
Abstract
Aims
During the first days after harvest of Lolium perenne L., N remobilized from roots and stubble forms the main N source for regrowth. Low N uptake from the soil during this period may lead to N loss if N fertilizer is applied too soon. Furthermore, temporary N deprivation has been found to stimulate root growth. We therefore hypothesized that a strategic delay in N application after harvest may improve N-use efficiency of L. perenne grassland by increasing root biomass and reducing N loss.
Methods
In a laboratory and field experiment with L. perenne, we delayed N fertilizer application after harvest for 0, 3, 6, 9 and 12 days, repeated this for up to six harvest cycles, and determined effects on herbage yield, herbage N uptake and root biomass.
Results
In both experiments, delaying N application for up to 12 days had no significant effect on root biomass or total herbage N uptake, but it significantly reduced total herbage yield in the laboratory experiment. Total yield tended to be highest when N application was delayed for 3 days. Two growth periods in the field experiment showed significantly higher N uptake when N application was delayed, possibly due to rainfall-induced N losses in the treatments with shorter delay.
Conclusions
Our results do not provide evidence that delaying N application improves N-use efficiency of L. perenne grassland by increasing root biomass. However, strategic timing of N fertilizer application based on rainfall forecasts could contribute to improve N-use efficiency by reducing N losses from leaching and denitrification.
Similar content being viewed by others


Introduction
Root characteristics, such as rooting density and rooting depth, play a critical role in the interception and uptake of nitrogen (N) by plants, including grasses (Atkinson et al. 2005). Higher N uptake results in higher N-use efficiency (at a given amount of available N), may increase herbage yield, and reduces environmental pollution from nitrate leaching (Kristensen and Thorup-Kristensen 2004; Crush et al. 2007), denitrification (Misselbrook et al. 2014) and volatilization (Bouwman et al. 2002; Smith et al. 2012).
The N-use efficiency of inorganic fertilizers on grassland is generally assumed to be 80–100 % (www.rvo.nl); however, in reality it is often as low as 60 to 65 % on sandy soils (Schröder et al. 2007; van Eekeren et al. 2010a). This means that up to 40 % of applied fertilizer N can be lost. Specific N losses for commonly used nitrate based fertilizers (e.g. calcium ammonium nitrate) applied to grassland are estimated to be approximately 2 % for NH3 emission (Misselbrook et al. 2000), 1 % for N2O emission (Smith et al. 2012; Misselbrook et al. 2014) and 6 % for NO3 leaching (Stark and Richards, personal comm.). Heavy rainfall just after fertilizer application can increase losses through these pathways; for instance, Esala and Leppänen (1998) reported a NO3-loss of 28 % of applied NO3-N when heavy rainfall was simulated one day after fertilizer application. De Klein and van Logtestijn (1994) reported that denitrification losses strongly increased when fertilizer was applied on a wet soil, ranging between 6 and 9 % of N applied.
Strategic timing of N fertilizer application during the growing season can be a valuable management tool to reduce N loss and increase N-use efficiency of grassland, by synchronizing N supply with N demand (Di and Cameron 2002). Well-known and well-researched practices are the timing of the first fertilizer application in Spring (Stevens et al. 1989), the strategic distribution of fertilizer applications over the growing season (Reid 1984; Bittman and Kowalenko 2000), and the timing of the final fertilizer application towards the end of the growing season (Laidlaw et al. 2000; O’Donovan et al. 2004).
Less is known about the optimal timing of N application after a grass harvest event (by grazing or cutting). In practice, it is often recommended (Sheldrick et al. 1994) that the time between harvest and fertilizer application should be minimized to immediately boost grass regrowth and maximize utilization of fertilizer N before the next harvest. However, various grass defoliation experiments have shown that during the first days of regrowth, most of the N comes from remobilized organic N (proteins) stored in the stubble and roots, and not from current root uptake of fertilizer N. For example, Ourry et al. (1989) and Thornton et al. (1993) found that the contribution of remobilized N to total N in new leaves of Lolium perenne L. was 70 % after one week and 40 to 45 % after two weeks of regrowth. Low uptake of fertilizer N during the first week after application increases the risk of N loss to the environment, especially with heavy rainfall.
