
Abstract
Background
In the Mediterranean climate, plants have evolved under conditions of low soil-water and nutrient availabilities and have acquired a series of adaptive traits that, in turn exert strong feedback on soil fertility, structure, and protection. As a result, plant-soil systems constitute complex interactive webs where these adaptive traits allow plants to maximize the use of scarce resources.
Scope
It is necessary to review the current bibliography to highlight the most know characteristic mechanisms underlying Mediterranean plant-soil feed-backs and identify the processes that merit further research in order to reach an understanding of the plant-soil feed-backs and its capacity to cope with future global change scenarios. In this review, we characterize the functional and structural plant-soil relationships and feedbacks in Mediterranean regions. We thereafter discuss the effects of global change drivers on these complex interactions between plants and soil.
Conclusions
The large plant diversity that characterizes Mediterranean ecosystems is associated to the success of coexisting species in avoiding competition for soil resources by differential exploitation in space (soil layers) and time (year and daily). Among plant and soil traits, high foliar nutrient re-translocation and large contents of recalcitrant compounds reduce nutrient cycling. Meanwhile increased allocation of resources to roots and soil enzymes help to protect against soil erosion and to improve soil fertility and capacity to retain water. The long-term evolutionary adaptation to drought of Mediterranean plants allows them to cope with moderate increases of drought without significant losses of production and survival in some species. However, other species have proved to be more sensitive decreasing their growth and increasing their mortality under moderate rising of drought. All these increases contribute to species composition shifts. Moreover, in more xeric sites, the desertification resulting from synergic interactions among some related process such as drought increases, torrential rainfall increases and human driven disturbances is an increasing concern. A research priority now is to discern the effects of long-term increases in atmospheric CO2 concentrations, warming, and drought on soil fertility and water availability and on the structure of soil communities (e.g., shifts from bacteria to fungi) and on patching vegetation and root-water uplift (from soil to plant and from soil deep layers to soil superficial layers) roles in desertification.
Similar content being viewed by others

Introduction
Mediterranean climates occur in five occidental coastal regions of the continents between latitudes 34–45º in both hemispheres, i.e., the Mediterranean Basin, California, central Chile, the Cape region of South Africa, and southwestern and southern Australia (Di Castri and Mooney 1973; Di Castri 1981). Moderately wet and cold winters are coupled with dry and hot summers, but the intensity of the drier periods, increasing from high to low latitudes within these areas, can vary widely (Di Castri and Mooney 1973; Di Castri 1981). Several studies have improved our understanding of the functional and structural traits of Mediterranean plant communities, e.g., Sclerophylly, low growth rate and nutrient concentrations, and high presence of sprouting species (Kruger 1979; Specht 1979; Mooney 1989). The typical Mediterranean communities are mostly dominated by woody plants with conservative traits, such as slow growth and high sclerophylly associated with low water and nutrient availabilities.
Some important chemical and physical properties of the soils, such as nutrient availability, erodibility, moisture content, infiltration capacity, the quantity and nutritional quality of soil organic carbon (SOC), cation exchange capacity, and pH, are strongly determined by vegetative cover. Changes in these important soil variables can, in turn, affect plant cover. Close and very broad relationships, with multiple feedbacks, thus exist between plant communities and soil structure and function in this type of ecosystem.
Although some of these plant-soil interactions are well understood, other plant-soil interactions present some contrasting results such as the impact of increased atmospheric CO2 concentrations on plant litter and soil activity and others interactions remain poorly studied such as the role of roots in water transport among different soil layers. Consequently, a review of these main interactions is clearly needed. Current predictions suggest that drought and warming will increase in Mediterranean areas (IPCC 2007). Prolonged drought coupled to less predictable torrential rainfall (Frei et al. 1998) opens a scenario of shifts in the feedbacks and equilibrium within the plant-soil system. Several studies have explored the effects of increasing atmospheric CO2, drought, and warming on Mediterranean soils and plants, observing diverse responses, some of which involve feedback mechanisms between plants and soils. Some of these studies report contradictory results. Most results suggest that the impacts of global change threaten to increase the degradation and desertification process. In this context, the current knowledge suggests that plant-soil feedbacks can play an outstanding role in the capacity of these ecosystems to adapt to future scenarios. It is thus urgent to review and ordinate the current knowledge to highlight the best known characteristic mechanisms underlying Mediterranean plant-soil feedbacks and to identify the processes that merit further research to reach a better understanding of the plant-soil feedbacks and their capacity to cope with future global change scenarios.
By gathering data and reviewing the published studies (in the international literature) over the last two decades, this review aims (i) to characterize the functional and structural plant-soil relationships and feedbacks in Mediterranean regions, (ii) to review the impacts of increasing atmospheric CO2, drought, and warming on the Mediterranean plant-soil system, and (iii) to suggest directions for future research needed to improve our knowledge of the plant-soil system in changing Mediterranean environments.
Mediterranean plants and soils
Mediterranean plants: an evolution under low water and nutrient availabilities
Mediterranean plant communities are generally dominated by sclerophyllous woody plants with an herbaceous or shrubby understory (Specht 1981; Orshan 1983). When large data sets of Mediterranean plants have been studied across climatic gradients, mean annual precipitation has been the most determinant climatic variable associated with plant growth and community distribution (Sardans et al. 2011). From humid to semi-arid regions, understory vegetation grades from annual and perennial grasses and herbs (savanna understory) to evergreens and sclerophylls (heathland understory). The most specific structural characteristics of Mediterranean plants are those related to conservative mechanisms linked to the avoidance of water stress but frequently also to the scarcity of soil nutrients. The lack of soil resources has led to a narrow evolution of plant-soil systems.
Mediterranean plants have acquired a set of morphological and physiological adaptations in response to deficits of soil water (Table 1). The development of deep and extensive root systems is among the most characteristic traits of plants in Mediterranean ecosystems. Deep root systems enable the uptake of water from deep soil layers in drought seasons when the upper layers are water depleted (Veneklas and Poot 2003; Padilla and Pugnaire 2007; Baldocchi and Xu 2007; Hernández-Santana et al. 2008). Moreover, in Mediterranean plant communities, plant species with root-systems that tend to occupy different soil layer usually coexist avoiding the root systems overlapping and consequently diminishing the competition intensity, and also allowing to exploit the sources throughout all the soil depth (Castell et al. 1994; Silva and Rego 2003; Lefi et al. 2004a, b; Mattia et al. 2005; Moreno et al. 2005; Filella and Peñuelas 2003b; Silva and Rego 2003; Mattia et al. 2005; Mereu et al. 2009). Plants with deep-roots have proved to resist better drought events than species with shallow-root systems (Padilla et al. 2007; West et al. 2012). Furthermore, Mediterranean plants have high root plasticity in the early stages of life (Padilla et al. 2007). At the foliage morphological level, Mediterranean plants improve their capacity of drought avoidance increasing their foliar sclerophylly by developing thick cuticule and increasing leaf mass area (LMA) (Table 1), high density of foliar trichomes (Table 1), and high plasticity of foliar morphology and size (Table 1).
General adaptations to drought by Mediterranean plants at physiological level include a large capacity to maintain water flux and hydraulic lift (water conduction from soil to plant tissues) in the soil-plant continuum (Table 1), including a large resistance to xylem cavitation (Table 1) and high stomatal control (Table 1). Thus, Mediterranean plants also respond to drought by both decreasing xylem cavitation vulnerability and adapting and protecting leaf function and structure (De Mico and Aronne 2009; Vilagrosa et al. 2010). Some studies suggest that these two responses are coordinated (Vilagrosa et al. 2010). Drought changes woody architecture of woody plants increasing vessel density, decreasing total hydraulic diameter (Corcuera et al. 2004; Gea-Izquierdo et al. 2012), increasing vessel wall thickness (De Mico and Aronne 2009) and decreasing cambial activity (De Luis et al. 2011). Other general adaptation strategies to drought include the accumulation of metabolites and/or water soluble sugars in tissues to increase turgor (Table 1), a large capacity to reduce metabolic activity during drought periods (Table 1), and efficient photoprotective and antioxidant mechanisms (Table 1). Moreover, antioxidant mechanisms to cope with summer drought have already been listed (Table 1). The desiccation tolerance strategy observed in some Mediterranean species allows maintaining high stomatal conductance despite very low water potentials (Fotelli et al. 2000; Volaire et al. 2009) In these drought tolerant species the predawn leaf relative water content decreases simultaneously with stomatal closure as drought progresses (Gulías et al. 2002). However, the most stress-tolerant plant species are able to develop very low water potentials with markedly reduced stomatal aperture (Fotelli et al. 2000).
Apart from water availability, the capacity of the soil nutrient supply has often been shown to be an important factor in the growth, structure, and distribution of plant communities (Kruger 1979; Specht 1979; Carreira et al. 1992; Sardans et al. 2004; Henkin et al. 1998). Mediterranean plants share characteristics such as slow growth, sclerophylly, and low nutrient contents that are present in plants of other non-Mediterranean ecosystems adapted to poor soils (Bussotti et al. 2000). The importance of nutrients has been appreciated in many experiments of nutrient manipulation by fertilization in different Mediterranean zones of the world, for example in Californian chaparral (McMaster et al. 1982), South African fynbos (Witkowski et al. 1990; Herppich et al. 2002), Australian jarrah and Eucalyptus forests (Dell et al. 1987), and the Mediterranean Basin (Mayor and Rodà 1994; Henkin et al. 1998; Sardans and Peñuelas 2004; Sardans et al. 2004, 2005a, b). Mediterranean plants frequently change foliar chemical and physical traits in response to both water deficit and nutrient stress (Gratani et al. 2003; Sardans et al. 2006a, 2008a, b). Proteaceae plant communities that dominate several Mediterranean climatic regions of South Africa and southwestern Australia have the capacity to accumulate more or less P in seedlings when P is most limiting (Groom and Lamont 2010). Due to the prevalence of soils with low nutrient content in Mediterranean areas, Mediterranean plants have high levels of investment in root systems to enhance nutrient uptake, in addition to the C invested for adaptation to low water availability (Martínez et al. 1998; Sardans et al. 2005a, 2006b). Resprouting capacity is also determined by the availability of soil nutrients (Sardans et al. 2006c). A higher retranslocation of nutrients from leaves before falling is another characteristic of Mediterranean plants helping them to achieve a higher nutrient use efficiency (NUE) (Schlesinger and Hasey 1981; Fioretto et al. 2003; Sardans et al. 2005a) and having a further effect on soil organic matter quality as commented below. García-Palacios et al. (2012) in an experiment of Mediterranean grasslands growing at different levels of soil N availability heterogeneity observed that soil N availability heterogeity modulated the effects of root length, LMA and nutrient availability on ecosystem function. The nutrient content of soil is thus a key factor for the structure and function of Mediterranean plants that, in turn, influence nutrient content by their adaptations that improve NUE.
A positive interaction has been observed between soil-water and nutrient availabilities (Matías et al. 2011). Increases in the availability of water increases plant-production capacity, produces more litter of higher nutritional quality (Dirks et al. 2010), and protects the soil better. All these enhancements further enable higher microbial activity and availability of nutrients (Dirks et al. 2010). In fact, the competition for soil resources is a very important and widespread phenomenon in Mediterranean plant communities (Vilà and Sardans 1999), suggesting a significant interdependence of soil-plant systems, mainly to improve water and nutrient availability and the capacity to withstand limitations.
Species-specific adaptations of plant species and communities to resist drought
The capacity of Mediterranean plants to withstand drought varies for the different species within and between plant communities. For example, Californian perennial grasses are more tolerant of drought than annuals of the Mediterranean Basin but less tolerant than perennials of the Mediterranean Basin, despite the more intense summer drought of California’s Central Valley compared to the Mediterranean Basin (Vaughn et al. 2011). Other studies have observed better adaptation to drought in evergreen oaks of the Mediterranean Basin compared to those of California (Grünzweig et al. 2008). Different morphological and physiological adaptive strategies to drought have been observed in comparisons between typical groups of Mediterranean plants (Medrano et al. 2009). Different growth forms display specific morphological adjustments in response to shortages of water. Different coexisting species of the same community can also respond differently. When drought is enhanced, some species survive and grow less well than others in the short and medium terms (Ogaya et al. 2003; Ogaya and Peñuelas 2007a) due to species differences in some of the previously mentioned traits for resisting drought and increasing water use efficiency (WUE). Species also differ in their phenotypic capacity to change some traits (Lloret et al. 2004; Ogaya and Peñuelas 2008; Bell et al. 2007). Significant differences in the capacity to take up water from different soil depths (Filella and Peñuelas 2003b; Lefi et al. 2004a, b), a trait that is especially important in severe droughts, are frequently observed. Moreover, coexisting species in the same community have different strategies for the uptake of water, from plants with leaf water potentials depending on soil water status and evaporative demands (isohydric) to plants whose leaf water potentials do not depend on soil water status and evaporative demand (anisohydric). This combination of hydraulic strategies and also different strategies to capture of water at different times allows plants to avoid competition for water (Peñuelas et al. 2011; Quero et al. 2011).
Mediterranean species also differ in their capacity to regenerate. In Mediterranean ecosystems, some species are able to regenerate by sprouting, and others can only regenerate from seeds. These two strategies are widely represented in Mediterranean communities and frequently coexist in competitive equilibrium. The high cost of biomass production together with high levels of natural disturbance (severe dry periods, recurrent fires, and herbivore pressure) are proposed as general explanations for the elevated number of species with resprouting capacity (Mooney and Dunn 1970; Lloret et al. 1999) that adds complexity and diversity to Mediterranean plant communities.
Mediterranean soils
Mediterranean areas have a large variety of soils, but they generally have medium-low fertility (Yaalon 1997). The Mediterranean region of central Chile is relatively nutrient rich, the vegetation is mainly limited by water and leaf turn-over rates is relatively high (Miller 1981). California and Europe Mediterranean areas are less nutrient rich and climate is the most limiting factor but nutrients have proved to exert some importance, and in Mediterranean areas of Australia and South-Africa soils are old and nutrient-poor, nutrients thus having an outstanding role in ecosystems structure and function (Miller 1981). The most frequent soil types range from Alfisols (Luvisols) in the wetter regions with mesic environments to Inceptisols and Entisols in more xeric and semi-arid environments or on slopes. A majority of calcareous rock allows the formation of Xerolls (Mollisols under xeric environments) due to the protection of humic organic matter by the aggregates formed with Ca2+ (Yaalon 1997). In extreme xeric conditions in the transition to sub-desertic environments, the arid conditions and the salinization of the upper horizon can form Aridisols. Rubefication (by the precipitation of Fe2O3 during summer drought from the Fe2+ released by the weathering of minerals during wetter seasons) and incomplete lixiviation of carbonates and nutrients (due to the lack of sufficient rainfall) from exchangeable complexes (clays and humus) are general features of Mediterranean soils (Yaalon 1997).
Apart from the limiting water content during summer drought, Mediterranean soils are often poor in nutrients (Specht 1973; Di Castri 1981; Carreira et al. 1991). Along with nitrogen (Mayor and Rodà 1994), phosphorus is a frequent limiting nutrient in Mediterranean ecosystems (McMaster et al. 1982; Witkowski et al. 1990; Lebourgeois et al. 1997; Henkin et al. 1998; Sardans et al. 2004, 2005a, 2006d). Soil moisture and nutrient supply and their availability for plant growth thus strongly interact in Mediterranean ecosystems (Everard et al. 2010).
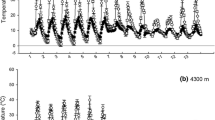
The large variation in soil moisture throughout the year significantly impacts soil chemistry and activity, causing large seasonal changes in the activities of soil enzymes (Sardans and Peñuelas 2005 and 2010; Sardans et al. 2006d, 2008c and d). A comparison of studies from the last two decades that have used similar methods to estimate activities of extracellular soil enzymes indicates that the activities of several important extracellular enzymes, such as urease, protease, β-glucosidase, and acid and alkaline phosphatases, in Mediterranean soils are similar to those of other temperate ecosystems, such as temperate grasslands, temperate wet forests, and tropical forests (Fig. 1, Table S1 in the supplementary material). Soil enzyme activity is considered one of the best proxies of health and activity of soils (Dick 1997), so the similar activities suggest that even though Mediterranean ecosystems are generally less productive than other temperate and tropical ecosystems, they invest similar amounts of resources to produce soil enzymes. For example, the activity of root phosphatases in Mediterranean shrubs is equal to or even higher than in shrubs of other temperate areas of Europe (Estiarte et al. 2008). These similarities are probably related to the limiting role of nutrients and to the advantage provided by nutrient availability to improve WUE. For example, higher N availability improves the mechanisms of stomatal control reducing the unproductive water losses allowing a higher biomass production by unit of water absorbed (Brueck 2008).

Soil extracellular activities of urease, protease, β-glucosidase, and acid and alkaline phosphatases expressed in mg substrate released g−1 soil dry weight h−1 in Mediterranean soils compared with temperate and tropical soils. The reports have analyzed urease activity by the Kandeler and Gerber (1988) method or by comparable methods such as that of Tabatabai and Bremner (1969), protease activity by the Ladd and Butler (1972) method, β-glucosidase activity by the Eivazi and Tabatabai (1988) or related methods (Dick et al. 1996), and acid and alkaline phosphatase activities by the Tabatabai and Bremner (1969) and comparable methods (Tabatabai 1994). The values presented are those of controls when diverse treatments were applied to the studied soils and of the annual mean when several measurements were conducted throughout the year. The number of studies reviewed, by biome and enzyme, are depicted above the bars. The bars indicate mean (+ S.E.). For detailed information, see Table S1 in the supplementary material
Soil enzyme activity in Mediterranean soils is mainly determined by moisture. Many studies have reported decreased activity with decreasing soil moisture, both throughout the year (Grierson and Adams 2000; Quilchano and Marañon 2002; Criquet et al. 2002; Bell and Adams 2004; Sardans et al. 2008c, d; Rutigliano et al. 2009; Sardans and Peñuelas 2010) and along gradients of soil moisture or experimental drought (Garcia et al. 1994; Sardans and Peñuelas 2005 and 2010; Sardans et al. 2006d, 2008c and d). Long summer droughts decrease soil enzyme activity and plant uptake and change soil C:N:P stoichiometry relative to other seasons (Aponte et al. 2010). Moreover, soil enzyme activity depends more on soil moisture than on SOC nutritional quality (Sardans and Peñuelas 2010), although both are important factors controlling rates of mineralization (García-Pausas et al. 2004; Rutigliano et al. 2009; Incerti et al. 2011). For example, the C:N and C:P ratios of litter are inversely correlated with rates of decomposition (Bengtsson et al. 2012). Similar results are observed when soil activity has been measured by soil respiration. Positive relationships are found between soil respiration and soil moisture (Rey et al. 2002 and 2011; Inglima et al. 2009; Carbone et al. 2011). In Mediterranean ecosystems when volumetric soil water content dropped below certain values, ranging from 13 % (Rey et al. 2011) to 20 % (Xu and Qi 2001; Rey et al. 2002) depending on reports, there is a strong soil respiration limitation. However, litter decomposition rates of Mediterranean forests are similar to those of temperate deciduous forest (Van Wesemael and Veer 1992). The values are very variable, ranging from 0.12 to 2.22 year−1 depending on the species (Van Wesemael 1993; Fioretto et al. 2003; Rodríguez-Pleguezuelo et al. 2009). Frequently N concentration increases during decomposition process (Gallardo and Merino 1992; Fioretto et al. 2003), specially in the first months of decomposition (Gallardo and Merino 1999). Thereafter, during more advanced stages of decomposition, N release can decrease increasing litter N concentration (Maisto et al. 2011), observing in some cases a shift from N limitation to P limitation throughout litter decomposition process (Gallardo and Merino 1999). Not all studies have detected this trend in N concentrations during decomposition process (Fioretto et al. 2001). N litter concentration is also correlated with litter decomposition rates (Moro and Domingo 2000; García-Pausas et al. 2004), but lignin content and leaf mass area (LMA) of litter are also determinant in Mediterranean plants litter decomposition rates (Cortez et al. 1996, 2007; Kazakou et al. 2006, 2009). Furthermore, plant species of early succession stages with low LMA, and high growth rate and litter N content have faster litter decomposition rates and N release from litter than species of later successional stages with opposite traits (García-Pausas et al. 2004; Kazakou et al. 2006 and 2009). Litter resulting from mixing litter of different species have proved to decompose at different rates (in general faster) than that expected from the decomposition rates observed by each species litter alone (De Marco et al. 2011). The importance and changes of P concentration during litter decomposition process has been less studied than those of N. Some studies have observed a decrease of P concentrations during litter decomposition (Gallardo and Merino 1999; Fioretto et al. 2001) with the consequent increases in litter N:P ratio (Gallardo and Merino 1999). In addition to litter quality, soil temperature and moisture have also an outstanding role in litter decomposition rates in Mediterranean ecosystems (Fioretto et al. 2000; Dirks et al. 2010; Incerti et al. 2011). Dirk et al. (2010) observed that litter decomposition is related with atmospheric humidity during summer, showing that during dry periods a significant part of water to leaf litter decomposition can come from atmosphere. The community density of soil macroinvertebrate decomposers is also related to precipitation through its effects on the physical properties of soil and the quality of plant litter (Morón-Ríos et al. 2010). These studies demonstrate that in Mediterranean ecosystems water availability is the most important factor determining soil biological functioning.
