
Abstract
Dinitrogen fixation by cyanobacteria is of particular importance for the nutrient economy of cold biomes, constituting the main pathway for new N supplies to tundra ecosystems. It is prevalent in cyanobacterial colonies on bryophytes and in obligate associations within cyanolichens. Recent studies, applying interspecific variation in plant functional traits to upscale species effects on ecosystems, have all but neglected cryptogams and their association with cyanobacteria. Here we looked for species-specific patterns that determine cryptogam-mediated rates of N2 fixation in the Subarctic. We hypothesised a contrast in N2 fixation rates (1) between the structurally and physiologically different lichens and bryophytes, and (2) within bryophytes based on their respective plant functional types. Throughout the survey we supplied 15N-labelled N2 gas to quantify fixation rates for monospecific moss, liverwort and lichen turfs. We sampled fifteen species in a design that captures spatial and temporal variations during the growing season in Abisko region, Sweden. We measured N2 fixation potential of each turf in a common environment and in its field sampling site, in order to embrace both comparativeness and realism. Cyanolichens and bryophytes differed significantly in their cyanobacterial N2 fixation capacity, which was not driven by microhabitat characteristics, but rather by morphology and physiology. Cyanolichens were much more prominent fixers than bryophytes per unit dry weight, but not per unit area due to their low specific thallus weight. Mosses did not exhibit consistent differences in N2 fixation rates across species and functional types. Liverworts did not fix detectable amounts of N2. Despite the very high rates of N2 fixation associated with cyanolichens, large cover of mosses per unit area at the landscape scale compensates for their lower fixation rates, thereby probably making them the primary regional atmospheric nitrogen sink.
Similar content being viewed by others

Introduction
Through the quantification of interspecific variation in plant functional traits, comparative plant ecology has gained momentum in explaining species distributions, tradeoffs in biochemistry and physiology, as well as whole-ecosystem processes (Westoby and Wright 2006). Lavorel et al. (2007) considered vascular plant functional traits as predictors of key vegetation responses to environmental variation and vegetation effects on ecosystem functions at diverse spatial and temporal scales. In contrast to the effort put into developing extensive international methodological protocols and databases for quantitative analysis of vascular plant traits (Cornelissen et al. 2003), non-vascular plants such as cryptogams have remained mostly out of the scientists’ trait research scope to date, even though the application of bryophyte and lichen traits as drivers of large-scale biogeochemistry holds much promise. Indeed, bryophytes and lichens are of particular importance in cold biomes such as the Arctic tundra (Longton 1997), where they are key contributors to green biomass and control soil hydrology, temperatures and chemistry (Cornelissen et al. 2007). One aspect of the latter is their ability to form symbiotic relationships with cyanobacteria (Dalton and Chatfield 1985; During and Van Tooren 1990; Henriksson et al. 1987). Most bryophytes have a high water retention capacity and thus provide a stable and favourable habitat for cyanobacterial growth and N2 fixation activity (Dickson 2000). In lichens, cyanobacteria are a symbiont which provide their fungal partner with nitrogen from fixed atmospheric N2 in exchange for physical protection and—in the case of tripartite lichens—from carbohydrates in the fungi (Nash 1996). Since nitrogen is a principal limiting environmental factor for plant growth and soil organic matter turnover in polar regions (Longton 1988) and biological nitrogen fixation there occurs mostly in cyanobacteria (Solheim et al. 1996), the interaction between cryptogams and cyanobacteria forms an important field for ecological investigations.
A great deal of variation in nitrogen fixation of cyanobacterial bryophytes has been attributed to abiotic factors; Zielke et al. (2002) reported that in the High Arctic there is a strong dependency of fixation rates on site hydrology, temperature and light intensity. In regions with low precipitation during the growing season, for instance, N2 fixation is limited either to the period of snowmelt or to sites which stay wet during summer, e.g. peat bogs, fens, or dense moss mats (Zielke et al. 2005). Experimental long-term increase in UV-B radiation, simulating atmospheric ozone depletion, reduces N2 fixation activity in Arctic mosses, but does not affect it in sub-Arctic ones (Solheim et al. 2002, 2006). Fixation rates also depend on the successional age of boreal forests via differences in environmental moisture, nutrition and possible other factors (DeLuca et al. 2008; Lagerström et al. 2009; Zackrisson et al. 2004). In contrast to this emphasis on abiotic control over N2-fixation, only a few studies have investigated the role of host specificity in the interaction between cyanobacteria and bryophytes (see During and Van Tooren 1990). For mosses the types of association probably range from simply fortuitous, through more or less specific epiphytism, to intracellular colonisation of dead moss cells or hollow hyaline cells (Solheim and Zielke 2002). Even though extensive information is available on cyanobacteria-plant symbioses (including therein liverworts and hornworts) (Rai et al. 2000), there is hardly any information on how moss-cyanobacteria associations are formed, what genes are involved if any, and the extent of nitrogen exchange between host and symbiont. At an ecosystem level, a single study seldom combines more than a couple of host species and those are often closely related phylogenetically. This poses problems when estimations of regional biological atmospheric nitrogen fixation are attempted. Even the widely applied technique for measuring N2 fixation rates by means of acetylene reduction capacity provides idiosyncratic data, unless calibrated for each analysis by means of 15N2 labelled gas (DeLuca et al. 2002).