Additionally, there is evidence that delayed N application after harvest may stimulate root growth. Ennik and Hofman (1983) found an increase in root volume of Lolium multiflorum Lam. grown in nutrient solution, when N supply was exhausted four days prior to harvest and the following renewal of the nutrient solution. Jarvis and Macduff (1989) found an increased root growth rate when L. perenne, grown in flowing solution culture, was deprived of nitrate-N for 11 days.
Based on this evidence, it can be hypothesized that temporary N deprivation by delaying N application after a harvest event may increase root biomass, and possibly also rooting depth, of L. perenne. If this hypothesis can be proven under field conditions, farmers could use delayed N application after harvest as a management strategy to increase root growth, and hence N uptake and herbage yield in grasslands. Increased root growth and rooting depth may also reduce drought sensitivity of grasslands by giving grass plants access to a larger soil volume and thus to potentially more soil moisture (Kramer and Coile 1940; Garwood and Williams 1967).
So far, only few field studies have been conducted on using delayed N application after grass harvest as a management practice to increase N uptake and herbage yield of permanent grassland. Research by Wolton (1973) on permanent grasslands in the UK showed that herbage yield was highest when N application was delayed for 1 day, as compared to 8 or 15 days. In another UK field study, Sheldrick et al. (1994) found that a delay of more than 10 days had a significant (P < 0.01) negative effect on herbage yield. Nitrogen-use efficiency was significantly (P = 0.001) higher after a 7-day delay compared to a delay for 0, 3 or 14 days. In the latter study, experimental plots were irrigated after N application to avoid effects of drought on fertilizer N availability. The effect of delayed N application on root characteristics was not determined in either of these studies.
The objectives of the present study were to assess the effects of delaying N application after harvest on root biomass, herbage N uptake and herbage yield of L. perenne, where herbage N uptake was used as a proxy for N-use efficiency. These effects were assessed both in a laboratory column experiment using intact cores taken from permanent grassland, as well as in a field experiment on permanent grassland. Our hypothesis was that delaying N application after harvest increases L. perenne root biomass, resulting in increased N uptake and possibly also herbage yield.
Materials and methods
Experimental design
For the laboratory column experiment, intact grassland soil cores were taken from an L. perenne sward (four years old, cutting only) on a sandy soil (Typic Haploquod [USDA]) in the Netherlands in June 2010 (van Eekeren et al. 2010b). Cores were collected by hammering open-end PVC tubes (400 mm long, 115 mm diameter) into the soil and then digging them out, keeping soil and tube together. Cores were placed in a controlled growth room (day: 16 h of 20°C; night: 8 h of 18°C; 80 % relative humidity; 85 W m−2 light intensity). After one week, grass regrowth was cut to 6 cm height, as a ‘clearing cut’ before the start of the experiment. Treatments consisted of N fertilizer application, delayed for either 0, 3, 6, 9 or 12 days following the clearing cut (first growth period) or harvest cut (second, third and fourth growth period, each period lasting 21 days). The experiment was carried out in four replicates per treatment, for a total of 20 cores.
The incubated grassland cores were fertilized with N at a rate equivalent to 100 kg N ha−1 for each growth period, resulting in a total application of 400 kg N ha−1 over four growth periods. Applications of P, K and S were also equally distributed over these periods, at a total rate equivalent to 34, 498 and 20 kg ha−1, respectively. Nutrients were added via a stock solution, containing 32 ml of 1 M (NH4)2SO4, 59 ml of 1 M (NH4)2HPO4, 662 ml of 1 M KNO3, 80 ml of 1 M NH4NO3 and 482 ml of 1 M NH4Cl in 2 L of de-ionized water. Weight loss of the cores by evapotranspiration was compensated daily using de-ionized water. Furthermore, every four days a 10 mm rainfall event was simulated by adding the equivalent amount of water (for a total of 20 rainfall events over the entire experiment).
In the field experiment we tested the same treatments as in the laboratory experiment, i.e. N applications delayed for either 0, 3, 6, 9 or 12 days following a harvest event. The field experiment had a randomized block design with six replicate plots (10 × 2.5 m) in six blocks, with each block located in a different field of permanent L. perenne grassland on a drought-sensitive sandy soil in the southern part of the Netherlands (51°32’N, 5°51’E). Ranges of basic reference soil properties of the different fields were (0–10 cm depth): clay 2–5 %, silt 6–24 %, fine sand 39–55 %, coarse sand 23–51 %, organic matter 2.3–6.3 %, bulk density 1.23–1.52 kg L−1 and pH-KCl 4.7–5.8.