Topographic factors, due to solar incidence and slope variation, and soil texture, due to water-infiltration capacity, have key roles in plant cover by determining the retention and storage of soil water, soil enzyme activity, and rates of soil erosion (Bastida et al. 2008). Soils with sandy structures are particularly critical in Mediterranean regions because of their high permeability and low capacity to retain water (Kooijman et al. 2005; Prieto et al. 2010a, b). Soil organic matter (SOM) plays a key role in soil-water content by allowing the formation of stable aggregates that decrease the index of soil erosion (Barthes and Roose 2002). Calcareous Mediterranean soils have some singular differences from other soils regarding soil fertility. These soil types severely immobilize phosphate due to precipitation with Ca2+ (Carreira and Latjha 1997), which is frequently critical because of the natural low P content of Mediterranean soils. On the other hand, calcareous soils have a good capacity to physically stabilize the SOM (Rovira and Vallejo 2003; Rovira et al. 2010), which should be beneficial due to the importance of stable soil aggregates in preventing soil erosion (Barthes and Roose 2002). Mediterranean soils have a low-moderate humification index (Simón et al. 1994) as a result of lower plant production and soil microbial activity imposed by summer drought and cold winters in continental and alpine areas (Simón et al. 1994). Soils of Mediterranean woodlands normally contain considerable amounts of SOC, with moder the main frequent type of humus (Vacca 2000; van Wesemael and Verstraten 1993; Sevink et al. 1989), although mull humus is also widely represented (Peltier et al. 2001; Sadaka and Ponge 2003; Andreetta et al. 2011 and 2012). The constraints and feedback processes resulting from the long and highly variable summer droughts have large effects on humus properties (Sevink et al. 1989; van Wesemael and Verstraten 1993; Vacca 2000; Peltier et al. 2001; Ponge 2003; Andreetta et al. 2011) and microbial communities (Monokrousos et al. 2004; Goberna et al. 2006; Aponte et al. 2010) in Mediterranean areas. Amounts of SOM in Mediterranean ecosystems depend on levels of precipitation (Boix-Fayos et al. 1998; Gómez-Rey et al. 2010) and type of plant community (Lucas-Borja et al. 2010). Rewetting events during summer droughts are particularly important by suddenly increasing the mineralization of the SOC, ecosystem carbon fluxes (Jarvis et al. 2007) and the release of nutrients (Rey et al. 2005; Almagro et al. 2009).
Plant-soil relationships in Mediterranean ecosystems
Several studies have observed a strong interdependence between changes in plant community and changes in soil properties in Mediterranean ecosystems (Garcia et al. 2002; Goberna et al. 2007a, b; Enright and Goldblum 1999; Dias et al. 2011). These close relationships are observed through both time and space. We here discuss the most relevant plant-soil relationships summarized from the literature. Most feedbacks between Mediterranean plant communities and soils are linked to their mutual protection.
Plant cover: avoiding competition for soil resources and improving the physical protection of soil
Despite the low water content and nutrient availability of Mediterranean soils, Mediterranean plant communities frequently have a notable cover and species diversity, reflecting the large capacity of coexisting species to avoid competition for soil resources. The avoidance of overlapping structures of the root system has been widely observed among different types of plants, such as between trees and grasses (Silva and Rego 2003; Lefi et al. 2004a, b; Mattia et al. 2005; Moreno et al. 2005) or among more similar types, such as between woody evergreens (Filella and Peñuelas 2003b; Silva and Rego 2003; Mattia et al. 2005; Mereu et al. 2009). This differential capacity of plant species to exploit soil sources at different depths of soil has beneficial effects for the coexistence of species. Filella and Peñuelas (2003b) observed higher water uptake in evergreen shrubs growing under large pine trees than in those growing under smaller pines or in those separated from large pine trees, an effect very likely linked to the hydraulic lift, driven by a deep root system of big trees, from deep soil layers to shallower layers, effect that improves the water status of shrubs that grew under big pine trees (Filella and Peñuelas 2003a). The possibility that the deep roots can pump water to the surface layers, thereby helping to maintain higher moisture levels in the upper richer soil layers, is a phenomenon that needs to be investigated in depth. The extensive and deep root systems of Mediterranean plant communities also protect the soil against erosion from torrential rain (Andreu et al. 1998; Chirino et al. 2006), mainly on slopes (Mattia et al. 2005; De Baets et al. 2007 and 2008). This widespread presence of roots throughout the soil layers has several other notable effects on soil properties and traits, such as decomposer fauna and physical soil texture (Maestre and Cortina 2002, 2003). The presence of Mediterranean plant communities with more diverse species compositions improves soil quality and structure (higher levels of SOM, microbial biomass, and nutrient content; more stable aggregates; a more equilibrated texture; and greater permeability) (Andreu et al. 1998; Pinzari et al. 1999; Chirino et al. 2006).
Plant cover and soil fertility and water availability
Most studies suggest a significant positive and rapid feedback between soil properties linked to fertility (water-storing capacity, permeability, nutrient availability, depth) and plant cover (Gallardo et al. 2000; Johnson-Maynard et al. 2002; Castaldi et al. 2009; Ochoa-Hueso et al. 2011). An increase in plant cover thus has a direct positive effect on soil porosity, which increases water-infiltration capacity and decreases runoff (Johnson-Maynard et al. 2002; Goberna et al. 2007a; Garcia-Estringana et al. 2010). This capacity to improve water infiltration generally occurs in different types of plant community but is higher in grasses and angiosperm evergreen shrub and tree communities than in pine forests (Johnson-Maynard et al. 2002; Garcia-Estringana 2010). These differences are probably linked to the observed differences in the formation of sand-sized organic matter in different communities, such as between evergreen shrubs and pines (Quideau et al. 1998). A positive feedback between plant cover and soil properties is consequently established. The improvement of soil quality generated by plant cover improves soil moisture and water availability, fertility (Gallardo et al. 2000), and soil enzyme activity (Garcia et al. 2002). These effects are related to the improvements in the physical and chemical properties of soil and thus to higher water availability that, in turn, has a positive effect on Mediterranean plants (Sadaka and Ponge 2003; Ruiz-Sinoga et al. 2011). This relationship between plant cover and soil protection and fertility tends to be stronger in dry areas than in wetter areas (Ruiz-Sinoga et al. 2011).
Mediterranean plants, by having some of their typical properties linked to mechanisms of stress avoidance, can affect soil properties linked to nutrient cycling and availability. A reduction in nutrient losses in litterfall could be a strategy employed by sclerophyllous plants adapted to poor soils (Aerts 1995) and could partially compensate for decreased nutrient uptake in dry conditions. This conservative use of water and nutrients, though, can also retard nutrient cycling as a result of the larger content of structural compounds and the more recalcitrant litter that also has lower nutrient concentration due to a high level of retranslocation. Mediterranean plants can cover up to 84 % of the N demand throughout leaf expansion from retranslocation from old tissues (Silla and Escudero 2003). High levels of N, P and K retranslocation from senescing leaves have been widely observed in Mediterranean plants (Grubb 1977; Pugnaire and Chapin 1993; Fioretto et al. 2003; Milla et al. 2005; Sardans et al. 2005a; Fife et al. 2008). Sclerophyllous leaves are moreover rich in structural compounds (Rundel 1982; Gallardo and Merino 1992; Turner 1994) that also produce litter of low nutritional quality and lower rates of litter decomposition, which can then lead to more occluded humic compounds that retain nutrients in unavailable forms to plants in the soil. However, photodegradation could contribute to decrease lignin concentration in litter of Mediterranean soils such as observed in other semiarid environments (Austin and Vivanco 2006; Austin and Ballaré 2010; Ballaré et al. 2011). A fast release of N under high UV radiation intensity allowing increases in soil microbial activity is one of the probable mechanisms underlying this effect (Foereid et al. 2010). Mediterranean plants are rich in secondary metabolites such as phenolics (Castells and Peñuelas 2003; Hernandez et al. 2004; Skerget et al. 2005; Coulis et al. 2009; Bettaieb et al. 2011), compounds that decrease N mineralization rates in litter (Castells and Peñuelas 2003; Castells et al. 2004) and retard decomposition (Castaldi et al. 2009). Foliar extracts of Arbutus unedo rich in phenolic compounds strongly inhibit soil nitrification (Castaldi et al. 2009). Mediterranean plants have higher levels of N and P retranslocation (Nuñez-Oliveira et al. 1993; Fioretto et al. 2003; Sardans et al. 2005a) that lead to leaf litter with low nutrient concentrations. This strategy of retranslocation has further consequences on the rates of litter decomposition because the concentrations of litter N and P appear to be determinants in the rates of litter decomposition (Schlesinger and Hasey 1981) and in the community structure of soil decomposers (Almagro and Martínez-Mena 2012). More retranslocation should thus lead to a low release of N and P from litter and lower availability of soil N and P. The high capacity for erosion of Mediterranean rain may have further favored the evolution of plants toward higher retranslocation to assure better control of nutrient stocks, thereby improving the capacity of the plant-soil system to retain nutrients. The high retranslocation is related to the observed higher percentage of nutrients in the stand biomass of Mediterranean forests compared with other forest types (Rodà et al. 1999a, b; Sardans and Peñuelas 2012; Sardans et al. 2012a).
The capacity of plants to absorb nutrients in the early phases of regeneration may be a key factor to prevent the degradation of soil. Resprouting capacity should be particularly important in nutrient-poor environments and in environments, such as Mediterranean ecosystems, where the risk of nutrient losses is high after fires or from torrential rainfall. The sprouting capacity during the initial phases of regeneration after disturbances such as fires thus provides resprouting species with a higher competitive ability for nutrient uptake than the species that can only regenerate with seeds (Lloret et al. 2004; Sardans et al. 2004). However, in Mediterranean ecosystems sprouter-seeders strategies present clear trade off, with sprouters having higher allocation to root system and less to flowering and seed production, and seeders having higher aboveground growth capacity, higher flowering and seed production (Enright and Goldblum 1999; Bell 2001). Sprouters tend to have a higher conservative use of nutrients than seeders (Saura-Mas and Lloret 2009), but it is not clear that in general sprouters increase their cover with fires. Contrarily, regional surveys have reported an increase in the relative abundance of non-resprouters in burnt areas (Lloret et al. 2005). Nevertheless, other reports suggest that the presence of resprouters can be enhanced when the frequency of disturbances increases (Bon and Midgley 2003). However, some studies have suggested new possibilities that explain the high prevalence of sprouter plant species in Mediterranean ecosystems. Lamont and Wiens (2003) suggest that the great genet resilience associated with frequent new branching by fire, drought-rewetting and herbivore might benefit the somatic mutations in the meristematic tissues favoring ecotypic differentiation and speciation in sprouters. Moreover, within a single species resprouts have proved to better resist drought than genets grown from seeds (Peña-Rojas et al. 2004).
Plant communities with high numbers of species produce litter with higher rates of decomposition and a better capacity to retain soil N than communities with lower species diversity (Bonanomi et al. 2010; Maisto et al. 2011). This finding suggests that a mixed litter from different species constitutes a more balanced and complete source of food for the community of soil decomposers than litter from only one species. In Mediterranean ecosystems that produce litter of low nutritional quality, the mixing of species in species-rich communities is thus favored because the diversity can improve soil activity and nutrient mineralization, conferring greater stability to the soil. Baraza et al. (2009) have observed that foliar nutritional quality is highly variable among different Mediterranean plants of the same and of different communities, frequently with different nutritional properties, creating a scenario of differential nutritional supplies in space (different species) and time (seasons) in which herbivores must select their diet.
In Mediterranean arid areas with high risk of desertification, usually water infiltration capacity is higher and run-off erosion is lower in slopes with less insolation and evaporation capacity (Boix-Fayos et al. 1998; Kutiel et al. 1998). In slopes of more arid Mediterranean areas the variability of water and nutrients availability is related to the distribution of vegetation patches with respect to bare soil patches that can be considered as runoff sinks and sources, respectively (Boix-Fayos et al. 1998; Kutiel et al. 1998; Ruiz-Sinoga and Martínez-Murillo 2009; Mayor et al. 2009; Mayor and Bautista 2012; Gabarrón-Galeote et al. 2012; Merino-Martin et al. 2012). But some few studies have not observed these relationships so clearly (González-Pelayo et al. 2010). This effect of patches is crucial in the maintenance of vegetation in more arid sites. The patches of vegetation also influence hydrological and erosion behavior (Ruiz-Sinoga et al. 2010, 2011), and are related with soil properties (Ruiz-Sinoga et al. 2010; Ochoa-Hueso et al. 2011) such as infiltration capacity and biological activity throughout the slopes (Maestre and Cortina 2002; Agra and Ne’eman 2012). When adjacent patches have different species composition, with different root depth and density, ones can act as runoff sinks and others as runoff sources (Merino-Martin et al. 2012). Grazing, particularly from mammals, contribute to maintain plant diversity between different patches of vegetation (Golodets et al. 2011). The presence of patches with very different species composition can exert a key role in plant invasive success to maintain large soil variability (Harrison et al. 2001; Prober and Wiehl 2011). Moreover, the presence of large trees in patches exerts an important role in maintaining high level of herbaceous species richness by affecting light and water competition intensity and availability (Blank and Carmel 2012).
The Mediterranean agroforest systems (Dehesa) that cover wide areas in south Europe and constitute a particular system where human activities help to maintain the ecosystem with a particular plant cover with savanna structure (Gallardo 2003; Gallego Fernández et al. 2004) that maintain great differences in the horizontal spatial distribution of soil nutrient availability (Gallardo 2003; Gea-Izquierdo et al. 2010), soil organic content (Howlett et al. 2011) and soil water availability (Joffre et al. 1999). All these soil properties are positively related with tree canopy. Regional scale evidences of the tree density and mean annual precipitation suggest that the structure of these man-made ecosystems have adjusted over long-time and corresponds to an optimal functional equilibrium with water cycle (Joffre et al. 1999). Frequently the major problem to maintain these ecosystems and maintain tree production is the shrub competition that strongly impacts soil nutrient availability (Rolo et al. 2012).
Changes during successional events: soil properties and plant diversity
The positive feedbacks between plant cover and soil fertility properties also drive plant succession. An increase in the growth of tree seedlings is observed when shrub cover increases, with shrubs acting as nursery species for the seedlings by enhancing soil properties such as infiltration, retention capacity, and soil enzyme activity (Duponnois et al. 2011). In the driest Mediterranean areas with sparse vegetation, soil patches under vegetation present higher levels of water permeability, SOM, microbial biomass, and enzyme activity than bare soil (Goberna et al. 2007b). Moreover, when plant cover is enhanced, the SOM, soil microbial biomass, and activity linked to higher biomass production and soil moisture quickly increase (García et al. 2005; Muñoz et al. 2007; Duponnois et al. 2011; Maestre et al. 2011). The composition and activity of soil microbial communities in Mediterranean areas are strongly determined by changes in plant communities that cover the soil (Bonkowski and Roy 2012), and soil microbial activity is generally negatively affected by the degradation of the plant cover (Garcia et al. 2002). Other studies have also observed a close relationship between the spatial distribution of plants and soil NH4 + concentration (Gallardo et al. 2000; Castillo-Monroy et al. 2010). More diverse plant communities are associated with soils higher in SOM and N concentration than are plant communities with fewer species (Lucas-Borja et al. 2012). In the meadow-shrubland-maquis succession in Mediterranean climates, the aboveground biomass and overall soil activity increase, although with high seasonal fluctuations (Fioretto et al. 2009). In fact, when soil properties have been studied in different plant communities from grassland to evergreen shrubland and forest, larger differences in soil variables related to soil fertility have been observed among seasons than among different plant communities (Monokrousos et al. 2004), illustrating the strong link between seasonal changes of aboveground vegetation with seasonal changes in soil properties. Anyway, regarding nutrient availability in soils the current studies suggest a strong mechanisms of nutrient conservation during succession in Mediterranean ecosystems. Intense nutrient uptake and accumulation capacities are observed in the early successional especies (Carreira et al. 1992). The presence of legumes is high in different Mediterranean successional communities both in early succesional stages with high presence of herbaceous legumes and in latter successional stages with high presence of perennial legume shrubs (Arianoutsou and Thanos 1996). Regarding drought response strategy, early successional species escape drought by dying before summer while later herbaceous species maintain a favourable water status in relation to leaf shedding during summer; finally, late successional trees with a large body allow access to a large pool of resources, producing dense leaves that can tolerate desiccation (Navas et al. 2010). In this respect, seed banks of herbaceous plants are less sensible to decrease than those of woody plants if the frequency of severe drought episodes increases (Del Cacho and Lloret 2012). The protection of soil moisture and the improvement of soil texture under shrubs can facilitate tree seedling survival in most wet Mediterranean ecosystems (Gómez-Aparicio et al. 2005).
Studies during the course of succession of plant communities in Mediterranean areas show a progressive replacement of species with low leaf mass area (LMA) and high concentrations of foliar nutrients by species with opposite traits (Cortez et al. 2007). Fast-growing species are replaced by slower growing species associated with the conservative use of soil resources. This trend in Mediterranean areas is closely associated with changes in the availability of soil N and P and also with the ratio between bioavailable N and P (Dias et al. 2011). The large capacity of the fast-growing, nutrient-rich species to absorb nutrients in the first phases of succession of the plant community should be particularly important in Mediterranean areas by hindering the loss of soil nutrients by erosion and leaching.
Mycorrhizae and their role in Mediterranean plant-soil relationships
Mycorrhizae improve the resistance of plant communities against environmental stresses including nutrient deficiency, drought, and soil disturbance. Therefore, they play a key role in Mediterranean plant communities that frequently experience all these stresses that threaten to increase in the scenario of global change. Several studies in Mediterranean plant communities have observed that mycorrhizal fungi play an important role in the capacity of plants to take up water (Bell and Adams 2004) and nutrients (Martínez-García et al. 2011). Mycorrhizal activity in Mediterranean ecosystems is often specifically associated with an improvement in the uptake of P (Bell and Adams 2004; Domíngues-Nuñez et al. 2006; Martínez-García et al. 2011). Frequently a positive correlation between total percentatge of mycorrhizal root-length colonization and soil organic matter contents and soil N concentrations has been observed in Mediterranean ecosystems (Çakan and Karatay 2006). Studies of mycorrhizal inoculation in Mediterranean plants have shown a largely improved N, P and K uptake by inoculated plants (Azcón and Barea 1997; Medina and Azcón 2010) improving plant survival, diversity and functionally of these drought-adapted plant communities (Barea et al. 2011). These studies have demonstrated that the role of mycorrhization in more dry and nutrient poor-Mediterranean ecosystems is critical to improve the drought resistance capacity and to prevent the aggravation of desertification process.