This study is the first to examine the effects of cryptogam host specificity on rates of N2 fixation, independent of abiotic environmental factors. We hypothesised principal differences in N2 fixation rates (1) between lichens and bryophytes, and (2) among bryophyte plant functional types. Thereto, measurements were done in monospecific cryptogam patches both in the field and in a common environment, using a standard method for 15N2 incubation. We surveyed a list of contrasting, abundant sub-arctic species to account for the phylogenetic, morphological and habitat diversity of hosts to cyanobacteria in the region. This host specificity could possibly arise from differences in surfaces available for bacterial colonisation, density of vegetation patches, and consequently in microhabitat moisture. On a larger scale, we compared the relative contributions of cyanobacterial bryophytes versus cyanolichens to the regional input of atmospheric nitrogen in the Subarctic. We hypothesised (3) that, because of the overall dominance in the regional vegetation cover, bryophytes could play an important role in overall N2-fixation in the Subarctic. To test the latter hypothesis, we did not limit our analyses to comparisons on dry weight or area basis, but also included regional abundance.
Methods
Site description and plant material sampling
Field work took place in the summer of 2007 off the southern shore of Lake Torneträsk, North Sweden. Three distinct valleys in the vicinity of Abisko were considered for collecting cryptogamsy4—one in Stordalen Nature Reserve and two in Abisko National Park. Each of the three locations included three vegetation types characteristic of the lowland Subarctic: nutrient poor birch forest with ericaceous undergrowth, oligotrophic bog dominated by Sphagnum fuscum hummock communities, and minerotrophic mire (fen). Thus each of the studied species was collected from three spatially separated habitats. In total 15 cryptogam species from different taxonomic groups, with varying morphologies and from contrasting habitats were considered for this study. These included 11 mosses—Aulacomnium palustre (Hedw.) Schwaeger., Tomentypnum nitens (Hedw.) Loeske, Drepanocladus cossonii (Schimp.) Loeske, Paludella squarrosa (Hedw.) Brid. Muscol., Sphagnum lindbergii Schimp. ex Lindb, Sphagnum riparium Aongstr., Sphagnum fuscum (Schimp.) Klinggr., Hylocomnium splendens (Hedw.) B.S.G., Pleurozium schreberi (Brid.) Mitt., Polytrichum strictum Brid., Polytrichum commune Hedw.; two liverworts—Lophozia lycopodioides (Wallr.) Cogn., Ptilidium ciliare (L.) Hampe; and two cyanolichens—Nephroma arcticum (L.) Torss., Peltigera aphthosa (L.) Willd. Nomenclature followed recent checklists and identification guides: Drepanocladus sp. (Hedenäs 2003), Sphagna (Daniels and Eddy 1990), other musci (Hill et al. 2006), liverworts (Damsholt 2002), and lichens (Dobson 2005). We sampled monospecific patches of vegetation typical for the particular species’ habitat. For each species four samples were gathered from each valley—two for incubation with 15N2 and two as controls. Collection from the same sites was performed twice, in mid-June and then in mid-July, to account for possible temporal variation through the growing season. However, due to logistic constraints, six of the moss species were collected and incubated only once, either during the first or the second round (for details see section “Statistical analysis”).
The bryophyte species were grouped into two functional types according to their shoot growth form, and mat architecture (Buck and Goffinet 2000): acrocarpous (erect shoots, growing in dense turfs) and pleurocarpous (crawling shoots, growing in mats).
In situ microhabitat moisture was recorded for each species, since we considered water availability a likely limiting factor for cyanobacterial activity. We monitored moisture availability in monospecific patches by means of air-dried wooden cylinders (r = 1.25 cm, h = 7 cm) nested for a week (July, 15–21) in the bryophyte turf at 5 cm depth. The cylinders were weighed before and after field placement to determine the relative increase in water content during a period without major rainfall.