The experiment included six harvest cycles (growth periods), with 4 to 6 weeks between each harvest. N applications over the six growth periods were 80, 60, 60, 50, 40 and 30 kg N ha−1, respectively, resulting in a total application of 320 kg N ha−1. N fertilizer was applied as calcium ammonium nitrate (CAN, 27 % N) and evenly distributed over each experimental plot with a custom-made, small-scale fertilizer spreader (Europart, Austria). Fertilizer application in the control (0-day delay) treatments took place on March 2, April 29, June 1, July 4, August 16 and September 23 of 2011. Plots were not additionally fertilized with phosphorus (P) or potassium (K), because soil concentrations of plant-extractable P and K were considered to be sufficient: ammonium lactate-extractable P (P-Al) (Reijneveld et al. 2014) ranged from 16 to 31 mg P 100 g−1 dry soil, and 0.01 M CaCl2 extractable K (Houba et al. 1990) ranged from 61 to 134 mg K kg−1 dry soil. Additional fertilization with S was not considered in the field experiment. Experimental plots within one field (block) were all harvested at the same date, irrespective of the number of days by which N application had been delayed. Block 1 was harvested one to two weeks later than the other blocks during the entire experimental period, due to lagging grass growth.
In contrast to our laboratory experiment and the field experiment by Sheldrick et al. (1994), we did not irrigate our field plots after fertilizer application, as this is not common farming practice in the Netherlands. To take potential rainfall effects on N availability into account, daily rainfall data from the nearest weather station (5 to 10 km away) were collected from the Dutch Royal Meteorological Institute (KNMI, Bilthoven, the Netherlands).
Measurements
In the laboratory experiment, herbage yield was determined by cutting the grass to 6 cm height with scissors. In the field experiment, plots were mowed with a Haldrup grass harvester (J. Haldrup a/s, Løgstør, Denmark), also to a cutting height of 6 cm. The harvested material was weighed and sampled to determine dry weight (24 h at 70°C) and total N content (to calculate herbage N uptake). In the laboratory experiment, total N was determined using the micro-Kjeldahl method (Houba et al. 1997); in the field experiment, a Dumas-based method (NEN 16634–1 2008) was used.
In both experiments, roots were sampled after the final harvest. In the laboratory experiment, root samples were taken by removing the cores from their PVC tubes and dividing the soil into four layers: 0–10, 10–20, 20–30 and 30–40 cm depth. One tube was excluded from root analysis due to damage caused by Melolontha melolontha larvae. In the field experiment, roots were sampled by taking three soil cores (8.5 cm diameter) from three soil layers (10–20, 20–30 and 30–40 cm depth) in each plot, using a root auger (Eijkelkamp, Giesbeek, the Netherlands). These cores were pooled into one sample per soil layer per plot for further analysis. We did not sample the top layer (0–10 cm depth) because previous field measurements had shown large random variation in root biomass and root length density in the 0–10 cm layer (Deru et al. 2012) and because we were primarily interested in treatment effects on deeper rooting. Furthermore, root samples were not taken in block 1 and 2, due to lagging grass growth and hence divergent harvest dates (block 1), or due to damage caused by Elateridae larvae (block 2).
All soil cores were carefully washed through a 2 mm mesh screen, and non-root particles were hand-picked from the washed samples. Samples were then oven-dried (24 h at 70 °C) to determine dry weight and calculate root biomass (kg DM ha−1). Root samples from the laboratory experiment were also analyzed for root length density, prior to drying. To this end, root length per diameter class was measured by image analysis using WinRHIZO-software (Bouma et al. 2000) (settings: scan resolution 400 dpi; filtering out of particles smaller than 0.2 cm2 and with a length/width ratio lower than 6; twenty diameter classes: from 0.00 to 0.95 mm in steps of 0.05 mm, and >0.95 mm). From this data, root length density (root length per soil volume, cm cm−3) and mean root diameter (mm) were calculated.