Mycorrhizal fungal communities are sensitive to processes of soil degradation and to summer drought (Martínez-García et al. 2012). Both reduce mycorrhizal density, but the communities do not disappear, suggesting a certain degree of adaptation to stress (Barea et al. 2011).
Impacts of increased atmospheric CO2 concentrations
Mediterranean shrublands have the lowest capacity to store C compared to other temperate-climate shrublands throughout Europe (Beier et al. 2009). However, several studies have hypothesized an increase in plant and microbial biomass under high atmospheric CO2 in Mediterranean ecosystems (Dhillion et al. 1996). An overview of the current literature identifies three basic and direct effects of increased CO2 concentrations on Mediterranean ecosystems: changes in plant water conductance, a trend to increase the accumulation of biomass, and a change in the chemistry of plant tissues (Table 2, Table S2 in the supplementary material).
Under elevated levels of atmospheric CO2, the foliar conductance of water decreases and WUE increases (Bettarini et al. 1995; Tognetti et al. 1999a, b, 2000; Roumet et al. 2000; Maroco et al. 2002; Morgan et al. 2004) (Table 2). Some studies (Peñuelas and Azcón-Bieto 1992; Scarascia-Mugnozza et al. 1996; Tognetti et al. 1998; Nijs et al. 2000; Morgan et al. 2004; Peñuelas et al. 2008a; Andreu-Hayles et al. 2011; Maseyk et al. 2011), and mathematical models (Osborne et al. 2000) suggest that increases in concentrations of atmospheric CO2, by increasing WUE, can counteract the effects of the projected increase in drought in the coming decades (IPCC 2007). Long-term experiments have not tested whether Mediterranean plants could acclimate to elevated CO2 to buffer these higher WUEs. Moscatelli et al. (2001) observed that the effects of elevated CO2 on plant growth and microbial biomass occurred only at the beginning of their study and progressively diminished as monitoring continued. In this context, Rey and Jarvis (1997) showed that the effects on relative growth rates (RGR) had disappeared after 4.5 years of plants exposed to increased atmospheric CO2 concentrations and that the increased biomass accumulation was the result of an early effect of RGR. A recent study has observed that elevated atmospheric CO2 (712 ppm) during a strong summer drought did not prevent a decline in water use and the capacity of soil to store water in the most critical dry periods (Hernández-Santana et al. 2009). Moreover, the results of other studies manipulating atmospheric CO2 and conditions of drought have suggested that these two variables may have compensatory effects; the effects of short-term increases in CO2 on several plant variables that improve WUE can be counteracted by enhanced conditions of drought at the levels projected by most climatic models (Lopez et al. 1998; Nijs et al. 2000; Loreto et al. 2001; Hernández-Santana et al. 2009).
An overview of the current bibliography (Table 2) on the effects of increased atmospheric CO2 on plant growth and photosynthetic capacity indicates a short-term enhancement of plant growth and photosynthetic rates (Scarascia-Mugnozza et al. 1996; Lopez et al. 1998; Nijs et al. 2000; Marek et al. 2001), but this positive effect is frequently only observed in nutrient-rich soils (Midgley et al. 1995; Cruz et al. 2003). The positive effect on growth generally decreased with time (Körner and Miglietta 1994; Jones et al. 1995; Peñuelas et al. 2008a; Andreu-Hayles et al. 2011), suggesting acclimation to increase of atmospheric CO2 concentrations (Navas et al. 1995; Roumet et al. 2000; Saurer et al. 2003). The increased growth in the short-term is linked to increases in WUE associated with an atmosphere richer in CO2 (Nijs et al. 2000). Moreover, increased atmospheric CO2 concentrations can favor species that fix N2 over those that do not (Navas et al. 1997; Grünzweig and Dumbur 2012) and can increase the nutrient uptake capacity by enhancing mycorrhizal associations (Rillig et al. 1998a). The higher production of reproductive and vegetative biomass of legumes than non N2-fixing plants under increased atmospheric CO2 concentrations related to higher protein synthesis capacity could give competitive advantage to legumes in a CO2–rich atmosphere (Navas et al. 1997; Grünzweig and Dumbur 2012).
Most data from Mediterranean regions show that when plants are grown in increased atmospheric CO2 concentrations a decrease in plant N and P concentrations is observed and consequently the C:N and C:P ratios of plant biomasses increase (Sardans et al. 2012b and Table 2). Under elevated CO2, plant production and the C:N ratio increase, diminishing the concentrations of N and other nutrients (De Angelis et al. 2000; Peñuelas et al. 2001; Bassirirad et al. 2003; Tognetti and Peñuelas 2003) and increasing phenolic compounds (Peñuelas and Estiarte 1998; Castells et al. 2002) in foliar biomass. Mediterranean plants can partially buffer the effects of higher C:N ratios in plant tissues under increased atmospheric CO2 concentrations by increasing the retranslocation, fixing, and mineralization of N (Peñuelas and Estiarte 1997). Despite the capacity of plants and ecosystems to increase nutrient availability under increased atmospheric CO2 concentrations, most studies have observed higher C:N ratios and lower nutrient contents in plant tissues under increased atmospheric CO2 concentrations than in current levels of atmospheric CO2 concentration (Bettarini et al. 1995; Midgley et al. 1995; Gahrooee et al. 1998; Cotrufo et al. 1999; Roumet et al. 1999; De Angelis et al. 2000; Blaschke et al. 2001; Peñuelas et al. 2001; Tognetti and Peñuelas 2003; Cruz et al. 2003) (Table 2). This decrease in N concentrations could impact on plant-insect relationships (Roumet et al. 1999). The effects of increased atmospheric CO2 concentrations can have opposite feedbacks on the capacity of soil to mineralize. The increases of plant biomass and litter and microbial biomass under increased atmospheric CO2 concentrations suggest a possible increase in soil enzyme activity linked to higher soil microbial biomass (Dhillion et al. 1996). On the other hand, several reportas have suggested a slow-down of litter decomposition and mineralization rates under increased atmospheric CO2 concentrations in Mediterranean ecosystems (Gahrooee et al. 1998; Cotrufo et al. 1999; De Angelis et al. 2000) linked to an increase of litter C:N and C:lignin (De Angelis et al. 2000) and C:P (Cotrufo et al. 1999) ratios that can counteract the higher soil enzyme activity. In fact, high concentrations of atmospheric CO2 over 2 years had no important direct effect on soil nitrification and denitrification (Pinay et al. 2007). Higher concentrations of atmospheric CO2 can increase the short-term turnover of C due to higher growth and litter production, but this effect decreases in the medium term by the limitation of nutrients (Cotrufo et al. 2005). Differential effects of the enhancement of atmospheric concentrations of CO2 in different species of plants have been observed in mycorrhizal-plant symbioses where the presence of mycorrhizae increases in the roots of some species and decreases in others (Rillig et al. 1998b) (Table 2). An increase in phenolics in leaf litter decreases N mineralization (Castells et al. 2004), also suggesting a change in the nutritional quality of plant biomass for herbivores. Nevertheless, all these chemical changes can be buffered under long-term exposure to elevated CO2, as observed in plants in the vicinity of CO2 springs, suggesting an evolutionary adaptive response in Mediterranean plants to changes in levels of CO2 (Peñuelas et al. 2002b).
Climatic changes impact on plant-soil relationships
The largest effects of climatic change in Mediterranean ecosystems (IPCC 2007) may well be those related to increased drought, since water stress is already the principal constraint in the Mediterranean region (Specht 1979; Mooney 1989). Enhanced drought is expected to be accompanied by more severe torrential rainfall (Frei et al. 1998), already frequent events (Romero et al. 1998). Over the last century, temperatures have shown an overall trend toward warming (Kutiel and Maheras 1998; Peñuelas et al. 2002a). Precipitation has exhibited either a long-term downward trend, principally in the dry season (Kutiel et al. 1996; Esteban-Parra et al. 1998), or no significant change (Piñol et al. 1998; Peñuelas and Boada 2003), but potential evapotranspiration has risen in all cases.
Warming
The effects of climatic change on nutrient supply in Mediterranean areas may depend on a trade-off between warming and the effects of drought. Concentrations of nutrient in the biomass result from the balance of nutrient uptake and growth. Increased temperatures in these water-limited ecosystems will further decrease the availability of soil water by enhancing evapotranspiration (Larcher 2000), which will very likely result in a decrease in the uptake of nutrients by roots and in the movement of these nutrients to shoots (Bradford and Hsiao 1982) alltogether having a general negative effect on soil activity and plant growth. Warming can also have a direct effect during cold and wetter winters by increasing soil enzyme activities (Sardans et al. 2006d), extending growth periods (Peñuelas and Filella 2001; Peñuelas et al. 2002a), and raising photosynthetic rates (Llorens et al. 2003b). This positive effect during the cold season could be counterbalanced by a lower availability of water in the summer and by shortening the growing season by extending the duration of water deficits (Llorens et al. 2003b; Peñuelas et al. in preparation). The effect of warming on plant growth depends on total annual precipitation. Warming has positive effects on growth in wetter years and no or even negative effects in drier years (Swarbreck et al. 2011). In a study analyzing long-term temporal metadata in Mediterranean areas, changes in precipitation may even have had a relatively higher importance than changes in temperature in explaining phenological shifts in some Mediterranean plant communities (Peñuelas et al. 2004). As mentioned above, some studies have observed a threshold value of temperature showing that during periods with temperatures above 20 °C soil activity is strongly dependent of soil moisture (Rey et al. 2011). In Mediterranean regions, warming only increases soil enzyme activities (Sardans et al. 2008c; Sardans and Peñuelas 2010) and SOM mineralization in the year periods in which soil moisture is high (Bottner et al. 2000; Qi and Xu 2001; Sardans et al. 2006d). The effects of warming at the levels projected by climatic models (IPCC 2007) in arid environments such as Mediterranean areas are thus related to the indirect effect of decreasing soil-water content (Peñuelas et al. 2004; Swarbreck et al. 2011; Sardans et al. 2012b).
Some studies have observed that N mineralization is enhanced more rapidly than P mineralization under warming (Rinnan et al. 2007). This study did not refer specifically to Mediterranean ecosystems but in Mediterranean areas this could also occur, and the increases in available soil N:P ratios could further increase in some Mediterranean areas where the N deposition is increasing in the last years (Vourlitis and Pasquini 2009; Pinho et al. 2012). In fact, experiments of N fertilization in Mediterranean terrestrial ecosystems have shown an increased water-use efficiency (WUE) (Guerrieri et al. 2010), a response that could have positive effect on plant production under warming. However, other possible impacts such as changes in competitive relationships between species adapted to N-limitation and adapted to P-limitation remain to be investigated. In this respect some studies have observed that the changes in nutrient cycling in Mediterranean ecosystems under global change are strongly related to community composition shifts (Maestre et al. 2012).
Drought
The decline in total rainfall and/or availability of soil water projected for the coming decades (IPCC 2007) may prove to be even more drastic under warmer conditions with a CO2-rich atmosphere and higher demand for water (Piñol et al. 1998; Peñuelas et al. 2005). Some studies investigating the impact on Mediterranean ecosystems of increased atmospheric CO2 concentrations, drought and warming suggest that most future scenarios of drought combined with warming and/or with increased atmospheric CO2 concentrations may involve strong impacts on soil-water deficits and water use in trees of Mediterranean forests by increasing soil water deficit (Hernández-Santana et al. 2008).
Plants
Increases in drought intensity at the level projected for the coming decades (IPCC 2007) could reduce soil moisture (Sardans et al. 2008a, b), plant growth and cover in the short term (Boix-Fayos et al. 1998; Goldstein et al. 2000; Beier et al. 2004; Serrano et al. 2005; Tsiafouli et al. 2005; Ogaya and Peñuelas 2007a; Sardans et al. 2008a, b; Prieto et al. 2009a, b; Pérez-Ramos et al. 2010; Cotrufo et al. 2011; Talmon et al. 2011) (Table 3, Table S3 in the supplementary material). Moreover, plant cover can be seriously threatened by the increase of fire risk projected in the Mediterranean area in the context of enhanced drought (Mouillot et al. 2002; Moriondo et al. 2006).
Drought also induces changes in foliar nutrient contents, with a general trend to reduce foliar N (Sardans et al. 2008a, b), P (Sardans and Peñuelas 2004, 2007a; Sardans et al. 2008g), and micronutrients (Sardans et al. 2008e, f), which can also change in other plant tissues (Sardans and Peñuelas 2004, 2007a, 2008b: Sardans et al. 2006b, 2008a, b, e, f, g; Sardans and Peñuelas 2008). The nutrient content of aboveground biomass, mainly in leaves, tends to decrease with drought (Sardans et al. 2008a; b), with the exception of K that increases in some aboveground tissues of some plant species (Sardans and Peñuelas 2007a; Sardans et al. 2008g). Drought increases the C:N and C:P ratios of Mediterranean shrubs and trees, associated with a protective and conservative mechanism (Inclan et al. 2005; Sardans et al. 2008b, c, d; Matías et al. 2011) and with sclerophylly and increases in lignin content (Bussotti et al. 2000; Sardans et al. 2006a). Drought enhances the allocation of N to the root system, which increases the capacity to absorb water (Sardans et al. 2008a; González et al. 2010). Drought also increases the oxidative stress of Mediterranean plants (Munné-Bosch and Peñuelas 2003 and 2004). Under moderate conditions of drought, photosynthetic pigments remain more or less constant, and anti-stress compounds such as α-tocopherol increase in concentration. When stress from drought is severe, the concentration of photosynthetic pigments falls, and anti-stress pigments and secondary compounds can increase more (Munné-Bosch and Peñuelas 2004; Hernandez et al. 2004) or even decrease (Bettaieb et al. 2011) due to the loss of production capacity. Drought can induce foliar senescence that contributes to the remobilization of nutrients during stress, thus allowing the rest of the plant, including new leaves, to benefit from the nutrients accumulated during the foliar life span (Munné-Bosch and Alegre 2004). Sclerophylly is a typical and general trait of evergreen Mediterranean plants and usually increases when the environment evolves toward drier conditions (Dunn et al. 1977; Sabaté et al. 1992; Oliveira et al. 1994).
During drought, different plant species of the same community exhibit differential decreases in the growth of aboveground biomass (Mamolos et al. 2001; Ogaya and Peñuelas 2007b; Prieto et al. 2009a, b), changes in foliar nutrient concentrations and contents (Peñuelas et al. 2008b; Sardans et al. 2008a, b, e, f; Sardans and Peñuelas 2008), and effects on hydraulic conductivity (Mereu et al. 2009) (Table 3). These differences in growth and nutrient uptake during drought are related to species differences in the capacity to maintain ecophysiological traits, such as photosynthetic rate, WUE, regulation of foliar gas exchange (Peñuelas et al. 2000; Llorens et al. 2003a, b; Ogaya and Peñuelas 2003; Ozturk et al. 2010; Vilagrosa et al. 2010; Galle et al. 2011), capacity to protect against oxidative damage (Munné-Bosch and Peñuelas 2004), root depth (Padilla and Pugnaire 2007; West et al. 2012) and capacity to reabsorb N (Sanz-Pérez et al. 2009).
The relationships between hydraulic architecture and leaf area are a key factor determining drought adaption success. Drought-induced mortality in Mediterranean pines has proved to be related to lower hydraulic conductivity per unit of leaf area (Martínez-Vilalta and Piñol 2002). The resistance to xylem embolism allows to maintain higher stomatal conductance and sap flow even at lower water potentials thus permitting large WUE (Martínez-Vilalta et al. 2003). The survival capacity to drought has been proved to be also related to carbon reserves (Volaire 1995; Galiano et al. 2012). Under drought, the Mediterranean tree Olea europea stops growth but not photosynthetic activity that allows it to store assimilates, produce antioxidants, antiherbivore compounds, such as phenolics, and osmoprotective substances, mainly water soluble sugars allowing to maintain cell turgor (Volaire 1995; Sofo et al. 2008). Mediterranean trees usually present lower stomatal sensitivity to drought in seedling phase than in adult phase, probably in benefit of an increase in growth under competition with understory vegetation, making them more vulnerable to drought events in initial phases of growth (Mediavilla and Escudero 2004).
Soils
As discussed above, moderate reductions in rainfall imply significant decreases in soil moisture. In addition to thes direct effect, reduced rainfall can indirectly affect plants by reducing growth and increasing C:nutrient ratios. Some studies have also reported increases in SOM (Talmon et al. 2011). Most reports recommend the differentiation between short-term and medium- and long-term effects of drought on SOM (Ogaya and Peñuelas 2006; Martí-Roure et al. 2011; Navarro-García et al. 2012). In the short term, drought can increase SOM by increasing the total amount of litterfall (Oliveira and Peñuelas 2002; Ogaya and Peñuelas 2006; Martí-Roure et al. 2011) and dead roots (Lloret et al. 1999; Padilla and Pugnaire 2007). Few long-term experiments on the effect of drought on SOM have been conducted, but observational studies under prolonged and sustained drought or across environmental gradients suggest that SOM decreases by the reduction of plant cover, implying a decrease in litterfall and an increase in soil erosion by diminishing soil protection and permeability (Boix-Fayos et al. 1998; Li et al. 2011; Navarro-García et al. 2012).
SOM mineralization is very sensitive to drought in Mediterranean ecosystems (Table 3). Decreases in soil enzyme activity (Zaman et al. 1999; Fioretto et al. 2000; Li and Sarah 2003; Sardans and Peñuelas 2005, 2010; Sardans et al. 2006d; Sardans et al. 2008c, d; Hueso et al. 2011), soil respiration (Bottner et al. 2000; Qi and Xu 2001; Emmett et al. 2004; Asensio et al. 2007), and root enzyme activity (Sardans et al. 2007) have been widely observed under drought conditions. The decrease in soil enzyme activity is mainly due to the direct effect of the reduction in soil moisture (Sardans et al. 2008c: Sardans and Peñuelas 2010) but also to the higher C:nutrient ratio in plant litter (Wessel et al. 2004). This decrease in the nutritional quality of the SOM can have a significant effect on the rates of litter decomposition (Coûteaux et al. 2002).
Another interesting but poorly studied aspect is the effect of drought on the structure of the community of decomposers. Curiel-Yuste et al. (2011) observed that fungi became more abundant and active than bacteria when soils were drier. More associations of arbuscular mycorrhizal fungi increase the tolerance of plants to drought (Benabdellah et al. 2011). Changes in the community of soil decomposers under drought should be the focus of new studies because these changes can be very important for understanding the capacity of Mediterranean ecosystems to adapt to increasing conditions of drought. Increases in fungal biomass can increase the capacity of plants to take up water and/or of soil to retain more water but can also change the capacity of nutrient cycling. Further research is needed to fill this gap.