Sample processing and analysis of N2 fixation
Pilot work determined the active part in N2 fixation along moss’ shoots. Moss was cut into previous year’s growth, older photosynthetically active part, and senesced part according to specific moss morphology and coloration. Segments below previous-year’s growth were the most prominent in nitrogen fixation activity (see Fig. 1). Additionally, a Confocal Laser Scanning Microscopy, CLSM (Bio-Rad Radiance 2000), determined whether the measured rates of N2 fixation corresponded to the degree of moss colonisation by cyanobacteria. The technique, described in detail by Solheim et al. (2004), is non-destructive and allows for optical cross-section observations of intact moss samples. Detected cyanobacterial filaments were found present in leaf groves and moss axils, with largest colonies observed along the mid-length shoot segments (Fig. 2, see also the correspondingly high fixation rates for mid-part of Drepanocladus cossonii in Fig. 1). Field measurements of live (photosynthetically active) moss cushion thickness in the region (a minimum of 6 replicates per species) varied between 3.1 cm and 7.5 cm across moss species and were around 2.5 cm for leafy liverworts. Hence, to account for the contribution of each species to the process of nitrogen fixation, we opted for a correspondingly large vial size for the main part of the experiment.

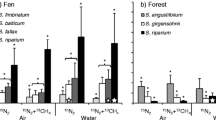
Daily N2 fixation rates by shoot segments of four moss species. In the imbedded legend “capitula” refers to previous year’s growth, “mid part” to the photosynthetically active part below the capitula, and “lower part” refers to the senesced part. Capital letters indicate overall statistical differences between species based on a Tukey post hoc test at the 0.05 significance level. Lower case letters indicate differences between moss segments in each respective species

A red filter CLSM micrograph of a Drepanocladus cossonii shoot showing (a), a colony of filamentous cyanobacteria detected on top of a folded leaf (magnification ×60) and (b), the same view (magnification ×200)
Collected plant material was temporarily stored in a dark climate chamber (5°C) and any living or dead material other than the target bryophyte or lichen carefully removed by hand. Mosses were cut to a maximal shoot length of 6 cm from the apex, liverworts were left intact, and for lichens three to four whole thallus lobes were used. Samples were saturated with water and placed in 135 ml glass vials. Right before incubation all samples were acclimated for 36 h outdoors in the experimental common environment, in a partly shaded spot just outside Abisko Research Station. Then the vials were sealed with rubber septa and in half of them 15 ml of headspace were replaced by 15 ml 15N2 (98% enriched, Cambridge Isotope Laboratories, Inc., U.S.A.), resulting in headspace enrichments of 10–19% depending on species [see Supplementary Materials S1 for note on calculation of final headspace enrichment]. The other half of the samples, the controls, was treated in the same manner except for the headspace substitution. All vials were placed randomly on three trays, representing statistical blocks (Quinn and Keough 2002), which corresponded to the three valleys the samples had been collected from. Incubation took 24 h during which the trays were rotated every 4 h to avoid possible unintended gradients in the common environment. Sunscreens were put up at noon to avoid direct sun illumination of the glass vials, in order to avoid any greenhouse effects (Zielke et al. 2002); this kept PAR at average levels of ca. 75 μEinsteins m−2 s−1 (ranging from 10 to 240 μEinsteins m−2 s−1). Additionally, temperature was recorded on Tinytag loggers coupled to probes inside and outside the vials and the data showed no significant difference within either of the incubation rounds (T-Test, for June P = 0.90; for July P = 0.64). June temperature averages inside and outside the vials were 18.0°C and 18.2°C respectively (13.5–22.7°C range). For July these averages were 10.5°C and 10.3°C (6.9–14.5°C range).
During each of the two phases of the experiment, samples from two moss species were nested in the field for acclimation and subsequent incubation in order to test for possible discrepancies between fixation rates in the experimental common environment and those in the field. We had indeed chosen species from contrasting environments to account for the range of habitats in this study—H. splendens from a dry birch forest, A. palustre from wet depressions within a birch forest, S. fuscum from hummocks in an oligotrophic bog, and D. cossonii from water pools in a minerotrophic fen.
After the incubations all plant and lichen material was oven-dried for 48 h at 70°C, weighed, and powdered in a ball mill. Nitrogen concentrations and atom% 15N values were determined using an Elemental Analyser (NC2500, ThermoQuest Italia, Rodana, Italy) coupled to a Continuous-flow Isotope Ratio Mass Spectrometer (Delta Plus, ThermoQuest Finnigan, Bremen, Germany). N2 fixation was determined by the 15N increment over the corresponding controls’ concentrations, corrected for each samples’ headspace volume and expressed per unit dry weight [see Supplementary Materials S1 for details on calculations].