Statistical analysis
Treatment effects on herbage yield, herbage N uptake and root characteristics were statistically analyzed using ANOVA in Genstat software (17th edition). Herbage yield and N uptake data were analyzed per harvest event and for all harvests together. Root biomass, root length density (measured in the laboratory experiment only) and mean root diameter (measured in the laboratory experiment only) were analyzed per soil layer and for all soil layers together.
Results
Root biomass
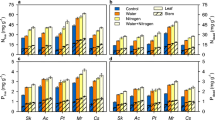
In the laboratory experiment, root biomass (as measured after four growth periods) was on average 3888, 631, 340 and 408 kg DM ha−1 in soil layers 0–10, 10–20, 20–30 and 30–40 cm, respectively. Delaying N application had no significant effect (P > 0.05) on root biomass (Fig. 1, Table 1), root length density or mean root diameter (data not shown). In the field experiment, root biomass (as measured after six growth periods) was on average 327, 182 and 118 kg DM ha−1 in soil layers 10–20, 20–30 and 30–40 cm, respectively. Although the data suggest a positive trend (Fig. 1), delayed N application had no significant effect (P > 0.05) on total root biomass (soil layer 10–40 cm). A significant treatment effect on root biomass was only found for soil layer 20–30 cm (P = 0.03), where root biomass was significantly higher after a 12-day delay when compared to shorter delays. Neither in the laboratory nor the field experiment a significant effect on root biomass in the deepest soil layer (30–40 cm) was detected, suggesting that delaying N application had no effect on rooting depth.

Root biomass of L. perenne (kg DM ha−1) in soil layers 10–20, 20–30 and 30–40 cm, as affected by delaying N application after harvest for 0, 3, 6, 9, or 12 days. Root biomass was measured either after four (laboratory experiment) or six (field experiment) harvest cycles. Error bars represent standard errors of total root biomass (soil layer 10–40 cm). Different letters in treatment averages within a soil layer indicate a significant difference (P < 0.05). NB root biomass in soil layer 0–10 cm not included. For the root biomass in soil layer 0–40 cm of the laboratory experiment, see Table 1
Herbage yield
In the laboratory experiment, delaying N application was found to have a significant (P < 0.01) negative effect on total herbage yield (cumulated over four harvests) (Table 1). Total herbage yield was highest after a 3-day delay (19,848 kg DM ha−1), but was significantly reduced when N application was delayed for 6 days or more, with lowest yields observed after a 12-day delay (12,133 kg DM ha−1). A similar, though non-significant, trend was observed in the field experiment (Table 2), where total herbage yield (cumulated over six harvests) was highest when N application was delayed for 3 days (11,446 kg DM ha−1) and lowest when it was delayed for 12 days (10,530 kg DM ha−1). Herbage yield at the second and third harvest tended to be lower when N application after the previous harvest had been more delayed, showing significantly lower values after a 12-day delay compared to a 0-day or 3-day delay in the third growth period. In contrast, herbage yield at the fourth and fifth harvest tended to be higher when N application had been delayed: at the fifth harvest, herbage yield was significantly higher for all delay treatments (except the 9-day delay), compared to the control (0-day delay).
Herbage N uptake
In the laboratory experiment, total herbage N uptake (cumulated over four growth periods) ranged from 247 to 304 kg N ha−1 (Table 1), and was not significantly affected by delaying N application (P = 0.19). Similarly, in the field experiment, no significant effect (P = 0.65) of delay treatments on total herbage N uptake (cumulated over six harvests) was detected (Table 3). However, when analyzing the data for individual growth periods, we found that N uptake during the fourth and fifth period was significantly higher (P = 0.01) when N application had been delayed for 12 days (fourth growth period) or 3 days or more (fifth growth period) (Table 3).
Rainfall in the field experiment
Rainfall varied considerably over the course of the field experiment (Table 4). The second growth period was particularly dry, with only 2 mm of rainfall during the first 12 days and only 10 mm of rainfall during the entire period. In contrast, the fourth growth period was very wet, with relatively heavy rainfall on the sixth, eighth and tenth day after harvest, and a total rainfall of 146 mm. The fifth growth period was also relatively wet and had unusually heavy rainfall (29 mm) on the first day after harvest.
Discussion
Effect of delayed N application on root biomass
In contrast to our hypothesis, we could not detect a significant effect of delayed N application on L. perenne root biomass, neither in the laboratory nor in the field experiment. The results obtained by Ennik and Hofman (1983) and Jarvis and Macduff (1989) with Lolium grown in nutrient solutions could therefore not be replicated in soil-based systems. The difference in results between these studies and our study can be attributed to differences in experimental conditions.