Mediterranean ecosystems under drought show no clear patterns of shifts in N:P ratios (Sardans et al. 2008a, b, 2012b). Different compensatory effects probably occur in the cycles of these two nutrients. P is frequently retranslocated more than N (Sardans et al. 2005a), but the comparative changes in N and P retranslocation with increasing conditions of drought is not known. On the other hand, increases of soil moisture decrease the fraction of P available in soils and increase the fraction of unavailable P (Sardans and Peñuelas 2004; Sardans et al. 2008g). N is also more soluble and easily leached than P and can be more lixiviated from soil by typical Mediterranean storms when plant cover is reduced under drought and in physically degraded soils. How increases in drought will affect Mediterranean ecosystems by changing N:P ratios remains unclear, but the N:P ratio is an important variable favoring species having different rates of growth (Sardans et al. 2012c).
Reductions in water uptake, mobility of nutrients in the soil, and release of nutrients from the SOM can exert a feedback contributing to reduce the capacity of plants under drought to take up nutrients by slowing the cycling of nutrients (Sardans et al. 2012b). Increasing nutrient concentrations in soils increase the risk of nutrient losses caused by runoff during torrential rainfalls (Sardans and Peñuelas 2004; Matías et al. 2011). Drought projected for the coming decades should not sufficiently change the concentrations of trace elements in the medium term to constitute any environmental or health problems (Sardans and Peñuelas 2007b; Sardans et al. 2008h).
System adaptation capacity
The long-term evolutionary adaptation of Mediterranean plants to long dry summers allows them to cope with moderate increases of drought without an important loss of their production capacity and survival (Cotrufo et al. 2011). Mediterranean shrubs subject to drought increase their levels of antioxidative mechanisms to prevent injuries from increased oxidative stress (Munné-Bosch and Alegre 2000; Munné-Bosch and Peñuelas 2003 and 2004; Peguero-Pina et al. 2008; Nogues et al. 2012). Other processes enhanced by drought can contribute to increase the capacity of plants to take up water. For example, Benabdellah et al. (2011) observed an increase in arbuscular mycorrhizae that increased the capacity of plants to take up water. Nutrient retranslocation increased when drought increased (Correira and Martinez-Loucao 1997), which increased the concentrations of nutrients in plants and thus contributed to an improvement in WUE. Higher nutrient retranslocation, however, should increase the C:nutrient ratio in litter and contribute to the formation of more recalcitrant SOM and to the slowing of nutrient cycling. The advantages and disadvantages of nutrient retranslocation under increasing conditions of drought remain unclear and deserve further research.
Additional effects on C-cycling and emissions of biogenic volatile organic compounds
Drought usually increases emissions of plant terpenes, although this effect strongly depends on drought intensity (Pegoraro et al. 2005; Plaza et al. 2005; Llusia et al. 2006, 2008, 2010). Studies suggest that severe drought decreases the emission of terpenes (Loreto et al. 2001; Pegoraro et al. 2007; Peñuelas and Staudt 2010) but reduces sink strength of the soil for atmospheric isoprene (Pegoraro et al. 2007). The effects of drought on the emission of terpenes from Mediterranean soils remain unclear and vary for different VOC compounds (Asensio et al. 2008).
Rewetting events after a period of drought have a large impact on soil respiration and microbial activity and increase the loss of C from soil (Navarro-García et al. 2012). After rewetting events, as for example after a rain event in a drought period, an increase of soil heterotrophic respiration from litter and other soil organic fractions (Rey et al. 2005; Jarvis et al. 2007; Casals et al. 2011; Placella et al. 2012) and increases in the activity of some soil enzymes (Zornoza et al. 2006) have been widely observed. The capacity to faster resuscitation capacity of certain microbial groups after wet-up of dry soils may be a phylogenetically conserved ecological property in Mediterranean soils (Placella et al. 2012). This process can imply a sudden increase in nutrient availability, Butterly et al. (2009) observed a 35–40 % increase of available P. However, this effect drops during the first days after rain (Butterly et al. 2009). These effects are due to the increased water availability and to the increase of SOC made accessible to microorganisms by the physical destruction of soil structure by drying/wetting events (Inglima et al. 2009; Unger et al. 2010; Navarro-García et al. 2012) and by the increases of osmoregulatory substances released from microorganisms in response to hypo-osmotic stress (in order to avoid cell lyses) that increasses the organic source to respiration (Unger et al. 2010). In the short term, rewetting causes a cascade of effects, from increases of SOC mineralization and availability of nutrients (Nguyen and Marschner 2005; Inglima et al. 2009) to short periods of plant growth (Xiang et al. 2008).
Desertification
Climate change scenarios projected for the coming decades predict a reduction in total precipitation and a higher frequency and severity of torrential rainfall (Frei et al. 1998; Romero et al. 1998; IPCC 2007 The most threatening phenomenon for Mediterranean soils, especially in the most xeric areas, is desertification linked to a continual positive feedback of higher frequency and intensity of torrential rainfall (associated to climatic change) with an increase of soil erosion, which in turn leads to a loss of soil fertility and thus plant cover. These facts can also be aggravated by human activities (such as excessive livestock pressure) and by the increasing frequency and extend of fires, specially in European Mediterranean region (Shakesby 2011), that then leads to an increased risk of erosion and desertification. The increases of fires intensity and frequency is a serious concern (Shakesby 2011). Moreover the investigations of community succession and soil processes after fires in Mediterranean plant communities create plant patches (Baeza et al. 2007) and soil degradation, reducing water infiltration (Coelho et al. 2004), all of them leading towards desertification process. During the last decades, several Mediterranean areas of the EU countries have been subjected to land-use change (Millan et al. 2005; Zucca et al. 2006; Calatrava et al. 2011; Salvati et al. 2012), whereby unproductive areas were forested or abandoned or submitted to overexploitation (Calatrava et al. 2011). It appears that these processes favour a general trend towards an increased cover of semi-arid shrublands, and in the worst cases to desertification (Zucca et al. 2006) and may even decrease rain events (Millan et al. 2005). This cascade of positive feedbacks among these four variables has been observed in some studies in southern Spain (Garcia et al. 2002; Moreno-de las Heras et al. 2011; Ruiz-Sinoga et al. 2011 and 2012) and is exponentially favored and accelerated by the increase in aridity (Ruiz-Sinoga et al. 2011). The erosion of soils in Mediterranean areas is strongly related to torrential rainfall (González-Hidalgo et al. 2007). Slopes and higher erodible bedrock accelerate this process (Moreno-de las Heras et al. 2011). Moreover, photodegradation of litter rises when plant cover decreases (Barnes et al. 2012), thus increasing the SOM losses and contributing to soil degradation. Soil erosion is becoming the primary environmental concern in human economic balances in Mediterranean regions (Riera et al. 2007).
Future research directions
Mediterranean plants resist moderate increases in drought without significant changes in their production capacity and survival and are able to maintain levels of soil fertility. These capacities are species specific. The mortality observed in some forested Mediterranean areas after severely dry summers varies in different species of the same community (Hernández-Santana et al. 2009). While we do not know the real level of future droughts, we can expect an increasing relative cover of more drought-adapted species within communities. The impact of this situation on the soil is less clear because if the drought is not severe, a new equilibrium in the plant-soil system may be attained. Therefore, further research is needed.
The interaction between atmospheric CO2 and increased drought should be studied in long-term experiments, because a short-term counteracting effect has been observed. At long-term, the acclimation of plants to increased atmospheric CO2 concentrations could diminish the compensatory effect of the increasing drought intensity. Similarly, warming can have a positive effect on ecosystems such as alpine areas by reducing the length of the cold season that can compensate the negative effects on production capacity resulting from the increased water stress during summer. These last contradictory effects have not been thoroughly studied, at least to the best of our knowledge, and should be a clear objective for further studies. If drought is more severe, and longer dry summers coincide with more severe torrential rainfall, a strong influence on soil erosion can be expected, as occurs in some areas of southeastern Spain (Ruiz-Sinoga et al. 2011 and 2012). Moreover, the projected increased drought could be even more detrimental for Mediterranean ecosystems if it occurs in spring (Allard et al. 2008). In addition to the environmental impact, an increase in the area of shrubland affected by erosion is a key concern, for several reasons. An increase of 1 % in the current area of shrubland of the Mediterranean Basin could cause an enormous economic impact (Riera et al. 2007).
Summarizing the information in the literature, we can identify two different groups of feedbacks in Mediterranean plant-soil systems in response to the scarcity of water and nutrients in soils. Firstly, a high allocation to a deep, strong, and dense root system together with a notable plant cover and investment in soil enzyme production exerts a positive effect on soil fertility and protection capacity. Secondly but in contrast, a high nutrient retranslocation, high retention of nutrients in standing biomass, and high C:nutrient content of litter constitute constraints for a rapid release of nutrients from the SOM, lowering fertility and slowing the soil processes that produce high quality SOM. Current studies indicate that drought enhancement slows the first group of positive properties for soil fertility and protection but intensifies the second group of negative properties.
Plant cover, soil biological activity, and fertility are interconnected. The causes underlying the high soil enzyme activity observed in Mediterranean ecosystems warrants future research. Most efforts to date have investigated the relationships between soil moisture, temperature, and food quality of litter in terms of lignin content or C:nutrient ratio, but Mediterranean plants may possess mechanisms for stimulating soil enzyme production. Because the current data suggest that extensive root systems can significantly impact soil enzymes, the introduction of soil genomics, proteomics, and metabolomics should be encouraged to gain knowledge of the role of plants, through rhizospheric activity and root exudates, on soil microbial communities, on soil enzyme activity, soil mineralization, and soil texture. Moreover, these studies should investigate the role of metabolites at different levels and in different processes, such as the allelopathic inhibition of growth of competing vegetation, stimulation of microbial and especially fungal growth, or changes in soil structure by contributing to the formation of soil aggregates.
Studies in Mediterranean areas suggest a key role for the root system mediated by the availability of water. A more thorough knowledge of root structure and distribution of entire plant communities, from forests in mesic areas to sparse shrublands in drier areas, should be another main objective for research in this area. This research would provide a better understanding of the shifts that occur in plant communities when climate evolves to a different pattern of precipitation. The study of how root systems and particularly the deepest roots, which frequently enter fractures in the bedrock, pump water from the deepest to the upper soil layers warrants urgent research. How root systems maintain higher levels of soil mineralization and activity also warrants further research. Isotopic methods should be a useful tool in these studies by allowing the monitoring of the movement of water in soil-plant systems.
We also lack knowledge of the interacting effects of drought and warming in realistic field scenarios and in different conditions, from cold alpine forested areas to dry, warm lowland areas. The presence of large gradients of water availability and temperature in Mediterranean alpine areas, frequently over short distances, constitutes an excellent playground for investigating the role of soil moisture and temperature and their interactions in modulating the active and passive mechanisms of plants that affect soil texture and fertility. Moreover, these studies could help us to better understand the capacity of response of the different Mediterranean ecosystems to the projected increase in drought. In the context of future scenarios of global change, field experiments are needed for studying the long-term effects of high concentrations of atmospheric CO2 combined with drier conditions, because this future scenario is very likely and because the current but incomplete results strongly suggest that the effects of increasing concentrations of atmospheric CO2 on plants could decline with time.
References
Aerts R (1995) The advantages of being evergreen. Trees 10:402–406
Agra HE, Ne’eman G (2012) Composition and diversity of herbaceous patches in woody vegetation: the effects of grazing, soil seed bank, patch spatial properties and scale. Flora 207:310–317
Allard V, Ourcival JM, Rambal S, Joffre R, Rocheteau A (2008) Seasonal and annual variation of carbon exchange in an evergreen Mediterranean forest in southern France. Global Change Biol 14:714–715
Almagro M, Martínez-Mena M (2012) Exporting short-term leaf-litter decomposition dynamics in a Mediterranean ecosystem: dependence on litter type and site conditions. Plant Soil. doi: 10.1007/s11104-012-1187-6
Almagro M, López J, Querejeta JI, Martínez-Mea M (2009) Temperatura dependence of soil CO2 efflux is strongly modulated by seasonal petterns of moisture availability in a Mediterranean ecosystem. Soil Biol Biochem 41:594–605
Andreetta A, Ciampalini R, Moretti P, Vingiani S, Poggio G, Matteucci G, Tescari F, Carnicelli S (2011) Forest humus forms as potential indicators of soil carbon storage in Mediterranean environments. Biol Fertil Soils 47:31–40
Andreetta A, Macci C, Ceccherini MT, Cecchini G, Masciandaro G, Pietramellara G, Carnicelli S (2012) Microbial dynamics in Mediterranean moder humus. Biol Fertil Soils 48:259–270
Andreu V, Rubio JL, Gimeno-García E, Llinares JV (1998) Testing three Mediterranean shrub species in runoff reduction and sediment transport. Soil Tillage Res 45:441–454
Andreu-Hayles L, Planells O, Gutierrez E, Muntan E, Helle G, Anchukaitis KJ, Schleser GH (2011) Long tree-ring chronologies reveal 20th century increases in water-use efficiency but no enhancement of tree growth at five Iberian pine forest. Global Change Biol 17:2095–2112
Aponte C, Marañón T, García LV (2010) Microbial C, N and P in soils of Mediterranean oak forest: influence of season, canopy cover and soil depth. Biogeochemistry 101:77–92
Arianoutsou M, Thanos CA (1996) Legumes in the fire-prone Mediterranean regions: an example from Greece. Wedland Fire 6:77–82
Asensio D, Peñuelas J, Ogaya R, Llusia J (2007) Seasonal soil and CO2 exchange rates in a Mediterranean holm oak forest and their responses to drought conditions. Atmos Env 41:2447–2455
Asensio D, Peñuelas J, Prieto P, Estiarte M, Filella I, Llusia J (2008) Interannual and seasonal changes in the soil exchange rates of monoterpenes and other VOCs in a Mediterranean shrubland. Eu J Soil Sci 59:878–891
Austin AT, Ballaré CL (2010) Dual role of lignin in plant litter decomposition in terrestrial ecosystems. Proc Natl Acad Sci U S A 107:4618–4622
Austin AT, Vivanco L (2006) Plant litter decomposition in a semi-arid ecosystem controlled by photodegradation. Nature 442:555–558
Azcón R, Barea JM (1997) Mycorrhizal dependency of a representative plant species in mediterranean shrublands (Lavandula spica L.) as a key factor to its use for revegetation strategies in desertification-threatened areas. Appl Soil Ecol 7:83–92
Baeza MJ, Valdecantos A, Alloza JA, Vallejo R (2007) Human disturbance and environmental factors as drivers of long-term post-fire regeneration patterns in Mediterranean forest. J Veg Sci 18:243–252
Baldocchi DD, Xu L (2007) What limits evapotranspiration from Mediterranean oak woodlands-The supply of moisture in the soil, physiological control by plants or the demand by the atmosphere? Adv Water Resour 30:2113–2122
Ballaré CL, Caldwell MM, Flint SD, Robinson SA, Bornman JF (2011) Effects of solar ultraviolet radiation on terrestrial ecosystems. Patterns mechanisms, and interactions with climate change. Photochem Phtobiol Sci 10:226–241
Baraza E, Hodar JA, Zamora R (2009) Species, site and seasonal variation in leaf-chemistry diversity of woody Mediterranean plants. Rev D’Ecol – Terre Vie 64:135–144
Barea JM, Palenzuela J, Cornejo P, Sánchez-Castro I, Navarro-Fernández C, López-García A, Estrada B, Azcón R, Ferrol N, Azcón-Aguilar C (2011) Ecological and functional roles of mycorrhizas in semi-arid ecosystems of Southeast Spain. J Arid Env 75:1292–1301
Barnes PW, Throop HL, Hewins DB, Abbene ML, Archer SR (2012) Soil coverage reduces photodegradation and promotes the development of soil-microbial films on dryland leaf litter. Ecosystems 15:311–321
Barthes B, Roose E (2002) Aggregate stability as an indicator of soil susceptibility to runoff and erosion: validation at several levels. Catena 47:133–149
Bassirirad H, Constable JVH, Lussenhof J, Kimball BA, Norby RJ, Oechel WC, Reich PB, Schlesinger WH, Zitzer S, Sehtiya HL, Silim S (2003) Widespread foliage δ15N depletion under elevated CO2 inferences for the nitrogen cycle. Global Change Biol 9:1582–1590
Bastida F, Barberá GG, García C, Hernández T (2008) Influence of orientation, vegetation and season on soil microbial and biochemical characteristics under semiarid conditions. Appl Soil Ecol 38:62–70
Beier C, Emmett B, Gundersen P, TIetema A, Peñuelas J, Estiarte M, Gordon C, Gorissen A, Llorens L, Rodà F, Williams D (2004) Novel Approaches to study Climate Change effects on terrestial ecosystems in the field: drought and passive nighttime warming. Ecosystems 7:583–597
Beier C, Emmett BA, Tietema A, Schmidt IK, Peñuelas J, Lang EK, Duce P, De Angelis P, Gorissen A, Estiarte M, de Pato GD, Sowerby A, Kröel-Dulay G, Lellei-Kovacs E, Kull O, Mand P, Petersen H, Gjelstrup P, Spano D (2009) Carbon and nitrogen balances for six shrublands across Europe. Global Biogeochem Cycles 23:GB4008
Bell DT (2001) Ecological response syndromes in the flora of southern western Australia: fire resprouters versus reeseders. Bot Rev 67:417–440
Bell TL, Adams MA (2004) Ecophysiology of ectomycorrhizal fungi associated with Pinus app. In low rainfall areas of western Australia. Plant Ecol 171:35–52
Bell LW, Williams AH, Ryan MH, Ewing MA (2007) Water relations and adaptations to increasing water deficit in three perennial legumes, Medicago sativa, Dorycnium hirsutum and Dorycnium rectum. Plant Soil 290:231–243
Benabdellah K, Abbas Y, Abourough M, Aroca R, Azcón R (2011) Influence of two bacterial isolates from degraded and non-degraded soils and arbuscular mycorrhizae fungi isolated from semi-arid zone growth of Trifolium repens under drought conditions: Mechanisms related to bacterial effectiveness. Eu J Soil Biol 47:303–309
Bengtsson J, Janion C, Chown SL, Leinaas HP (2012) Litter decomposition in fynbos vegetation, south Africa. Soil Biol Biochem 47:100–105
Bettaieb I, Mamrouni-Sallami I, Bourgou S, Limam F, Marzouk B (2011) Drought effects on polyphenol composition and antioxidant activities in aerial parts of Salvia officinalis L. Acta Phisiol Plantarum 33:1103–1111
Bettarini I, Calderoni G, Miglieta F, Rasch A, Ehleringer J (1995) Isotopic carbon discrimination and leaf nitrogen content of Erica arborea L. along a CO2 concentration gradient in a CO2 spring in Italy. Tree Physiol 15:327–332
Blank L, Carmel Y (2012) Woody vegetation patch types affect herbaceous species richness and composition in a Mediterranean ecosystems. Commun Ecol 13:72–81
Blaschke L, Schulte M, Raschi A, Slee N, Rennenberg H, Polle A (2001) Photosynthesis, soluble and structural carbon compounds in two Mediterranean oak species (Quercus pubescens and Q. ilex) after lifetime growth at naturally elevated CO2 concentrations. Plan Biol 3:288–298
Boix-Fayos C, Calvo-Cases A, Imeson AC, Soriano-Soto MD, Tiemessen IR (1998) Spatial and short-term temporal variations in runoff, soil aggregation and other soil properties along a Mediterranean climatological gradient. Catena 33:123–128
Bon WJ, Midgley JI (2003) The evolutionary ecology of sprouting in woody plants. Int J Plant Sci 164:5105–5114
Bonanomi G, Incerti G, Antignani V, Capodilupo M, Mazzoleni S (2010) Decomposition and nutrient dynamics in mixed litter of Mediterranean species. Plant Soil 331:481–496
Bonkowski M, Roy J (2012) Decomposer community complexity affects plant competition in a model early successional grassland community. Soil Biol Biochem 46:41–48
Bottner P, Couteaux MM, Anderson JM, Berg B, Billès G, Bolger T, Casablanca H, Romanyá J, Rovira P (2000) Decomposition of 13C-labelled plant material in a European 65–40º latitudinal transect of coniferous forest soils: simulation of climate change by translocation of soils. Soil Biol Biochem 32:527–543
Bradford KJ, Hsiao TC (1982) Physiological responses to moderate water stress. In: Lange OL, Lange PS, Nobel CB, Osmond J, Ziegler H (eds) Physiological Plant Ecology II. Water Relations and Carbon Assimilation, vol 12 B. Springer, Berlin, pp 263–342
Brueck H (2008) Effects of nitrogen supply on water-use efficiency of higher plants. J Plant Nutr Soil Sci 171:210–219
Bussotti F, Borghini F, Celesti C, Leonzio C, Bruschi P (2000) Leaf morphology and macronutrients in broadleaved trees in central Italy. Trees 14:361–368
Butterly CR, Bünemann EK, McNeill AM, Baldock JA, Marschner P (2009) Carbon pulses but not phosphorus pulses are related to decreases in microbial biomass during repeated drying and rewetting of soils. Soil Biol Biochem 41:1406–1416
Çakan H, Karatay C (2006) Interactions between mycorrhizal colonization and plant life forms along the successional gradient of coastal sand dunes in the eastern Mediterranean, Turkey. Ecol Res 21:301–310
Calatrava J, Barbera GG, Castillo VM (2011) Farming practiques and policy measures for agricultural soil conservation in semi-arid Mediterranean areas: the case of the Guadalentin basin in Southeast Spain. Land Degr Develop 22:58–69
Carbone MS, Still CJ, Ambrose AR, Dawson TE, Williams AP, Boot CM, Schaeffer SM, Schimel JP (2011) Seasonal and episodic moisture controls on plant and microbial contributions to soil respiration. Oecologia 167:265–278
Carreira JA, Latjha K (1997) Factors affecting phosphate sorption along a Mediterranean, dolomitic soil and vegetation chronosequence. Eur J Soil Sci 48:139–149
Carreira JA, Asensi A, Niell FX (1991) Sustitución de comunidades y concentración de nutrientes en plantas tras incendios forestales en una cuenca subárida del S.E. de España. Orsis 6:199–214
Carreira JA, Sánchez-Vázquez F, Niell FX (1992) Short-termand small-scale patterns of post-fire regeneration in a semi-ariddolomitic basin of Southern Spain. Acta Oecol 13:241–253
Casals P, López-Sangil L, Carrara A, Gimeno C, Nogues S (2011) Autotrophic and heterotrophic contributions to short-term soil CO2 efflux following simulated summer precipitation pulses in a Mediterranean dehesa. Global Biogeochemical Cycles 25:GB3012.