Estimation of regional N2 fixation
In order to get a better understanding of the regional significance of N2 fixation associated with subarctic cryptogams, fixation rates were expressed on an area basis. To this aim, data on bryophyte cushion density and thickness per dry weight was gathered in the Abisko region with a minimum of six replicates per species. For lichens, oven-dry weight of individual thalli was expressed per unit surface area of fully expanded thalli (thallus specific weight) as in Dahlman and Palmqvist (2003). Variances of these measurements were incorporated in the final regional-scale N2 fixation budgets. The latter were consecutively upscaled to the landscape level by integrating species abundance and cover percentage data from former biodiversity studies in the Abisko region.
Statistical analysis
Data were analysed using the SPSS 13.0 software package. Rank transformations were carried out when both Levene’s test and visual exploration of residual plots for homogeneity of variances violated the assumptions of the statistical tests used (Quinn and Keough 2002). A Repeated Measures ANOVA for the two consecutive measurements at each location could not be performed due to violations of the basic assumptions of the test (homogeneity of variances and normality) and lack of useful biologically meaningful data transformations. Thus the two observations in time (in June and July) were treated as a nested factor within the main factor—species, to account for the fact that they were performed on the same plots. The local effect of valley location (sampling site), the effect of species and their interaction were tested. This analysis was performed on the reduced list of nine species which had been replicated over the two rounds of the experiment. Since no temporal variation was detected (see below), the whole list of 15 species (thus including S. riparium, P. schreberi and P. strictum from the first round; and S. lindbergii, H. spendens, and P. commune from the second round) was tested with One-Way ANOVA followed by a Tukey’s test for differential effects of each species, and the two moss functional type groups (acrocarps and pleurocarps), respectively. Field measurements of nitrogen fixation were tested against common environment measurements on the same species coming from corresponding valleys and collection times by a Two-Way ANOVA model. At last, data on field moisture levels for each species were compared to corresponding rates of N2 fixation and analysed by means of a linear regression model.
Results
N2 fixation across cryptogam species
Natural variation in amounts of stable isotope nitrogen in cryptogams proved to be considerable (average δ15N = 0.68, range from −5.29 to 25.01) and in some cases exceeded the enrichment after incubation (average δ15N = 7.31, range from −5.49 to 86.63). Hence, several control samples were richer in 15N than their incubation counterparts and this rendered expressing daily species-specific N2 fixation rates difficult (see Table 1, negative fixation rates). However, the intercept of our statistical model for differences in fixation rates (the one testing the effects of valleys (collecting sites), species, their interaction, and collection time as a nested factor in species) was significantly different from zero (P = 0.005) and positive (B = 57.0). This provided evidence that incubation significantly increased the amounts of labeled N2 in the samples altogether. When tested separately, this scenario also held true for lichens (P = 0.010; B = 2,631) and mosses (P = 0.044; B = 57.0), but not for liverworts which showed no significant 15N uptake (P = 0.75; B = 152). We found no significant effects of valleys (P = 0.17) or collection time (P = 0.47) on the nitrogen fixation rates and concluded that within the studied region and time scale there were no consistent spatial or temporal variations in fixation rates. There was a highly significant effect of species (P < 0.001), but none of the species by valleys interaction (P = 0.59).
In order to include all of the species (together with the six species tested for N2 fixation only in one of the collection rounds) we had to average the two observations in time (June and July) for the rest of the species, which was justified based on the non-significant influence of time from the first test. The design included two factors—valleys and species, with a post-hoc test on species. Species had a significant effect (P = 0.020) on 15N accumulation, which was not influenced by valleys (P = 0.409). The only two cryptogam species that were fixing atmospheric nitrogen at significantly different rates from one another and higher than the bryophytes were the two lichens P. aphthosa and N. arcticum (see Table 1). Correspondingly, the tissue nitrogen contents of these two species (25.2 mg N g−1 dry weight in P. aphthosa and 20.4 in N. arcticum) were also much higher than those of the mosses (a range of 3.4–9.6 mg N g−1 dry weight for all the studied species).
For the four species nested in the field we found no significant differences in the pairwise comparisons between fixation rates in the experimental common environment and those in the ‘home site’ of each species (P = 0.32). Thus we concluded that exhibited rates of N2 fixation in the common environment were realistic field values.
The linear regression model of species N2 fixation rates and environmental moisture did not explain much of the observed variation (R 2 = 0.15, P = 0.24). Whilst the higher fixation rates were mostly seen among the wetter species, there were also wet species with very low rates. Furthermore, grouping of bryophytes into functional types according to their morphology did not reduce the variation within mosses and liverworts (data not shown).