Firstly, root growth under suboptimal field conditions is likely to differ from root growth in nutrient solutions under optimal conditions (Davidson 1969; Powell 1977). Physical obstruction by soil particles (Houlbrooke et al. 1997; Steingrobe et al. 2000) may have prevented a rapid increase in root growth and root volume such as observed by Ennik and Hofman (1983) in nutrient solution.
Secondly, in the experiments by Ennik and Hofman (1983) and Jarvis and Macduff (1989) treatments consisted of controlled interruptions of a continuous N supply. This was not the case in our experiments, where N was supplied only at the start of each growth period. It is possible that this source of N was exhausted before the next harvest, particularly in treatments where N application was delayed for only a few days (leaving more days for N uptake before the next harvest) and in the second half of the field experiment when fertilizer application levels per growth period were lower. In the latter case, all delay treatments may have experienced N shortage, masking effects of delayed N application on root biomass. In the laboratory experiment, average N uptake between the moment of N application and harvest varied between 7.2 and 9.7 kg N ha−1 day−1 in the 12-day delay treatment. This level of uptake rates suggests that added N could indeed be depleted before the next harvest and that N shortage could arise sooner in treatments with a short delay compared to treatments with a long delay.
A third explanation for the absence of treatment effects on root biomass in our experiments could be that effects arising early in the experiment had disappeared by the end of the experiment, when root samples were taken. However, this seems unlikely, as Ennik and Hofman (1983) found that increases in root biomass were larger after repeated intervals of N deprivation. Based on these results, effects of delayed N application on root biomass should have increased, rather than disappeared, over the course of our experiments, given that delays in N application were repeated four (laboratory experiment) to six times (field experiment).
Finally, root biomass is a rather general indicator of root presence and activity, as it includes young, old and dead roots. In our experiments, relatively small changes in root growth arising from delayed N application may have been masked by the large standing root biomass in the grassland cores and field plots, as compared to a relatively small standing biomass in the nutrient solution experiments of Ennik and Hofman (1983) and Jarvis and Macduff (1989). The use of the ingrowth-core method (Steingrobe et al. 2000) might be more suitable to detect relatively small changes in root biomass, as this method allows to measure newly developed roots (root production) in the root zone.
Effect of delayed N application on herbage yield and herbage N uptake
In the field experiment, a delay of N application after harvest for up to 12 days had no significant (P > 0.05) effect on total herbage yield and total herbage N uptake over six growth periods. However, the results do suggest that total herbage yield was highest when N application was delayed for 3 days, with yields decreasing only when this delay was 6 days or more (Table 2). This trend was not observed for herbage N uptake. Field experiments in the UK by Wolton (1973), carried out at three different sites during three growing seasons with 28-day harvest cycles, showed that herbage yield was highest after a 1-day delay compared to an 8- or 15-day delay in N application. The effect of a 3-day delay was not tested in Wolton’s (1973) study, but their conclusion was that N fertilizer most effectively increased herbage yield when applied within 8 days after harvest. In another UK study, carried out at one location during one growing season with 42-day harvest cycles, Sheldrick et al. (1994) found that a delay of up to 10 days had no significant (P > 0.05) effect on herbage yield. Nitrogen-use efficiency, however, was significantly higher after a 7-day delay compared to a 0-day, 3-day, or 14-day delay. In the latter study, irrigation after N application reduced the effect of rainfall events on N availability. The findings of Sheldrick et al. (1994) are in agreement with the results of Ourry et al. (1989) and Thornton et al. (1993), who found that only a small amount of N is taken up from the soil during the first week of regrowth, implying that fertilizer N added within 7 days after harvest is susceptible to loss, reducing N-use efficiency. Without irrigation and with N applied under dry conditions, a rainfall event may kick-start N uptake in all treatments simultaneously, reducing treatment effects of delayed N application on N-use efficiency. This may have played a role in our experiment.