Castaldi S, Carfora A, Natale A, Messere A, Miglieta F, Cotrufo MF (2009) Inhibition of net nitrification activity in a Mediterranean woodland: possible role of chemicals produced by Arbutus unedo. Plant Soil 315:273–283
Castell C, Terradas J, Tenhunen JD (1994) Water relations, gas exhange, and growth of resprouts and mature plant shoots of Arbutus Unedo L. and Quercus ilex L. Oecologia 98:201–211
Castells E, Peñuelas J (2003) Is there a feedback between N availability in siliceous and calcareous soil and Cistus albidus leaf chemical composition? Oecologia 136:183–192
Castells E, Roumet C, Peñuelas J, Roy J (2002) Intraspecific variability of phenolic concentrations and their responses to elevated CO2 in two Mediterranean perennial grasses. Env Exp Bot 47:205–216
Castells E, Peñuelas J, Valentine DW (2004) Are phenolic compounds released from the Mediterranean shrub Cistus albidus responsible for changes in N cycling in siliceous and calcareous soils? New Phytol 162:187–195
Castillo-Monroy AP, Maestre FT, Delgado-Baquerizo M, Gallardo A (2010) Biological soil crust modúlate nitrogen availability in semi-arid ecosystems: insights from a Mediterranean grassland. Plant Soil 333:21–34
Chirino E, Bonet A, Bellot J, Sánchez JR (2006) Effects of 30-year-old Aleppo pine plantations on runoff, soil erosion, and plant diversity in a semi-arid landscape in south eastern Spain. Catena 65:19–29
Coelho CDOA, Ferreira AJD, Boulet AK, Keizer JJ (2004) Overland flow generation processes, erosion yields and solute loss following different intensity fires. Quar J Engin Geol Hydrogeol 37:233–240
Corcuera L, Camarero JJ, Gil-Pelegrín E (2004) Effects of a severe drought on Quercus ilex radial growth and xylem abatomy. Trees 18:83–92
Correira PJ, Martinez-Loucao MA (1997) Leaf nutrient variation in mature carob (Ceratonia siliqua) trees in response to irrigation and fertilization. Tree Phisiol 17:813–819
Cortez J, Demard JM, Bottner P, Monrozier J (1996) Decomposition of Mediterranean leaf litters: a microcosm experiment investigating relationships between decomposition rates and litter quality. Soil Biol Biochem 28:443–452
Cortez J, Garnier E, Pérez-Harguindeguy N, Debussche M, Gillton D (2007) Plant traits, litter quality and decomposition in a Mediterranean old-field succession. Plant Soil 296:19–34
Cotrufo MF, Raschi A, Lanini M, Ineson P (1999) Decomposition and nutrient dynamics of Quercus pubescens leaf litter in a naturally enriched CO2 Mediterranean ecosystem. Funct Ecol 13:343–351
Cotrufo MF, De Angelis P, Polle A (2005) Leaf litter production and decomposition in a poplar short-term coppice exposed to free air CO2 enrichmentr (POPFACE). Global Change Biol 11:971–982
Cotrufo MF, Alberti G, Inglima I, Marjanovic H, LeCain D, Zaldei A, Peressotti A, Miglietta F (2011) Decreased summer drought affects plant productivity and soil carbon dynamics in a Mediterranean woodland. Biogeosciences 8:2729–2739
Coulis M, Hattenschwiler S, Rapior S, Coq S (2009) The fate of condensed tannins during litter consumption by soil animals. Soil Biol Biochem 41:2573–2578
Coûteaux MM, Aloui A, Kurz-Besson C (2002) Pinus halepensis litter decomposition in laboratory microcosms as influenced by temperature and a millipede, Glomeris marginata. Appl Soil Ecol 20:85–96
Criquet S, Tagger S, Vogt G, Le Petit J (2002) Endoglucanase and b-glycosidase activities in an evergreen oak litter: annual variation and regulationg factors. Soil Biol Biochem 34:1111–1120
Cruz C, Lips H, Martins-Louçao MA (2003) Nitrogen use efficiency by a slow-growing species as affected by CO2 levels, root temperature, N source and availability. J Plant Physiol 160:1421–1428
Curiel-Yuste JC, Peñuelas J, Estiarte M, Garcia-Mas J, Mattana S, Ogaya R, Pujol M, Sardans J (2011) Drought-resistant fungi control soil organic matter decomposition and its response to temperature. Global Change Biol 17:1475–1486
David TS, Henriques K-B, Nunes VF, Vaz M, Pereira JS, Siegwole R, Chaves MM, Gazarini LC, David JS (2007) Water-use strategies in two co-occuring Mediterranean evergreen oaks: surviving the summer drought. Tree Physiol 27:793–803
De Angelis P, Chigwerewe KS, Scarascia-Mugnozza G (2000) Litter quality and decomposition in a CO2-enriched Mediterranean forest. Plant Soil 224:31–41
De Baets S, Poesen J, Knapen A, Barberá GG, Navarro JA (2007) Root cheracteristics of representative Mediterranean plant species and their erosion-reducing potential during concentrated runoff. Plant Soil 294:169–183
De Baets S, Poesen J, Reubens B, Wemans K, De Baerdemaeker J, Muys B (2008) Root tensile strength and root distribution of typical Mediterranean plant species and their contribution to soil shear strength. Plant Soil 305:207–236
De Luis M, Novak K, Raventós J, Gricar J, Prislan P, Cufar K (2011) Cambial activity, wood formation and sapling survival of Pinus halepensis exposed to different irrigation regimes. For Ecol Manag 262:1630–1638
De Marco A, Mcola A, Maisto G, Giordano M, Virzo de Santo A (2011) Non-additive effects of litter mixtures on decomposition of leaf litters in a Mediterranean maquis. Plant Soil 344:305–317
De Mico V, Aronne G (2009) Seasonal dimorphism in wood anatomy of the Mediterranean Cistus incatus L. subsp. incatus. Trees 23:981–989
Del Cacho M, Lloret F (2012) Resilence of Mediterranean shrubland to a severe drought spisode: the role of seed bank and seedling emergence. Plant Biol 14:458–466
Delfine S, Loreto F, Alvino A (2001) Drought-stress effects on physiology, growth and biomass production of rainfed and irrigated bell pepper plants in the Mediterranean region. J Am Soc Horticul Sci 126:297–304
Dell B, Jones S, Wilson SA (1987) Phosphorus-nutrition of Jarrah (Eucapyptus-marginata) seedlings-use of bark for diagnosing phosphorus deficiency. Plant Soil 97:369–379
Dhillion SS, Roy J, Abrams M (1996) Assessing the impact of elevated CO2 on soil microbial activity in a Mediterranean model ecosystem. Plant Soil 182:333–342
Di Castri F (1981) Mediterranean-type shrublands of the world. In: Di Castri F, Goodall DW, Specht RL (eds) Mediterranean- Type Shrublands. Elsevier, Amsterdam, pp 1–52
Di Castri F, Mooney HA (1973) Mediterranean Type Ecosystems: Origin and Structure. Springer Verlag, New York, pp 11–19
Dias T, Malveiro S, Martins-Louçao MA, Sheppard LJ, Cruz C (2011) Linking N-driven biodiversity changes with soil N availability in a Mediterranean ecosystem. Plant Soil 341:125–126
Díaz-Barradas MC, Zunzunegui M, Ain-Lhout F, Jáuregui J, Boutaleb S, Álvarez-Cansino L, Esquivas MP (2010) Seasonal physiological responses of Arginia spinosa tree from Mediterranean to semi-arid climate. Plant Soil 337:217–231
Dick RP (1997) Soil enzyme activities as integrative indicators of soil health. In: Pankurst CE, Doube BM, Gupta VVSR (eds) Biological Indicators of Soil Health. CAB International, Wellingford, pp 121–157
Dick RP, Breakwill D, Turco R (1996) Soil enzyme activities and biodiversity measurements as integrating biological indicators. In: Doran JW, Jones AJ (eds) Handbook of Methods for Assessment of Soil Quality. SSSA, Madison, pp 247–272
Dirks I, Navon Y, Kanas D, Dumbur R, Grünzweig JM (2010) Atmospheric water vapor as driver of litter decomposition in Mediterranean shrublands and grassland during rainless seasons. Global Change Biol 16:2799–2812
Domíngues-Nuñez JA, Selva Serrano J, Rodríguez Barreal JA, de Omeñaca S, González JA (2006) The influence of mycorrhization with Tuber melanosporum in the afforestation of a Mediterranean site with Quercus ilex and Quercus faginea. For Ecol Manag 231:226–233
Dunn EL, Shropshire FM, Strong LC, Mooney HA (1977) The water factor and convergent evolution in Mediterranean-type vegetation. In: Lange L, Kappen L, Schulze E (eds) Water and Life. Problems and Modern Approaches. Springer, Berlin, pp 492–505
Duponnois R, Ouahmane L, Kane A, Thioulouse J, Hafidi M, Boumezzough A, Prin Y, Baudoin E, Galiana A, Dreyfus B (2011) Nurse shrubs increased the early growth of Cupressus seedlings by enhancing belowground mutualism and soil microbial activity. Soil Biol Biochem 43:2160–2168
Eivazi F, Tabatabai MA (1988) Glucosidases and galactosidases in soils. Soil Biol Biochem 20:601–606
Emmett BA, Beier C, Estiarte M, Tietema A, Kristensen HL, Williams D, Peñuelas J, Schmidt I, Sowerby A (2004) The response of soil processes to climate change: results from manipulation studies of shrublands across an environmental gradient. Ecosystems 7:625–637
Enright NJ, Goldblum D (1999) Demography of a non-sprouting and resprouting Hakea species (Proteaceae) in fire-prone Eucalyptus woodlands of southeastern Australia in relation to stand age, drought and disease. Plant Ecol 144:71–82
Esteban-Parra MJ, Rodrigo FS, Castro-Diez Y (1998) Spatial and temporal patterns of precipitation in Spain for the period 1880–1992. Int J Clim 18:1557–1574
Estiarte M, Peñuelas J, Sardans J, Emmett BA, Sowerby A, Beier C, Schmidt IK, Tietema A, Van Meeteren MJM, Lang EK, Mathe P, De Angelis P, De Dato G (2008) Root-surface phosphatase activity in shrublands across a European gradient: effects of warming. J Env Biol 29:25–29
Everard K, Seabloom EW, Harpole WS, de Mazancourt C (2010) Plant use affects competition for nitrogen: why drought favors invasive species in California. Am Nat 175:85–97
Fife DN, Nambiar EKS, Saur E (2008) Retranslocation of foliar nutrients in evergreen tree species planted in a Mediterranean environment. Tree Physiol 28:187–196
Filella I, Peñuelas J (1999) Altitudinal differences in UV absorbance, UV reflectance and related morphological traits of Quercus ilex and Rhododedron ferrugineum in the Mediterranean region. Plant Ecol 145:157–162
Filella I, Peñuelas J (2003a) Indications of hydraulic lift by Pinus halepensis and its effects on the water relations of neighbour species. Biol Plantarum 47:209–214
Filella I, Peñuelas J (2003b) Partitioning of water and nitrogen in co-occuring Mediterranean woody shrub species of different evolutionary history. Oecologia 137:51–61
Fioretto A, Papa S, Curcio E, Sorrentino G, Fuggi A (2000) Enzyme dynamics on decomposing leaf litter of Cistus incanus and Myrtus communis in a Mediterranean ecosystem. Soil Biol Biochem 32:1847–1855
Fioretto A, Papa S, Sorrentino G, Fuggi A (2001) Decomposition of Cistus incatus leaf litter in a Mediterranean maquis ecosystem: mass loss, microbial enzyme activities and nutrient changes. Soil Biol Biochem 33:311–321
Fioretto A, Papa S, Fuggi A (2003) Litter-fall and litter decomposition in a low Mediterranean shrubland. Bio Fert Soils 39:37–44
Fioretto A, Papa S, Pellegrino A, Ferrigno A (2009) Microbial activities in soils of a Mediterranean ecosystem in different successional stages. Soil Biol Biochem 41:2061–2068
Foereid B, Bellarby J, Meier-Augenstein W, Kemp H (2010) Does light exposure make plant more degradable? Plant Soil 333:275–285
Fotelli MN, Radoglou KM, Constantinodou HIA (2000) Water stress responses of seedlings of four Mediterranean oak species. Tree Pysiol 20:1065–1075
Frei C, Schär C, Lüthi D, Davies HC (1998) Heavy precipitation processes in a warmer climate. Geophys Res Lett 25:1431–1434
Gabarrón-Galeote MA, Martínez-Murillo JF, Ruiz-Sinoga JD (2012) Relevant effects of vegetal cover and litter on the soil hydrological response of two contrasting Mediterranean hillslopes at the end of the dry season (south of Spain). Hydrol Proc 26:1729–1738
Gahrooee FR, van Breemen N, Buurman P (1998) Litter quality of Quercus plants growing in enhanced atmospheric CO2 in Mediterranean-type ecosystems. Impacts of Global Change on Tree Physiology and Forest Ecosystems. For Sci 52:233–237
Galiano L, Martínez-Vilalta J, Sabaté S, Lloret F (2012) Determinants of drought effects on crown conditions and their relationship with depletion of carbon reserves in a Mediterranean holm oak forest. Tree Physiol 32:478–489
Gallardo A (2003) Effect of tree canopy on the spatial distribution of soil nutrients in a Mediterranean Dehesa. Pedobiologia 47:117–125
Gallardo A, Merino J (1992) Nitrogen immobilization in leaf litter at two Mediterranean ecosystems of S.W. Spain. Biogeochemistry 15:213–228
Gallardo A, Merino J (1999) Control of leaf litter decomposition rate in a Mediterranean shrubland as indicated by N, P and lignin concentrations. Pedobiologia 43:64–72
Gallardo A, Rodríguez-Saucedo JJ, Covelo F, Fernández-Alès R (2000) Soil nitrogen heterogeneity in a Dehesa ecosystem. Plant Soil 222:71–82
Galle A, Florez-Sarasa I, El Aououad H, Flexas J (2011) The Mediterranean evergreen Quercus ilex and the semi-deciduous Cistus albidus differ in their leaf gas exchange regulation and acclimatrion to repeated drought and re-watering cycles. J Exp Bot 62:5207–5216
Gallego Fernández JB, García Mora MR, García Novo F (2004) Vegetation dynamics of Mediterranean shrublands in former cultural landscape at Grazalema Mountains, south Spain. Plant Ecol 172:83–94
Galmés J, Flexas J, Savé R, Medrano H (2007a) Water relations and stomatal characteristics of Mediterranean plants with different growth forms and leaf habits: responses to water stress and recovery. Plant Soil 290:139–155
Galmés J, Medrano H, Flexas J (2007b) Photosynthesis and photoinhibition in response to drought in a pubescent (var. minor) and a glabrous (var. palaui) variety of Digitalis minor. Env Exp Bot 60:105–111
Garcia C, Hernandez T, Costa F (1994) Microbial activity in soils under Mediterranean environmental conditions. Soil Biol Biochem 26:1185–1191
Garcia C, Hernandez T, Roldan A, Martin A (2002) Effect of plant cover decline on chemical and microbiological parameters under Mediterranean climate. Soil Biol Biochem 34:635–642
García C, Roldan A, Hernandez T (2005) Ability of different plant species to promote microbiological processes in semiarid soil. Geoderma 124:193–202
Garcia-Estringana P, Alonso-Blázquez N, Marques MJ, Bienes R, Alegre J (2010) Direct and indirect effects of Mediterranean vegetation on runoff and soil loss. Eur J Soil Sci 61:174–185
García-Palacios P, Maestre FT, Milla R (2012) Community-aggregated plant traits interact with soil nutrient heterogeneity to determine ecosystem functioning. Plant Soil. doi:10.1007/s11104-012-1349-6
García-Pausas J, Casals P, Romanyà J (2004) Litter decomposition and faunal activity in Mediterranean forest soils: effects of N content and the moss layer. Soil Biol Biochem 56:989–997
Gea-Izquierdo G, Allen-Díaz B, San Miguel A, Cañellas I (2010) How do trees affect spatio-temporal heterogeneity of nutrient cycling in Mediterranean annual grasslands? Ann For Sci 67:112
Gea-Izquierdo G, Fonti P, Cherubini P, Martín-Benito D, Chaar H, Cañellas I (2012) Xylem hydraulic adjustment and growth response of Quercus canariensis Wild. To climate variability. Tree Physiol 32:401–413
Goberna M, Sánchez J, Pasqual JA, García C (2006) Surface and subsurface organic carbon, microbial biomass and activity in a forest soil sequence. Soil Biol Biochem 38:2233–2243
Goberna M, Sánchez J, Pascual JA, García C (2007a) Pinus halepensis Mill. Plantations did not restore organic carbón, microbial biomass and activity levels in a semi-arid Mediterranean soil. Appl Soil Ecol 36:107–115
Goberna M, Pascual JA, García C, Sánchez J (2007b) Do plant clumps constitute microbial hotspots in semiarid Mediterranean patchy landscapes? Soil Biol Biochem 39:1047–1054
Goldstein AH, Hultman NE, Fracheboud JM, Bauer MR, Panek JA, Xu M, Qi Y, Guenther AB, Baugh W (2000) Effects of climate variability on the carbon dioxide, water, and sensible heat fluxes above a ponderosa pine plantation in the Sierra Nevada (CA). Agric For Metreol 101:113–129
Golodets C, Kigel J, Sternberg M (2011) Plant diversity partitioning in grazed Mediterranean grassland at multiple spatial and temporal scales. J Appl Ecol 48:1260–1268
Gómez-Aparicio L, Gómez JM, Zamora R, Boettinger JL (2005) Canopy vs. soil effects of shrub facilitating tree seedlings in Mediterranean montane ecosystems. J Veg Sci 16:191–198
Gómez-Rey MX, Madeira M, González-Prieto SJ, Coutinho J (2010) Soil C and N dynamics within a precipitation gradient in Mediterranean eucalypt plantations. Plant Soil 336:157–171
González E, Muller E, Comín FA, González-Sachis M (2010) Leaf nutrient concentration as an indicator of Populus and Tamarix response to flooding. Persp Plant Ecol Evol Syst 12:257–266
González-Hidalgo JC, Peña-Monné JL, de Luís M (2007) A review of daily soil erosion in Western Mediterranean areas. Catena 71:193–199
González-Pelayo O, Andreu V, Gimeno-García E, Campo J, Rubio JL (2010) Effects of fire and vegetation cover on hydrological characteristics of a Mediterranean shrubland soil. Hydrobiol Proc 24:1504–1513
Grant OM, Tronina L, Ramalho JC, Kurz Besson C, Lobo-do-Vale R, Santos Pereira J, Jones HG, Chaves MM (2010) The impact of drought on leaf physiology of Quercus suber L. trees: comparison of an extreme drought event with chronic rainfall reduction. J Exp Bot 61:4361–4371
Gratani L, Meneghini M, Pesoli P, Cresence MF (2003) Structural and functional plasticity of Q. ilex seedlings of different provinances in Italy. Trees 17:515–521
Grierson PF, Adams MA (2000) Plant species affect acid phosphatase, ergosterol and microbial P in a Jarrah (Eucapyptus marginata Donn ex Sm.) forest in south-western Australia. Soil Biol Biochem 32:1817–1827
Groom PK, Lamont BB (2010) Phosphorus accumulation in Proteaceae seeds: a synthesis. Plant Soil 334:61–71