Discussion
This study is the first to compare N2 fixation rates of a range of lichen and bryophyte species in a direct and standardised way in order to detect and quantify species contributions to ecosystem-level N2 fixation. It is also the first attempt at classifying cryptogams into functional types according to their potential to sustain colonies of symbiotic cyanobacteria. We have presented evidence for differential N2 fixation rates among high-level taxonomic lineages of Subarctic cryptogams and tested underlying patterns which might contribute to understanding the process. These include phylogenetic relationships among genera, extent of symbiotic interaction, growth form and morphology, and moisture. The study was carried out in standardised conditions for all species and, considering the large incubation vial size, provides relatively realistic estimates of the atmospheric nitrogen inputs to local ecosystems.
N2 fixation as a cryptogam functional trait?
We found significant differences in absolute rates of bivalent nitrogen fixation between bryophytes and lichens, and between the two lichens we considered in this study. Within bryophytes, mosses and liverworts showed a clear dissimilarity in their fixation rates as well. The results were highly variable from sample to sample, as also reported in studies from the high-Arctic (Solheim et al. 1996) and the Subarctic (Basilier et al. 1978), but we found no consistent temporal or spatial pattern within the range of our study. There were clear differences in the maximal amounts of fixed nitrogen among the considered species, but the within-species variation masked such patterns in the species means. The latter factor also hampered our attempt to report underlying patterns for the observed diversity in fixation rates. We found no correlation between growth form or habitat moisture and N2 fixation. Hence the only meaningful grouping factors were higher-level taxonomic lineages and specificity of the symbiotic relationship.
Clearly, lichens demonstrated outstanding capabilities of N2 fixation on a dry weight basis and this is possibly due to the symbiotic association between mycobiont and cyanobiont. It is interesting to note that the two lichen species showed also, correspondingly to their individual fixation rates, the highest tissue N-content, which can be a result of intense incorporation of ammonium derived from cyanobacteria by the mycobiont (Rai 1988). They are both tripartite ones and the primary photobiont green algae form the main body throughout the lichen thalli. Cyanobacteria are the secondary symbiont and they are enclosed in cephalodia, where they behave as heterotrophs, providing nitrogen to their host in return for sugars (Nash 1996). This heterotrophic strategy allows for a much higher ratio of heterocysts (the actual site of nitrogen fixation) to vegetative cells in the cyanobacterial filaments. A study by Hitch and Millbank (1975) reports for Peltigera aphthosa and Nephroma arcticum 21.1% and 14.1% heterocyst frequencies respectively, whereas in bipartite cyanolichens heterocysts form on average 4% of the total algal cell population. Hence, it would be interesting to screen a wide range of bi- and tri-partite lichens and check whether these consistent morphological differences are manifested at the level of N2 fixation rates.
Moss species in this study formed a group with rates of N2 fixation one or two orders of magnitude lower than those in lichens. This corroborates findings of Solheim et al. (2002) who estimated fixation rates under natural conditions in Abisko at 2,760 μmol N kg−1 d−1 for the cyanolichen P. aphthosa and 64 μmol N kg−1 d−1 for the forest moss H. splendens. The differences we found in fixation rates among different moss species, although non-significant, appeared consistent. We expected these to be correlated with different hydrology regimes at a microhabitat scale for species specific patches. However the big variation in fixation rates within species did not allow us to build a strong regression model. Subdivision of mosses into groups based on their morphology or habitat did not succeed in explaining the observed N2 fixation rate differences either, owing to the same complications.
The two liverworts in our study, Lophozia lycopodioides and Ptilidium ciliare, showed no fixation at all. They even showed negative values of fixation (see Table 1), as a likely artefact of our method of subtracting 15N concentrations in control samples from those in incubated ones. Thus, in practice these values show a lack of nitrogen fixation activity. Despite the frequent abundance of the two birch forest species in the subarctic region, the results for degree of colonisation by cyanobacteria should not be surprising. Although in scientific literature close (internal) symbiotic relationships between a few liverwort species and cyanobacteria have been reported (see Adams 2002), none of them listed L. lycopodioides or P. ciliare. Liverworts are known for rich composition of secondary compounds (Asakawa 1995, 2004; Mues 2000), many of which exhibit antimicrobial properties (Asakawa 2008). We speculate these compounds might also thwart colonization of liverworts by epiphytic cyanobacteria.
Methodological issues
Throughout the survey we experienced difficulties analysing the data on 15N accumulation due to the great natural variation of this isotope in the Abisko non-vascular vegetation. A possible source of this signal noise (i.e. high 15N levels in control samples) could have arisen due to the sampling approach. Moss was cut out to a depth of 6 cm below the surface, and thus potentially included old litter material, which might be enriched with the heavier nitrogen isotope as a result of discrimination during litter decomposition (Nadelhoffer et al. 1996). Nevertheless, we believe that the results shown here are robust and conclusive on the actual amount of atmospheric nitrogen fixation at the level of higher taxa. We consider techniques involving acetylene reduction activity (ARA) to be less laborious but also less informative especially when correlation to N2 fixation and ammonia production is attempted. Theoretical stoichiometric ratio of ethylene to nitrogen (3:2) is rarely attained and must always be corrected by parallel uptake of labelled 15N2 for each organism and possibly location (Basilier 1980; Millbank 1981). Thus we advocate the use of labelled nitrogen gas in further studies and tentatively propose a more extensive sampling for natural abundance of 15N isotope in the study sites.