In contrast to our field experiment, our laboratory experiment showed a significant negative effect on herbage yield when N application after harvest was delayed for 6 days or more. This difference in results may be explained by the shorter harvest cycles in the laboratory experiment (three weeks) compared to the field experiment (four to six weeks), which potentially increased the impact of delayed N application in the former: for example, a 12-day delay reduced the number of days available to grass plants to utilize fertilizer N before the next harvest by 34 %, compared to 57 % in the laboratory experiment. The impact of this reduction was potentially exacerbated by the more favourable growing conditions (higher N application rate, temperature, light intensity) in the laboratory experiment compared to the field experiment.
Effect of rainfall on herbage yield and herbage N uptake in the field experiment
Heavy or prolonged rainfall after a fertilizer application may result in N leaching, leading to lower N uptake and lower herbage yield (Mancino and Troll 1990; Adomaitis et al. 2008). In our field experiment, rainfall patterns may thus have influenced the effects of delayed N application on herbage yield and N uptake, particularly during the first 12 days of each growth period. For example, in the fourth growth period, rainfall was relatively high on days 6, 8 and 10 (Table 4), coinciding with increased N uptake in treatments where N application was delayed to day 9 and 12 (Table 3). Possibly, leaching potential was lower on the latter days (particularly day 12), resulting in significantly higher N uptake. Rainfall effects may have played an even larger role during the fifth growth period, when both N uptake and herbage yield were significantly higher in all delay treatments, compared to the control (0-day delay). This effect may be explained by the extremely heavy rainfall on the first day of this growth period, which may have caused significant leaching of the N applied immediately after harvest (0-day delay treatment). The difference in N uptake between the treatment with a 0-day delay and treatments with longer delay suggest that at least 10 kg of the 40 kg N applied on day 0 was lost for plant uptake (Table 3).
Given the influence of rainfall on the fate of fertilizer N applied under field conditions, effects of delayed N application will likely depend on the fertilizer formulation used. In our field experiment we used CAN, which contains 50 % ammonium-N and 50 % nitrate-N. Wolton (1973) and Sheldrick et al. (1994) did not report the type of N fertilizer used in their experiments. As nitrate is more sensitive to leaching than urea or ammonium (Mancino and Troll 1990; Adomaitis et al. 2008), fertilizer formulation may influence the effect of delayed N application, particularly when rainfall is high.
Finally, the relatively large effects of rainfall events on herbage N uptake and herbage yield observed in our experiment suggest that N-use efficiency and herbage yield of grassland may be increased by minimizing the risk of fertilizer N leaching. Weather predictions should therefore be taken into account to avoid N application right before heavy or prolonged rainfall. Furthermore, using fertilizer formulations with a higher percentage of ammonium during relatively wet periods of the growing season could increase herbage yield and N-use efficiency, while reducing environmental pollution from N leaching and possibly also denitrification (de Klein and van Logtestijn 1994).
Conclusions
In both our experiments, a delay in N application of up to 12 days after harvest had no significant effect on L. perenne root biomass or total N uptake, but did significantly reduce total herbage yield in the laboratory experiment, possibly due to the relatively short harvest cycles. The observed positive effects of delayed N application on N uptake and herbage yield in the fourth and fifth growth period of our field experiment appear to be the result of the avoidance of leaching, caused by heavy rainfall shortly after fertilizer N application. This suggests that strategic timing of N application based on rainfall forecasts could contribute to reduce N losses from leaching.