Grubb PJ (1977) Control of forest growth and distribution on wet tropical mountains: with special reference to mineral nutrition. Ann Rev Ecol System 8:83–107
Grünzweig JM, Dumbur R (2012) Seed traits, seed-reserve utilization and offspring performance across pre-industrial to future CO2, concentrations in a Mediterranean community. Oikos 121:579–588
Grünzweig JM, Carmel V, Riov J, Sever N, McCreary DD, Flather CH (2008) Growth, resource storage, and adaptation to drought in California and eastern Mediterranean oak seedlings. Can J For Res 38:331–342
Guerrieri R, Siegwolf R, Saurer M, Ripullone F, Mencuccini M, Borghetti M (2010) Anthropogenic NOx emissions after the intrinsic water-use efficiency (WUEi) for Quercus cerris stands under Mediterranean climate conditions. Environ Pollut 158:2841–2847
Gulías J, Flexas J, Abadía A, Medrano H (2002) Photosynthetic responses to water deficit in six Mediterranean sclerophyll species: possible factors explaining the declining distribution of Rhamnus ludovici-salvatoris, an endemic Balearic species. Tree Physiol 22:687–697
Harrison S, Rice K, Maron J (2001) Habitat patchness promotes invasion by alien grasses on serpentine soil. Biol Conser 100:45–53
Henkin Z, Seligman NG, Kafkafi U, Noy-Meir I (1998) Effective growing days: a simple predictive model of response of herbaceous plant growth in a Mediterranean ecosystem to variation in rainfall and phosphorus availability. J Ecol 86:137–148
Hernandez I, Alegre L, Munne-Bosch S (2004) Drought-induced changes in flavonoids and other low molecular weight antioxidants in Cistus clussi grown under Mediterranean field conditions. Tree Physiol 24:1303–1311
Hernández-Santana V, David TS, Martínez-Fernández J (2008) Environmental and plant-based controls of water use in a Mediterranean oak stand. For Ecol Manag 255:3707–3715
Hernández-Santana V, Martínez-Vilalta J, Martínez-Fernández J, Williams M (2009) Evaluating the effect of drier and warmer conditions on water use by Quercus pyrenaica. For Ecol Manag 258:1719–1730
Herppich M, Herppich WB, von Willert DJ (2002) Leaf nitrogen content and photosynthetic activity in relation to soil nutrient availability in coastal and mountain fynbos plants (South Africa). Basic Appl Ecol 3:329–337
Hoff C, Rambal S (2003) An examination of the interaction of the interaction between climate, soil and leaf area index in a Quercus ilex ecosystem. Am For Sci 60:153–161
Howlett DS, Moreno G, Mosquera MR, Nair PKR, Nair VD (2011) Soil carbon storage as influenced by trwee cover in the Dehesa coprk oak silvopastorure of central-western Spain. J Env Monit 13:1897–1904
Hueso S, Hernández T, García C (2011) Resistance and resilence of the soil microbial biomass to severe drought in semiarid soils: The importance of organic amendments. Appl Soil Ecol 50:27–36
Incerti G, Bonanomi G, Giannino F, Rutigliano FA, Piermatteo D, Castaldi S, De Marco A, Fierro A, Fioretto A, Maggi O, Papa S, Persiani AM, Feoli E, Virzo de Santo A, Mazzoleni S (2011) Litter decomposition in Mediterranean ecosystems: modeling the controlling role of climatic conditions and litter quality. Appl Soil Ecol 49:148–157
Inclan R, Gimeno BS, Dizengremel P, Sanchez M (2005) Compensation processes of Aleppo pine (Pinus halepensis Mill.) to ozone exposure and drought stress. Environ Pollut 137:517–524
Inglima I, Alberti G, Bertolini T, Vaccari FP, Gioli B, Miglietta F, Cotrufo MF, Peressotti A (2009) Precipitation pulses enhance respiration of Mediterranean ecosystems: the balance between organic and inorganic components of increased soil CO2 efflux. Global Change Biol 15:1289–1301
IPCC (2007) Climate Change 2007: the physical science basis. Contribution of Working Group I. In: Solomon S, Qin D, Manning M, Chen Z, Marquis M, Averyt KB, Tignor M, Miller HL (eds) Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge, pp 849–940
Jarvis P, Rey A, Petsikos C, Wingate L, Rayment M, Pereira J, Banza J, David J, Miglietta F, Borghetti M, Manca G, Valentini R (2007) Drying and wetting of Mediterranean soils stimulates decomposition and carbon dioxide emission: the “Birch effect”. Tree Physiol 27:929–940
Joffre R, Rambal S, Ratte JP (1999) The dehesa system of southern Spain and Portugal as a natural ecosystem mimic. Agrofor Syst 45:57–79
Johnson-Maynard JL, Graham RC, Wu L, Shouse PJ (2002) Modification of soil structural and hydraulic properties after 50 years of imposed chaparral and pine vegetation. Geoderma 110:227–240
Jones MB, Brown JC, Raschi A, Miglietta F (1995) The effects on Arbutus unedo L. of long-term exposure to elevated CO2. Global Change Biol 1:295–302
Kandeler E, Gerber H (1988) Short-term assay of soil urease activity using colorimetric determination of ammonium. Biol Fertil Soils 6:68–72
Kazakou E, Vile D, Shipley B, Gallet C, Garnier E (2006) Co-variations in litter decomposition, leaf traits and plant growth in species from a Mediterranean old-field succession. Funct Ecol 20:21–30
Kazakou E, Violle C, Roumet C, Pintor C, Gimenez O, Garnier E (2009) Litter quality and decomposability of species from a Mediterranean succession depend on leaf traits but not on nitrogen supply. An Bot 104:1151–1161
Kooijman AM, Jongejans J, Sevink J (2005) Parent material effects on Mediterranean woodland ecosystems in NE Spain. Catena 59:55–68
Körner C, Miglietta F (1994) Long term effects of naturally elevated CO2 on mediterranean grassland and forest trees. Oecologia 99:343–351
Kruger FJ (1979) South African heathlands. In: Specht RL (ed) Ecosystems of the world. Heathlands and Related Shrublands. Elsevier, Amsterdam, pp 19–80
Kurz-Besson C, Otieno D, do Vale Lobo R, Siegwoll R, Schmidt M, Herd A, Nogueira C, David TS, Tenhunen J, Pereira JS, Chaves M (2006) Hydraulic lift in cork oak trees in a savannah-type Mediterranean ecosystem and its contribution to the local balance. Plant Soil 282:361–378
Kutiel H, Maheras P (1998) Variations in the temperature regime across the Mediterranean during the last century and their relationship with circulation indices. Theor Appl Climatol 61:39–53
Kutiel H, Maheras P, Guika S (1996) Circulation and extreme rainfall conditions in the Eastern Mediterranean during the last century. Int J Clim 16:72–92
Ladd JN, Butler JHA (1972) Short-term assays of soil proteolytic enzyme activities using protein and dipeptide derivatives as substrate. Soil Biol Biochem 4:19–30
Lamont BB, Wiens D (2003) Are seed set and speciation rates always low among species that resprout after fire, and why? Evolutionary Ecol 17:277–292
Larcher W (2000) Temperature stress and survival ability of Mediterranean sclerophyllous plants. Plant Biosyst 134:279–295
Lebourgeois F, Levy G, Becker M, Lefevre Y (1997) Effects of mineral nutrition and soil water conditions on radial growth of Corsican pine in western France. An Sci For 54:279–300
Lefi E, Medrano H, Cifre J (2004a) Water uptake dynamics, photosynthesis and water use efficiency in field-grown Medicago arborea under prolonged Mediterranean drought conditions. Ann Appl Biol 144:299–307
Lefi E, Medrano H, Cifre J (2004b) Water dynamics, photosynthesis and water use efficiency in field-grown Medicago arborea and Medicago citrine under prolonged Mediterranean drought conditions. Am Appl Biol 144:299–307
Li X, Sarah P (2003) Enzyme activities along a climate change transect in the Judean Desert. Catena 53:349–363
Li XY, Contreras S, Solé-Benet A, Cantón Y, Domingo F, Lázaro R, Lin H, Van Wesemael B, Puigdefábregas J (2011) Controls of infiltration-runoff processes in Mediterranean karst rangelands in SE Spain. Catena 86:98–109
Limousin JM, Misson L, Lavoir LM, Martin NK, Rambal S (2010) Do photosynthetic limitations of evergreen Quercus ilex leaves change with long-term increased drought severity? Plant Cell Env 33:863–875
Llorens L, Peñuelas J, Estiarte M (2003a) Ecophysiological responses of two Mediterranean shrubs, Erica multiflora and Globularia alypum, to experimentally drier and warmer conditions. Physiol Plantarum 119:231–243
Llorens L, Peñuelas J, Filella I (2003b) Diurnal and seasonal variations in the photosynthetic performance and water relations of two co-occuring Mediterranean shrubs, Erica multiflora and Globularia alypum. Physiol Plantarum 118:84–95
Lloret F, Casanovas C, Peñuelas J (1999) Seedling survival of Mediterranean shrubland species in relation to root:shoot ratio, seed size and water and nitrogen use. Funct Ecol 13:210–216
Lloret F, Peñuelas J, Ogaya R (2004) Establishment of co-existing Mediterranean tree species under a varying soil moisture regime. J Veg Sci 15:237–244
Lloret F, Estevan H, Vayreda J, Terradas J (2005) Fire regenerative syndromes of forest woody species across fire and climatis gradients. Oecologia 146:461–468
Llusia J, Peñuelas J, Alessio GA, Estiarte M (2006) Seasonal contrasting changes of foliar concentrations of terpenes and other volatile organic compound in four dominant species of a Mediterranean shrubland submitted to a field experimental drought and warming. Physiol Plantarum 127:632–649
Llusia J, Peñuelas J, Alessio G, Estiarte M (2008) Contrasting species-specific, compound-specific, seasonal, and interannual responses of foliar isoprenoid emissions to experimental drought in a Mediterranean shrubland. Int J Plant Sci 169:637–645
Llusia J, Peñuelas J, Ogaya R, Alessio G (2010) Annual and seasonal changes in foliar terpene content and emission rates in Cistus albidus L. submitted to soil drought in Prades forest (Catalonia, NE Spain). Acta Physiol Plantarum 32:387–394
Lo Gullo MA, Salleo S, Rosso R, Trifilò F (2003) Drought resistance of 2-year-old saplings of Mediterranean forest trees in the field: relations between water relations, hydraulics and productivity. Plant Soil 250:250–272
Lopez B, Sabate S, Ruiz I, Gracia C (1998) Effects of elevated CO2 and decreased water availability on holm-oak seedlings in controlled environmental chambers. Impacts of global change on tree physiology and forest ecosystems. For Sci 52:125–133
Loreto F, Fischbach RJ, Schnitzler JP, Ciccioli P, Brancaleoni E, Calfapietra C, Seufert G (2001) Monoterpene emission and monoterpene synthase activities in the Mediterranean evergreen oak Quercus ilex L. grown at elevated CO2 concentrations. Global Change Biol 7:709–717
Lucas-Borja ME, Bastida F, Nicolás C, Moreno JL, del Cierro A, Andrés M (2010) Influence of forest cover and herbaceous vegetation on the microbiological and biochemical properties of soil under Mediterranean humid climate. Eur J Soil Biol 46:273–279
Lucas-Borja ME, Candel D, Jindo K, Moreno JL, Andrés M, Bastida F (2012) Soil microbial community structure and activity in monospecific and mixed forest stands, under Mediterranean humid conditions. Plant Soil. In press
Maestre FT, Cortina J (2002) Spatial patterns of surface soil properties and vegetation in a Mediterranean semi-arid steppe. Plant Soil 241:279–291
Maestre FT, Cortina J (2003) Small-scale spatial variation in soil CO2 effux in a Mediterranean semiarid steppe. Appl Soil Ecol 23:199–209
Maestre FT, Puche MD, Guerrero C, Escudero A (2011) Shrub encroachment does not reduce the activity of some soil enzymes in Mediterranean semiarid grassland. Soil Biol Biochem 43:1746–1749
Maestre FT, Castillo-Monroy AP, Bowker MA, Ochoa-Hueso R (2012) Species richness effects on ecosystem multifunctionality depend on eveness, composition and spatial pattern. J Ecol 100:317–330
Maisto G, De Marco A, Meola A, Sessa L, De Santo AV (2011) Nutrient dynamics in litter mixtures four Mediterranean maquis species decomposing in situ. Soil Biol Biochem 43:520–530
Mamolos AP, Veresoglou DS, Noitaskis V, Gerakis A (2001) Differential drought tolerance of five coexisting plant species in Mediterranean lowland grasslands. J Arid Env 49:329–341
Marek MV, Sprtova M, De Angelis P, Scarascia-Mugnozza G (2001) Spatial distribution of photosynthetic response to long-term influence of elevated CO2 in a mediterranean macchia mini-ecosystem. Plant Sci 160:1125–1136
Maroco JP, Breia E, Faria T, Pereira JS, Chaves MM (2002) Effects of long-term exposure to elevated CO2 and N fertilization on the development of photosynthetic capacity and biomass accumulation in Quercus suber L. Plant Cell Env 25:105–113
Martínez F, Merino O, Martin A, García Martín D, Merino J (1998) Beloground structure and production in a Mediterranean sand dune shrub community. Plant Soil 201:209–216
Martínez-García LB, Armas C, Miranda JD, Padilla FM, Pugnaire FI (2011) Shrubs influence arbuscular mycorrgizal fungi communities in a semi-arid environment. Soil Biol Biochem 43:682–689
Martínez-García LB, Miranda JD, Pugnaire FI (2012) Impacts of changing rainfall patterns on mycorrhizal status of a shrub from arid environments. Eu J Soil Biol 50:64–67
Martínez-Vilalta J, Piñol J (2002) Drought-induced mortality and hydraulic architecture in pine populations of the NE Iberian Peninsula. For Ecol Manag 161:247–256
Martínez-Vilalta J, Mangirón M, Ogaya R, Sauret M, Serrano L, Peñuelas J, Piñol J (2003) Sap flow of three co-occuring Mediterranean woody species under varying atmospheric and soil water conditions. Tree Physiol 23:747–758
Martí-Roure M, Casals P, Romanyà J (2011) Temporal changes in soil organic C under Mediterranean shrublands and grasslands: impact of fire and drought. Plant Soil 338:289–300
Maseyk K, Grünzweig JM, Rotenberg E, Yakir D (2008) Respiration acclimation contributes to high carbon-use efficiency in a seasonally dry pine forest. Global Change Biol 14:1553–1567
Maseyk K, Hemming D, Angert A, Leavitt SW, Yakir D (2011) Increase in water-use efficiency and underlying processes in pine forest across a precipitation gradient in the dry Mediterranean region over the past 30 years. Oecologia 167:573–585
Matías L, Castro J, Zamora R (2011) Soil-nutrient availability under a global-change scenario in a Mediterranean mountain ecosystem. Global Chane Biol 17:1646–1657
Mattia C, Bischetti GB, Gentile F (2005) Biotechnical characteristics of root systems of typical Mediterranean species. Plant Soil 278:23–32
Mayor X, Rodà F (1994) Effects of irrigation and fertilization on stem diameter growth in a Mediterranean holm oak forest. For Ecol Manage 68:119–126
Mayor AG, Bautista S, Bellot J (2009) Factors and interactions controling infiltration, runoff, and soil loss at the microscale in a patchy Mediterranean semiarid landscape. Earth Surf Process Landforms 34:1702–1711
Mayor AG, Bautista S (2012) Multi-scale evaluation of soil functional indicators for the assessment of water and soil retention in Mediterranean semiarid landscapes. Ecol Indic 20:332–336
McMaster GS, Jow WM, Kummerow J (1982) Response of Adenostoma fasciculatun and Ceanothus gregii chaparral to nutrient additions. J Ecol 70:745–756
Mediavilla S, Escudero A (2004) Stomatal responses to drought of mature trees and seedlings of two co-occuring Mediterranean oaks. For Ecol Manag 187:281–294
Medina A, Azcón R (2010) Effectiveness of the application of arbuscular mycorrhiza fungi and organic amendments to improve soil quality and plant performance under stress conditions. J Soil Plant Nutr 10:354–372
Medrano H, Flexas J, Galmés J (2009) Variability in water use efficiency at the leaf level among Mediterranean plants with different growth forms. Plant Soil 317:17–29
Mereu S, Salvatori E, Fusaro L, Gerosa G, Muys B, Manes F (2009) An integrated approach shows different use of water resources from Mediterranean maquis species in a coastal dune ecosystem. Biogeosciences 6:2599–2610
Merino-Martin L, Moreno-de las Heras M, Pérez-Domingo S, Espigarea T, Nicolau JM (2012) Hydrological heterogeneity in Mediterranean reclaimed slopes: runoff and sediment yield at the patch and slope scales along a gradient of overland flow. Hydrol Earth Syst Sci 16:1305–1320
Midgley GF, Stock WD, Juritz JM (1995) Effects of elevated CO2 on Cape fynbos species adapted to soils of different nutrient status:nutrient and CO2-responsiveness. J Biogeography 22:185–191
Milla R, Castro-Díaz P, Maestro-Martínez M, Montserrat-Martí G (2005) Relationships between phenology and the remobilization of nitrogen, phosphorus and potassium in branches of eight Mediterranean evergreens. New Phytol 168:167–178
Millan MM, Estrella MJ, Sanz MJ, Mantilla E, Martin M, Pastor F, Salvador R, Vallejo R, Alonso L, Gangoiti G, Ilardia JL, Navazo M, Albizuri A, Artinano B, Ciccioli P, Kallos G, Carvalho RA, Andres D, Hoff A, Werhahn J, Seufert G, Versino B (2005) Climatic feedbacks and desertification: The Mediterranean model. J Clim 18:684–701
Miller PC (1981) Nutrients and water relations in Mediterranean-type ecosystems. Gen Tech Rep PSW-58. Berkeley USA
Misson L, Limousin JM, Rodriguez R, Letts MG (2010) Leaf physiological responses to extreme droughts in Mediterranean Quercus ilex forest. Plant Cell Environ 33:1898–1910
Monokrousos N, Papatheodorou EM, Diamantopoulos JD, Stamou GP (2004) Temporal and spatial variability of soil chemical and biologicval variables in a Mediterranean shrubland. For Ecol Manag 202:83–91
Mooney H (1989) Chaparral physiological ecology-paradigms reexamined. In: Keely SC (ed) The California chaparral. Paradigms reexamined no 34, science series. Natural History Museum of Los Angeles County, Los Angeles
Mooney HA, Dunn EL (1970) Convergent evolution of Mediterranean-climate evergreen sclerophyll shrubs. Evolution 24:292–303
Moreno G, Obrador JJ, Cubera E, Dupraz C (2005) Fine root distribution in Dehesas of Central-western Spain. Plant Soil 277:153–162