Regional upscaling of the results
The process of bivalent nitrogen fixation associated with subarctic cryptogams affects nutrient cycling within the ecosystem in a number of ways, but based on our experimental study alone we cannot ascertain how much nitrogen enters the biocycle. Indeed, we need to account for the processes of denitrification, leaching, and relative contribution compared to nitrogen input from other sources (i.e. wet deposition, free-living cyanobacteria on bare soil, nodulated vascular plants). Since these are out of the scope of the current study we remain cautious in extrapolating the results. Here we use former biodiversity studies in the Abisko region on species and vegetation type cover percentage estimations for the studied habitats. These data, nevertheless, are not meant to serve as a ground for total nitrogen input in the area, but rather as a means to estimate the relative spatial contribution of each of the life forms we investigated to N2 fixation. In order to achieve this we reported (Table 1) nitrogen fixation both on area and biomass bases, thus accounting for the high area-to-dry weight ratio of the lichens (Nash 1996). The information presented in Table 2 reflects the total nitrogen that enters a respective habitat per day, based on the ground cover of abundant habitat-characteristic species.
The two common cyanolichens we surveyed, N. arcticum and P. aphthosa, appear to contribute as much as the forest mosses to the total ecosystem input of atmospheric nitrogen into birch-heath woodland. Cyanolichens are virtually absent from the other two habitats studied, fens and bogs, where mosses fix substantial N2 on a landscape area basis. Thus we claim that mosses probably play a key role in the process of nitrogen fixation there. A similar pattern emerges from older studies done in the region on total yearly nitrogen input. The two cyanolichens S. paschale and N. arcticum fix 0.38 g N m−2 yr−1 in similar Fennoscandian woodland (Longton 1997) and N. arcticum alone fixes 0.1 g N m−2 yr−1 in Fennoscandian tundra (Sonesson et al. 1975), whereas the two mosses S. riparium and Drepanocladus exannulus together fix as much as 0.3–4.3 g N m−2 yr−1 in the Stordalen fen, Abisko (Basilier 1979). In comparison, atmospheric deposition levels of nitrate (NOy) and ammonia (NHx), as reported in a survey by the Swedish Environmental Research Institute, are less than 0.4 g N m−2 yr−1 for the region of North Sweden (Binkley and Hogberg 1997). These data together ascertain the significance of biological N2 fixation to the N-limited vegetation in the Subarctic (Crittenden and Kershaw 1978).
In conclusion, our multispecies comparison of N2-fixation rates of cryptogams, both under standardised conditions and in situ, applying a direct nitrogen detection method, has provided strong evidence for differential nitrogen input by cryptogam taxa in the Subarctic, and substantiates their importance to the nitrogen economy of these cold biomes. Furthermore we demonstrate that high rates of nitrogen fixation shown by cyanolichens on a small scale can indeed be moderate when extrapolated to an ecosystem level. We conclude that mosses, with their high biomass and overall abundance, are at least equally important drivers of large-scale atmospheric nitrogen input to the Subarctic.