References
Adomaitis T, Vaisvila Z, Mazvila J, Staugaitis G, Fullen MA (2008) Influence of mineral fertilizer on nitrogen leaching. Acta Agric Scand Sect B Soil Plant Sci 58:199–207. doi:10.1080/09064710701593012
Atkinson D, Black KE, Dawson LA, Dunsiger Z, Watson CA, Wilson SA (2005) Prospects, advantages and limitations of future crop production systems dependent upon the management of soil processes. Ann Appl Biol 146:203–215. doi:10.1111/j.1744-7348.2005.040061.x
Bittman S, Kowalenko CG (2000) Within-season grass herbage crude-protein- and nitrate-N concentrations as affected by rates and seasonal distribution of fertilizer nitrogen in a high yearly rainfall climate. Can J Plant Sci 80:277–285
Bouma TJ, Nielsen KL, Koutstaal B (2000) Sample preparation and scanning protocol for computerised analysis of root length and diameter. Plant Soil 218:185–196. doi:10.1023/A:1014905104017
Bouwman AF, Boumans LJM, Batjes NH (2002) Estimation of global NH3 volatilization loss from synthetic fertilizers and animal manure applied to arable lands and grasslands. Glob Biogeochem Cycles 16:8–1–8-14. doi:10.1029/2000gb001389
Crush JR, Easton HS, Waller JE, Hume DE, Faville MJ (2007) Genotypic variation in patterns of root distribution, nitrate interception and response to moisture stress of a perennial ryegrass (Lolium perenne L.) mapping population. Grass Forage Sci 62:265–273. doi:10.1111/j.1365-2494.2007.00583.x
Davidson RL (1969) Effects of soil nutrients and moisture on root/shoot ratios in Lolium perenne L. and Trifolium repens L. Ann Bot 33:571–577
De Klein CAM, van Logtestijn RSP (1994) Denitrification in the top soil of managed grasslands in the Netherlands in relation to soil type and fertilizer level. Plant Soil 163:33–44
Deru JGC, van Eekeren N, de Boer HC (2012) Rooting density of three grass species and eight Lolium perenne varieties. In: Grassland - a European resource? 24th General meeting of the European Grassland Federation. Lublin, Poland, pp. 604–606
Di HJ, Cameron KC (2002) Nitrate leaching in temperate agroecosystems: sources, factors and mitigating strategies. Nutr Cycl Agroecosyst 64:237–256. doi:10.1023/A:1021471531188
Ennik GC, Hofman TB (1983) Variation in the root mass of ryegrass types and its ecological consequences. Neth J Agric Sci 31:325–334
Esala M, Leppänen A (1998) Leaching of 15N-labeled fertilizer nitrate in undisturbed soil columns after simulated heavy rainfall. Commun Soil Sci Plant Anal 29:1221–1238. doi:10.1080/00103629809370022
Garwood EA, Williams TE (1967) Soil water use and growth of a grass sward. J Agric Sci 68:281–292
Houba VJG, Novozamsky I, Lexmond TM, van der Lee JJ (1990) Applicability of 0.01 M CaCl2 as a single extraction solution for the assessment of the nutrient status of soils and other diagnostic purposes. Commun Soil Sci Plant Anal 21:2281–2290. doi:10.1080/00103629009368380
Houba VJG, van der Lee JJ, Novozamsky I (1997) Soil and plant analysis. Part 1: soil analysis procedures. Department of Soil Quality, Wageningen University, the Netherlands
Houlbrooke DJ, Thom ER, Chapman R, McLay CDA (1997) A study of the effects of soil bulk density on root and shoot growth of different ryegrass lines. N Z J Agric Res 40:429–435
Jarvis SC, Macduff JH (1989) Nitrate nutrition of grasses from steady-state supplies in flowing solution culture following nitrate deprivation and or defoliation. 1. Recovery of uptake and growth and their interactions. J Exp Bot 40:965–975
Kramer PJ, Coile TS (1940) An estimation of the volume of water made available by root extension. Plant Physiol 15:743–747
Kristensen HL, Thorup-Kristensen K (2004) Root growth and nitrate uptake of three different catch crops in deep soil layers. Soil Sci Soc Am J 68:529–537
Laidlaw AS, Watson CJ, Mayne CS (2000) Implications of nitrogen fertilizer applications and extended grazing for the N economy of grassland. Grass Forage Sci 55:37–46
Mancino CF, Troll J (1990) Nitrate and ammonium leaching losses from N-fertilizers applied to ‘Pencross’ creeping bentgrass. Hortscience 25:194–196
Misselbrook TH, van der Weerden TJ, Pain BF, Jarvis SC, Chambers BJ, Smith KA, Phillips VR, Demmers TGM (2000) Ammonia emission factors for UK agriculture. Atmos Environ 34:871–880