Moreno-de las Heras M, Espigares T, Merino-Martín L, Nicolau JM (2011) Water-related ecological impacts of rill erosion processes in Mediterranean-dry reclaimed slopes. Catena 84:114–124
Morgan JA, Pataki DE, Körner C, Clark H, Del Gross SJ, Grünzweig JM, Knapp AK, Mosier AR, Newton PCD, Niklaus AK, Nippert JB, Nowak RS, Parton WJ, Polley HW, Shaw MR (2004) Water relations in grassland and desert ecosystems exposed to elevated atmospheric CO2. Oecologia 140:11–25
Moriondo M, Good P, Durao R, Bindi M, Giannakopoulos C, Corte-Real J (2006) Potential impact of climate change on fire risk in the Mediterranean area. Clim Res 31:85–95
Moro MJ, Domingo F (2000) Litter decomposition in four woody species in a Mediterranean climate: weight loss, N and P dynamics. An Bot 86:1065–1071
Morón-Ríos A, Rodríguez MA, Pérez-Camacho L, Rebollo S (2010) Effects of seasonal grazing and precipitation regime on the soil macroinvertebrates of a Mediterranean old-field. Eur J Soil Biol 46:91–96
Moscatelli MC, Fonck M, De Angelis P, Larbi H, Macuz A, Rambelli A, Grego S (2001) Mediterranean natural forest living at elevated carbón dioxide: soil biological properties and plant biomass growth. Soil Use Manag 17:195–202
Mouillot F, Rambal S, Joffre R (2002) Simulating climate change impacts on fire frequency and vegetation dynamics in a Mediterranean-type ecosystem. Global Change Biol 8:423–437
Munné-Bosch S, Alegre L (2000) Changes in carotenoids, tocopherols and diterpenes during drought and recovery, and the biological significance of chlorophyll loss in Rosmarinus officinalis plants. Planta 210:925–931
Munné-Bosch S, Alegre L (2004) Die and let live: leaf senescence contributes to plant survival under drought stress. Funct Plant Biol 31:203–216
Munné-Bosch S, Lalueza P (2007) Age-related changes in oxidative stress markers and abscisic acid levels in a drought-tolerant shrub, Cistus clusii grown under Mediterranean field conditions. Planta 225:1039–1049
Munné-Bosch S, Peñuelas J (2003) Photo- and antioxidative protection, and a role for salicylic acid during drought and recovery in field-grown Phillyrea angustifolia plants. Planta 217:758–766
Munné-Bosch S, Peñuelas J (2004) Drought-induced oxidative stress in strawberry tree (Arbutus unedo L.) growing in Mediterranean field conditions. Plant Sci 166:1105–1110
Muñoz C, Zagal E, Ovalle C (2007) Influence of trees on soil organic matter in Mediterranean agroforestry systems: an example from the “Espinal” of central Chile. Eur J Soil Sci 58:728–735
Nardini A, Salleo S, Lo Gullo MA, Pitt F (2000) Different responses to drought and freeze stress of Quecus ilex L. growing along a latitudinal gradient. Plant Ecol 148:139–147
Navarro-García F, Casermeiro MA, Schimel JP (2012) When structure means conservation: Effect of aggregate structure in controlling microbial responses to rewetting events. Soil Biol Biochem 44:1–8
Navas ML, Guillerm JL, Fabreguettes J, Roy J (1995) The influence of elevated CO2 on community structure, biomass and carbon balance of mediterranean old-field microcosms. Global Change Biol 1:325–335
Navas ML, Sonie L, Richarte J, Roy J (1997) The influence of elevated CO2 on species phenology, growth and reproduction in a Mediterranean old-field community. Global Change Biol 3:523–530
Navas ML, Roumet C, Bellmann A, Laurent G, Garnier E (2010) Suites of plant traits in species from different stages of a Mediterranean secondary succession. Plant Biol 12:183–196
Nguyen BT, Marschner P (2005) Effect of drying and rewetting on phosphorus transformations in red brown soils with different soil organic matter content. Soil Biol Biochem 37:1573–1576
Nijs I, Roy J, Salager JL, Fabreguettes J (2000) Elevated CO2 alters carbon fluxes in early successional Mediterranean ecosystems. Global Change Biol 6:981–994
Nogues I, Peñuelas J, Llusia J, Estiarte M, Munné-Bosch S, Sardans J, Loreto F (2012) Physiological and antioxidant responses of Erica multiflora to drought and warming through different seasons. Plant Ecol 213:649–661
Nuñez-Oliveira E, Martínez-Zabaigar J, Escudero-Garcia JC (1993) Litterfall and nutrient flux in Cistus-ladanifer L. shrubland in SW Spain. Acta Oecol 14:361–369
Ochoa-Hueso R, Hernandez RR, Pueyo JJ, Manrique E (2011) Spatial distribution and physiology of biological soil crust from semi-arid central Spain are related to soil chemistry and shrub cover. Soil Biol Biochem 43:1894–1901
Ogaya R, Peñuelas J (2003) Comparative seasonal gas exhange and chlorophyll fluorescence of two dominant woody species in a holm oak Forest. Flora 198:132–141
Ogaya R, Peñuelas J (2006) Contrasting foliar responses to drought in Quercus ilex and Phillyrea latifolia. Biol Plantarum 50:373–382
Ogaya R, Peñuelas J (2007a) Tree growth, mortality, and above-ground biomass accumulation in a holm oak forest under a five-year experimental field drought. Plant Ecol 189:291–299
Ogaya R, Peñuelas J (2007b) Species-specific drought effects on flower and fruit production in a Mediterranean holm oak forest. Forestry 80:351–357
Ogaya R, Peñuelas J (2008) Changes in leaf d13C and d15N for three Mediterranean tree species in relation to soil water availability. Acta Oecol 34:331–338
Ogaya R, Peñuelas J, Martínez-Vilalta J, Mangirón M (2003) Effect of drought on diameter increment of Quercus ilex, Phillyrea latifolia, and Arbutus unedo in a holm oak forest of NE Spain. For Ecol Manag 180:175.184
Oliveira G, Peñuelas J (2002) Comparative protective strategies of Cistus albidus and Quercus ilex facing photoinhibitory winter conditions. Env Exp Bot 47:281–289
Oliveira G, Correira O, Martins-Louçäo MA, Catarino FMI (1994) Phenological and growth patterns of the Mediterranean oak Quercus suber. Trees 9:41–49
Oliveras I, Martínez-Vilalta J, Jiménez-Ortiz T, Lledó MJ, Escarré A, Piñol J (2003) Hidrailic properties of Pinus halepensis, Pinus pinea and Tetrachinus articulate in a dune ecosystem of eastern Spain. Plant Ecol 169:131–141
Orshan G (1983) Approaches to the definition of Mediterranean growth forms. Ecol Studies 43:86–100
Osborne CP, Mitchell PL, Sheehy JE, Woodward FI (2000) Modelling the recent historial impacts of atmospheric CO2 and climate change on Mediterranean vegetation. Global Change Biol 6:445–458
Otieno DO, Kurz-Besson C, Liu J, Schmidt MWT, do Vale-Lobo R, David TS, Siegwoll R, Pereira JS, Tenhunen JD (2006) Seasonal variations in soil and plant water status in a Quercus suber L. stand: roots as determinants of tree productivity and survival in the Mediterranean-type ecosystem. Plant Soil 283:119–135
Ozturk M, Dogan Y, Sakcali MS, Doulis A, Karem F (2010) Ecophysiological responses of some maquis (Ceratonia siliqua L., Olea Oleaster Hoffm. & Link, Pistacea lentiscus and Quercus coccifera L.) plant species to drought in the east Mediterranean ecosystem. J Env Biol 31:233–245
Padilla FM, Pugnaire FI (2007) Rooting depth and soil moisture control Mediterranean woody seedling survival during drought. Funct Ecol 21:489–495
Padilla FM, de Dios MJ, Pugnaire FI (2007) Early root growth plasticity in seedlings of three Mediterranean woody species. Plant Soil 296:103–113
Pegoraro E, Rey A, Barron-Gafford G, Monson R, Malhi Y, Murthy R (2005) The interacting effects of elevated atmospheric CO2 concentration, drought and leaf-to-air vapour pressure deficit on ecosystem isoprene fluxes. Oecologia 146:120–129
Pegoraro E, Potosnak MJ, Monson RK, Rey A, Barron-Gafford G, Osmond CB (2007) The effect of elevated CO2, soil and atmospheric water deficit and seasonal phenology on leaf and ecosystem isoprene emission. Funct Plant Biol 34:774–784
Peguero-Pina JJ, Morales F, Flexas J, Gil-Pelegrín E, Moya I (2008) Photochemistry, remotely sensed physiological reflectance index and de-epoxidation state of the xanthophyll cycle in Quercus coccifera under intense drought. Oecologia 156:1–11
Peltier A, Ponge JF, Jordana R, Ariño A (2001) Humus forms in Mediterranean scrubland with Aleppo pine. Soil Sci Soc Am J 65:884–896
Peña-Rojas K, Aranda X, Fleck I (2004) Stomatal limitation to CO2 assimilation and down-regulation of photosynthesis in Quercus ilex resprouts in response to slowly imposed drought. Tree Physiol 24:813–822
Peñuelas J, Azcón-Bieto J (1992) Changes in lead Δ13C of herbarium plant species during the last 3 centuries of CO2 increase. Plant Cell Environ 15:485–489
Peñuelas J, Boada M (2003) A global change-induced biome shift in the Montseny mountains (NE Spain). Global Change Biol 9:131–140
Peñuelas J, Estiarte M (1997) Trends in plant carbon concentration and plant demand for N throughout this century. Oecologia 109:69–73
Peñuelas J, Estiarte M (1998) Can elevated CO2 affect secondary metabolism and ecosystem functioning? Trends Ecol Evol 13:20–24
Peñuelas J, Filella I (2001) Phenology: responses to a warming world. Science 294:793–795
Peñuelas J, Staudt M (2010) BVOCs and global change. Trends Plant Sci 15:133–144
Peñuelas J, Filella I, Lloret F, Piñol J, Siscart D (2000) Effects of a severe drought on water and nitrogen use by Quercus ilex and Phillyrea latifolia. Biol Plantarum 43:47–53
Peñuelas J, Filella I, Tognetti R (2001) Leaf mineral concentrations of Erica arborea, Junniperus communis and Myrtus communis growing in the proximity of a natural CO2 spring. Global Change Biol 7:291–301
Peñuelas J, Filella I, Comas P (2002a) Changed plant and animal life cycles from 1952 to 2000 in the Mediterranean region. Glob Change Biol 8:531–544
Peñuelas J, Castells E, Joffre R, Tognetti R (2002b) Carbon-based secondary and structural compounds in Mediterranean shrubs growing near a natural CO2 spring. Global Change Biol 8:281–288
Peñuelas J, Filella I, Zhang X, Llorens L, Ogaya R, Lloret F, Comas P, Estiarte M, Terradas J (2004) Complex spatiotemporal phonological shifts as a response to rainfall changes. New Phytol 161:837–846
Peñuelas J, Filella I, Sabate S, Gracia C (2005) Natural systems: terrestrial ecosystems. In: Llebot JE (ed) Report on Climate Change in Catalonia. Institut d’Estudis Catalans, Barcelona, pp 517–553
Peñuelas J, Hunt JM, Ogaya R, Jump AS (2008a) Twentieth century changes of tree-ring δ13C at the southern range-edge of Fagus sylvatica: increasing water-use efficiency does not avoid the growth decline induced by warming at low latitudes. Global Change Biol 14:1076–1088
Peñuelas J, Sardans J, Ogaya R, Estiarte M (2008b) Nutrient stoichiometry relations and biogeochemical niche in coexisting plant species: effects of simulated climate change. Polish J Ecol 56:613–622
Peñuelas J, Terradas J, Lloret F (2011) Solving the conundrum of plant species coexistence: water in space and time matters most. New Phytol 18:3–6
Pérez-Ramos IM, Ourcival JM, Limousin JM, Rambal S (2010) Mast seedling under increasing drought: results from a long-term data set and from a rainfall exclusión experiment. Ecology 91:3057–3068
Pinay G, Barbera P, Carreras-Palou A, Fromin N, Sonié L, Couteaux MM, Roy J, Philippot L, Lensi R (2007) Impact at atmospheric CO2 and plant life forms on soil microbial activities. Soil Biol Biochem 39:33–42
Pinho P, Theobald MR, Días T, Tang YS, Cruz C, Martins-Louçao MA, Máguas C, Sutton M, Branquinho C (2012) Critical loads of nitrogen deposition and critical levels of atmospheric ammonia for semi-natural Mediterranean evergreen woodlands. Biogeosciences 9:1205–1215
Piñol J, Terradas J, Lloret F (1998) Climate warming, wildfire hazard, and wildfire occurrence in coastal eastern Spain. Clim Change 38:347–357
Pinzari F, Trinchera A, Benedetti A, Sequi P (1999) Use of biochemical indices in the Mediterranean environment: comparison among soils under different forest vegetation. J Microbiol Met 36:21–28
Placella SA, Brodie EL, Firestone MK (2012) Rainfall-induced carbon dioxide pulses result from se quential resuscitation of phylogenetically clustered microbial groups. Proc Natl Acad Sci U S A 109:10931–10936
Plaza J, Nuñez L, Pujadas M, Pèrez-Pastor R, Bermejo V, García-Alonso S, Elvira S (2005) Field monoterpene emission of Mediterranean oak (Quercus ilex) in the central Iberian Peninsula measured by enclosure and micrometeorological techniques: Observation of drought stress effect. J Geophys Res 110:D01105
Ponge F (2003) Humus forms in terrestrial ecosystems: a framework to biodiversity. Soil Biol Biochem 35:935–945
Prieto P, Peñuelas J, Lloret F, Llorens L, Estiarte M (2009a) Experimental drought and warming decrease diversity and slow down post-fire succession in a Mediterranean shrubland. Ecography 32:623–636
Prieto P, Peñuelas J, Llusia J, Asensio D, Estiarte M (2009b) Effects of experimental warming and drought on biomass accumulation in a Mediterranean shrubland. Plant Ecol 205:179–191
Prieto I, Kikvidze Z, Pugnaire FI (2010a) Hydraulic lift: soil processes and transpiration in the Mediterranean leguminous shrub Retama sphaerocarpa (L.) Boiss. Plant Soil 329:447–456
Prieto I, Martínez-Tilleria K, Martínez-Manchego L, Montecinos S, Pugnaire FI, Squeo FA (2010b) Hydraulic lift through transpiration suppression in shrubs from two arid ecosystems: patterns and control mechanisms. Oecologia 163:855–865
Prober SM, Wiehl G (2011) Resource heterogeneity and persistance of exotic annuals in long-ungrazed Mediterranean-climate woodlands. Biol Invasions 13:2009–2022
Pugnaire FI, Chapin FS (1993) Controls over nutrient resorption from leaves of evergreen Mediterranean species. Ecology 74:124–129
Qi Y, Xu M (2001) Separating the effects of moisture and temperature on soil CO2 efflux in a coniferous forest in the Sierra Nevada. Plant Soil 237:15–23
Quero JL, Sterck FJ, Martínez-Vilalta J, Villar R (2011) Water-use strategies of six co-existing Mediterranean woody species during a summer drought. Oecologia 166:45–57
Quideau SA, Graham RC, Chadwick OA, Wood HB (1998) Organic carbon sequestration under chaparral and pine after four decades of soil development. Geoderma 83:227–242
Quilchano C, Marañon T (2002) Dehydrogenase activity in Mediterranean forest soils. Biol Fert Soils 35:102–107
Rey A, Jarvis PG (1997) Growth response of young birch trees (Betula pendula Roth.) after four and a half years of CO2 exposure. Ann Bot 80:809–816
Rey A, Pegoraro E, Tedeschi V, De Parri I, Jarvis PG, Valentini R (2002) Annual variation in soil respiration and its components in a coppice oak forest in central Italy. Globa Change Biol 8:851–866
Rey A, Petsikos C, Jarvis PG, Grace J (2005) Effect of temperature and moisture on rates of carbon mineralization in a Mediterranean oak forest under controlled and field conditions. Eur J Soil Sci 56:589–599
Rey A, Pegoraro E, Oyonarte C, Were A, Escribano P, Raimundo J (2011) Impact of land degradation on soil respiration in a steppe (Stipa tenacissima L.) semi-arid ecosystem in the SE of Spain. Soil Biol Biochem 43:393–403
Riera P, Peñuelas J, Farreras V, Estiarte M (2007) Valuation of climate-change effects on Mediterranean shrublands. Ecol Appl 17:91–100
Rillig MC, Allen MF, Klironomos JN, Field CB (1998a) Arbuscular mycorrhizal percent root infection and infection intensity Bromus hordaceous grown in elevated atmospheric CO2. Mycologia 90:199–205
Rillig MC, Allen MF, Klironomos JN, Chiariello NR, Field CB (1998b) Plant species-specific changes in root-inhabiting fungi in a California annual grassland: responses to elevated CO2 and nutrients. Oecologia 113:252–259
Rinnan R, Michelsen A, Baath E, Jonasson S (2007) Mineralization and carbon turnover in subartic heath soil as affected by warming and additional litter. Soil Biol Biochem 39:3014–3023
Ripullone F, Borghetti M, Raddi S, Vicinelli E, Baraldi R, Guerrieri MR, Nolè A, Magnani F (2009) Physiological and structural changes in response to altered precipitation regimes in a Mediterranean macchia ecosystem. Trees 23:823–834
Rivas-Ubach A, Sardans J, Pérez-Trujillo M, Estiarte M, Peñuelas J (2012) Strong relationship between elemental stoichiometry and metabolome in plants. Proc Natl Acad Sci U S A 109:4181–4186
Rodà F, Retana J, Gracia CA, Bellot J (1999a) Ecology ofMediterranean evergreen oak forest. Springer, Berlin
Rodà F, Mayor X, Sabaté S, Diego V (1999b) Water and nutrient limitations to primary production. In: Rodà F, Retana J, Gracia C, Bellot J (eds) Ecology of Mediterranean evergreen oak forests. Springer, Berlin, pp 183–194
Rolo V, López-Díaz ML, Moreno G (2012) Shrubs affect soil nutrients availability with contrasting consequences for pasture understory and tree overstory production and nutrient status in Mediterranean grazed open woodlands. Nutr Cycl Agroecosyst 93:89–112
Rodríguez-Pleguezuelo CR, Durán Zuazo VH, Fernández JLM, Martín Peinado FJ, Franco Tarifa D (2009) Litter decomposition and nitrogen release in a sloping Mediterranean subtropical agroecosystem on the coast of Granada (SE, Spain): effects of floristic and topographic alteration on the slope. Agric Ecosys Environ 134:79–88
Romero R, Guijarro JA, Ramis C, Alonso S (1998) A 30-year (1964–1993) daily data base for the Spanish Mediterranean regions: first exploratory study. Int J Climatol 18:541–560
Roumet C, Laurent G, Roy J (1999) Leaf structure and chemical composition as affected by elevated CO2: genotypic responses of two perennial grasses. New Phytol 143:73–81
Roumet C, Garnier E, Suzor H, Salager JL, Roy J (2000) Short and long-term responses of whole-plant gas exchange to elevated CO2 in four herbaceous species. Env Exp Bot 43:155–169