References
Adams DG (2002) Cyanobacteria in symbiosis with hornworts and liverworts. In: Rai AN, Bergman B, Rasmussen U (eds) Cyanobacteria in symbiosis. Kluwer, Dordrecht, pp 117–135
Asakawa Y (1995) Chemical constituents of the bryophytes. In: Herz W, Kirby G, Moore R, Steglich W, Tamm C (eds) Progress in the chemistry of organic natural products. Springer, New York, pp 1–618
Asakawa Y (2004) Chemosystematics of the Hepaticae. Phytochemistry 65:623–669
Asakawa Y (2008) Recent advances of biologically active substances from the marchantiophyta. Nat Prod Commun 3:77–92
Basilier K (1979) Moss-associated nitrogen-fixation in some mire and coniferous forest environments around Uppsala, Sweden. Lindbergia 5:84–88
Basilier K (1980) Fixation and uptake of nitrogen in sphagnum blue-green-algal associations. Oikos 34:239–242
Basilier K, Granhall U, Stenstrom TA (1978) Nitrogen-fixation in wet minerotropic moss communities of a Sub-Arctic mire. Oikos 31:236–246
Binkley D, Hogberg P (1997) Does atmospheric deposition of nitrogen threaten Swedish forests? For Ecol Manag 92:119–152
Buck WR, Goffinet B (2000) Morphology and classification of mosses. In: Shaw AJ, Goffinet B (eds) Bryophyte biology. Cambridge University Press, Cambridge, pp 71–123
Cornelissen JHC, Lavorel S, Garnier E, Diaz S, Buchmann N, Gurvich DE, Reich PB, ter Steege H, Morgan HD, van der Heijden MGA, Pausas JG, Poorter H (2003) A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Aust J Bot 51:335–380
Cornelissen JHC, Lang SI, Soudzilovskaia NA, During HJ (2007) Comparative cryptogam ecology: a review of bryophyte and lichen traits that drive biogeochemistry. Ann Bot 99:987–1001
Crittenden PD, Kershaw KA (1978) Discovering role of lichens in nitrogen cycle in boreal-arctic ecosystems. Bryologist 81:258–267
Dahlman L, Palmqvist K (2003) Growth in two foliose tripartite lichens. Nephroma arcticum and Peltigera aphthosa: empirical modelling of external vs. internal factors. Funct Ecol 17:821–831
Dalton DA, Chatfield JM (1985) A new nitrogen-fixing cyanophyte-hepatic association—Nostoc and Porella. Am J Bot 72:781–784
Damsholt K (2002) Illustrated flora of nordic liverworts and hornworts. Nordic Bryological Society, Lund
Daniels RE, Eddy A (1990) Handbook of European Sphagna. Institute of Terrestrial Ecology, Huntingdon
DeLuca TH, Zackrisson O, Nilsson MC, Sellstedt A (2002) Quantifying nitrogen-fixation in feather moss carpets of boreal forests. Nature 419:917–920
DeLuca TH, Zackrisson O, Gundale MJ, Nilsson MC (2008) Ecosystem feedbacks and nitrogen fixation in boreal forests. Science 320:1181–1181
Dickson LG (2000) Constraints to nitrogen fixation by cryptogamic crusts in a polar desert ecosystem, Devon Island, NWT, Canada. Arctic Antarct Alpine Res 32:40–45
Dobson F (2005) Lichens: An illustrated guide to the British and Irish species. Richmond, Richmond
During HJ, Van Tooren BF (1990) Bryophyte interactions with other plants. Bot J Linn Soc 104:79–98
Hedenäs L (2003) The European species of the Calliergon-Scorpidium-Drepanocladus complex, including some related or similar species. Meylania 28:1–116
Henriksson E, Henriksson LE, Norrman JO, Nyman PO (1987) Biological dinitrogen fixation (acetylene-reduction) exhibited by blue-green-algae (cyanobacteria) in association with mosses gathered on Surtsey, Iceland. Arctic Alpine Res 19:432–436
Hill MO, Bell N, Bruggeman-Nannenga MA, Brugues M, Cano MJ, Enroth J, Flatberg KI, Frahm JP, Gallego MT, Garilleti R, Guerra J, Hedenas L, Holyoak DT, Hyvonen J, Ignatov MS, Lara F, Mazimpaka V, Munoz J, Soderstrom L (2006) An annotated checklist of the mosses of Europe and Macaronesia. J Bryol 28:198–267
Hitch CJB, Millbank JW (1975) Nitrogen-metabolism in lichens: 7. Nitrogenase activity and heterocyst frequency in lichens with blue-green phycobionts. New Phytol 75:239–244
Kallio P (1975) Kevo, Finland. In: Rosswall T, Heal OW (eds) Structure and function of tundra ecosystems. Ecological Bulletins 20, Stockholm, pp 193–223
Lagerström A, Esberg C, Wardle DA, Giesler R (2009) Soil phosphorus and microbial response to a long-term wildfire chronosequence in northern Sweden. Biogeochemistry 95:199–213
Lavorel S, Diaz S, Conelissen JHC, Garnier E, Harrison SP, McIntyre S, Pausas JG, Perez-Harguindeguy N, Urcelay C (2007) Plant functional types: are we getting any closer to the Holy Grail? In: Pitelka LF, Canadell J, Pataki D (eds) Terrestrial ecosystems in a changing world. Springer, Berlin, pp 171–186
Longton RE (1988) The biology of polar bryophytes and lichens. Cambridge University Press, Cambridge
Longton RE (1997) The role of bryophytes and lichens in polar ecosystems. In: Woodin SJ, Marquiss M (eds) Ecology of arctic environments. Blackwell Science, Oxford, pp 69–96
