weeks. So, even in a generally wet year, a few short periods of intense solar radiation (bright days) might substantially inhibit Sphagnum growth for the rest of the year, thus confounding any relationship between abundance and average water tables. That the water table is not always a primary control on peatland plant distributions may explain the somewhat complicated picture that emerges in the studies by Stewart and Lance (1991) and Bellamy et al. (2012) as reviewed in the Introduction.
Finally, it might be argued that any treatment effects at our study site have not had long enough to be expressed in the vegetation and that a longer period of investigation may reveal responses to blocking. This is, of course, possible, and it is clear from other studies, including Stewart and Lance (1991), that changes in abundance can occur over many years (decades). However, we did find significant inter-annual changes in the two plant groups that we studied, showing that blanket bog vegetation can change substantially between years. And, although we found year-on-year changes in Sphagnum abundance, these were not limited to the drain blocking treatments.
End note
To arrive at this estimate, we used a blanket peatland area of 15,736 km2, assumed a mean peat depth of 1.5 m, a mean dry bulk density of 100 kg m−3, and a mean carbon content of peat dry mass of 50%.
References
Bain CG, Bonn A, Stoneman R, Chapman S, Coupar A, Evans M, Gearey B, Howat M, Joosten H, Keenleyside C, Labadz J, Lindsay R, Littlewood N, Lunt P, Miller CJ, Moxey A, Orr H, Reed M, Smith P, Swales V, Thompson DBA, Thompson PS, Van de Noort R, Wilson JD, Worrall F (2011) IUCN UK commission of inquiry on peatlands. IUCN UK Peatland Programme, Edinburgh
Baird AJ, Holden J, Chapman P (2009) A literature review of evidence on emissions of methane in peatlands. Defra Project SP0574, University of Leeds, Leeds, and Defra, London. Downloadable at: goo.gl/QSkM7L
Bellamy PE, Stephen L, Maclean IS, Grant MC (2012) Response of blanket bog vegetation to drain-blocking. Appl Veg Sci 15:129–135. doi:10.1111/j.1654-109X.2011.01151.x
Belyea LR (1999) A novel indicator of reducing conditions and water-table depth in mires. Funct Ecol 13:431–434
Belyea LR, Clymo RS (2001) Feedback control of the rate of peat formation. P Roy Soc Lond B 268:1315–1321
Campeau S, Rochefort L (1996) Sphagnum regeneration on bare peat surfaces: field and greenhouse experiments. J Appl Ecol 33:599–608
Clymo RS (1973) The growth of Sphagnum: some effects of environment. J Ecol 61:849–869
Cooper MDA, Evans CD, Zielinski P, Levy PE, Gray A, Peacock M, Norris D, Fenner N, Freeman C (2014) Infilled ditches are hotspots of landscape methane flux following peatland re-wetting. Ecosystems 17:1227–1241. doi:10.1007/s10021-014-9791-3
Evans CD, Page SE, Jones T, Moore S, Gauci V, Laiho R, Hruška J, Allott TEH, Billett MF, Tipping E, Freeman C, Garnett MH (2014) Contrasting vulnerability of drained tropical and high-latitude peatlands to fluvial loss of stored carbon. Global Biogeochem Cy 28:1215–1234. doi:10.1002/2013GB004782
Gallego-Sala AV, Prentice CI (2012) Blanket peat biome endangered by climate change. Nature Climate Change 3:152–155. doi:10.1038/NCLIMATE1672
Green SM, Baird AJ (2012) A mesocosm study of the role of the sedge Eriophorum angustifolium in the efflux of methane—including that due to episodic ebullition—from peatlands. Plant Soil 351:207–218. doi:10.1007/s11104-011-0945-1
Haapalehto TO, Vasander H, Jauhiainen S, Tahvanainen T, Kotiaho JS (2011) The effects of peatland restoration on water-table depth, elemental concentrations, and vegetation: 10 years of changes. Restor Ecol 19:587–598
Heijmans MPD, Klees H, Berendse F (2002) Competition between Sphagnum magellanicum and Eriophorum angustifolium as affected by raised CO2 and increased N deposition. Oikos 97:415–425
Holden J, Chapman PJ, Labadz JC (2004) Artificial drainage of peatlands: hydrological and hydrochemical process and wetland restoration. Prog Phys Geog 28:95–123