Rovira P, Vallejo R (2003) Physical protection and biochemical quality of organic matter in Mediterranean calcareous forest soils: a density fractionation approach. Soil Biol Biochem 35:245–261
Rovira P, Jorba M, Romanyà J (2010) Active and passive organic matter fractions in Mediterranean forest soils. Biol Fertil Soils 46:355–360
Ruiz-Sinoga JD, Martínez-Murillo JF (2009) Effects of soil surface components on soil hydrological behavior in a dry Mediterranean environment (Southern Spain). Geomorphology 108:234–245
Ruiz-Sinoga JD, Diaz AR, Bueno EF, Martínez-Murillo JF (2010) The role of soil surface conditions in regulating runoff and erosion processes on a metamosphic hillslope (Southern Spain). Soil surface conditions, runoff and erosion in Southern Spain. Catena 80:131–139
Ruiz-Sinoga JD, Gabarón Galeote MA, Martínez Murillo JF, Garcia Marín R (2011) Vegetation strategies for soil consumption along a pluviometric gradient in southern Spain. Catena 84:12–20
Ruiz-Sinoga JD, Pariente S, Romero Díaz A, Martínez Murillo JF (2012) Variability of relationships between soil organic carbon and some soil properties in Mediterranean rangelands under different climatic conditions (South of Spain). Catena. In press
Rundel PW (1982) Water balance in the Mediterranean. In Symposium on the Environmental Consequences of Fire and Fuel Management in Mediterranean Ecosystems. (Eds) H A Mooney and C E Conrad. pp. 95–106. USDA Forest Service Gen. Tech. Rep. PSW 3
Rutigliano FA, Castaldi S, D’Ascoli R, Papa S, Carfora A, Marzaioli R, Fioretto A (2009) Soil activities related to nitrogen cycle under three plant cover types in Mediterranean environment. Appl Soil Ecol 43:40–46
Sabaté S, Calvet S, Gràcia CA (1992) Preliminary results of a fertilization-irrigation experiment in a Quercus ilex L. ecosystems at Prades mountains in Tarragona (NE Spain) in relation to leaves and twigs. Vegetatio 99–100:283–287
Sadaka N, Ponge JF (2003) Climatic effects on soil trophic networks and the resulting humus profiles in holm oak (Quercus rotundifolia) forests in the high Atlas of Morocco as revealed by correspondence analysis. Eur J Soil Sci 54:767–777
Salvati L, Perini L, Sabbi A, Bajocco S (2012) Climate aridity and land use changes: a regional-scale analysis. Geogr Res 50:193–203
Sanz-Pérez V, Castro-Díaz P, Millard P (2009) Effects of drought and shade on nitrogen cycling in the leaves and canopy of Mediterranean Quercus seedlings. Plant Soil 316:45–56
Sardans J, Peñuelas P (2004) Increasing drought decreases phosphorus availability in an evergreen Mediterranean forest. Plant Soil 267:367–377
Sardans J, Peñuelas J (2005) Drought decreases soil enzyme actiyity in a Mediterranean holm oak forest. Soil Biol Biochem 37:455–461
Sardans J, Peñuelas J (2007a) Drought changes phosphorus and potassium accumulation patterns in an evergreen Mediterranean forest. Funct Ecol 21:191–201
Sardans J, Peñuelas J (2007b) Drought changes the dynamics of trace elements accumulation in a Mediterranean Quercus ilex forest. Environ Pollut 147:567–583
Sardans J, Peñuelas J (2008) Drought changes nutrient sources, content and stoichiometry in the bryophyte Hypnum cupressiforme Hedw. growing in a Mediterranean forest. J Bryol 30:59–65
Sardans J, Peñuelas J (2010) Soil enzyme activity in a Mediterranean forest after six years of drought. Soil Sci Soc Am J 74:838–851
Sardans J, Peñuelas J (2012) Tree nutrient allocation to leaves and wood changes with climate, forest type and growth. Global Ecol Biogeogr. In press
Sardans J, Rodà F, Peñuelas J (2004) Phosphorus limitation and competitive capacities of Pinus halepensis and Quercus ilex subsp. rotundifolia on different soils. Plant Ecol 174:305–317
Sardans J, Peñuelas J, Rodà F (2005a) Changes in nutrient use efficiency, status and retranslocation in young post-fire regeneration Pinus halepensis in response to sudden N and P input, irrigation and removal of competing vegetation. Trees 19:233–250
Sardans J, Rodà F, Peñuelas J (2005b) Effects of water and a nutrient pulse supply on Rosmarinus officinalis growth, nutrient content and flowering in the field. Env Exp Bot 53:1–11
Sardans J, Peñuelas J, Rodà F (2006a) Plasticity of leaf morphological traits, leaf nutrient content, and water capture in the Mediterranean evergreen oak Quercus ilex subsp. ballota in response to fertilization and changes in competitive conditions. Ecoscience 13:258–270
Sardans J, Rodà F, Peñuelas J (2006b) Effects of a nutrient pulse supply on nutrient status of the Mediterranean trees Quercus ilex subsp. ballota and Pinus halepensis on different soils and under different competitive pressure. Trees 20:619–632
Sardans J, Peñuelas J, Rodà F (2006c) The effects of nutrient availability and removal of competing vegetation on resprouter capacity and nutrient accumulation in the shrub Erica multiflora. Acta Oecol 29:221–232
Sardans J, Peñuelas J, Estiarte M (2006d) Warming and drought alter soil phosphatase activity and soil P availability in a Mediterranean shrubland. Plant Soil 289:227–238
Sardans J, Peñuelas J, Estiarte M (2007) Seasonal patterns of root-surface phosphatase activities in a Mediterranean shrubland. Response to experimental warming and drought. Biol Fertil Soils 43:779–786
Sardans J, Peñuelas J, Estiarte M, Prieto P (2008a) Warming and drought alter C and N concentration, allocation and accumulation in a Mediterranean shrubland. Global Change Biol 14:2304–2316
Sardans J, Peñuelas J, Ogaya R (2008b) Drought-induced changes in C and N stoichiometry in a Quercus ilex Mediterranean forest. For Sci 54:513–522
Sardans J, Peñuelas J, Ogaya R (2008c) Experimental drought reduced acid and alkaline phosphatase activity and increased organic extractable P in soil in a Quercus ilex Mediterranean forest. Eu J Soil Biol 44:509–520
Sardans J, Peñuelas J, Estiarte M (2008d) Changes in soil enzymes related to C and N cycle and in soil C and N content under prolonged warming and drought in a Mediterranean shrubland. Appl Soil Ecol 39:223–235
Sardans J, Peñuelas J, Ogaya R (2008e) Drought’s impact on Ca, Fe, Mg, Mo and S concentration and accumulation patterns in the plants and soil of a Mediterranean evergreen Quercus ilex forest. Biogeochemistry 87:49–69
Sardans J, Peñuelas J, Prieto P, Estiarte M (2008f) Changes in Ca, Fe, Mg, Mo, Na, and S content under warming and drought. J Geophys Res 113:G03039
Sardans J, Peñuelas J, Prieto P, Estiarte M (2008g) Drought and warming induced changes in P and K concentration and accumulation in plant biomass and soil in a Mediterranean shrubland. Plant Soil 306:261–271
Sardans J, Peñuelas J, Estiarte M (2008h) Warming and drought change trace element bioaccumulation patterns in a Mediterranean shrubland. Chemosphere 70:874–885
Sardans J, Rivas-Ubach A, Peñuelas J (2011) Factors affecting nutrient concentration and stoichiometry of forest trees in Catalonia (NE Spain). For Ecol Manag 262:2024–2034
Sardans J, Peñuelas J, Coll M, Vayreda J, Rivas-Ubach A (2012a) Stoichiometry of potassium is largely determined by water availability and growth in Catalonian forests. Funct Ecology. In press.
Sardans J, Rivas-Ubach A, Peñuelas J (2012b) The C:N:P stoichiometry of organisms and ecosystems in a changing world: A review and perspectives. Persp Plant Ecol Evol Syst 14:33–47
Sardans J, Rivas-Ubach A, Peñuelas J (2012c) The elemental stoichiometry of aquatic and terrestrial ecosystems and its relationships with organismic lifestyle and ecosystem structure and function: a review and perspectives. Biogeochemistry. doi: 10.1007/s10533-011-9640-9
Saura-Mas S, Lloret F (2009) Linking post-fire regenerative strategy and leaf nutrient content in Mediterranean woody plants. Persp Plant Ecol Evol Syst 11:219–229
Saurer M, Cherubini P, Bonani G, Siegwolf R (2003) Tracing carbon uptake from a natural CO2 spring into tree rings: an isotope approach. Tree Physiol 23:997–1004
Scarascia-Mugnozza G, De Angelis P, Matteucci G, Valentini R (1996) Long-term excposure to elevated [CO2] in a natural Quercus ilex L. community: net photosynthesis and photochemical efficiency of PSII at different levels of water stress. Plant Cell Environ 19:643–654
Schlesinger WH, Hasey MM (1981) Decomposition of Chaparral shrub foliage: losses of organic and inorganic constituents from deciduous and evergreen leaves. Ecology 62:762–774
Serrano L, Peñuelas J, Ogaya R, Savé R (2005) Tissue-water relations of two co-occuring Evergreen Mediterranean species in response to seasonal and experimental drought conditions. J Plant Res 118:263–269
Sevink J, Imeson AC, Verstratem JM (1989) Humus form development and hillslope runoff, and the effects of fire and management, under Mediterranean forest in NE Spain. Catena 16:461–475
Shakesby RA (2011) Post-wildfire soil erosion in the Mediterranean: review and future research directions. Earth Sci Rev 105:71–100
Silla F, Escudero A (2003) Uptake, demand and internal cycling of nitrogen in saplings of Mediterranean Quercus species. Oecologia 136:28–36
Silva JS, Rego F (2003) Root distribution of a Mediterranean shrubland in Portugal. Plant Soil 255:529–540
Simón M, García I, Gil C, Polo A (1994) Characteristics of the organic matter of Mediterranean high-mountain soils. Geoderma 61:119–131
Skerget M, Kotnik P, Hadolin P, Hras HR, Simonic M, Knez Z (2005) Phenols, proanthocyanidins, flavones and flavonols in some plant materials and their antioxidant activities. Food Chem 89:191–198
Sofo A, Dichio B, Xiloyannis C, Masia A (2004) Lipoxygenase activity and proline accumulation in leaves and roots of olive trees in response to drought stress. Physiol Plantarum 121:58–65
Sofo A, Manfreda S, Fiorentino M, Dichio B, Xiloyannis C (2008) The olive tree: a paradigm for drought tolerance in Mediterranean climates. Hydrol Earth Syst Sci 12:293–301
Specht RL (1973) Structure and functional response of ecosystems in the Mediterranean climate of Australia. In: Chapman Di Castri F, Mooney HA (eds) Mediterranean-type ecosystems: origen and structure. Chapman and Hall, London, pp 113–120
Specht RL (1979) The sclerophyllous heath vegetation of Australia: the eastern and central states. In: Specht RL (ed) Heathlands and related shrublands. Descriptive Studies. Elsevier, Amsterdam, pp 125–210
Specht RL (1981) Primary production in Mediterranean-climate ecosystems regenerating after fire. In: Di Castri F, Goodwall DW, Specht RL (eds) Mediterranean-Type Ecosystems. Elsevier, Amsterdam, pp 257–268
Swarbreck S, Sudderth EA, StClair SB, Salve R, Casthanha C, Torn MS, Ackerly DD, Andersen GL (2011) Linking leaf transcript levels to whole plant analyses provides mechanistic insights to the impact of warming and altered water availability in an annual grass. Global Change Biol 17:1577–1594
Tabatabai MA (1994) Soil enzymes. In: Weaver RW, Angle JS, Bottomley PS (eds) Method of Soil Analysis. Part 2. Microbiological and biochemical properties. SSSA Book Series
Tabatabai MA, Bremner JM (1969) Use of p-nitrophenyl phosphate for assay of soil phosphatase activity. Soil Biol Biochem 1:301–307
Talmon Y, Sternberg M, Grünzweig JM (2011) Impact of rainfall manipulations and biotic controls soil respiration in Mediterraneanj and desert ecosystems along an aridity gradient. Global Change Biol 17:1108–1118
Tognetti R, Peñuelas J (2003) Nitrogen and carbon concentrations, and stable isotope ratios in Mediterranean shrubs growing in the proximity of a CO2 spring. Biol Plantarum 46:411–418
Tognetti R, Longobucco A, Miglietta F, Raschi A (1998) Transpiration and stomatal behaviour of Quercus ilex plants during the summer in a Mediterranean carbon dioxide spring. Plant Cell Environ 21:613–622
Tognetti R, Longobucco A, Raschi A, Miglietta F, Fumagalli I (1999a) Responses of two populus clones to elevated atmospheric CO2 concentration in the field. Ann For Sci 56:493–500
Tognetti R, Longobucco A, Raschi A (1999b) Seasonal embolism and xylem vulnerability in deciduous and evergreen Mediterranean trees influenced by proximity to a carbon dioxide spring. Tree Physiol 19:271–277
Tognetti R, Minnocci A, Peñuelas J, Raschi A, Jones MB (2000) Comparative field water relations of three Mediterranean shrub species co-occuring at a natural CO2 vent. J Exp Bot 51:1135–1146
Tsiafouli MA, Kallimanis AS, Katana E, Stamou GP, Sgardelis SP (2005) Responses of soil microarthropods to experimental short-term manipulation of soil moisture. Appl Soil Ecol 29:17–26
Turner IM (1994) Sclerophylly: primarily protective? Funct Ecol 8:669–675
Unger S, Máguas C, Pereira JS, David TS, Werner C (2010) The influence of precipitation pulses on soil respiration – assessing the “Birch effect” by stable carbon isotopes. Soil Biol Biochem 42:1800–1810
Vacca Z (2000) Effects of land use on forest floor and soil of a Quercus suber L. forest in Gallura (Sardinia, Italy). Land Degrad Develop 11:167–180
Van Wesemael B (1993) Litter decomposition and nutrient distribution in humus profiles in some Mediterranean forests in southern Tuscany. For Ecol Manag 57:99–114
Van Wesemael B, Veer M (1992) Soil organic-matter accumulation, litter decomposition and humus forms under Mediterranean-type forest in southern Tuscany. J Soil Sci 43:133–144
Van Wesemael B, Verstraten JM (1993) Organic acids in a morder type humus profile under a Mediterranean oak forest. Geoderma 59:75–88
Vaughn KJ, Biel C, Clary JJ, de Herralde F, Aranda X, Evans RY, Young TP, Savé R (2011) California perennial grasses are physiologically distinct from both Mediterranean annual and perennial grasses. Plant Soil 345:37–46
Vaz M, Pereira JS, Gazarini LC, David TS, David JS, Rodrigues A, Maroco J, Chaves MM (2010) Drought-induced photosynthetic inhibition and autumn recovery in two Mediterranean oak species (Quercus ilex and Quercus suber). Tree Physiol 30:946–956
Veneklas EJ, Poot P (2003) Seasonal patterns in water use and leaf turnover of different plant functional types in a species-rich woodland, south-western Australia. Plant Soil 257:295–304
Vilà M, Sardans J (1999) Plant competition in Mediterranean-type vegetation. J Veg Sci 10:281–294
Vilagrosa A, Morales F, Abadía A, Bellot J, Cochard H, Gil-Pelegrín E (2010) Are symplast tolerante to intense drought conditions and xylem vulnerability to cavitation coordinated? An integrated analysis of photosynthetic, hydraulic and leaf level processes in two Mediterranean drought-resistant species. Environ Exp Bot 69:233–242
Volaire F (1995) Growth, carbohydrate reserves and drought survival strategies of contrasting Dactylis glomerata populations in a Mediterranean environment. J Appl Ecol 32:56–66
Volaire F, Seddaiu G, Ledda L, Lelievre F (2009) Water deficit and induction of summer dormancy in perennial Mediterranean grasses. An Bot 103:1337–1346
Vourlitis GL, Pasquini SC (2009) Experimental dry-season N deposition alters species composition in southern California Mediterranean-type shrublands. Ecology 90:2183–2189
Wessel WW, Tietema A, Beier C, Emmett BA, Peñuelas J, Riis-Nielsen T (2004) A qualitative ecosystem assessment for different shrublands in western Europe under impact of climate change. Ecosystems 7:662–671
West AG, Dawson TE, February EC, Midgley GF, Bond WJ, Aston TL (2012) Diverse functional responses to drought in a Mediterranean-type shrublands in South Africa. New Phytol 195:396–407
Witkowski ETF, Mitchell DT, Stock WD (1990) Response of Cape fynbos ecosystem to nutrient additions: shoot growth and nutrient contents of a proteoid Leucospermun parile and an ericoid Phylica cephalantha evergreen shrub. Acta Oecol 11:311–326
Xiang SR, Doyle A, Holden PA, Schimel JP (2008) Drying and rewetting effects on C and N mineralizarion and microbial activity in surface and subsurface California grassland soils. Soil Biol Biochem 40:2281–2289
Xu M, Qi Y (2001) Soil-surface CO2 efflux and its spatial and temporal variations in a young ponderosa pine plantation in northern California. Global Change Biol 7:667–677
Yaalon DH (1997) Soils in the Mediterranean region: what makes them different? Catena 28:157–169
Zaman MDHJ, Cameron KC, Frampton CM (1999) Gross nitrogen mineralization and nitrification rates and their relationships to enzyme activities and the soil microbial biomass in soils treated with dairy shed effluent and ammonium fertilizer at different water potentials. Biol Fertil Soils 29:178–186
Zornoza R, Guerrero C, Mataix-Solera J, Arcenegui V, García-Orenes F, Mataix-Beneyto J (2006) Assessing air-drying and rewetting pre-treatment effect on some soil enzyme activities under Mediterranean conditions. Soil Biol Biochem 38:2125–2134
Zucca C, Canu A, Della Peruta R (2006) Effects of land use and landscape on spatial distribution and morphological features of gullies in an agropastoral area in Sardinia. Catena 88:87–95
Acknowledgments
This research was supported by the Spanish Government grants CGL2010-17172/BOS and Consolider-Ingenio Montes CSD2008–00040, and by the Catalan Government project SGR 2009–458.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Philippe Hinsinger.
Electronic supplementary material
Below is the link to the electronic supplementary material.
ESM 1
(DOC 343 kb)
Rights and permissions
About this article
Cite this article
Sardans, J., Peñuelas, J. Plant-soil interactions in Mediterranean forest and shrublands: impacts of climatic change. Plant Soil 365, 1–33 (2013). https://doi.org/10.1007/s11104-013-1591-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-013-1591-6