Millbank JW (1981) The assessment of nitrogen-fixation and throughput by lichens: 1. The use of a controlled environment chamber to relate acetylene-reduction estimates to nitrogen-fixation. New Phytol 89:647–655
Mues R (2000) Chemical constituents and biochemistry. In: Shaw A, Goffinet B (eds) Bryophyte biology. Cambridge University Press, Cambridge, pp 150–181
Nadelhoffer K, Shaver G, Fry B, Giblin A, Johnson L, McKane R (1996) 15N natural abundances and N use by tundra plants. Oecologia 107:386–394
Nash TH III (1996) Nitrogen, its metabolism and potential contribution to ecosystems. In: Nash TH III (ed) Lichen biology. Cambridge University Press, Cambridge, pp 121–135
Quinn GP, Keough GP (2002) Experimental design and data analysis for biologists. Cambridge University Press, Cambridge
Rai AN (1988) Nitrogen metabolism. In: Galun M (ed) Handbook of lichenology. CRC Press, Boca Raton, pp 201–237
Rai AN, Soderback E, Bergman B (2000) Cyanobacterium-plant symbioses. New Phytol 147:449–481
Rosswall T, Flower-Ellis JGK, Johansson LG, Jonsson S, Ryden BE, Sonesson M (1975) Stordalen (Abisko), Sweden. In: Rosswall T, Heal OW (eds) Structure and function of tundra ecosystems. Ecological Bulletins 20, Stockholm, pp 265–294
Solheim B, Zielke M (2002) Associations between cyanobacteria and mosses. In: Rai AN, Bergman B, Rasmussen U (eds) Cyanobacteria in symbiosis. Kluwer, Dordrecht, pp 137–152
Solheim B, Endal A, Vigstad H (1996) Nitrogen fixation in Arctic vegetation and soils from Svalbard, Norway. Polar Biol 16:35–40
Solheim B, Johanson U, Callaghan TV, Lee JA, Gwynn-Jones D, Bjorn LO (2002) The nitrogen fixation potential of arctic cryptogram species is influenced by enhanced UV-Beta radiation. Oecologia 133:90–93
Solheim B, Wiggen H, Roberg S, Spaink HP (2004) Associations between arctic cyanobacteria and mosses. Symbiosis 37:169–187
Solheim B, Zielke M, Bjerke JW, Rozema J (2006) Effects of enhanced UV-B radiation on nitrogen fixation in arctic ecosystems. Plant Ecol 182:109–118
Sonesson M (1967) Studies on mire vegetation in Torneträsk area Northern Sweden: I. Regional aspects. Botaniska Notiser 120:272–296
Sonesson M, Kvillner E (1980) Plant communities of the Stordalen mire. A comparison between numerical and non-numerical classification methods. In: Sonesson M (ed) Ecology of a subarctic mire. Ecological Bulletins 30, Stockholm, pp 113–126
Sonesson M, Wielgolaski FE, Kallio P (1975) Description of Fennoscandian tundra ecosystem. In: Wielgolaski FE (ed) Ecological studies. Analysis and Synthesis. p. Part 1
Westoby M, Wright IJ (2006) Land-plant ecology on the basis of functional traits. Trends Ecol Evol 21:261–268
Wielgolaski FE (1972) Vegetation types and primary production in tundra. In: Wielgolaski FE, Rosswall T (eds) Tundra biome. International Biological Programme, Lund, pp 9–34
Zackrisson O, DeLuca TH, Nilsson MC, Sellstedt A, Berglund LM (2004) Nitrogen fixation increases with successional age in boreal forests. Ecology 85:3327–3334
Zielke M, Ekker AS, Olsen RA, Spjelkavik S, Solheim B (2002) The influence of abiotic factors on biological nitrogen fixation in different types of vegetation in the High Arctic, Svalbard. Arctic Antarct Alpine Res 34:293–299
Zielke M, Solheim B, Spjelkavik S, Olsen RA (2005) Nitrogen fixation in the high arctic: Role of vegetation and environmental conditions. Arctic Antarct Alpine Res 37:372–378
Acknowledgements
We are grateful to Ellen Dorrepaal for collecting moss samples in Abisko for the pilot experiment, to Peter van Bodegom for statistical advice, to Wilfred Röling for granting us access to a CLSM, and to colleagues who made comments on earlier versions of the manuscript. This study benefited greatly from funding by NWO (Netherlands Organisation for Scientific Research) to JHCC through grants 047.017.010, 047019008 and 047.018.003. The experiments reported here comply with the current laws of Sweden and The Netherlands, where this study was carried out.
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Euan K. James.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Supplementary Materials S1
(DOC 27 kb)
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License (https://creativecommons.org/licenses/by-nc/2.0), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Gavazov, K.S., Soudzilovskaia, N.A., van Logtestijn, R.S.P. et al. Isotopic analysis of cyanobacterial nitrogen fixation associated with subarctic lichen and bryophyte species. Plant Soil 333, 507–517 (2010). https://doi.org/10.1007/s11104-010-0374-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-010-0374-6