Misselbrook TH, Cardenas LM, Camp V, Thorman RE, Williams JR, Rollett AJ, Chambers BJ (2014) An assessment of nitrification inhibitors to reduce nitrous oxide emissions from UK agriculture. Environ Res Lett 9:115006
NEN 16634-1 (2008) Food products - Determination of the total nitrogen content by combustion according to the Dumas principle and calculation of the crude protein content - Part 1: Oilseeds and animal feeding stuffs (www.nen.nl). Delft, the Netherlands
O’Donovan M, Delaby L, Stakelum G, Dillon P (2004) Effect of autumn/spring nitrogen application date and level on dry matter production and nitrogen efficiency in perennial ryegrass swards. Irish J Agri Food Res 43:31–41
Ourry A, Bigot J, Boucaud J (1989) Protein mobilization from stubble and roots, and proteolytic activities during post-clipping re-growth of perennial ryegrass. J Plant Physiol 134:298–303
Powell CL (1977) Effect of phosphate fertilizer and plant density on phosphate inflow into ryegrass roots in soil. Plant Soil 47:383–393. doi:10.1007/bf00011497
Reid D (1984) The seasonal distribution of nitrogen-fertilizer dressings on pure perennial ryegrass swards. J Agric Sci 103:659–669
Reijneveld A, Termorshuizen A, Vedder H, Oenema O (2014) Strategy for innovation in soil tests illustrated for P tests. Commun Soil Sci Plant Anal 45:498–515. doi:10.1080/00103624.2013.863909
Schröder JJ, Uenk D, Hilhorst GJ (2007) Long-term nitrogen fertilizer replacement value of cattle manures applied to cut grassland. Plant Soil 299:83–99. doi:10.1007/s11104-007-9365-7
Sheldrick RD, Lavender RH, Martyn TM (1994) Effects of delay in reapplication of nitrogen-fertilizer following cutting silage from a ryegrass sward. Grass Forage Sci 49:369–371. doi:10.1111/j.1365-2494.1994.tb02012.x
Smith KA, Dobbie KE, Thorman R, Watson CJ, Chadwick DR, Yamulki S, Ball BC (2012) The effect of N fertilizer forms on nitrous oxide emissions from UK arable land and grassland. Nutr Cycl Agroecosyst 93:127–149
Steingrobe B, Schmid H, Claassen N (2000) The use of the ingrowth core method for measuring root production of arable crops - influence of soil conditions inside the ingrowth core on root growth. J Plant Nutr Soil Sci 163:617–622. doi:10.1002/1522-2624(200012)163:6<617::aid-jpln617>3.0.co;2-0
Stevens RJ, Gracey HI, Kilpatrick DJ, Camlin MS, O’Neill DG, McLaughlan W (1989) Effect of date of application and form of nitrogen on herbage production in spring. J Agric Sci 112:329–337
Thornton B, Millard P, Galloway S (1993) The effects of temperature and form of nitrogen supply on relative contribution of root uptake and remobilization in supplying nitrogen for laminae regrowth of Lolium perenne L. J Exp Bot 44:1601–1606. doi:10.1093/jxb/44.10.1601
Van Eekeren N, de Boer H, Hanegraaf MC, Bokhorst JG, Nierop D, Bloem J, Schouten T, de Goede TGM, Brussaard L (2010a) Ecosystem services in grassland associated with biotic and abiotic soil parameters. Soil Biol Biochem 42:1491–1504. doi:10.1016/j.soilbio.2010.05.016
Van Eekeren N, Bos MM, de Wit J, Keidel H, Bloem J (2010b) Effect of individual grass species and grass species mixtures on soil quality as related to root biomass and grass yield. Appl Soil Ecol 45:275–283. doi:10.1016/j.apsoil.2010.05.003
Wolton KM (1973) Effect of time of N application in a growth period on herbage production. J British Grassland Soc 28:181–182
Acknowledgments
The authors acknowledge Jaap Nelemans and Marc Kroonen for their assistance in the laboratory and field experiment, respectively; Riekje Bruinenberg and Hans Dullaert for their assistance with root biomass measurements; and Jan de Wit and Christien Ettema for their valuable comments on an earlier version of this paper.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Funding
This work was financed by the Dutch Ministry of Infrastructure and Environment (Innovatieprogramma Kaderrichtlijn Water) (laboratory experiment) and the Dutch Dairy Board (field experiment).
Conflict of interest
The authors declare they have no conflict of interest
Additional information
Responsible Editor: Rafael S. Oliveira.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
de Boer, H., Deru, J., Hoekstra, N. et al. Strategic timing of nitrogen fertilization to increase root biomass and nitrogen-use efficiency of Lolium perenne L.. Plant Soil 407, 81–90 (2016). https://doi.org/10.1007/s11104-016-2917-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-016-2917-y