Holden J, Wallage ZE, Lane SN, McDonald AT (2011) Water table dynamics in undisturbed, drained and restored blanket peat. J Hydrol 402:103–114. doi:10.1016/j.jhydrol.2011.03.010
Holden J, Green SM, Baird AJ, Grayson RP, Chapman PJ, Evans C, Peacock M (2016) The impact of ditch blocking on blanket peatland hydrology. Appendix C in: Green SM, Baird AJ, Evans C, Ostle N, Holden J, Chapman PJ, McNamara N, Investigation of Peatland Restoration (Grip Blocking) Techniques to Achieve Best Outcomes for Methane and Greenhouse Gas Emissions/Balance: Field Trials and Process Experiments Final Report. Defra Project SP1202, University of Leeds, Leeds
Holden J, Green SM, Baird AJ, Grayson RP, Dooling GP, Chapman PJ, Evans CD, Peacock M, Swindles G (2017) The impact of ditch blocking on the hydrological functioning of blanket peatlands. Hydrol Process 31:525–539. doi:10.1002/hyp.11031
Jones JAA (1981) The nature of soil piping: a review of research. GeoBooks (Elsevier), Norwich, p 301
Komulainen V-M, Tuittila E-S, Vasander H, Laine J (1999) Restoration of drained peatlands in southern Finland: initial effects on vegetation change and CO2 balance. J Appl Ecol 36:634–648
Laine A, Wilson D, Kiely G, Byrne KA (2007) Methane flux dynamics in an Irish lowland blanket bog. Plant Soil 299:181–193. doi:10.1007/s11104-007-9374-6
Lindsay R (2010) Peatbogs and carbon: a critical synthesis to inform policy development in oceanic peat bog conservation and restoration in the context of climate change. RSPB Scotland with funding support from Scottish Natural Heritage, Scotland, p 315
Lynas BDT (1973) The Cambrian and Ordovician rocks of the Migneint area, North Wales. J Geol Soc London 129:481–503
Murray KJ, Tenhunen JD, Nowak RS (1993) Control on photosynthesis and production of Sphagnum mosses. Oecologia 96:200–207
Parry LE, Holden J, Chapman PJ (2014) Restoration of blanket peatlands. J Environ Manage 133:193–205
Peacock M, Evans CD, Fenner N, Freeman C (2013) Natural revegetation of bog pools after peatland restoration involving ditch blocking—the influence of pool depth and implications for carbon cycling. Ecol Eng 57:297–301
Stewart AJA, Lance AN (1991) Effects of moor-draining on the hydrology and vegetation of Northern Pennine blanket bog. J Appl Ecol 28:1105–1117
Wheeler BD, Money RP, Shaw SC (2002) Freshwater wetlands. In: Perrow MR, Davy AJ (eds) Handbook of Ecological Restoration, vol 2., Restoration in PracticeCambridge University Press, Cambridge, pp 325–354
Williamson J, Rowe E, Reed D, Ruffino L, Jones P, Dolan R, Buckingham H, Norris D, Astbury S, Evans CD (2017) Historical peat loss explains limited short-term response of drained blanket bogs to rewetting. J Environ Manag 188:278–286
Wilson L, Wilson J, Holden J, Johnstone I, Armstrong A, Morris M (2010) Recovery of water tables in Welsh blanket bog after drain blocking: discharge rates, time scales and the influence of local conditions. J Hydrol 391:377–386
Wilson L, Wilson JM, Johnstone I (2011) The effect of blanket bog drainage on habitat condition and on sheep grazing, evidence from a Welsh upland bog. Biol Conserv 144:193–201
Acknowledgements
The UK Government’s Department for Environment, Food and Rural Affairs (Defra) funded the research under grant SP1202 (goo.gl/H45Uuk). We are grateful to The National Trust for giving permission to work at the site, for paying for the experimental blocking and for providing assistance with setting up the experiment. The Countryside Council for Wales (now part of Natural Resources Wales) are also thanked for funding the vegetation survey.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Green, S.M., Baird, A.J., Holden, J. et al. An experimental study on the response of blanket bog vegetation and water tables to ditch blocking. Wetlands Ecol Manage 25, 703–716 (2017). https://doi.org/10.1007/s11273-017-9545-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11273-017-9545-z